
Substrate Selectivity of a Novel Amylo-α-1,6-glucosidase from Thermococcus gammatolerans STB12


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Expression and Purification of AMY
2.3. Enzyme Activity Assays
2.4. Analysis of Substrate Specificity
2.5. Preparation of the Debranched Dextrin and Starch
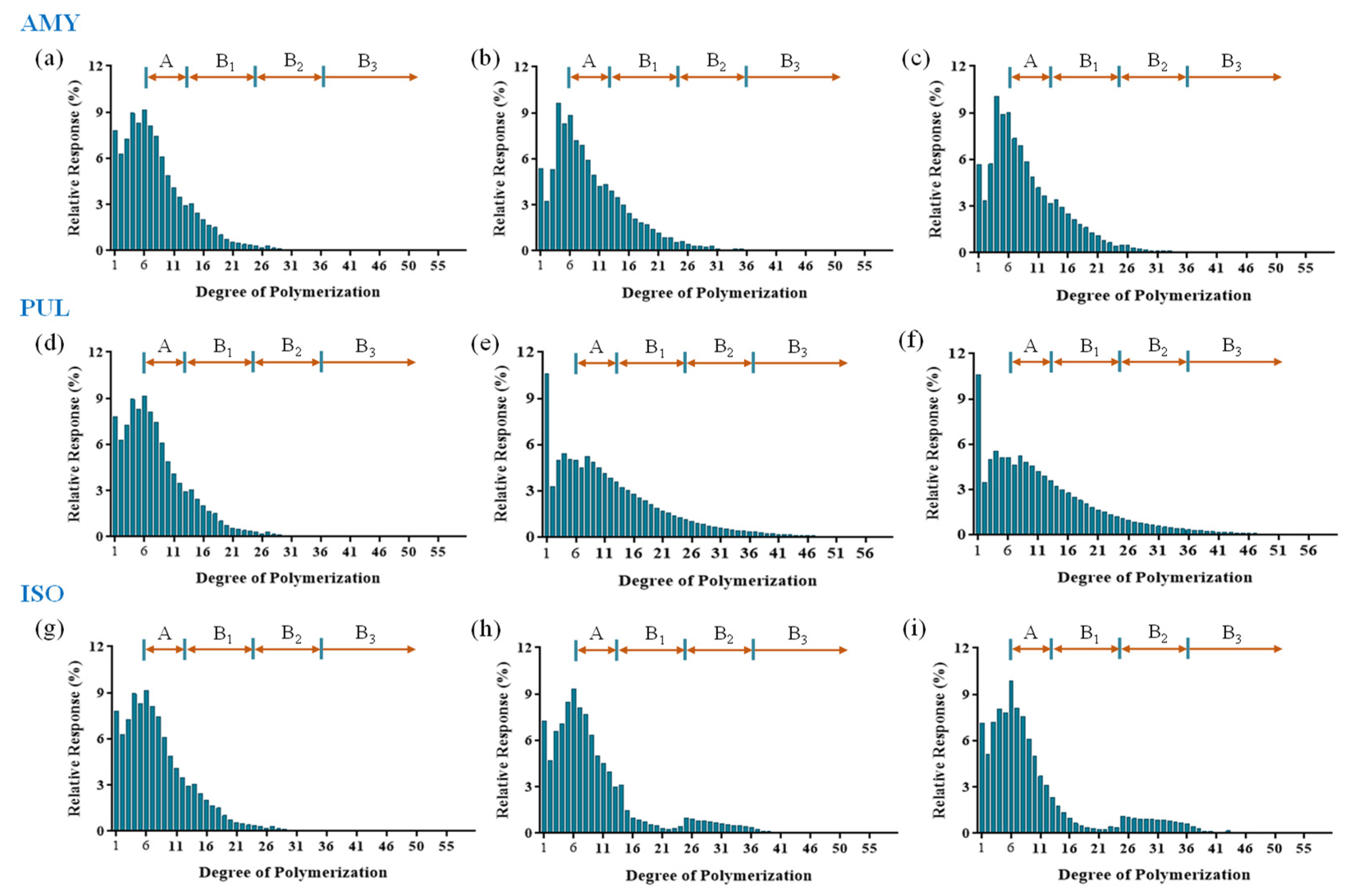
2.6. Chain Length Distribution Analysis
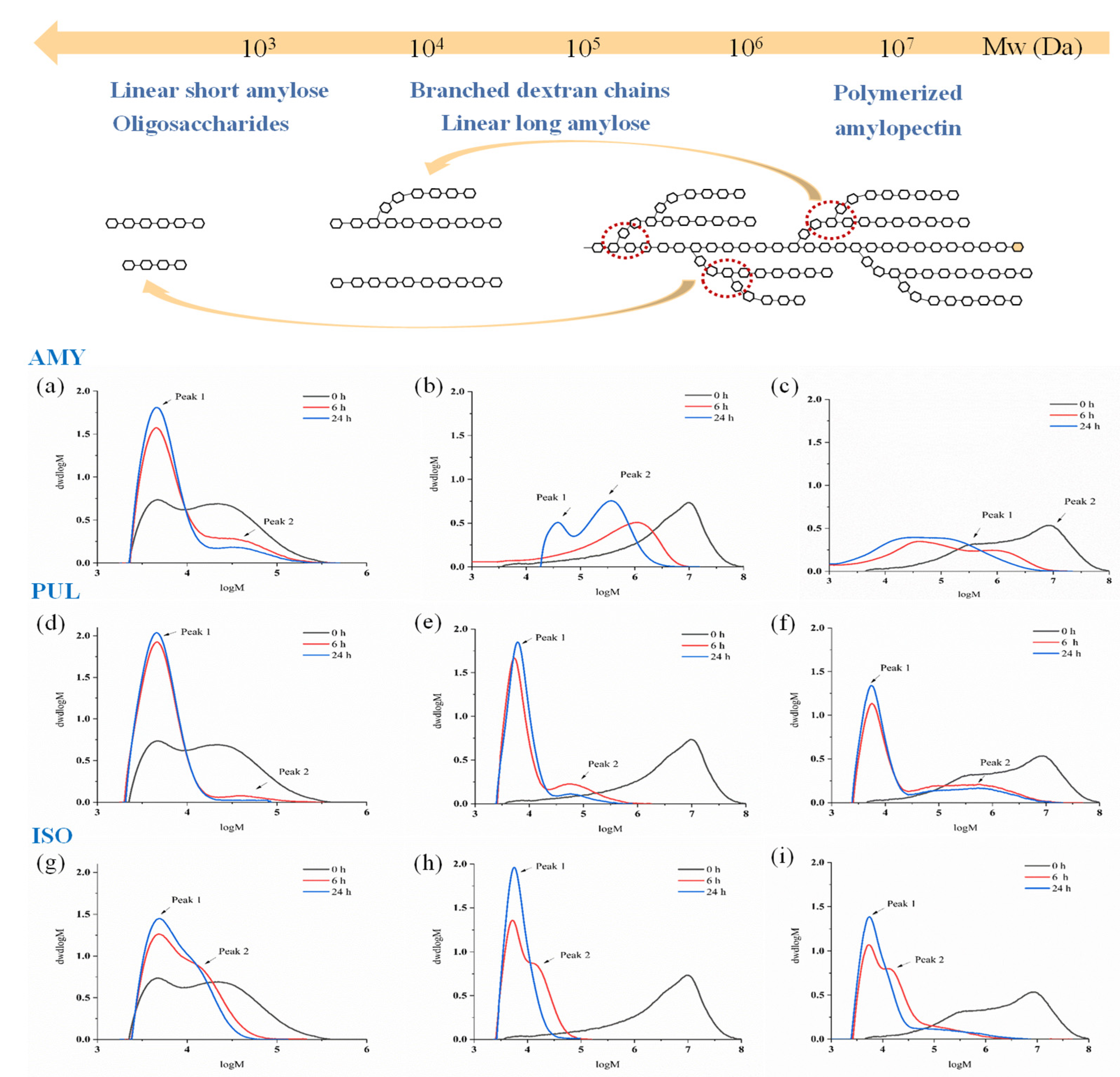
2.7. Molecular Weight Distribution Analysis
2.8. β-Amylolysis Limit Analysis
2.9. Statistical Analysis
3. Results and Discussion
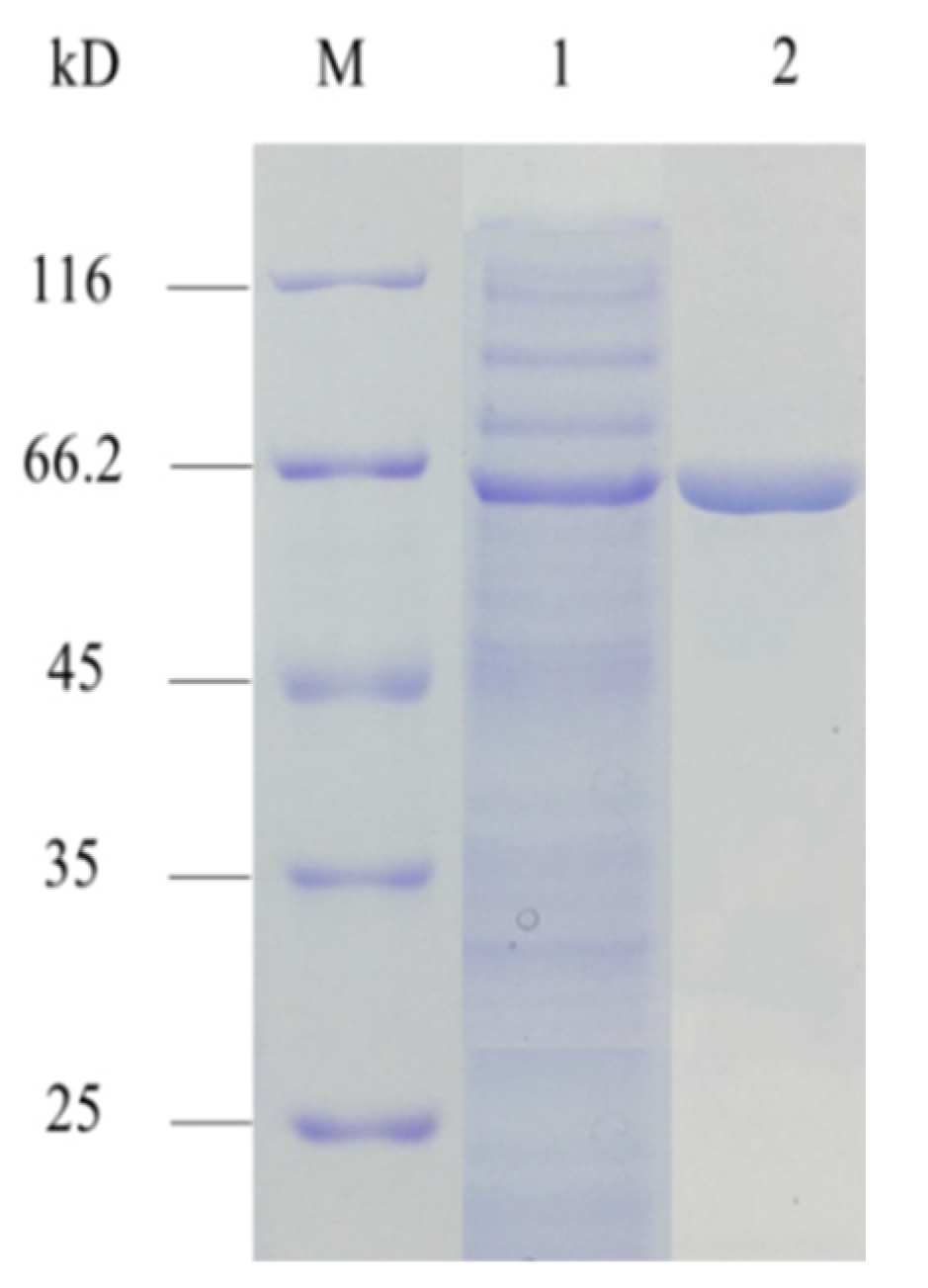
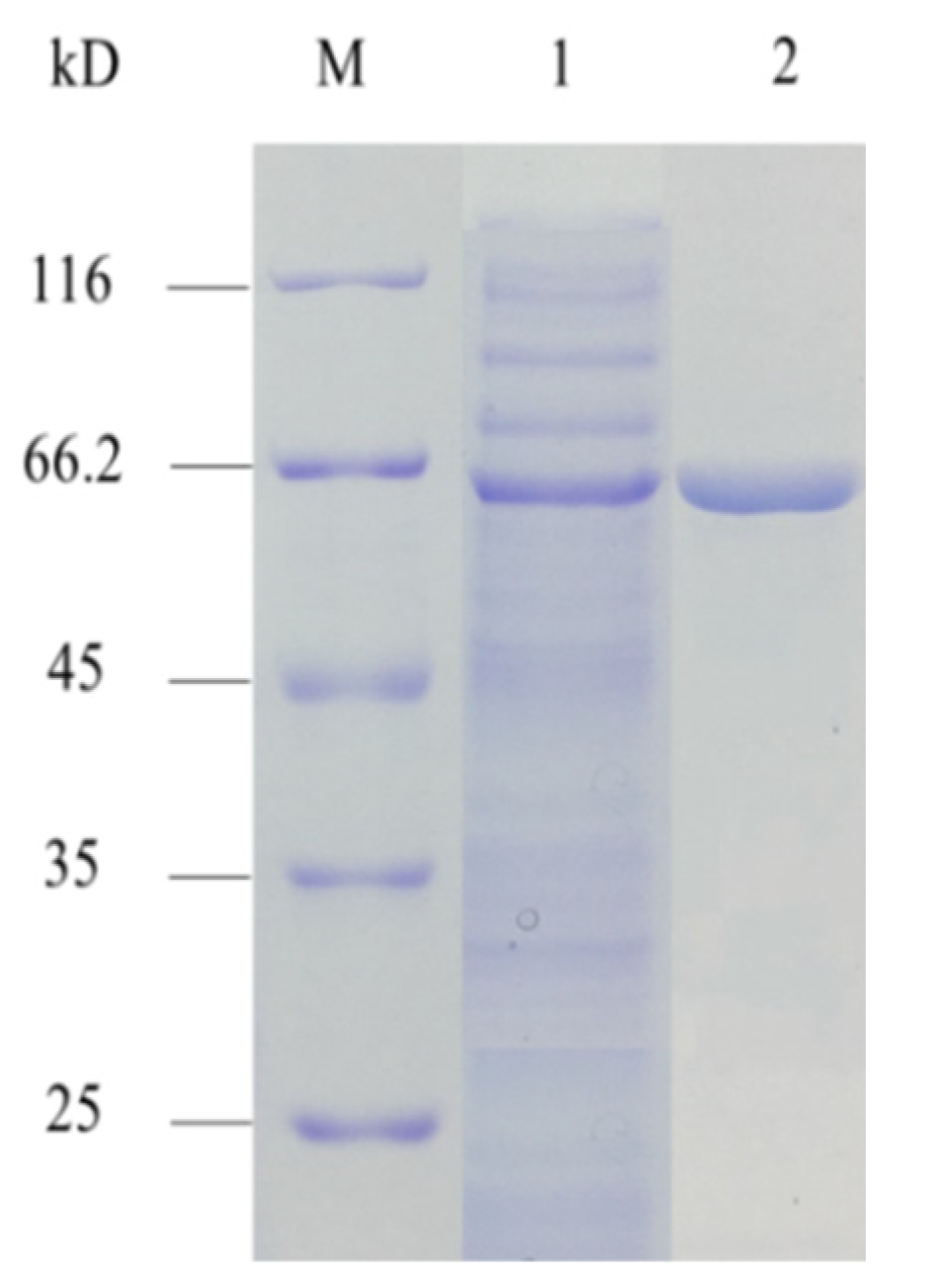
3.1. Expression and Purification of AMY
3.2. Analysis of Substrate Specificity
3.3. Chain Length Distribution
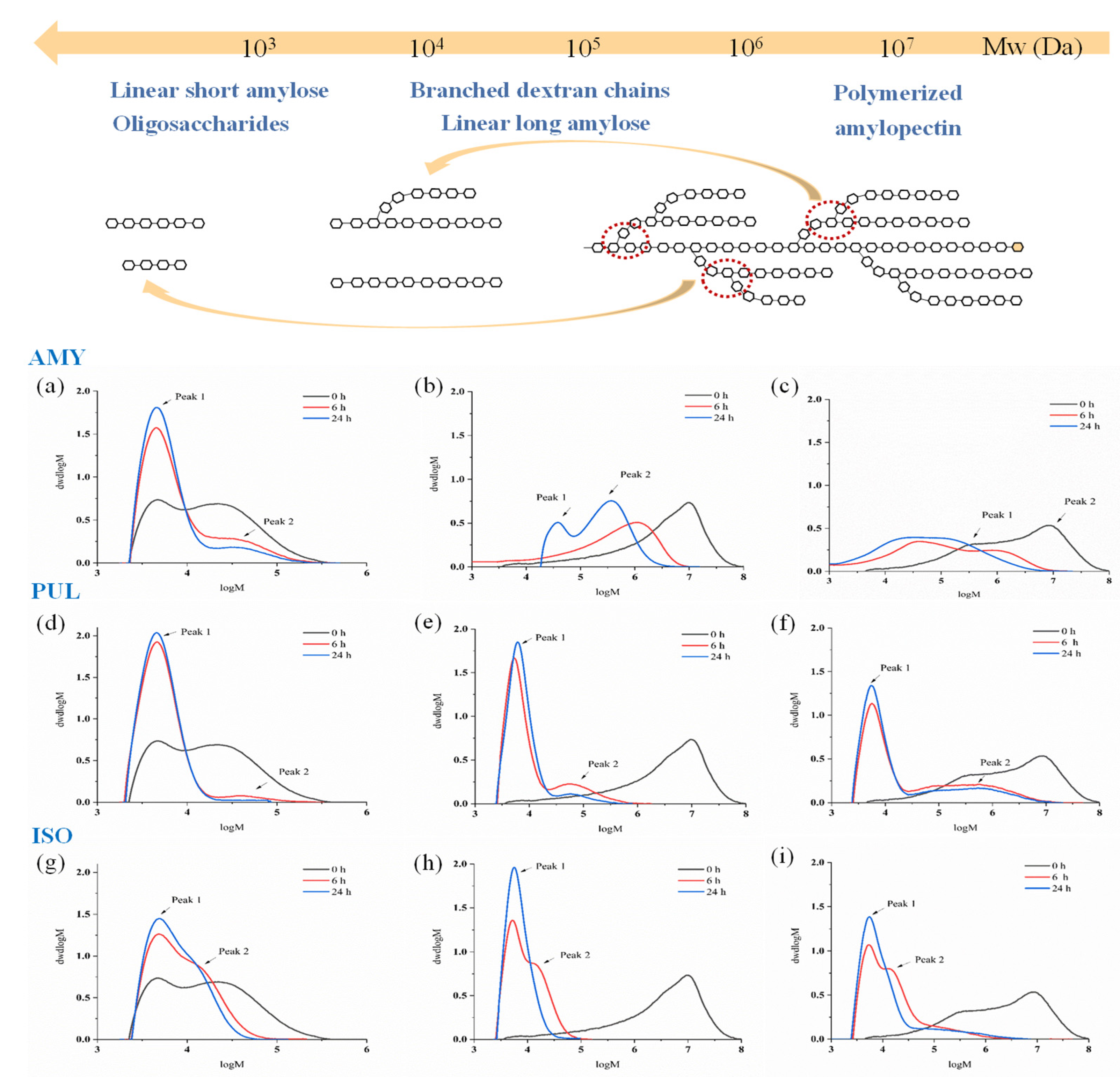
3.4. Molecular Weight Distribution
3.5. Analysis of Debranching Efficiency
3.6. Analysis of β-Amylolysis Limit
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, W.; Yao, N.; Mu, X.; Zhang, R.; Yan, X. Enhancement of glucose production from maltodextrin hydrolysis by optimisation of saccharification process using mixed enzymes involving novel pullulanase. Int. J. Food Sci. Nutr. 2016, 50, 2672–2681. [Google Scholar] [CrossRef]
- Du, Y.; Zang, Y.H.; Du, J. Effects of starch on latex migration and on paper coating properties. Ind. Eng. Chem. Res. 2011, 50, 9781–9786. [Google Scholar] [CrossRef]
- Meshram, M.W.; Patil, V.V.; Mhaske, S.T.; Thorat, B.N. Graft copolymers of starch and its application in textiles. Carbohydr. Polym. 2009, 75, 71–78. [Google Scholar] [CrossRef]
- Maarel, M.; Veen, B.; Uitdehaag, J.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the α-amylase family. J. Bacteriol. 2002, 94, 137–155. [Google Scholar]
- Adrianna, B.; Sylwia, S.; Amin, M.; Krystian, M. Health benefits of resistant starch: A review of the literature. J. Funct. Foods 2022, 93, 105094. [Google Scholar]
- Pandey, A.; Nigam, P.; Soccol, C.R.; Soccol, V.T.; Mohan, R. Advances in microbial amylases. Appl. Biochem. Biotech. 2000, 31, 2135–2152. [Google Scholar] [CrossRef]
- Vamadevan, V.; Bertoft, E. Structure-function relationships of starch components. Starch-Stärke 2015, 67, 55–68. [Google Scholar] [CrossRef]
- Wang, X.; Nie, Y.; Xu, Y. Industrially produced pullulanases with thermostability: Discovery, engineering, and heterologous expression. Bioresour. Technol. 2019, 278, 360–371. [Google Scholar] [CrossRef]
- Jensen, B.F.; Norman, B.E. Bacillus acidopullulyticus pullulanase: Application and regulatory aspects for use in the food industry. Process Biochem. 1984, 19, 129–134. [Google Scholar]
- Poliakoff, M.; Licence, P. Sustainable technology: Green chemistry. Nature 2007, 450, 810–812. [Google Scholar] [CrossRef]
- Duan, X.; Sheng, C.; Jian, C. Enhancing the cyclodextrin production by synchronous utilization of isoamylase and α-CGTase. Appl. Microbiol. Biotechnol. 2013, 97, 3467–3474. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dan, W.; Jian, C. Enhanced production of γ-cyclodextrin by optimization of reaction of γ-cyclodextrin glycosyltransferase as well as synchronous use of isoamylase. Food Chem. 2013, 141, 3072–3076. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Zhang, L.; Ding, Z.; Gu, Z.; Shi, G. Investigation of debranching pattern of a thermostable isoamylase and its application for the production of resistant starch. Carbohydr. Res. 2017, 446–447, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhou, X.; Hu, Y.; Zhang, Z. Resistant starch content and physicochemical properties of non-waxy rice starches modified by pullulanase, heat-moisture treatment, and citric acid. J. Cereal Sci. 2022, 105, 103472. [Google Scholar] [CrossRef]
- Takaku, H. Handbook of Amylase and Related Enzymes: Their Sources, Isolation Methods, Properties and Applications; Pergamon Press: Oxford, UK, 1988; pp. 215–217. [Google Scholar]
- Yokobayashi, K.; Akai, H.; Sugimoto, T.; Hirao, M.; Sugimoto, K.; Harada, T. Comparison of the kinetic parameters of Pseudomonas isoamylase and Aerobacter pullulanase. BBA Bioenergetics 1973, 293, 197–202. [Google Scholar]
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Ariff, A.B. Pullulanase: Role in starch hydrolysis and potential industrial applications. Enzyme Res. 2012, 2012, 921362. [Google Scholar] [CrossRef] [Green Version]
- Satyanarayana, T.; Littlechild, J.; Kawarabayasi, Y. Thermostable Archaeal and Bacterial Pullulanases and Amylopullulanases; Springer: Berlin, Germany, 2013; Volume 10, pp. 535–587. [Google Scholar]
- Harada, T. Isoamylase and its industrial significance in the production of sugars from starch. Biotechnol. Genet. Eng. 1984, 1, 39–64. [Google Scholar] [CrossRef]
- Park, J.T.; Park, H.S.; Kang, H.K.; Hong, J.S.; Cha, H.; Woo, E.J.; Kim, J.W.; Kim, M.J.; Boos, W.; Lee, S. Oligomeric and functional properties of a debranching enzyme (TreX) from the archaeon Sulfolobus solfataricus P2. Biocatal. Biotransformation 2009, 26, 76–85. [Google Scholar] [CrossRef]
- Dauvillee, D.; Kinderf, I.S.; Li, Z.; Kosar-Hashemi, B.; Samuel, M.S.; Rampling, L.; Ball, S.; Morell, M.K. Role of the Escherichia coli glgX gene in glycogen metabolism. J. Bacteriol. 2005, 187, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Kim, H.J.; Kim, Y.H.; Cha, H.; Kim, Y.W.; Kim, T.J.; Kim, Y.R.; Park, K.H.J.C.P. The action mode of Thermus aquaticus YT-1 4-α-glucanotransferase and its chimeric enzymes introduced with starch-binding domain on amylose and amylopectin. Carbohydr. Polym. 2007, 67, 164–173. [Google Scholar] [CrossRef]
- Suzuki, E.; Umeda, K.; Nihei, S.; Moriya, K.; Ohkawa, H.; Fujiwara, S.; Tsuzuki, M. Role of the GlgX protein in glycogen metabolism of the cyanobacterium, synechococcus elongatus PCC 7942. BBA Bioenerg. 2007, 1770, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Park, J.T.; Kang, H.K.; Cha, H.; Kim, D.S.; Kim, J.W.; Park, K.H. TreX from Sulfolobus solfataricus ATCC 35092 displays isoamylase and 4-α-glucanotransferase activities. Biosci. Biotechnol. Biochem. 2007, 71, 1348–1352. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, Y.; Nakamura, Y. Structural and enzymatic characterization of the isoamylase1 homo-oligomer and the isoamylase1–isoamylase2 hetero-oligomer from rice endosperm. Planta 2006, 225, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Mccleary, B.V.; Mcgeough, P.J. A comparison of polysaccharide substrates and reducing sugar methods for the measurement of endo-1,4-β-Xylanase. Appl. Biochem. Biotechnol. 2015, 177, 1152–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Qiu, G.; Zhang, Z.; Ban, X.; Gu, Z.; Li, C.; Hong, Y.; Cheng, L.; Li, Z. Importance of Trp139 in the product specificity of a maltooligosaccharide-forming amylase from Bacillus stearothermophilus STB04. Appl. Microbiol. Biotechnol. 2019, 103, 9433–9442. [Google Scholar] [CrossRef]
- Liu, G.; Hong, Y.; Gu, Z.; Li, Z.; Cheng, L. Pullulanase hydrolysis behaviors and hydrogel properties of debranched starches from different sources. Food Hydrocoll. 2015, 45, 351–360. [Google Scholar] [CrossRef]
- Liu, W.C.; Halley, P.J.; Gilbert, R.G. Mechanism of degradation of starch, a highly branched polymer, during extrusion. Macromolecules 2010, 43, 2855–2864. [Google Scholar] [CrossRef]
- Kong, H.; Yu, L.; Gu, Z.; Li, Z.; Ban, X.; Cheng, L.; Hong, Y.; Li, C. Fine structure impacts highly concentrated starch liquefaction process and product performance. Ind. Crop Prod. 2021, 164, 113347. [Google Scholar] [CrossRef]
- Yu, L.; Kong, H.; Gu, Z.; Li, C.; Ban, X.; Cheng, L.; Hong, Y.; Li, Z. Two 1,4-alpha-glucan branching enzymes successively rearrange glycosidic bonds: A novel synergistic approach for reducing starch digestibility. Carbohydr. Polym. 2021, 262, 117968. [Google Scholar] [CrossRef]
- Shen, X.; Bertoft, E.; Zhang, G.; Hamaker, B.R. Iodine binding to explore the conformational state of internal chains of amylopectin. Carbohydr. Polym. 2013, 98, 778–783. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Q.; Wang, L.; Gilbert, R.G.; Sullivan, M.A. Optimization of liver glycogen extraction when considering the fine molecular structure. Carbohydr. Polym. 2021, 261, 117887. [Google Scholar] [CrossRef] [PubMed]
- Rybicka, K.K. Glycosomes-the organelles of glycogen metabolism. Tissue Cell 1996, 28, 253–265. [Google Scholar] [CrossRef]
- Hanashiro, I.; Abe, J.I.; Hizukuri, S. A periodic distribution of the chain length of amylopectin as revealed by high-performance anion-exchange chromatography. Carbohydr. Res. 1996, 283, 151–159. [Google Scholar] [CrossRef]
- Kuakpetoon, D.; Wang, Y.J. Structural characteristics and physicochemical properties of oxidized corn starches varying in amylose content. Carbohydr. Res. 2006, 341, 1896–1915. [Google Scholar] [CrossRef]
- Charoenkul, N.; Uttapap, D.; Pathipanawat, W.; Takeda, Y. Molecular structure of starches from cassava varieties having different cooked root textures. Starch-Stärke 2006, 58, 443–452. [Google Scholar] [CrossRef]
- Nisha, M.; Satyanarayana, T. Recombinant bacterial amylopullulanases: Developments and perspectives. Bioengineered 2013, 4, 388–400. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Sweedman, M.C.; Shi, Y.C. Structural changes and digestibility of waxy maize starch debranched by different levels of pullulanase. Carbohydr. Polym. 2018, 194, 350–356. [Google Scholar] [CrossRef]
- Cai, L.; Shi, Y.C.; Rong, L.; Hsiao, B.S. Debranching and crystallization of waxy maize starch in relation to enzyme digestibility. Carbohydr. Polym. 2010, 81, 385–393. [Google Scholar] [CrossRef]
- Ji, H.; Li, X.; Bai, Y.; Shen, Y.; Jin, Z. Synergetic modification of waxy maize starch by dual-enzyme to lower the in vitro digestibility through modulating molecular structure and malto-oligosaccharide content. Int. J. Biol. Macromol. 2021, 180, 187–193. [Google Scholar] [CrossRef]
- Wang, Z.; Mhaske, P.; Farahnaky, A.; Kasapis, S.; Majzoobi, M. Cassava starch: Chemical modification and its impact on functional properties and digestibility, a review. Food Hydrocoll. 2022, 129, 107542. [Google Scholar] [CrossRef]
- Hizukuri, S. Relationship between the distribution of the chain length of amylopectin and the crystalline structure of starch granules. Carbohydr. Res. 1985, 141, 295–306. [Google Scholar] [CrossRef]
- Xia, W.; Zhang, K.; Su, L.; Wu, J. Microbial starch debranching enzymes: Developments and applications. Biotechnol. Adv. 2021, 50, 107786. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L. Structures and properties of commercial maltodextrins from corn, potato, and rice starches. Starch-Stärke 2021, 52, 8–9. [Google Scholar] [CrossRef]
- Wu, C.; Zhou, X.; Tian, Y.; Xu, X.; Jin, Z. Hydrolytic mechanism of α-maltotriohydrolase on waxy maize starch and retrogradation properties of the hydrolysates. Food Hydrocoll. 2017, 66, 136–143. [Google Scholar] [CrossRef]





{kind=link}
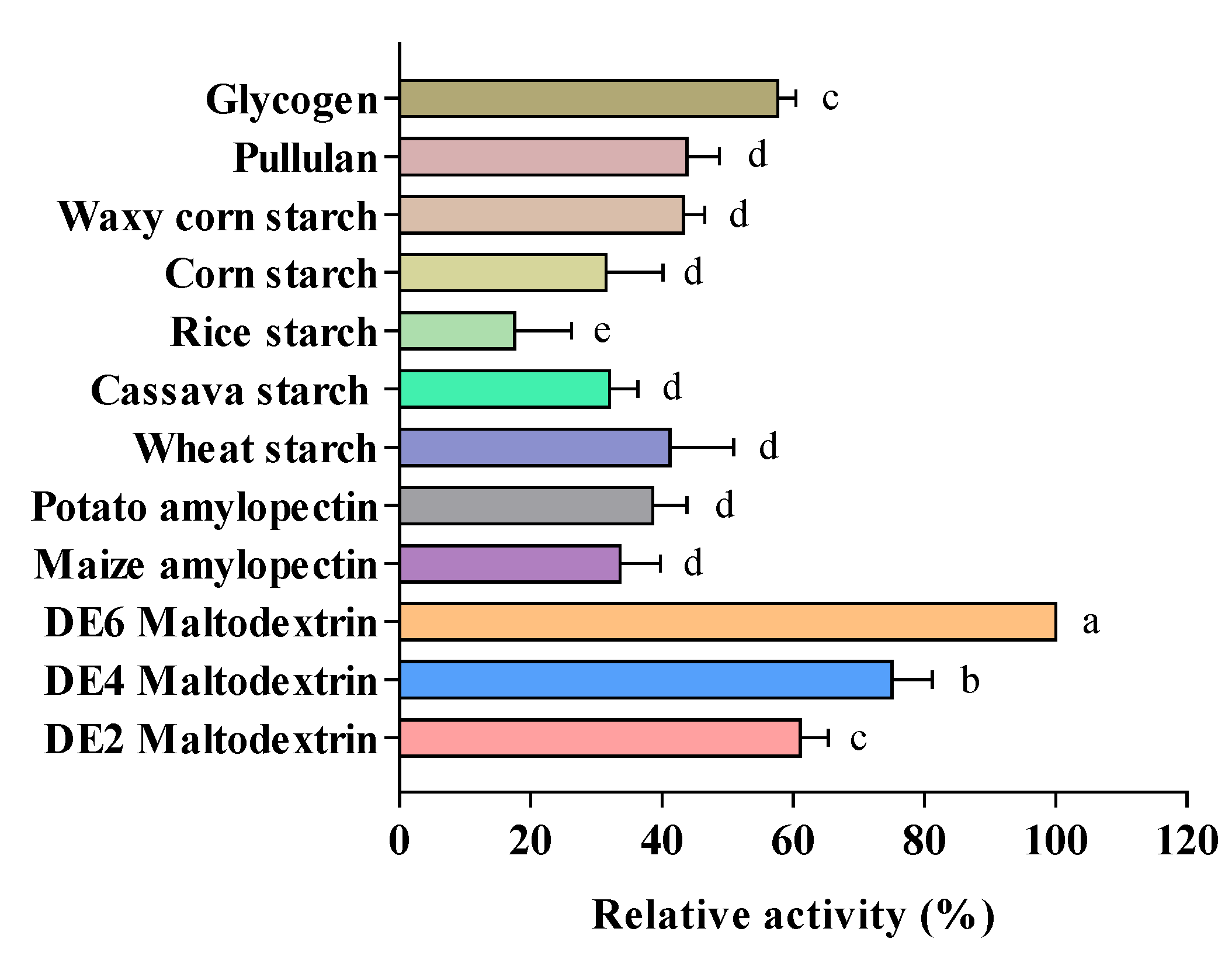{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Total Enzyme Activity (U) | Total Protein Content (mg) | Specific Enzyme Activity (U/mg) | Recovery Rate (%) |
|---|---|---|---|---|
| Crude enzyme | 9853.0 | 42.0 | 234.6 | - |
| HisTrap | 3666.5 | 5.0 | 733.3 | 37.2 |
| Sample | Time | Chain Length Distribution (%) 1 | CL 3 | ||||
|---|---|---|---|---|---|---|---|
| DP 2 < 6 | DP 6–12 | DP 13–24 | DP 25–36 | DP ≥ 37 | |||
| AMY | 0 h | 38.57 ± 0.27 a | 43.20 ± 0.15 b | 17.09 ± 0.34 d | 1.13 ± 0.12 e | 0.05 ± 0.01 d | 7.97 ± 0.27 c |
| 6 h | 31.74 ± 0.65 d | 42.20 ± 0.48 c | 23.12 ± 0.22 b | 2.76 ± 0.17 e | 0.19 ± 0.02 d | 9.27 ± 0.41 b | |
| 24 h | 33.68 ± 0.37 c | 41.89 ± 0.61 c | 21.88 ± 0.27 c | 2.39 ± 0.19 d | 0.14 ± 0.01 d | 9.37 ± 0.39 b | |
| PUL | 0 h | 38.57 ± 0.27 a | 43.20 ± 0.15 b | 17.09 ± 0.34 d | 1.13 ± 0.12 d | 0.05 ± 0.01 d | 7.97 ± 0.27 c |
| 6 h | 29.36 ± 0.31 e | 32.16 ± 0.28 d | 27.53 ± 0.16 a | 8.22 ± 0.47 b | 2.76 ± 0.38 a | 12.02 ± 0.62 a | |
| 24 h | 28.90 ± 0.42 e | 32.41 ± 0.37 d | 27.53 ± 0.39 a | 8.30 ± 0.27 b | 2.88 ± 0.19 a | 12.11 ± 0.50 a | |
| ISO | 0 h | 38.57 ± 0.27 a | 43.20 ± 0.15 b | 17.09 ± 0.34 d | 1.13 ± 0.12 e | 0.05 ± 0.01 d | 7.97 ± 0.27 c |
| 6 h | 34.10 ± 0.67 c | 44.94 ± 0.47 a | 12.45 ± 0.37 e | 7.76 ± 0.19 c | 0.75 ± 0.01 c | 9.33 ± 0.47 b | |
| 24 h | 35.29 ± 0.52 b | 43.45 ± 0.38 b | 9.61 ± 0.21 f | 10.27 ± 0.23 a | 1.38 ± 0.02 b | 9.86 ± 0.34 b | |
| Sample | Time | DE6 Maltodextrin | Waxy Corn Starch | Cassava Starch | |||
|---|---|---|---|---|---|---|---|
| Peak1 MW 1 (×103) | Peak2 MW (×104) | Peak1 MW (×105) | Peak2 MW (×106) | Peak1 MW (×105) | Peak2 MW (×106) | ||
| AMY | 0 h | 6.21 ± 0.13 | 5.30 ± 0.21 | n.d. 2 | 7.44 ± 0.67 | 3.40 ± 0.57 | 9.84 ± 0.76 |
| 6 h | 5.81 ± 0.16 | 5.24 ± 0.19 | 0.57 ± 0.09 | 2.16 ± 0.11 | 0.58 ± 0.86 | 1.28 ± 0.22 | |
| 24 h | 5.31 ± 0.11 | 4.26 ± 0.15 | 0.36 ± 0.13 | 0.94 ± 0.34 | 0.39 ± 0.23 | 0.94 ± 0.34 | |
| PUL | 0 h | 6.21 ± 0.13 | 5.30 ± 0.21 | n.d. | 7.44 ± 0.67 | 3.40 ± 0.57 | 9.84 ± 0.76 |
| 6 h | 5.48 ± 0.23 | 5.31 ± 0.18 | 0.07 ± 0.05 | 0.11 ± 0.08 | 0.07 ± 0.02 | 0.90 ± 0.42 | |
| 24 h | 5.51 ± 0.27 | n.d. | 0.07 ± 0.03 | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.87 ± 0.22 | |
| ISO | 0 h | 6.21 ± 0.13 | 5.30 ± 0.21 | n.d. | 7.44 ± 0.67 | 3.40 ± 0.57 | 9.84 ± 0.76 |
| 6 h | 5.52 ± 0.17 | 1.97 ± 0.24 | 0.01 ± 0.00 | 0.02 ± 0.01 | 0.09 ± 0.07 | 0.42 ± 0.15 | |
| 24 h | 5.53 ± 0.22 | 1.66 ± 0.35 | 0.01 ± 0.00 | n.d. | 0.06 ± 0.22 | 0.08 ± 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Tian, Y.; Ban, X.; Li, C.; Hong, Y.; Cheng, L.; Gu, Z.; Li, Z. Substrate Selectivity of a Novel Amylo-α-1,6-glucosidase from Thermococcus gammatolerans STB12. Foods 2022, 11, 1442. https://doi.org/10.3390/foods11101442
Wang Y, Tian Y, Ban X, Li C, Hong Y, Cheng L, Gu Z, Li Z. Substrate Selectivity of a Novel Amylo-α-1,6-glucosidase from Thermococcus gammatolerans STB12. Foods. 2022; 11(10):1442. https://doi.org/10.3390/foods11101442
Chicago/Turabian StyleWang, Yamei, Yixiong Tian, Xiaofeng Ban, Caiming Li, Yan Hong, Li Cheng, Zhengbiao Gu, and Zhaofeng Li. 2022. "Substrate Selectivity of a Novel Amylo-α-1,6-glucosidase from Thermococcus gammatolerans STB12" Foods 11, no. 10: 1442. https://doi.org/10.3390/foods11101442
APA StyleWang, Y., Tian, Y., Ban, X., Li, C., Hong, Y., Cheng, L., Gu, Z., & Li, Z. (2022). Substrate Selectivity of a Novel Amylo-α-1,6-glucosidase from Thermococcus gammatolerans STB12. Foods, 11(10), 1442. https://doi.org/10.3390/foods11101442