
The Potential of Phaeodactylum as a Natural Source of Antioxidants for Fish Oil Stabilization

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Raw Materials
2.1.2. Chemicals
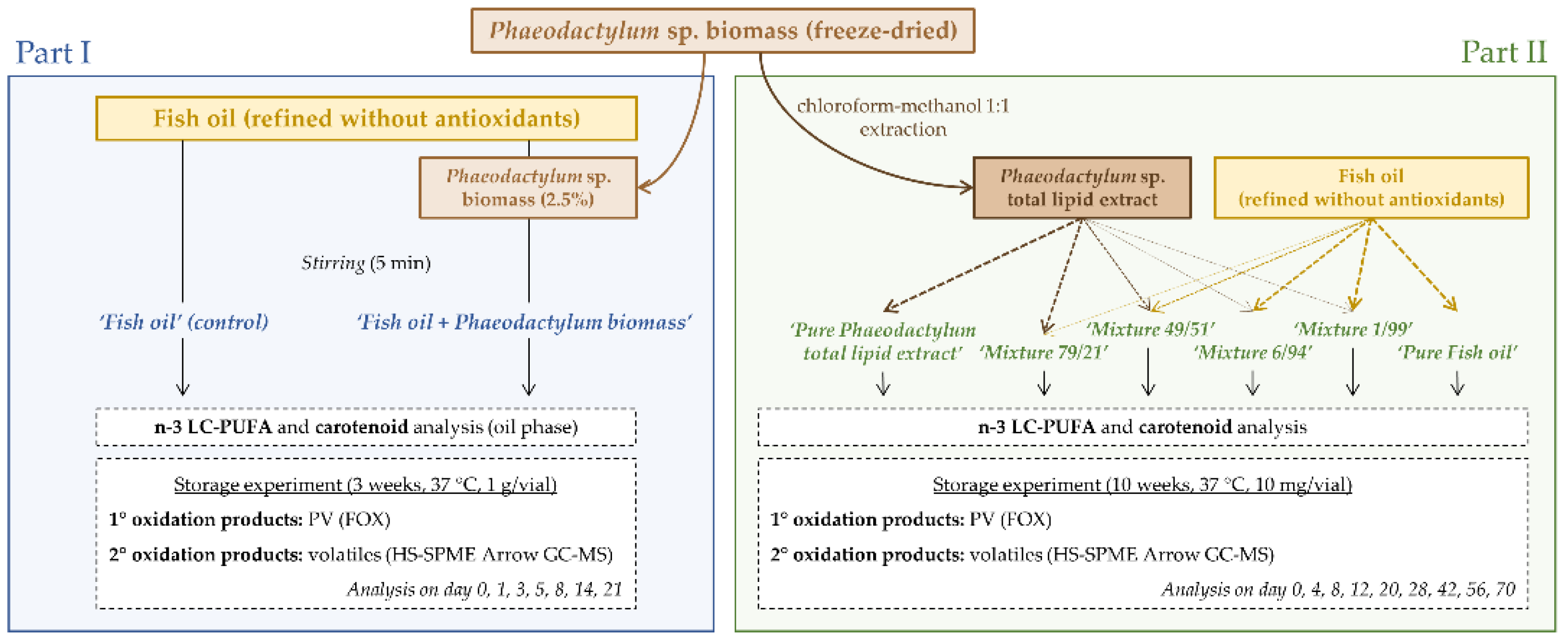
2.2. Experimental Set-Up
2.2.1. Part I
2.2.2. Part II
Phaeodactylum Total Lipid Extract
Preparation and Characterization of the Systems
Oxidative Stability (Storage Experiment)
2.3. Analyses
2.3.1. N-3 LC-PUFA Content
2.3.2. Carotenoids
2.3.3. Primary Oxidation Products: Peroxide Value
2.3.4. Secondary Oxidation Products: Volatile Compounds
2.4. Statistical Analysis
3. Results and Discussion
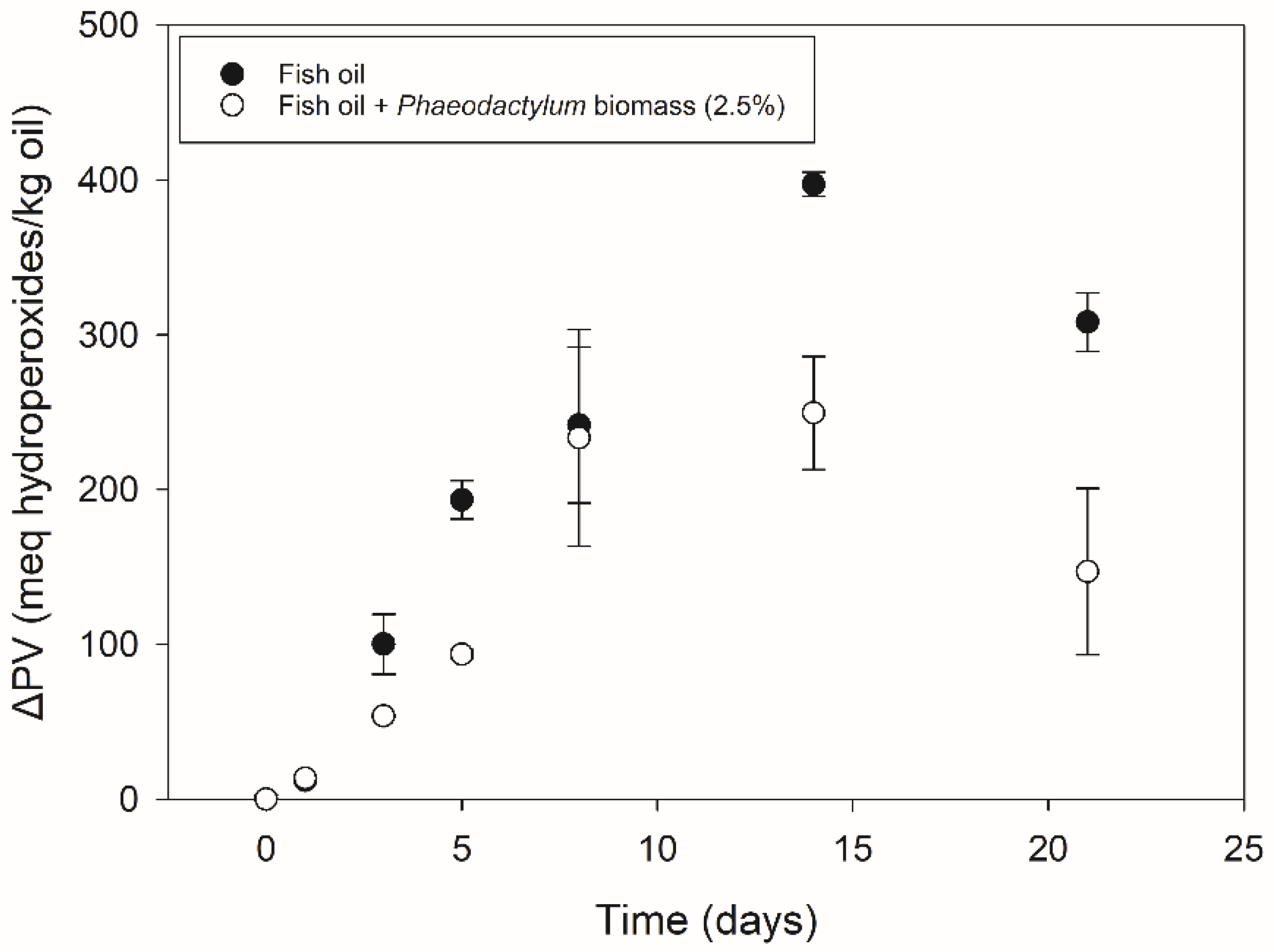
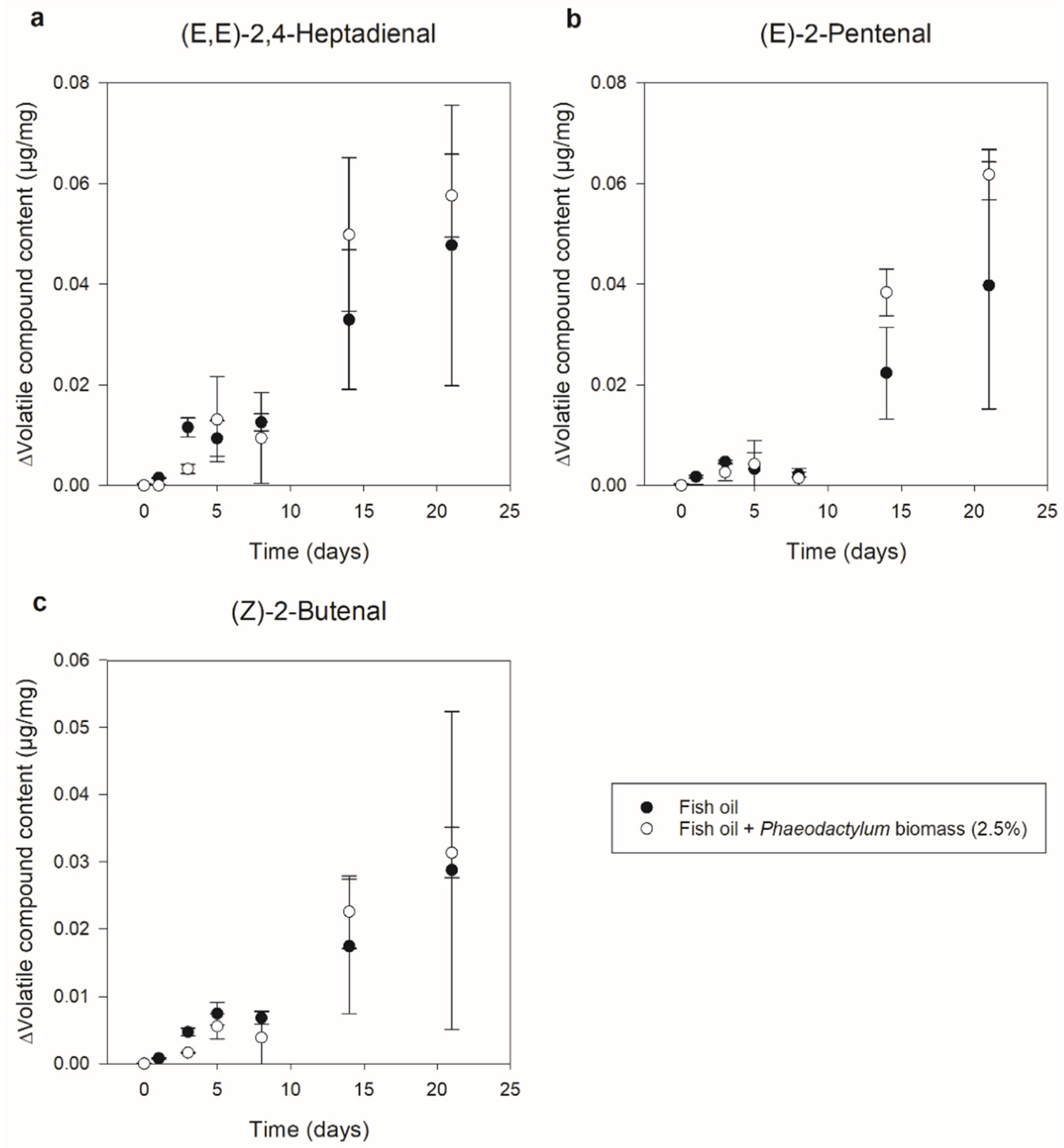
3.1. Phaeodactylum Biomass Addition (Part I)
3.1.1. Primary Oxidation
3.1.2. Secondary Oxidation
3.1.3. Antioxidative Capability
3.1.4. Carotenoid Content
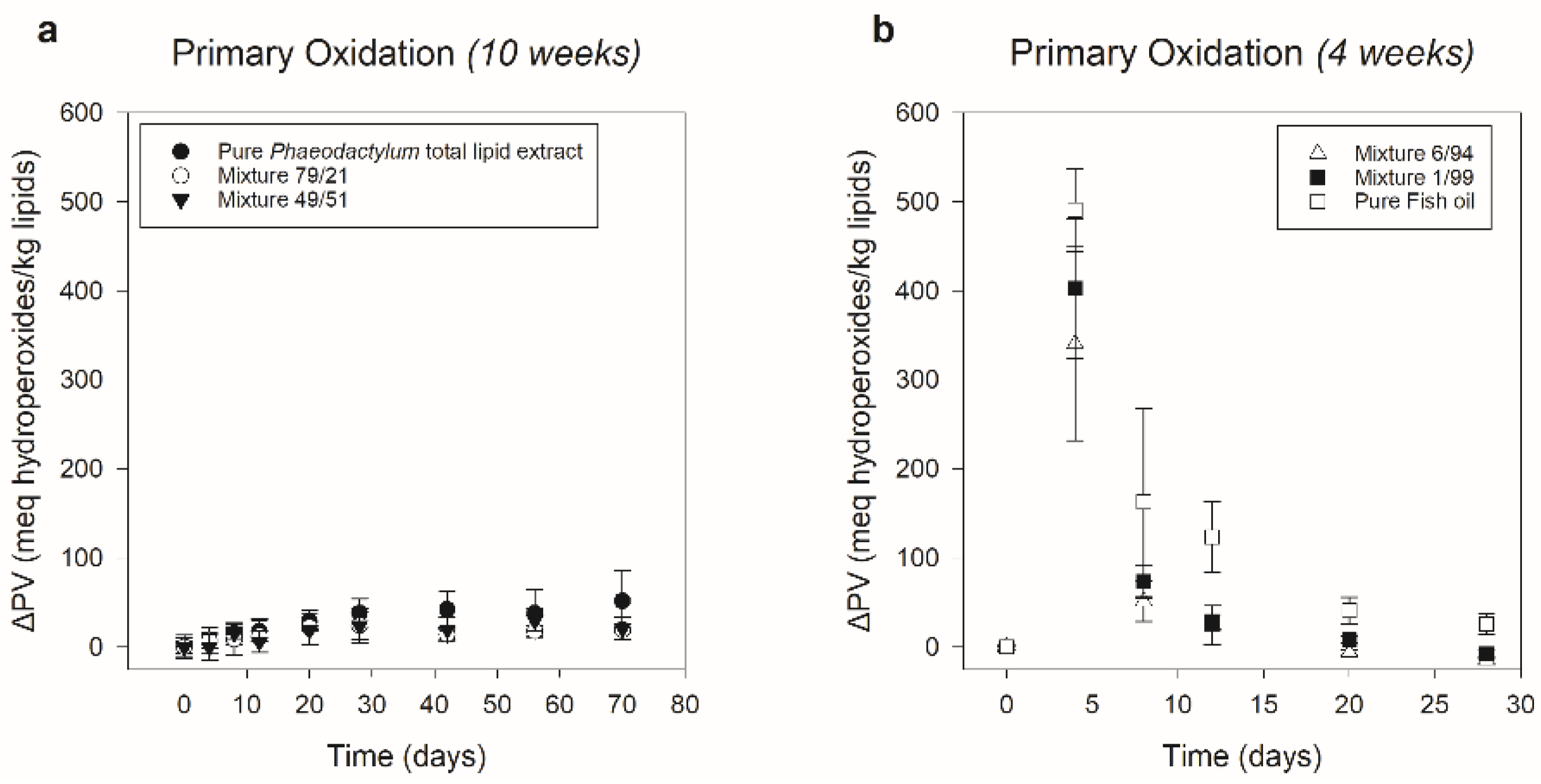
3.2. Phaeodactylum Total Lipid Extract–Fish Oil Mixtures (Part II)
3.2.1. Characterization (Fucoxanthin/n-3 LC-PUFA Ratio)
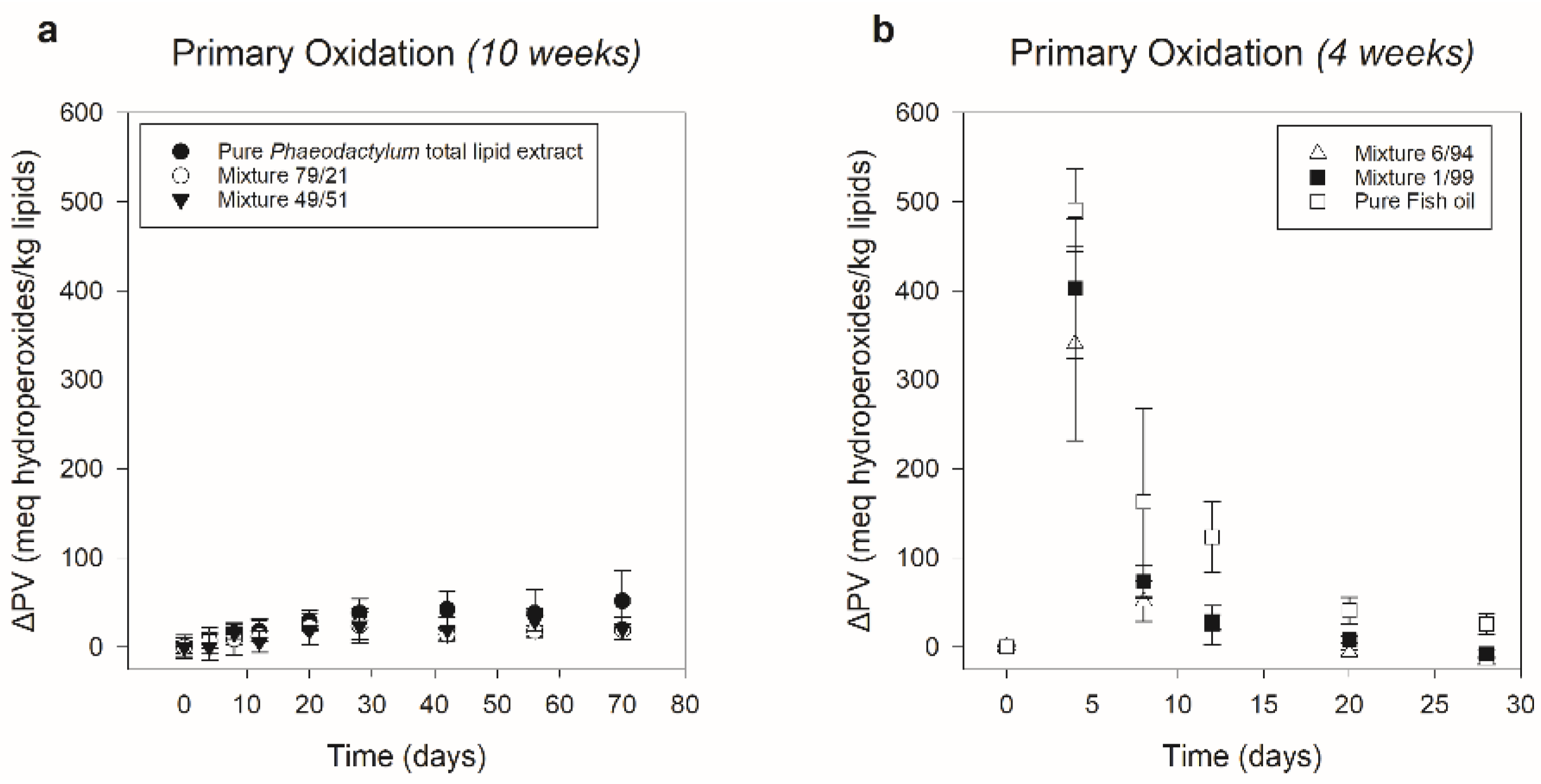
3.2.2. Primary Oxidation
3.2.3. Secondary Oxidation
3.2.4. Oxidative Stability and Antioxidative Capacity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phaeodactylum Total Lipid Extract (mg/g) | Fish Oil (mg/g) | |
|---|---|---|
| Fatty acid composition | ||
| Saturated | 69 (±1) | 226 (±4) |
| C14:0 | 9.1 (±0.6) | 53 (±2) |
| C16:0 | 60 (±1) | 134 (±4) |
| C18:0 | 28.9 (±0.9) | |
| C20:0 | 5.1 (±0.2) | |
| C22:0 | 5.4 (±0.1) | |
| Mono-unsaturated | 103 (±2) | 164 (±3) |
| C16:1 | 94 (±2) | 70 (±2) |
| C18:1 | 9.2 (±0.4) | 80 (±3) |
| C20:1 | 14.8 (±0.5) | |
| Poly-unsaturated | 140 (±1) | 259 (±6) |
| C18:2 | 8.6 (±0.1) | 8.5 (±0.1) |
| C18:3n-3 | 2.8 (±0.1) | 5.6 (±0.1) |
| C20:4n-6 | 9.0 (±0.4) | 7.7 (±0.2) |
| C20:3n-3 | 3.0 (±0.3) | 5.5 (±0.2) |
| C22:2 | 4.7 (±0.2) | |
| n-3 LC-PUFA | 117 (±1) | 227 (±6) |
| C20:5n-3 (EPA) | 106 (±1) | 129 (±5) |
| C22:5n-3 (DPA) | 13.8 (±0.7) | |
| C22:6n-3 (DHA) | 10.7 (±0.2) | 84 (±4) |
| Carotenoid composition | ||
| Fucoxanthin | 32.4 (±0.4) | |
| Diatoxanthin | 2.73 (±0.07) | |
| β-Carotene | 1.6 (±0.2) |
References
- Tocher, D.R. Omega-3 Long-Chain Polyunsaturated Fatty Acids and Aquaculture in Perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020; ISBN 9789251326923. [Google Scholar]
- Calder, P.C. Very Long-Chain n-3 Fatty Acids and Human Health: Fact, Fiction and the Future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Rodríguez, N.; Beltrán, S.; Jaime, I.; de Diego, S.M.; Sanz, M.T.; Carballido, J.R. Production of Omega-3 Polyunsaturated Fatty Acid Concentrates: A Review. Innov. Food Sci. Emerg. Technol. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Frankel, E.N. Lipid Oxidation, 2nd ed.; Oily Press: Bridgewater, UK, 2005; ISBN 9780953194988. [Google Scholar]
- Lima Araújo, K.G.; Domingues, J.R.; Sabaa Srur, A.U.O.; Da Silva, A.J.R. Production of Antioxidants by Anabaena PCC 7119 and Evaluation of Their Protecting Activity against Oxidation of Soybean Oil. Food Biotechnol. 2006, 20, 65–77. [Google Scholar] [CrossRef]
- Hrebień-Filisińska, A. Application of Natural Antioxidants in the Oxidative Stabilization of Fish Oils: A Mini-Review. J. Food Process. Preserv. 2021, 45, e15342. [Google Scholar] [CrossRef]
- Yan, X.; Li, X.; Zhou, C.; Fan, X. Prevention of Fish Oil Rancidity by Phlorotannins from Sargassum Kjellmanianum. J. Appl. Phycol. 1996, 8, 201–203. [Google Scholar] [CrossRef]
- Athukorala, Y.; Lee, K.-W.; Shahidi, F.; Heu, M.S.; Kim, H.-T.; Lee, J.-S.; Jeon, Y.-J. Antioxidant Efficacy of Extracts of an Edible Red Alga (Grateloupia filicina) in Linoleic Acid and Fish Oil. J. Food Lipids 2003, 10, 313–327. [Google Scholar] [CrossRef]
- Kindleysides, S.; Quek, S.Y.; Miller, M.R. Inhibition of Fish Oil Oxidation and the Radical Scavenging Activity of New Zealand Seaweed Extracts. Food Chem. 2012, 133, 1624–1631. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D.; Joseph, D. Changes in the Quality of Refined Fish Oil in an Accelerated Storage Study. J. Aquat. Food Prod. Technol. 2016, 25, 1155–1170. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D. Effects of Antioxidative Substances from Seaweed on Quality of Refined Liver Oil of Leafscale Gulper Shark, Centrophorus squamosus during an Accelerated Stability Study. Food Res. Int. 2018, 103, 450–461. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D. Effect of Antioxidant Compounds from Seaweeds on Storage Stability of C20-22 Polyunsaturated Fatty Acid Concentrate Prepared from Dogfish Liver Oil. Food Chem. 2018, 260, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Golmakani, M.-T.; Moosavi-Nasab, M.; Keramat, M.; Mohammadi, M.-A. Arthrospira platensis Extract as a Natural Antioxidant for Improving Oxidative Stability of Common Kilka (Clupeonella cultriventris caspia) Oil. Turkish J. Fish. Aquat. Sci. 2018, 18, 81–90. [Google Scholar] [CrossRef]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef] [PubMed]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Lemahieu, C.; Muylaert, K.; Van Durme, J.; Goiris, K.; Foubert, I. Stability of Omega-3 LC-PUFA-Rich Photoautotrophic Microalgal Oils Compared to Commercially Available Omega-3 LC-PUFA Oils. J. Agric. Food Chem. 2013, 61, 10145–10155. [Google Scholar] [CrossRef] [PubMed]
- Gheysen, L.; Bernaerts, T.; Bruneel, C.; Goiris, K.; Van Durme, J.; Van Loey, A.; De Cooman, L.; Foubert, I. Impact of Processing on N-3 LC-PUFA in Model Systems Enriched with Microalgae. Food Chem. 2018, 2268, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Gheysen, L.; Demets, R.; Devaere, J.; Bernaerts, T.; Goos, P.; Van Loey, A.; De Cooman, L.; Foubert, I. Impact of Microalgal Species on the Oxidative Stability of N-3 LC-PUFA Enriched Tomato Puree. Algal Res. 2019, 40, 101502. [Google Scholar] [CrossRef]
- Lee, Y.L.; Chuang, Y.C.; Su, H.M.; Wu, F.S. Freeze-Dried Microalgae of Nannochloropsis Oculata Improve Soybean Oil’s Oxidative Stability. Appl. Microbiol. Biotechnol. 2013, 97, 9675–9683. [Google Scholar] [CrossRef]
- Alavi, N.; Golmakani, M.T. Improving Oxidative Stability of Virgin Olive Oil by Addition of Microalga chlorella vulgaris Biomass. J. Food Sci. Technol. 2017, 54, 2464–2473. [Google Scholar] [CrossRef]
- Alavi, N.; Golmakani, M.T. Evaluation of Synergistic Effect of Chlorella vulgaris and Citric Acid on Oxidative Stability of Virgin Olive Oil. Food Sci. Biotechnol. 2017, 26, 901–910. [Google Scholar] [CrossRef]
- Alavi, N.; Golmakani, M.T. Antioxidant Properties of Whole-Cell Spirulina (Arthrospira platensis) Powder Expressed in Olive Oil under Accelerated Storage Conditions. J. Appl. Phycol. 2017, 29, 2971–2978. [Google Scholar] [CrossRef]
- Alavi, N.; Golmakani, M.T. Improving Oxidative Stability of Olive Oil: Incorporation of Spirulina and Evaluation of Its Synergism with Citric Acid. Grasas Aceites 2017, 68, e178. [Google Scholar] [CrossRef] [Green Version]
- Morsy, M.K.; Morsy, O.M.; Elbarbary, H.A.; Saad, M.A. Enhancing of Oxidative Stability and Quality Attributes of Olive Oil Using Spirulina (Arthrospira platensis) Nanoparticles. LWT 2019, 101, 444–455. [Google Scholar] [CrossRef]
- Wang, L.; Yang, B.; Yan, B.; Yao, X. Supercritical Fluid Extraction of Astaxanthin from Haematococcus pluvialis and Its Antioxidant Potential in Sunflower Oil. Innov. Food Sci. Emerg. Technol. 2012, 13, 120–127. [Google Scholar] [CrossRef]
- Rao, A.R.; Sarada, R.; Ravishankar, G.A. Stabilization of Astaxanthin in Edible Oils and Its Use as an Antioxidant. J. Sci. Food Agric. 2007, 87, 957–965. [Google Scholar] [CrossRef]
- Limón, P.; Malheiro, R.; Casal, S.; Acién-Fernández, F.G.; Fernández-Sevilla, J.M.; Rodrigues, N.; Cruz, R.; Bermejo, R.; Pereira, J.A. Improvement of Stability and Carotenoids Fraction of Virgin Olive Oils by Addition of Microalgae Scenedesmus ameriensis Extracts. Food Chem. 2015, 175, 203–211. [Google Scholar] [CrossRef]
- Gouveia, L.; Nobre, B.P.; Marcelo, F.M.; Mrejen, S.; Cardoso, M.T.; Palavra, A.F.; Mendes, R.L. Functional Food Oil Coloured by Pigments Extracted from Microalgae with Supercritical CO2. Food Chem. 2007, 101, 717–723. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty Acid Composition of 12 Microalgae for Possible Use in Aquaculture Feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.H.; Chan, K.W.; Ebrahimi, M. Antioxidant Capacities of Fucoxanthin-Producing Algae as Influenced by Their Carotenoid and Phenolic Contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef]
- Sullivan, J.C.; Budge, S.M.; St-Onge, M. Modeling the Primary Oxidation in Commercial Fish Oil Preparations. Lipids 2011, 46, 87–93. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative Antioxidant Activities of Carotenoids Measured by Ferric Reducing Antioxidant Power (FRAP), ABTS Bleaching Assay (ATEAC), DPPH Assay and Peroxyl Radical Scavenging Assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an Analytical Procedure for Extraction of Lipids from Microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Gheysen, L.; Dejonghe, C.; Bernaerts, T.; Van Loey, A.; De Cooman, L.; Foubert, I. Measuring Primary Lipid Oxidation in Food Products Enriched with Colored Microalgae. Food Anal. Methods 2019, 12, 2150–2160. [Google Scholar] [CrossRef]
- Lee, H.; Kizito, S.A.; Weese, S.J.; Craig-Schmidt, M.C.; Lee, Y.; Wei, C.I.; An, H. Analysis of Headspace Volatile and Oxidized Volatile Compounds in DHA-Enriched Fish Oil on Accelerated Oxidative Storage. J. Food Sci. 2003, 68, 2169–2177. [Google Scholar] [CrossRef]
- Noble, A.C.; Nawar, W.W. The Autoxidation of Highly Unsaturated Fatty Acids: Methyl 4,7,10,13,16,19-Docosahexaenoate. J. Am. Oil Chem. Soc. 1971, 48, 800–803. [Google Scholar] [CrossRef]
- Frankel, E.N. Volatile Lipid Oxidation Products. Prog. Lipid Res. 1982, 22, 1–33. [Google Scholar] [CrossRef]
- Hsieh, R.J.; Kinsella, J.E. Oxidation of Polyunsaturated Fatty Acids: Mechanisms, Products, and Inhibition with Emphasis on Fish. Adv. Food Nutr. Res. 1989, 33, 233–341. [Google Scholar] [CrossRef]
- Frankel, E.N.; Satué-Gracia, T.; Meyer, A.S.; German, J.B. Oxidative Stability of Fish and Algae Oils Containing Long-Chain Polyunsaturated Fatty Acids in Bulk and in Oil-in-Water Emulsions. J. Agric. Food Chem. 2002, 50, 2094–2099. [Google Scholar] [CrossRef]
- Junqueira, H.; Schroder, A.P.; Thalmann, F.; Klymchenko, A.; Mély, Y.; Baptista, M.S.; Marques, C.M. Molecular Organization in Hydroperoxidized POPC Bilayers. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183659. [Google Scholar] [CrossRef]
- Johnson, D.R.; Decker, E.A. The Role of Oxygen in Lipid Oxidation Reactions: A Review. Annu. Rev. Food Sci. Technol. 2015, 6, 171–190. [Google Scholar] [CrossRef]
- Prestegard, S.K.; Erga, S.R.; Steinrücken, P.; Mjøs, S.A.; Knutsen, G.; Rohloff, J. Specific Metabolites in a Phaeodactylum tricornutum Strain Isolated from Western Norwegian Fjord Water. Mar. Drugs 2016, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Pohnert, G.; Boland, W. The Oxylipin Chemistry of Attraction and Defense in Brown Algae and Diatoms. Nat. Prod. Rep. 2002, 19, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| System | Phaeodactylum Total Lipid Extract (Weight%) | Fish Oil (Weight%) |
|---|---|---|
| Pure Phaeodactylum total lipid extract | 100.0 | 0.0 |
| Mixture 79/21 | 78.6 | 21.4 |
| Mixture 49/51 | 49.2 | 50.8 |
| Mixture 6/94 | 6.4 | 93.6 |
| Mixture 1/99 | 0.7 | 99.3 |
| Pure Fish oil | 0.0 | 100.0 |
| Carotenoid | Content (µg/g Oil) | Carotenoid/n-3 LC-PUFA Ratio (w/w) |
|---|---|---|
| Fucoxanthin | 17 (±4) | 0.00007 (±0.00002) |
| Diatoxanthin | 4.4 (±0.7) | 0.000018 (±0.000003) |
| β-carotene | <LOQ (=5) | <0.00002 |
| System | N-3 LC-PUFA (mg/g) | Fucoxanthin (mg/g) | Fucoxanthin/n-3 LC-PUFA Ratio (w/w) |
|---|---|---|---|
| Pure Phaeodactylum total lipid extract | 117 (±1) | 32.4 (±0.4) | 0.277 (±0.003) |
| Mixture 79/21 | 145 (±3) | 24.06 (±0.04) | 0.1663 (±0.0003) |
| Mixture 49/51 | 165 (±4) | 16.7 (±0.3) | 0.101 (±0.002) |
| Mixture 6/94 | 222 (±3) | 1.73 (±0.04) | 0.0078 (±0.0002) |
| Mixture 1/99 | 232 (±3) | 0.211 (±0.004) | 0.00091 (±0.00002) |
| Pure Fish oil | 227 (±6) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demets, R.; Van Broekhoven, S.; Gheysen, L.; Van Loey, A.; Foubert, I. The Potential of Phaeodactylum as a Natural Source of Antioxidants for Fish Oil Stabilization. Foods 2022, 11, 1461. https://doi.org/10.3390/foods11101461
Demets R, Van Broekhoven S, Gheysen L, Van Loey A, Foubert I. The Potential of Phaeodactylum as a Natural Source of Antioxidants for Fish Oil Stabilization. Foods. 2022; 11(10):1461. https://doi.org/10.3390/foods11101461
Chicago/Turabian StyleDemets, Robbe, Simon Van Broekhoven, Lore Gheysen, Ann Van Loey, and Imogen Foubert. 2022. "The Potential of Phaeodactylum as a Natural Source of Antioxidants for Fish Oil Stabilization" Foods 11, no. 10: 1461. https://doi.org/10.3390/foods11101461
APA StyleDemets, R., Van Broekhoven, S., Gheysen, L., Van Loey, A., & Foubert, I. (2022). The Potential of Phaeodactylum as a Natural Source of Antioxidants for Fish Oil Stabilization. Foods, 11(10), 1461. https://doi.org/10.3390/foods11101461