
Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
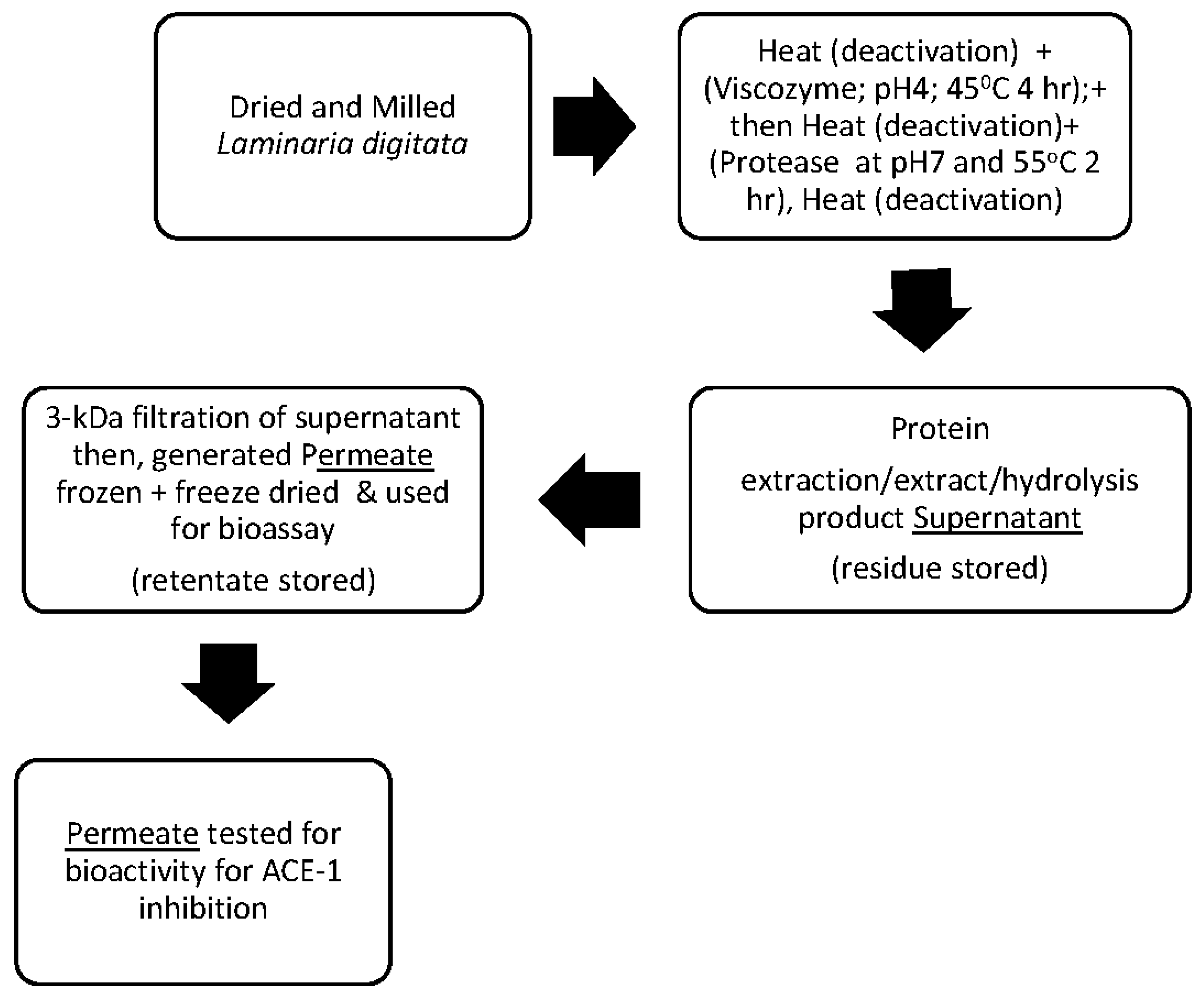
2.2. Generation of Peptide Hydrolysates from L. digitata
2.3. Peptide Enrichment Using Molecular Weight Cut Off (MWCO) Filtration
2.4. Chemical Characterisation of Seaweed and Seaweed Hydrolysates and Permeates
2.4.1. Protein Content
2.4.2. Fat and Moisture Content
2.4.3. Ash Content
2.5. Degree of Hydrolysis (DH) of L. digitata
2.6. Statistical Analysis
2.7. ACE-Inhibitory Activity Determination of L. digitata of 3 kDa Protein Hydrolysates, Permeates, and IC50
3. Results
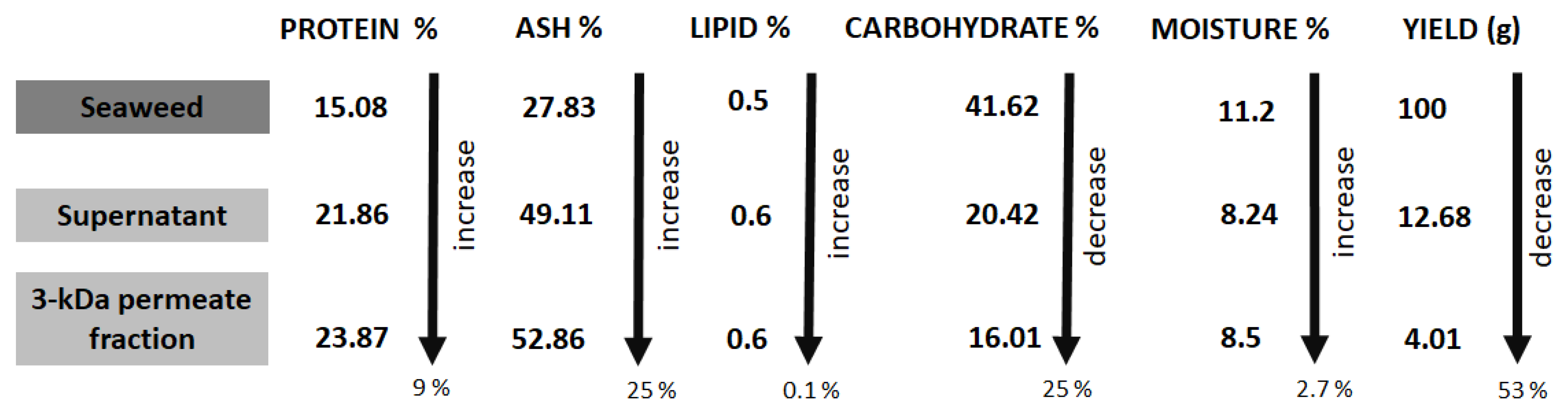
3.1. Extraction Yields
3.2. Nutritional Composition of Whole L. digitata, Hydrolysates and Permeates
3.3. Total Amino Acid (TAA) Profiles for Raw Seaweed (a), and 3 kDa Hydrolysate Permeate
3.4. ACE-1 Inhibition of Seaweed Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. In Globefish Research Programme; FAO: Rome, Italy, 2018; Volume 124. [Google Scholar]
- Koch, S.; van den Berg, S.; Nanta, R.; van der Werf, A. The role of seaweed in the future food system. Wagening. Econ. Res. 2021, 1, 1–15. [Google Scholar]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid liquid extraction method on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. Technol. 2012, 48, 860–869. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential Role of Seaweed Polyphenols in. Cardiovascular-Associated Disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermano, G.; Stoyanova, T.; Hennequart, F.; Wainwright, C.L. Seaweed-derived bioactives as potential energy regulators in obesity and type 2 diabetes. Adv. Pharmacol. 2020, 87, 205–256. [Google Scholar] [PubMed]
- O’Brien, R.; Hayes, M.; Sheldrake, G.; Tiwari, B.; Walsh, P. Macroalgal Proteins: A Review. Foods 2022, 11, S209–S212. [Google Scholar]
- Maruyama, S.; Segawa, Y.; Hashimoto, H.; Kitamura, S.; Kimura, M.; Osera, T.; Kurihara, N. Role of alginate in the mechanism by which brown seaweed Saccharina japonica intake alleviates an increase in blood pressure in 2-kidney, 1-clip renovascular hypertensive rats. Clin. Exp. Hypertens. 2022, 44, 72–82. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kang, M.C.; Kang, N.; Kim, H.-S.; Lee, S.-H.; Ahn, G.; Jung, W.-K.; Jeon, Y.-J. Effect of angiotensin I-converting enzyme (ACE) inhibition and nitric oxide (NO) production of 6, 6′-bieckol, a marine algal polyphenol and its anti-hypertensive effect in spontaneously hypertensive rats. Process Biochem. 2017, 58, 326–332. [Google Scholar] [CrossRef]
- Vijayan, R.; Chitra, L.; Penislusshiyan, S.; Palvannan, T. Exploring bioactive fraction of Sargassum wightii: In vitro elucidation of angiotensin-I-converting enzyme inhibition and antioxidant potential. Int. J. Food Prop. 2018, 21, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Nagappan, H.; Pee, P.P.; Kee, S.H.Y.; Ow, J.T.; Yan, S.W.; Chew, L.Y.; Kong, K.W. Malaysian brown seaweeds Sargassum siliquosum and Sargassum polycystum: Low density lipoprotein (LDL) oxidation, angiotensin converting enzyme (ACE)—Amylase, and-glucosidase inhibition activities. Food Res. Int. 2017, 99 Pt 2, 950–958. [Google Scholar] [CrossRef]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Abdelhedi, O.; Nasri, M. Basic and recent advances in marine antihypertensive peptides: Production, structure-activity relationship and bioavailability. Trends Food Sci. Technol. 2019, 88, 543–557. [Google Scholar] [CrossRef]
- Zhou, B.; Carrillo-Larco, R.M.; Danaei, G.; Riley, L.M.; Paciorek, C.J.; Stevens, G.A.; Gregg, E.W.; Bennett, J.E.; Solomon, B.; Singleton, R.K.; et al. Worldwide trends in hypertension prevalence and progress in treatment and control from 1990 to 2019: A pooled analysis of 1201 population-representative studies with 104 million participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef]
- Julius, S.; Nesbitt, S.D.; Egan, B.M.; Weber, M.A.; Michelson, E.L.; Kaciroti, N.; Black, H.R.; Grimm, R.H.; Messerli, F.H.; Oparil, S.; et al. Feasibility of treating prehypertension with an angiotensin-receptor blocker. N. Engl. J. Med. 2006, 354, 1685–1697. [Google Scholar] [CrossRef]
- Bhuyan, B.J.; Mugesh, G. Synthesis, characterization and antioxidant activity of angiotensin converting enzyme inhibitors. Org. Biomol. Chem. 2011, 9, 1356–1365. [Google Scholar] [CrossRef]
- Osterziel, K.J.; Dietz, R.; Harder, K.; Kübler, W. Comparison of captopril with enalapril in the treatment of heart failure: Influence on hemodynamics and measures of renal function. Cardiovasc. Drugs Ther. 1992, 6, 173–180. [Google Scholar] [CrossRef]
- Alan, S.L.A.; Yu, M.B.; Chir, B. Renovascular Hypertension and Ischemic Nephropathy. In Brenner & Rector’s the Kidney; Chertow, G., Luyckx, V., Marsden, P., Skorecki, K., Maarten, M., Yu, A., Eds.; Elsevier, Inc.: Philadelphia, PA, USA, 2020; pp. 1580–1621. [Google Scholar]
- Seca, A.M.L.; Pinto, D.C.G.A. Overview on the Antihypertensive and Anti-Obesity Effects of Secondary Metabolites from Seaweeds. Mar. Drugs 2018, 16, 237. [Google Scholar] [CrossRef] [Green Version]
- Pujiastuti, D.Y.; Ghoyatul Amin, M.N.; Alamsjah, M.A.; Hsu, J.-L. Marine Organisms as Potential Sources of Bioactive Peptides that Inhibit the Activity of Angiotensin I-Converting Enzyme: A Review. Molecules 2019, 24, 2541. [Google Scholar] [CrossRef] [Green Version]
- Soffer, R.L. Angiotensin-Converting Enzyme and the Regulation of Vasoactive Peptides. Annu. Rev. Biochem. 1976, 45, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lee, E.T.; Devereux, R.B.; Yeh, J.; Best, L.G.; Howard, B.V. Prehypertension, diabetes, and cardiovascular disease risk in a population-based sample: The Strong Heart Study. Hypertension 2006, 47, 410–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Bioactive Peptides Derived from Seaweed Protein and Their Health Benefits: Antihypertensive, Antioxidant, and Antidiabetic Properties. J. Food Sci. 2018, 83, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Alashi, A.M.; Young, J.F.; Therkildsen, M.; Aluko, R.E. Enzyme inhibition kinetics and molecular interactions of patatin peptides with angiotensin I-converting enzyme and renin. Int. J. Biol. Macromol. 2017, 101, 207–213. [Google Scholar] [CrossRef]
- Giromini, C.; Fekete, Á.A.; Givens, D.I.; Baldi, A.; Lovegrove, J.A. Short-Communication: A Comparison of the In Vitro Angiotensin-1-Converting Enzyme Inhibitory Capacity of Dairy and Plant Protein Supplements. Nutrients 2017, 9, 1352. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.A.; Hyun, S.K.; Kim, H.R.; Choi, J.S. Angiotensin-converting enzyme I inhibitory activity of phlorotannins from Ecklonia stolonifera. Fish. Sci. 2006, 72, 1292–1299. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.; Ko, S.C.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on angiotensin I-converting enzyme (ACE) inhibitory activity. Nutr Res. Pract. 2011, 5, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Olivares-Molina, A.; Fernández, K. Comparison of different extraction techniques for obtaining extracts from brown seaweeds and their potential effects as angiotensin I-converting enzyme (ACE) inhibitors. J. Appl. Phycol. 2016, 28, 1295–1302. [Google Scholar] [CrossRef]
- Hata, Y.; Nakajima, K.; Uchida, J.-i.; Hidaka, H.; Nakano, T. Clinical Effects of Brown Seaweed, Undaria pinnatifida (wakame), on Blood Pressure in Hypertensive Subjects. J. Clin. Biochem. Nutr. 2001, 30, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Xu, X.; Sun, X.; Zhang, X.; Chen, X.; Xu, N. Preparation and Identification of ACE Inhibitory Peptides from the Marine Macroalga Ulva intestinalis. Mar. Drugs 2019, 17, 79. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Enzymatic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: Essex, UK, 1986. [Google Scholar]
- Muramoto, K.; Sunahara, S.; Kamiya, H. Measurement of tryptophan in peptides by acid hydrolysis in the presence of phenol and its application to the amino acid sequence of a sea anemone toxin. Agric. Biol. Chem. 1987, 51, 1607–1616. [Google Scholar]
- Association of Official Analytical Chemists International (AOAC). Official Methods of Analysis of Association of Official Analytical Chemists International; Association of Official Analytical Chemists: Washington, DC, USA, 1995; Volume 1, p. 870. [Google Scholar]
- Jimenez, R.R.; Ladha, J.K. Automated elemental analysis: A rapid and reliable but expensive measurement of total carbon and nitrogen in plant and soil samples. Commun. Soil Sci. Plant. Anal. 1993, 24, 1897–1924. [Google Scholar] [CrossRef]
- Bikker, P.; Stokvis, L.; van Krimpen, M.M.; van Wikselaar, P.G.; Cone, J.W. Evaluation of seaweeds from marine waters in Northwestern Europe for application in animal nutrition. Anim. Feed Sci. Technol. 2020, 263, 114460. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; O-Nam, K.; Ko, J.-Y.; Lee, J.-H.; Kang, M.-C.; Kim, D.; Lee, J.B.; Lee, J.-S.; Jeon, Y.-J. Purification and identification of novel angiotensin-I converting enzyme (ACE) inhibitory peptides from cultured marine microalgae (Nannochloropsis oculata) protein hydrolysate. J. Appl. Phycol. 2013, 25, 1595–1606. [Google Scholar] [CrossRef]
- Lam le, H.; Shimamura, T.; Sakaguchi, K.; Noguchi, K.; Ishiyama, M.; Fujimura, Y.; Ukeda, H. Assay of angiotensin I-converting enzyme-inhibiting activity based on the detection of 3-hydroxybutyric acid. Anal. Biochem. 2007, 364, 104–111. [Google Scholar] [CrossRef]
- Lam le, H.; Shimamura, T.; Manabe, S.; Ishiyama, M.; Ukeda, H. Assay of angiotensin I-converting enzyme-inhibiting activity based on the detection of 3-hydroxybutyrate with water-soluble tetrazolium salt. Anal. Sci. 2008, 24, 1057–1060. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, J.; Kraan, S.; Guiry, M.D. A Guide to Commercially Important Seaweeds on the Irish Coast; Crofton House, Crofton Road, Dun Laoghaire, Co.: Dublin, Ireland, 2001; p. 67. [Google Scholar]
- Burtin, P. Nutritional Value of Seaweeds. Electron. J. Environ. Agric. Food Chem. 2003, 2, 498–503. [Google Scholar]
- Badmus, U.O.; Taggart, M.A.; Boyd, K.G. The effect of different drying methods on certain nutritionally important chemical constituents in edible brown seaweeds. J. Appl. Phycol. 2019, 31, 3883–3897. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Oba, T.; Yamaguchi, T.; Nakano, T.; Kahara, T.; Funayama, K.; Kobayash, A.; Nakano, T. Antihypertensive effects of hydrolysates of wakame (Undaria pinnatifida) and their angiotensin-I-converting enzyme inhibitory activity. Ann. Nutr. Metab. 2002, 46, 259–267. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Rajauria, G. Chapter 15—Seaweeds: A sustainable feed source for livestock and aquaculture. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 389–420. [Google Scholar]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.-G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kützing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, J.; Meaney, S.; Williams, G.A.; Hayes, M. Extraction of Protein from Four Different Seaweeds Using Three Different Physical Pre-Treatment Strategies. Molecules 2020, 25, 2005. [Google Scholar] [CrossRef]
- PubChem. Compound Summary for CID 614, DL-Proline. National Library of Medicine (US), National Center for Biotechnology Information. 2004. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/DL-Proline (accessed on 5 April 2022).
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Adams, M.A.; Johnsen, P.B.; Zhou, H.-Q. Chemical enhancement of feeding for the herbivorous fish Tilapia zillii. Aquaculture 1988, 72, 95–107. [Google Scholar] [CrossRef]
- Pavadi, P.; Murthy, H.S.; Naik, A.T.R.; Shivakumar, M.; Lakshmipathi, M.T.; Manjappa, N.; Ramesha, T.J.; Shankar, R.; Bank, S. Dietary administration of a feed attractant (Aquasavor) on growth, nutrient utilization and biochemical composition of Freshwater prawn (Macrobrachium rosenbergii). Int. J. Environ. Sci. Technol. 2012, 1, 439–446. [Google Scholar]
- Mommsen, T.P.; French, C.J.; Hochachka, P.W. Sites and patterns of protein and amino acid utilization during the spawning migration of salmon. Can. J. Zool. 1980, 58, 1785–1799. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Food and Drink Federation. Science behind Guideline Daily Amounts; Food and Drink Federation: London, UK, 2009. [Google Scholar]
- De Figueiredo, V.R.G.; Yamashita, F.; Vanzela, A.L.L.; Ida, E.I.; Kurozawa, L.E. Action of multi-enzyme complex on protein extraction to obtain a protein concentrate from okara. J. Food Sci. Technol. 2018, 55, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity, Antioxidant Properties, Phenolic Content and Amino Acid Profiles of Fucus spiralis L. Protein Hydrolysate Fractions. Mar. Drugs 2017, 15, 311. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Maekawa, K.; Chen, J.R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with α-Amylase Inhibitory Potential from Enzymatic Protein Hydrolysates of Red Seaweed (Porphyra spp.). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Cha, S.-H.; Ahn, G.-N.; Heo, S.-J.; Kim, K.-N.; Lee, K.-W.; Song, C.-B.; K Cho, S.-M.; Jeon, Y.-J. Screening of extracts from marine green and brown algae in Jeju for potential marine angiotensin-I converting enzyme (ACE) inhibitory activity. J. Korean Soc. Food Sci. Nutr. 2006, 35, 307–314. [Google Scholar]
- Hori, K.; Matsubara, K.; Fau-Miyazawa, K.; Miyazawa, K. Primary structures of two hemagglutinins from the marine red alga, Hypnea japonica. Biochim Biophys Acta. 2000, 1474, 226–236. [Google Scholar] [CrossRef]
- Thangam, R.; Senthilkumar, D.; Suresh, V.; Sathuvan, M.; Sivasubramanian, S.; Pazhanichamy, K.; Gorlagunta, P.K.; Kannan, S.; Gunasekaran, P.; Rengasamy, R.; et al. Induction of ROS-Dependent Mitochondria-Mediated Intrinsic Apoptosis in MDA-MB-231 Cells by Glycoprotein from Codium decorticatum. J. Agric. Food Chem. 2014, 62, 3410–3421. [Google Scholar] [CrossRef]
- García-Vaquero, M.; Hayes, M. Red and green macroalgae for fish and animal feed and human functional food development. Food Rev. Int. 2016, 32, 15–45. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Chen, J.-C.; Wang, J.; Zheng, B.-D.; Pang, J.; Chen, L.-J.; Lin, H.-t.; Guo, X. Simultaneous Determination of 8 Small Antihypertensive Peptides with Tyrosine at the C-Terminal in Laminaria japonica Hydrolysates by RP-HPLC Method. J. Food Process. Preserv. 2016, 40, 492–501. [Google Scholar] [CrossRef]
- McHugh, D.J. A Guide to the Seaweed Industry; FAO Fisheries: Rome, Italy, 2003. [Google Scholar]
- Van der Meer, J. Limits to food production from the sea. Nat. Food 2020, 1, 762–764. [Google Scholar] [CrossRef]
- Getachew, A.T.; Jacobsen, C.; Holdt, S.L. Emerging Technologies for the Extraction of Marine Phenolics: Opportunities and Challenges. Mar. Drugs 2020, 18, 389. [Google Scholar] [CrossRef]
- Costa, M.M.; Pestana, J.M.; Osório, D.; Alfaia, C.M.; Martins, C.F.; Mourato, M.; Gueifão, S.; Rego, A.M.; Coelho, I.; Coelho, D.; et al. Effect of Dietary Laminaria digitata with Carbohydrases on Broiler Production Performance and Meat Quality, Lipid Profile, and Mineral Composition. Animals 2022, 12, 1007. [Google Scholar] [CrossRef]
- Pihlanto-Leppälä, A.; Koskinen, P.; Piilola, K.; Tupasela, T.; Korhonen, H. Angiotensin I-converting enzyme inhibitory properties of whey protein digests: Concentration and characterization of active peptides. J. Dairy Res. 2000, 67, 53–64. [Google Scholar] [CrossRef]
- Choe, J.; Seol, K.H.; Kim, H.J.; Hwang, J.T.; Lee, M.; Jo, C. Isolation and identification of angiotensin I-converting enzyme inhibitory peptides derived from thermolysin-injected beef M. longissimus. Asian-Australas. J. Anim. Sci. 2019, 32, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Cunha, S.A.; de Castro, R.; Coscueta, E.R.; Pintado, M. Hydrolysate from Mussel Mytilus galloprovincialis Meat: Enzymatic. Hydrolysis, Optimization and Bioactive Properties. Molecules 2021, 26, 5228. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, Q.; Zhang, T.; Liu, M.; Duan, S.; Sun, X. The Antihypertensive Effects and Potential Molecular Mechanism of Microalgal Angiotensin I-Converting Enzyme Inhibitor-like Peptides: A Mini Review. Int J. Mol. Sci. 2021, 22, 4068. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Sample Name | % Protein | SD | % Lipid | SD | % Carbohydrate | SD | % Ash | SD | % Moisture | SD |
|---|---|---|---|---|---|---|---|---|---|---|
| Whole untreated L. digitata (a) | 15.08 | 0.2 | 0.5 | 0.25 | 41.62 | 3.28 | 27.83 | 2.81 | 11.2 | 0.03 |
| L. digitata supernatant fraction recovered following centrifugation (b) | 21.86 | 0.10 | 0.60 | 0.28 | 20.19 | 2.47 | 49.11 | 1.91 | 8.24 | 0.18 |
| L. digitata residual biomass fraction following centrifugation (b) | 14.88 | 0.33 | 0.72 | 0.35 | 50.85 | 2.75 | 27.84 | 1.91 | 5.73 | 0.16 |
| L. digitata permeate fraction recovered following MWCO filtration with a 3-kDa membrane (c) | 23.87 | 0.13 | 0.60 | 0.25 | 14.77 | 1.67 | 52.26 | 0.81 | 8.50 | 0.49 |
| L. digitata retentate fraction recovered following MWCO filtration with a 3-kDa membrane (c) | 18.59 | 0.18 | 1.56 | 0.62 | 31.13 | 4.19 | 38.82 | 1.61 | 9.90 | 1.79 |
| Amino Acid | L. digitata Raw Seaweed (g/100 g) | L. digitata 3-kDa Permeate (g/100 g) |
|---|---|---|
| Alanine | 32.78 | 63.69 |
| Arginine | 7.84 | 2.44 |
| Aspartic acid | 18.37 | 7.99 |
| Glutamic acid | 19.13 | 5.96 |
| Glycine | 9.53 | 3.72 |
| * Histidine | 2.79 | 0.62 |
| * Isoleucine | 7.73 | 3.34 |
| * Leucine | 13.25 | 5.57 |
| * Lysine | 8.92 | 2.99 |
| * Methionine | 1.49 | 0 |
| * Phenylalanine | 7.76 | 0 |
| Proline | 7.94 | 8.63 |
| Serine | 5.78 | 0.68 |
| * Threonine | 5.48 | 0.29 |
| Tyrosine | 2.17 | 0 |
| * Valine | 10.31 | 4.45 |
| Eta (2-aminoethanol) | 0.60 | 0.57 |
| ∑* EAA (%) | 35.67 | 32.22 |
| TAA(g/100 g) | 161.93 | 53.65 |
| Sample Name | IC50 |
|---|---|
| Permeate | 590 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purcell, D.; Packer, M.A.; Hayes, M. Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813. Foods 2022, 11, 1792. https://doi.org/10.3390/foods11121792
Purcell D, Packer MA, Hayes M. Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813. Foods. 2022; 11(12):1792. https://doi.org/10.3390/foods11121792
Chicago/Turabian StylePurcell, Diane, Michael A. Packer, and Maria Hayes. 2022. "Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813" Foods 11, no. 12: 1792. https://doi.org/10.3390/foods11121792
APA StylePurcell, D., Packer, M. A., & Hayes, M. (2022). Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813. Foods, 11(12), 1792. https://doi.org/10.3390/foods11121792