
Tieguanyin Oolong Tea Extracts Alleviate Behavioral Abnormalities by Modulating Neuroinflammation in APP/PS1 Mouse Model of Alzheimer’s Disease

 , ,
, ,  ,
,  ,
,
Abstract
:
1. Introduction
2. Results
2.1. Major Chemical Components in Three Different Types of Tieguanyin Extracts
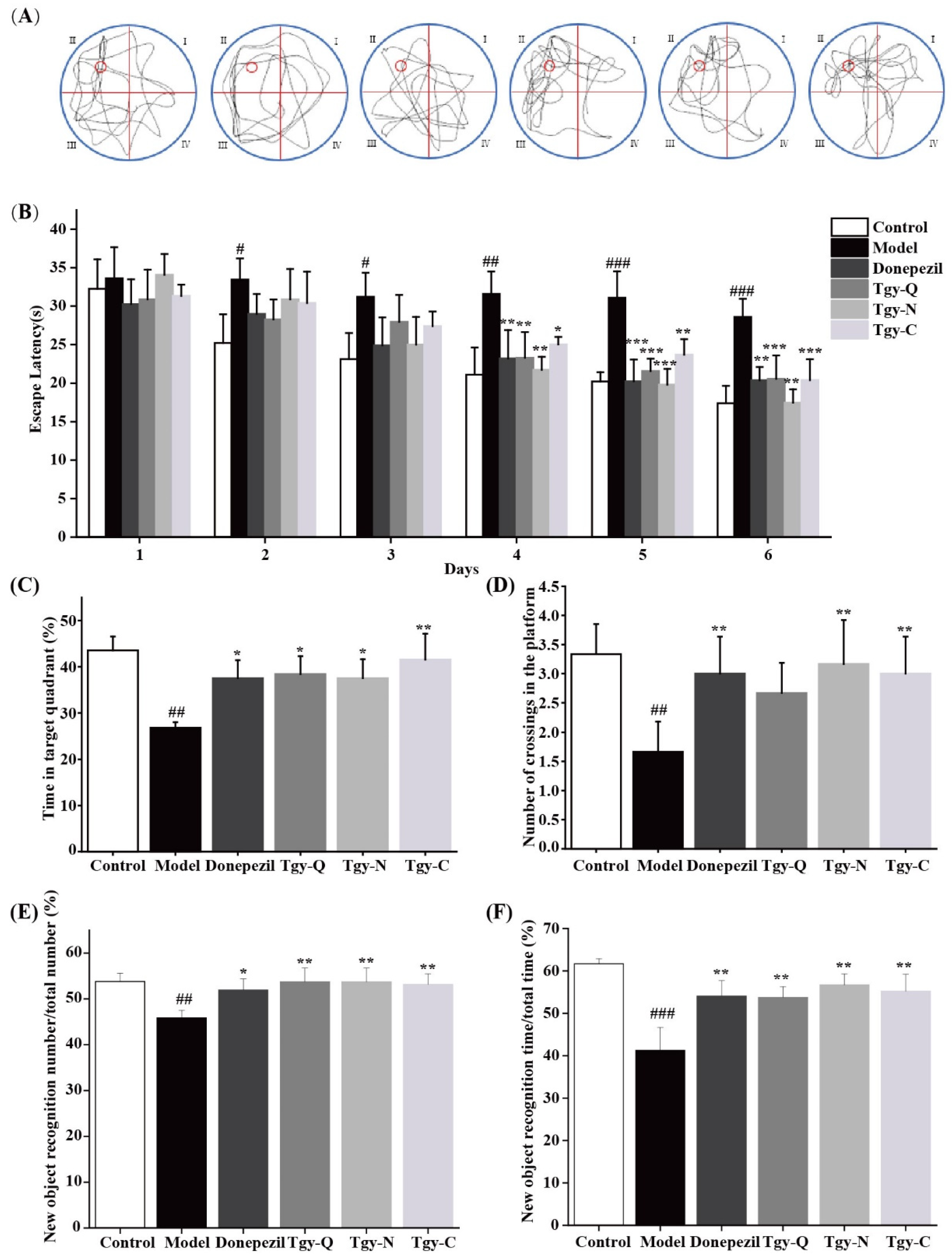
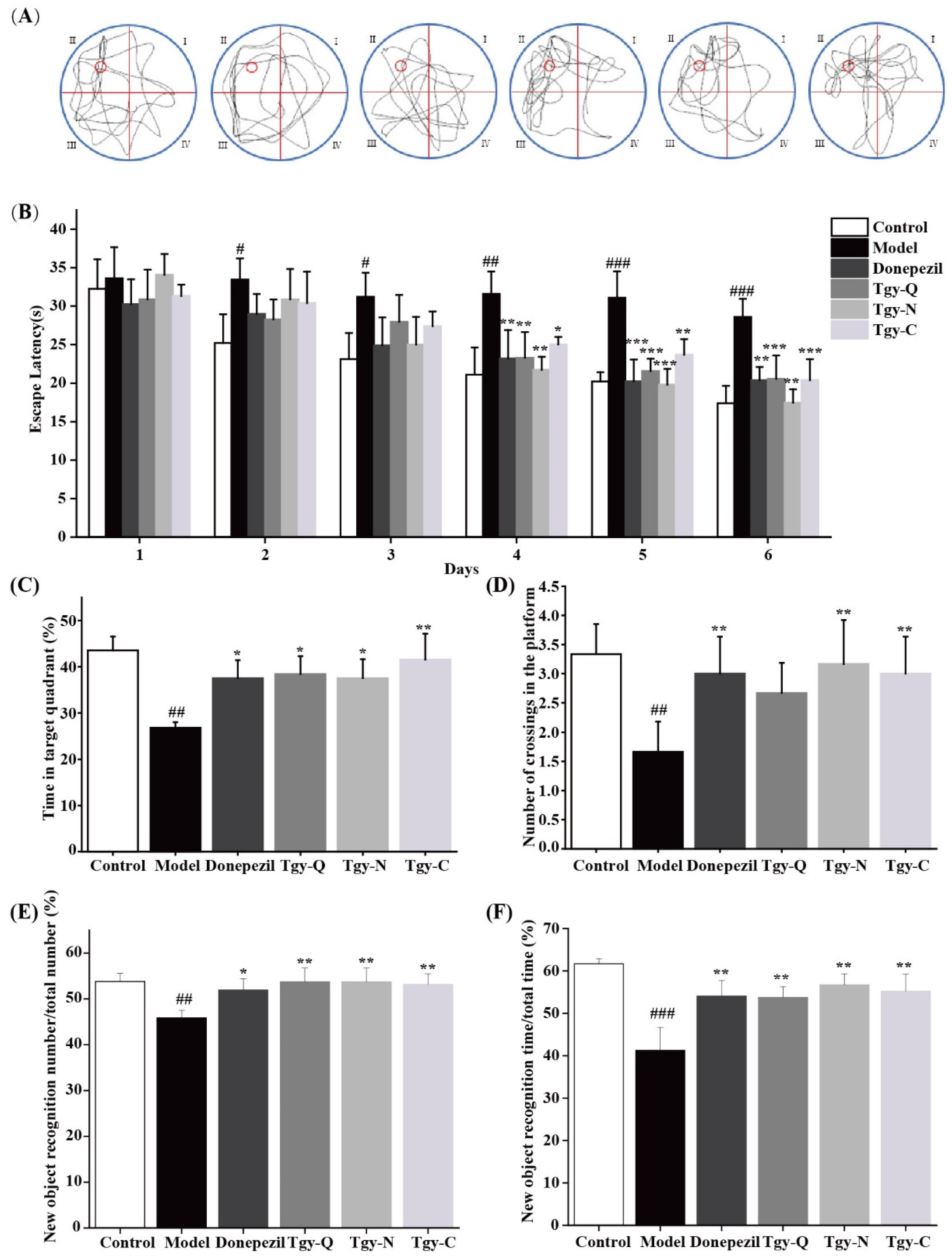
2.2. Tieguanyin Extracts Improves Cognitive Ability in APP/PS1 Mice
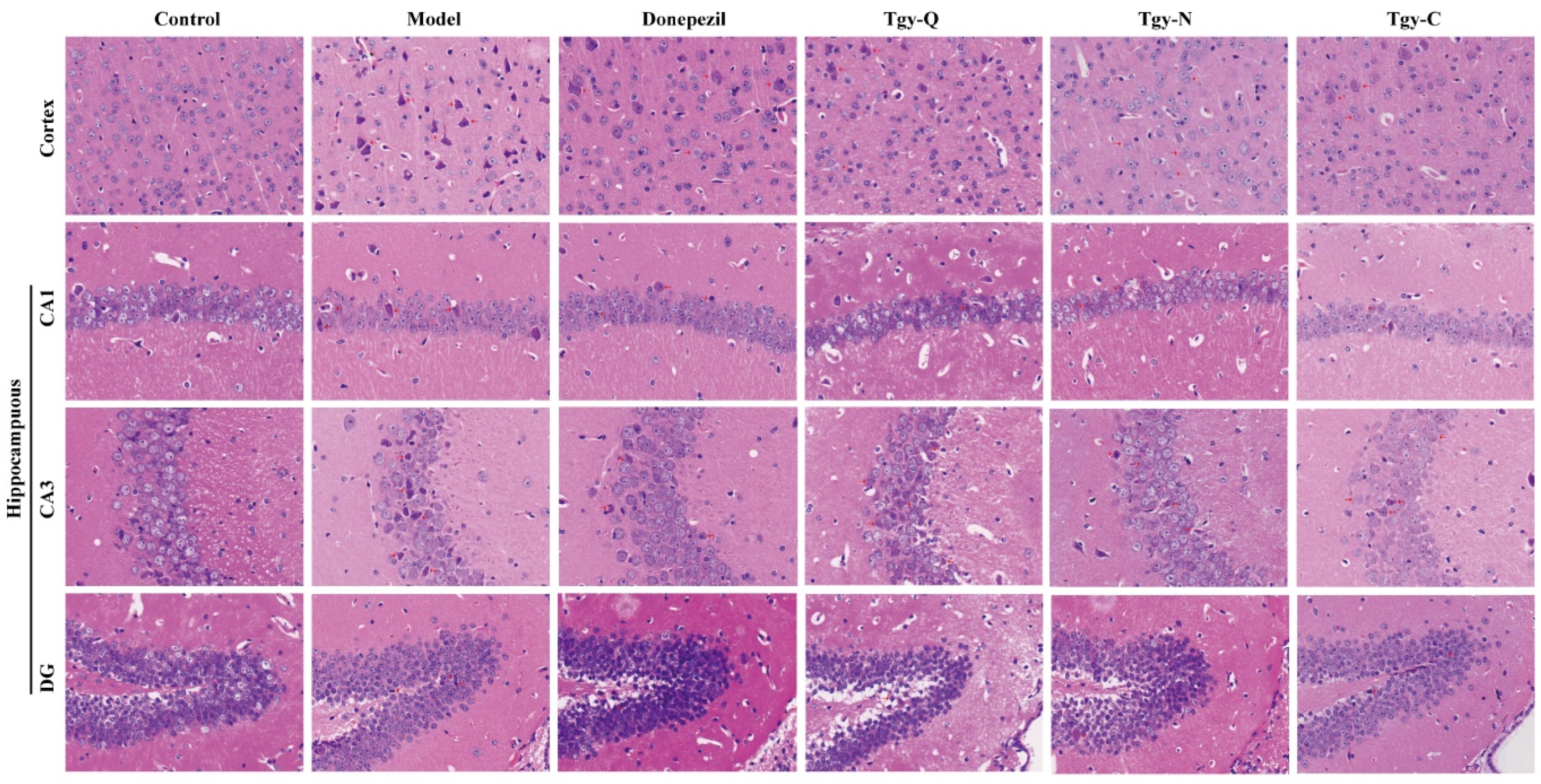
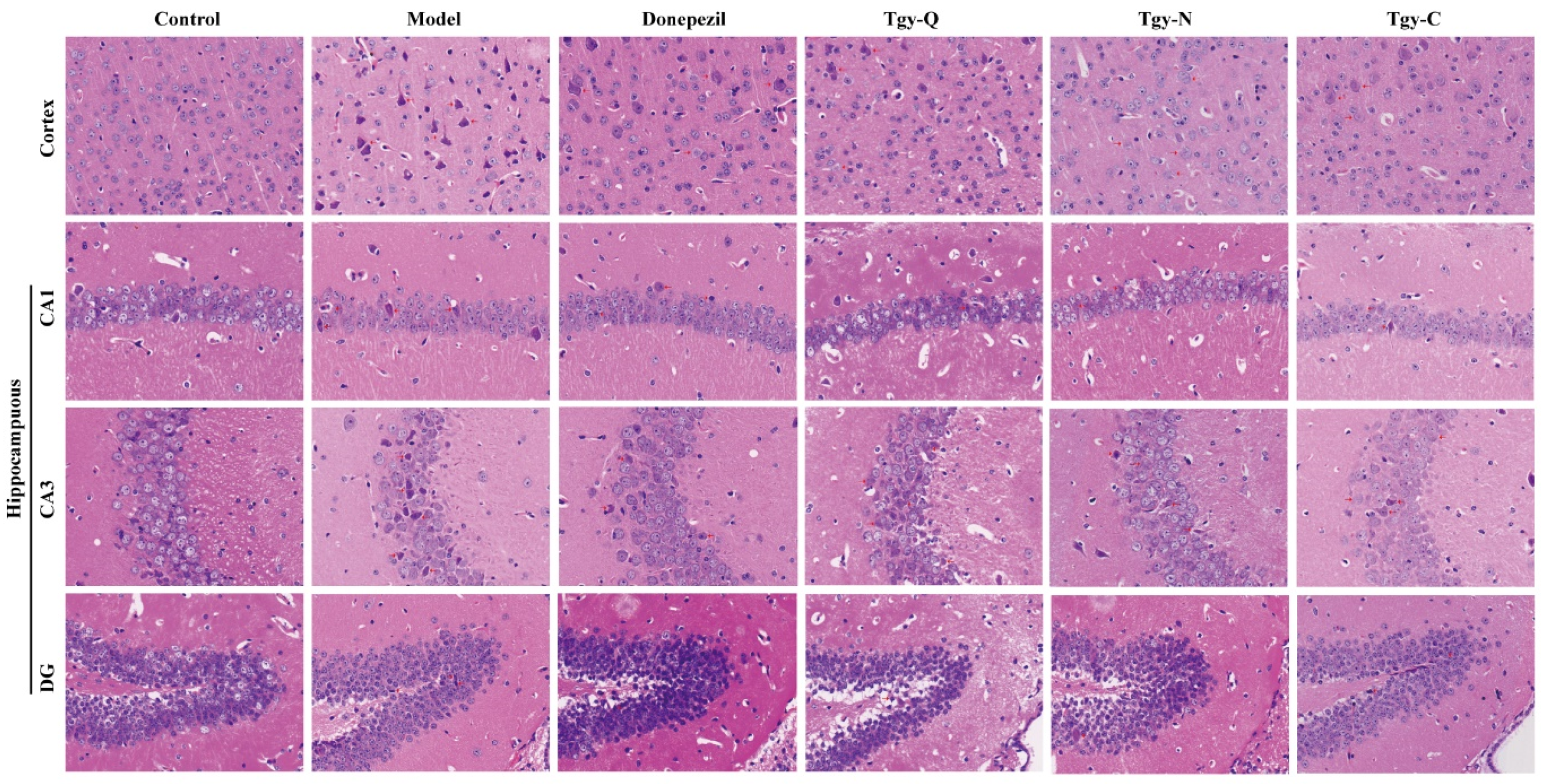
2.3. Tieguanyin Extracts Improves the Pathology of the Cortex and Hippocampus of APP/PS1 Mice
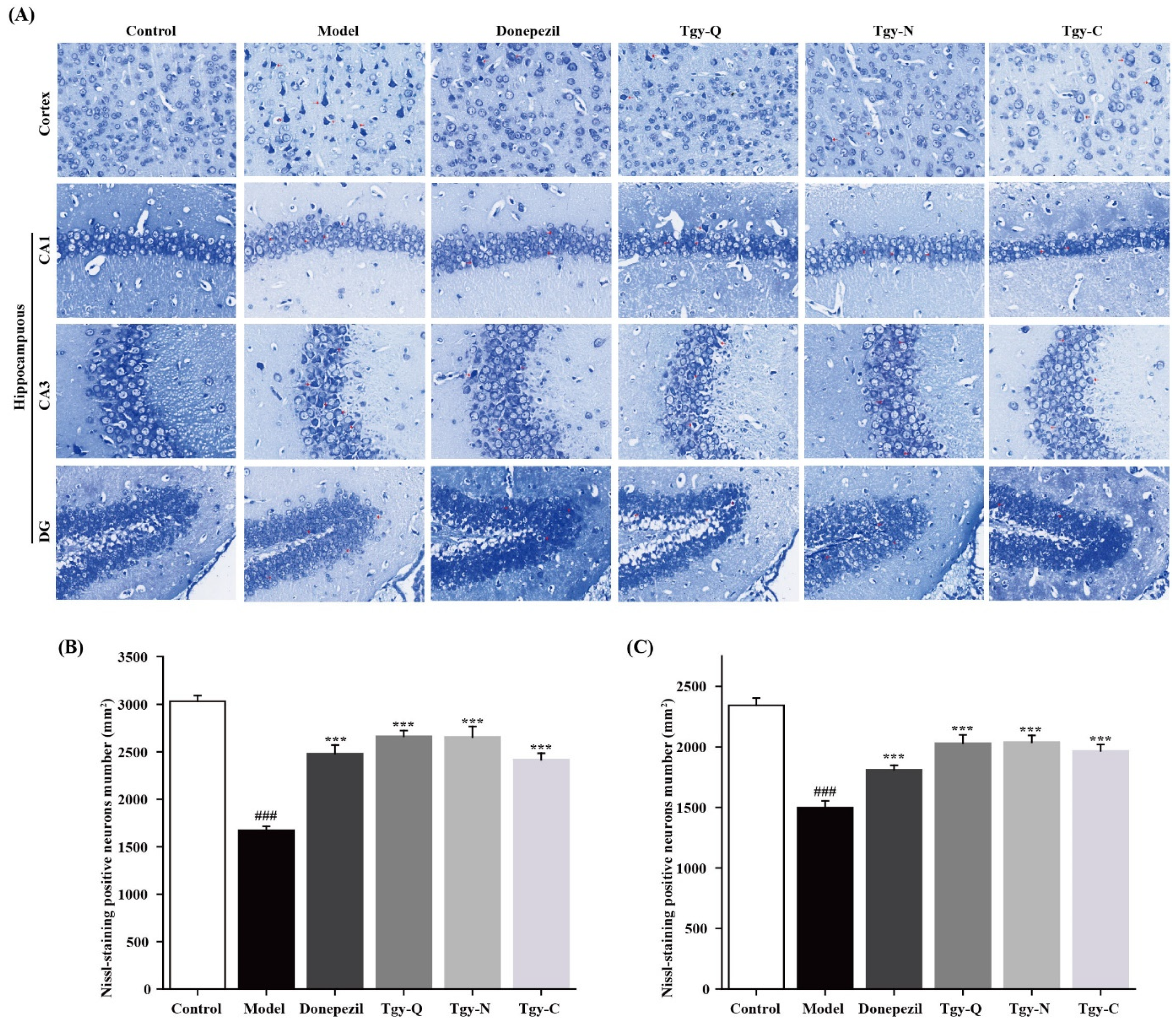
2.4. Effect of Tieguanyin Extracts on Cortical Neurons of APP/PS1 Mice
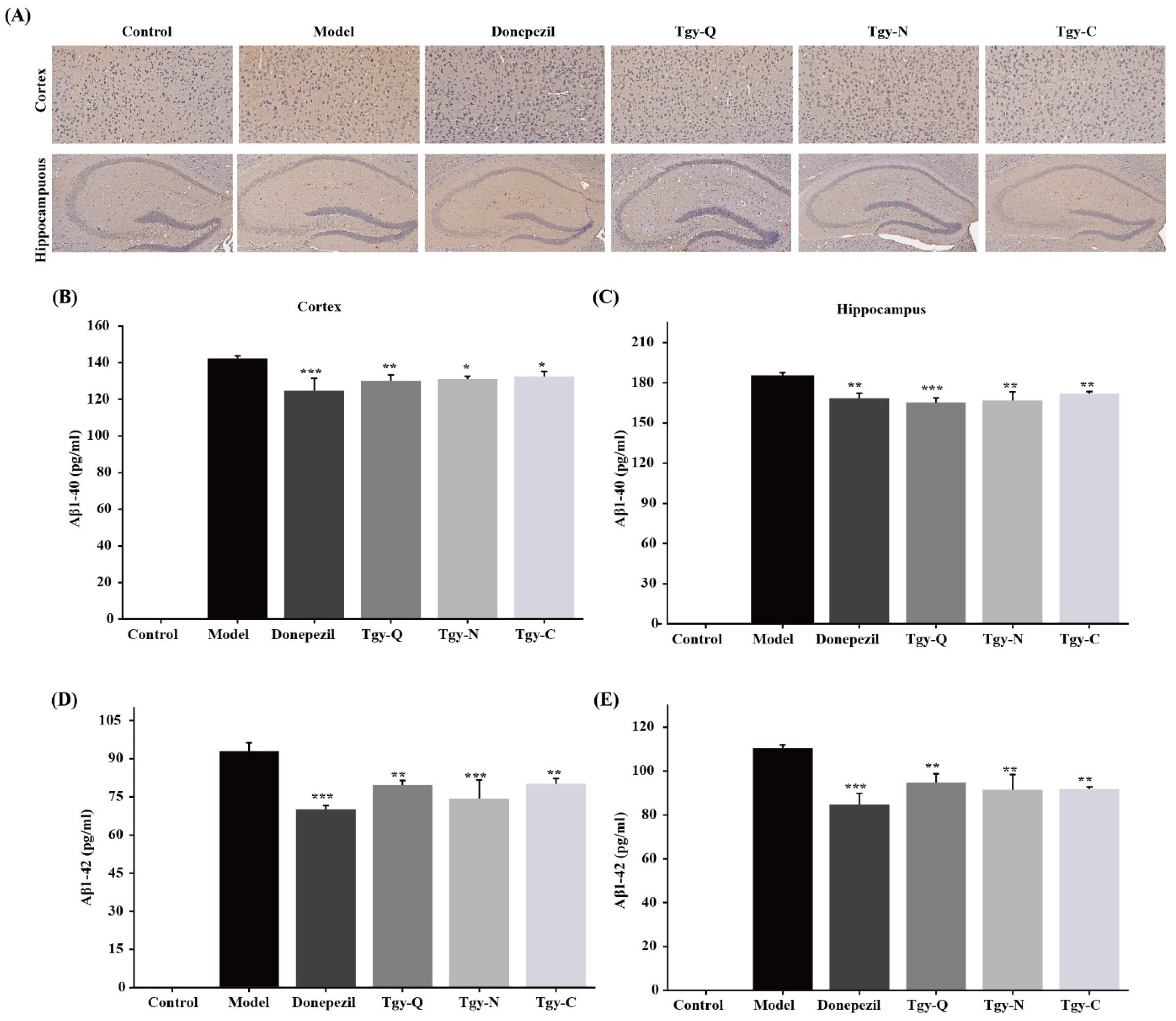
2.5. Tieguanyin Extracts Improves the Deposition of Aβ1-42 Amyloid Plaques in the Cortex and Hippocampus of APP/PS1 Mice
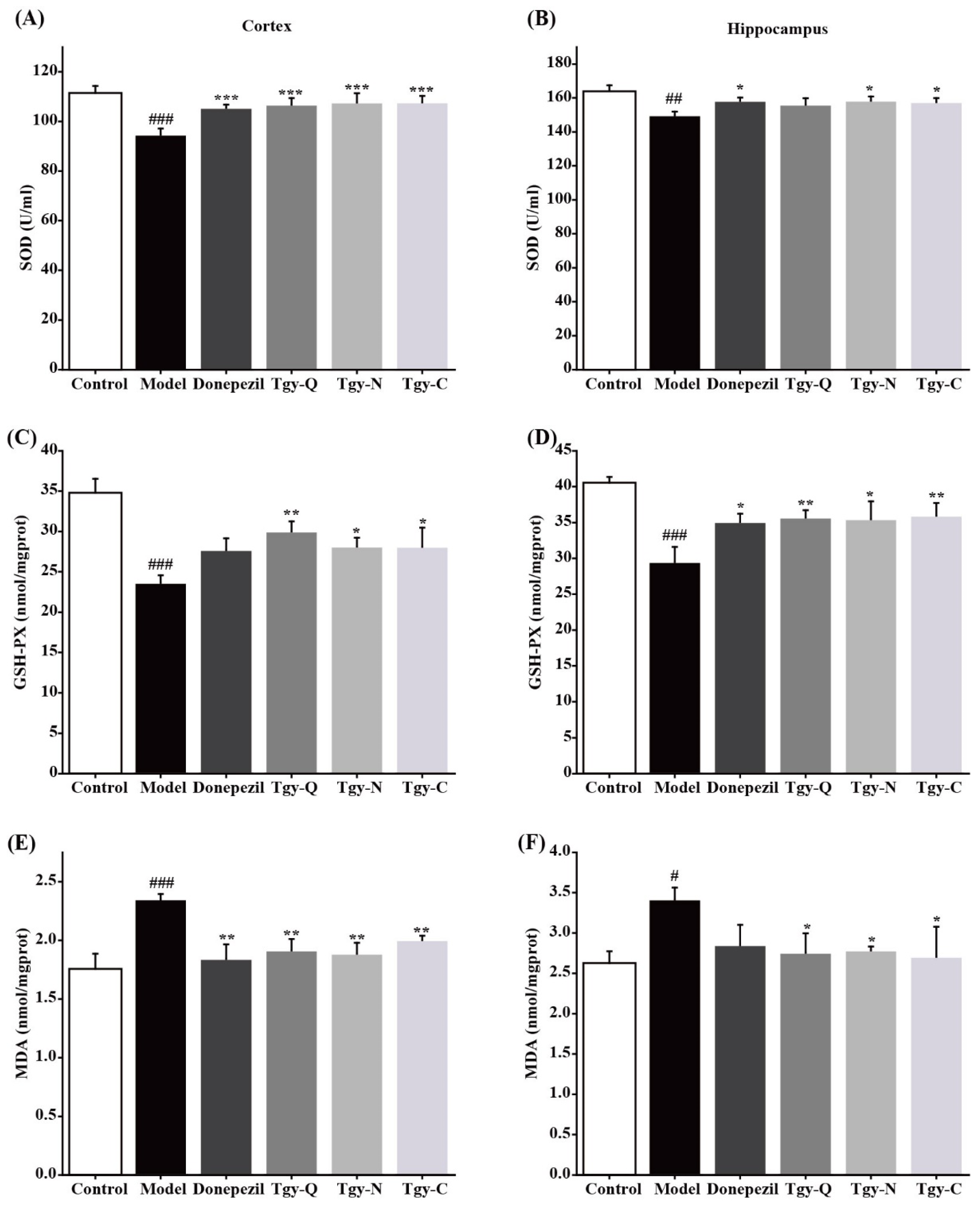
2.6. Tieguanyin Extracts Improves Brain Oxidative Stress in APP/PS1 Mice
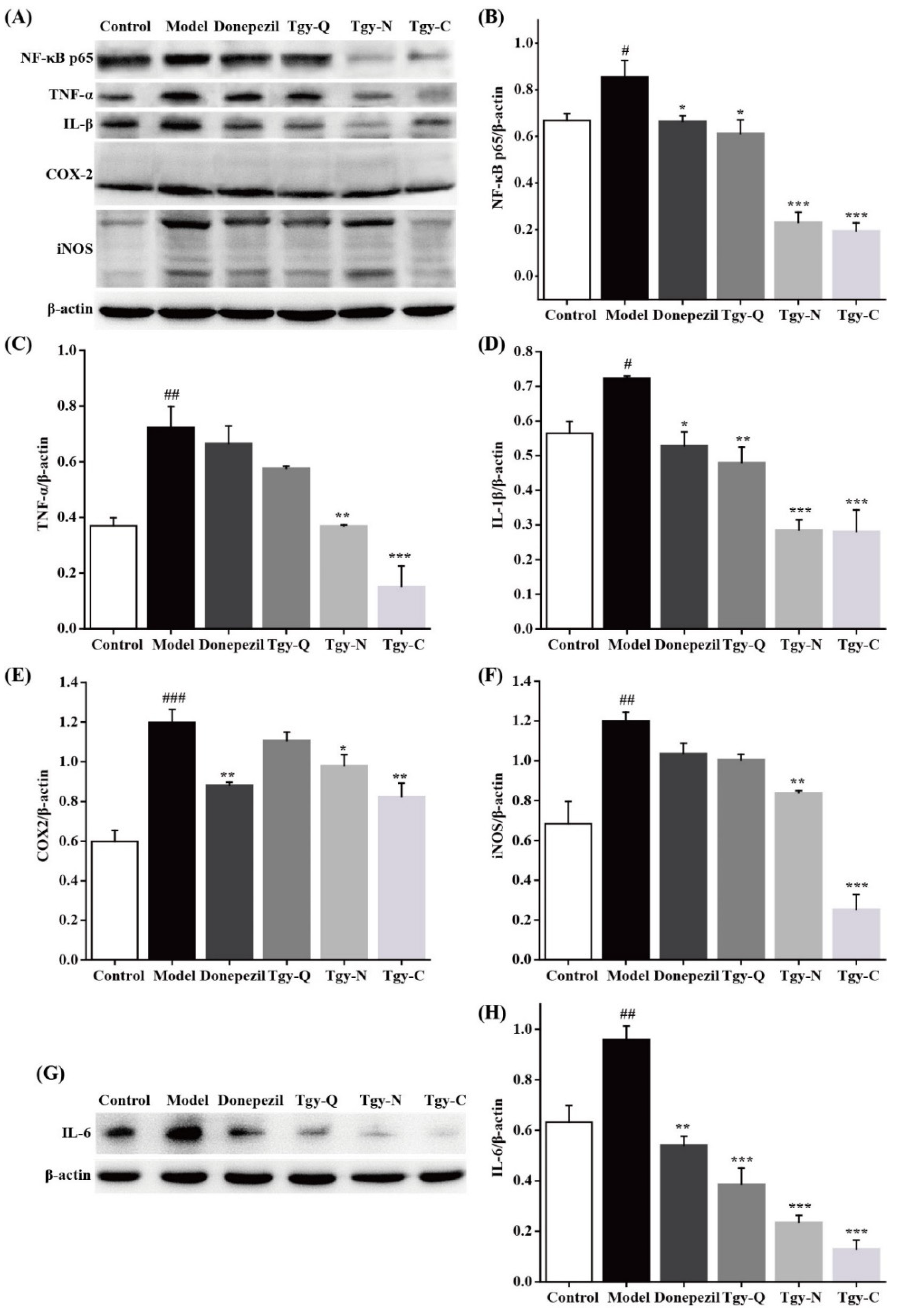
2.7. Tieguanyin Extracts Reduces Neuroinflammation in the Brain of APP/PS1 Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Preparation of Tieguanyin Extracts
4.3. Determination of Chemical Components of Tieguanyin Extracts
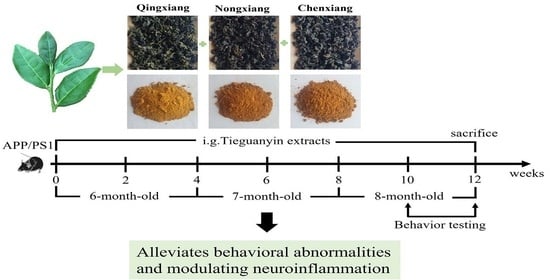
4.4. Animals
4.5. Morris Water Maze Test
4.6. Novel Object Recognition Test
4.7. Immunohistochemistry
4.8. Hematoxylin and Eosin (H&E) Staining
4.9. Quantification of Aβ1-40 and Aβ1-42 in Brain Tissues by Elisa
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spangenberg, E.E.; Green, K.N. Inflammation in Alzheimer’s disease: Lessons learned from microglia-depletion models. Brain Behav. Immun. 2017, 61, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C. World Alzheimer Report 2019: Attitudes to dementia, a global survey: Public health: Engaging people in ADRD research. Alzheimer Dement. 2020, 16, e038255. [Google Scholar] [CrossRef]
- Birks, J.S.; Harvey, R.J.J. Donepezil for dementia due to Alzheimer’s disease. Cochrane Database Syst. Rev. 2018, 6, CD001190. [Google Scholar] [CrossRef]
- Mao, X.-Y.; Yin, X.-X.; Guan, Q.-W.; Xia, Q.-X.; Yang, N.; Zhou, H.-H.; Liu, Z.-Q.; Jin, W.-L. Therapeutics. Dietary nutrition for neurological disease therapy. Curr. Status Future Dir. 2021, 226, 107861. [Google Scholar]
- Huang, L.; Su, T.; Li, X. Natural products as sources of new lead compounds for the treatment of Alzheimer’s disease. Curr. Top. Med. Chem. 2013, 13, 1864–1878. [Google Scholar] [CrossRef]
- Andrade, S.; Ramalho, M.J.; Loureiro, J.A.; Pereira, M.D.C. Natural compounds for Alzheimer’s disease therapy: A systematic review of preclinical and clinical studies. Int. J. Mol. Sci. 2019, 20, 2313. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-Q.; Wang, Z.-S.; Ma, Y.-X.; Zhang, W.; Lu, J.-L.; Liang, Y.-R.; Zheng, X.-Q. Neuroprotective effects and mechanisms of tea bioactive components in neurodegenerative diseases. Molecules 2018, 23, 512. [Google Scholar] [CrossRef] [Green Version]
- Mandel, S.A.; Amit, T.; Weinreb, O.; Reznichenko, L.; Youdim, M.B.H. Simultaneous manipulation of multiple brain targets by green tea catechins: A potential neuroprotective strategy for Alzheimer and Parkinson diseases. CNS Neurosci. Ther. 2008, 14, 352–365. [Google Scholar] [CrossRef]
- Schimidt, H.L.; Garcia, A.; Martins, A.; Mello-Carpes, P.; Carpes, F. Green tea supplementation produces better neuroprotective effects than red and black tea in Alzheimer-like rat model. Food Res. Int. 2017, 100, 442–448. [Google Scholar] [CrossRef]
- Xu, Y.-Q.; Liu, P.; Shi, J.; Gao, Y.; Wang, Q.-S.; Yin, J.-F. Quality development and main chemical components of Tieguanyin oolong teas processed from different parts of fresh shoots. Food Chem. 2018, 249, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Zhong-Zhen, Z.; Cao, Z.-J.; Chen, H.; Zhao, Z.-Z.; Zhu, L.; Yi, T. Oolong tea: A critical review of processing methods, chemical composition, health effects, and risk. Crit. Rev. Food Sci. Nutr. 2018, 58, 2957–2980. [Google Scholar]
- Garcia-Alloza, M.; Robbins, E.M.; Zhang-Nunes, S.X.; Purcell, S.M.; Betensky, R.A.; Raju, S.; Prada, C.; Greenberg, S.M.; Bacskai, B.J.; Frosch, M.P. Characterization of amyloid deposition in the APPswe/PS1dE9 mouse model of Alzheimer disease. Neurobiol. Dis. 2006, 24, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Yeo, I.J.; Hwang, C.J.; Choi, D.-Y.; Im, H.-S.; Kim, J.Y.; Choi, W.R.; Jung, M.H.; Han, S.-B.; Hong, J.T. Estrogen deficiency exacerbates Aβ-induced memory impairment through enhancement of neuroinflammation, amyloidogenesis and NF-ĸB activation in ovariectomized mice. Brain Behav. Immun. 2018, 73, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Q.; Ip, S.-P.; Yuan, Q.-J.; Zheng, G.-Q.; Tsim, K.K.; Dong, T.T.; Lin, G.; Han, Y.; Liu, Y.; Xian, Y.-F.; et al. Isorhynchophylline ameliorates cognitive impairment via modulating amyloid pathology, tau hyperphosphorylation and neuroinflammation: Studies in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2019, 82, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Benilova, I.; Karran, E.; De Strooper, B. The toxic Aβ oligomer and Alzheimer’s disease: An emperor in need of clothes. Nat. Neurosci. 2012, 15, 349–357. [Google Scholar] [CrossRef]
- Wiessner, C.; Wiederhold, K.-H.; Tissot, A.C.; Frey, P.; Danner, S.; Jacobson, L.; Jennings, G.T.; Lüönd, R.; Ortmann, R.; Reichwald, J.; et al. The second-generation active Aβ immunotherapy CAD106 reduces amyloid accumulation in APP transgenic mice while minimizing potential Side effects. J. Neurosci. 2011, 31, 9323–9331. [Google Scholar] [CrossRef]
- Wang, T.; Xie, X.-X.; Ji, M.; Wang, S.-W.; Zha, J.; Zhou, W.-W.; Yu, X.-L.; Wei, C.; Ma, S.; Xi, Z.-Y.; et al. Naturally occurring autoantibodies against Aβ oligomers exhibited more beneficial effects in the treatment of mouse model of Alzheimer’s disease than intravenous immunoglobulin. Neuropharmacology 2016, 105, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-W.; Lu, S.; Su, Y.-J.; Xue, D.; Yu, X.-L.; Wang, S.-W.; Zhang, H.; Xu, P.-X.; Xie, X.-X.; Liu, R.-T. Decreasing oxidative stress and neuroinflammation with a multifunctional peptide rescues memory deficits in mice with Alzheimer disease. Free Radic. Biol. Med. 2014, 74, 50–63. [Google Scholar] [CrossRef]
- Rezai-Zadeh, K.; Arendash, G.W.; Hou, H.; Fernandez, F.; Jensen, M.; Runfeldt, M.; Shytle, R.D.; Tan, J. Green tea epigallocatechin-3-gallate (EGCG) reduces β-amyloid mediated cognitive impairment and modulates tau pathology in Alzheimer transgenic mice. Brain Res. 2008, 1214, 177–187. [Google Scholar] [CrossRef]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Kan, Z.; Wang, Y.; Chen, Q.; Tang, X.; Thompson, H.J.; Huang, J.; Zhang, J.; Gao, F.; Shen, Y.; Wan, X. Green tea suppresses amyloid β levels and alleviates cognitive impairment by Inhibiting APP cleavage and preventing neurotoxicity in 5XFAD mice. Mol. Nutr. Food Res. 2021, 65, 2100626. [Google Scholar] [CrossRef]
- Lim, H.J.; Shim, S.B.; Jee, S.W.; Lee, S.H.; Lim, C.J.; Hong, J.T.; Sheen, Y.Y.; Hwang, D.Y. Green tea catechin leads to global improvement among Alzheimer’s disease-related phenotypes in NSE/hAPP-C105 Tg mice. J. Nutr. Biochem. 2013, 24, 1302–1313. [Google Scholar] [CrossRef]
- Ide, K.; Matsuoka, N.; Yamada, H.; Furushima, D.; Kawakami, K. Effects of tea catechins on Alzheimer’s disease: Recent updates and perspectives. Molecules 2018, 23, 2357. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.-M.; Han, Y.-W.; Han, X.-H.; Zhang, K.; Chang, Y.-N.; Hu, Z.-M.; Qi, H.-X.; Ting, C.; Zhen, Z.; Hong, W. Upstream regulators and downstream effectors of NF-κB in Alzheimer’s disease. J. Neurol. Sci. 2016, 366, 127–134. [Google Scholar] [CrossRef]
- Snow, W.M.; Albensi, B.C. Neuronal gene targets of NF-κB and their dysregulation in Alzheimer’s disease. Front. Mol. Neurosci. 2016, 9, 118. [Google Scholar] [CrossRef]
- Zhong, X.; Liu, M.; Yao, W.; Du, K.; He, M.; Jin, X.; Jiao, L.; Ma, G.; Wei, B.; Wei, M. Epigallocatechin-3-gallate attenuates microglial inflammation and neurotoxicity by suppressing the activation of canonical and noncanonical inflammasome via TLR4/NF-κB pathway. Mol. Nutr. Food Res. 2019, 63, 1801230. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Tai, Y.; Wei, C.; Yang, H.; Zhang, L.; Chen, Q.; Deng, W.; Wei, S.; Zhang, J.; Fang, C.; Ho, C.; et al. Transcriptomic and phytochemical analysis of the biosynthesis of characteristic constituents in tea (Camellia sinensis) compared with oil tea (Camellia oleifera). BMC Plant Biol. 2015, 15, 190. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, S.; Alam, S.; Saleem, M.; Ahmad, W.; Bibi, R.; Hamid, F.S.; Shah, H.U. Withering timings affect the total free amino acids and mineral contents of tea leaves during black tea manufacturing. Arab. J. Chem. 2019, 12, 2411–2417. [Google Scholar] [CrossRef] [Green Version]
- Laurentin, A.; Edwards, C.A. A microtiter modification of the anthrone-sulfuric acid colorimetric assay for glucose-based carbohydrates. Anal. Biochem. 2003, 315, 143–145. [Google Scholar] [CrossRef]
- Wei, X.; Mao, F.; Cai, X.; Wang, Y. Composition and bioactivity of polysaccharides from tea seeds obtained by water extraction. Int. J. Biol. Macromol. 2011, 49, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Chao, F.-L.; Zhang, L.; Zhang, Y.; Zhou, C.-N.; Jiang, L.; Xiao, Q.; Luo, Y.-M.; Lv, F.-L.; He, Q.; Tang, Y. Running exercise protects against myelin breakdown in the absence of neurogenesis in the hippocampus of AD mice. Brain Res. 2018, 1684, 50–59. [Google Scholar] [CrossRef]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object recognition test in mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and Eosin Staining of Tissue and Cell Sections. Cold Spring Harb. Protoc. 2008, 2008, pdb-prot4986. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, N.; Rankin, G.O.; Li, B.; Rojanasakul, Y.; Tu, Y.; Chen, Y.C. Anti-proliferative effect and cell cycle arrest induced by saponins extracted from tea (Camellia sinensis) flower in human ovarian cancer cells. J. Funct. Foods 2017, 37, 310–321. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Constituents | Tgy-Q | Tgy-N | Tgy-C |
|---|---|---|---|
| Tea polyphenol (mg/g) | 437.25 ± 2.13 a | 396.48 ± 3.81 b | 368.26 ± 2.92 c |
| Amino acids (mg/g) | 64.08 ± 0.60 a | 62.55 ± 0.74 b | 47.92 ± 0.28 c |
| Soluble proteins (mg/g) | 88.61 ± 3.01 b | 92.11 ± 2.79 b | 104.73 ± 5.25 a |
| Soluble sugars (mg/g) | 362.01 ± 8.19 a | 336.52 ± 4.35 b | 278.18± 4.17 c |
| Tea polysaccharides (mg/g) | 48.64 ± 2.11 b | 37.29 ± 1.00 c | 57.35 ± 1.52 a |
| Flavonoids (mg/g) | 24.18 ± 0.54 b | 26.56 ± 1.78 b | 32.71 ± 0.97 a |
| Caffeine (mg/g) | 71.36 ± 0.89 b | 77.12 ± 0.73 c | 77.49 ± 0.92 a |
| Gallic acid (mg/g) | 2.14 ± 0.21 c | 3.76 ± 0.17 b | 6.67 ± 0.44 a |
| Gallocatechin (mg/g) | 38.37 ± 1.08 a | 30.35 ± 1.62 c | 30.39 ± 1.76 b |
| Epigallocatechin (mg/g) | 49.06 ± 1.86 a | 45.80 ± 1.14 b | 38.47 ± 0.42 c |
| Catechin (mg/g) | 7.74 ± 1.61 a | 8.12 ± 0.76 a | 8.70 ± 1.72 a |
| Epicatechin (mg/g) | 24.59 ± 1.42 a | 16.32 ± 0.77 b | 5.41 ± 0.44 c |
| Epigallocatechin gallate (mg/g) | 105.21 ± 1.64 a | 86.99 ± 1.77 b | 77.42 ± 0.91 c |
| Gallocatechin gallate (mg/g) | 31.24 ± 1.07 b | 32.54 ± 1.29 b | 36.12 ± 1.31 a |
| Epicatechin gallate (mg/g) | 32.05 ± 0.84 a | 30.74 ± 1.34 a | 24.83 ± 1.24 b |
| Catechin gallate (mg/g) | 6.28 ± 0.64 a | 6.48 ± 1.47 a | 6.415 ± 1.52 a |
| Total catechins (mg/g) | 294.56 ± 10.91 a | 257.35 ± 10.20 b | 227.79 ± 9.36 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Zhou, H.; Ye, Y.; Yang, J.; Liu, Z.; He, P.; Li, B.; Wu, Y.; Wang, Y.; Tu, Y. Tieguanyin Oolong Tea Extracts Alleviate Behavioral Abnormalities by Modulating Neuroinflammation in APP/PS1 Mouse Model of Alzheimer’s Disease. Foods 2022, 11, 81. https://doi.org/10.3390/foods11010081
Kang H, Zhou H, Ye Y, Yang J, Liu Z, He P, Li B, Wu Y, Wang Y, Tu Y. Tieguanyin Oolong Tea Extracts Alleviate Behavioral Abnormalities by Modulating Neuroinflammation in APP/PS1 Mouse Model of Alzheimer’s Disease. Foods. 2022; 11(1):81. https://doi.org/10.3390/foods11010081
Chicago/Turabian StyleKang, Hyunuk, Hui Zhou, Yushan Ye, Jiangfan Yang, Zhonghua Liu, Puming He, Bo Li, Yuanyuan Wu, Yaomin Wang, and Youying Tu. 2022. "Tieguanyin Oolong Tea Extracts Alleviate Behavioral Abnormalities by Modulating Neuroinflammation in APP/PS1 Mouse Model of Alzheimer’s Disease" Foods 11, no. 1: 81. https://doi.org/10.3390/foods11010081
APA StyleKang, H., Zhou, H., Ye, Y., Yang, J., Liu, Z., He, P., Li, B., Wu, Y., Wang, Y., & Tu, Y. (2022). Tieguanyin Oolong Tea Extracts Alleviate Behavioral Abnormalities by Modulating Neuroinflammation in APP/PS1 Mouse Model of Alzheimer’s Disease. Foods, 11(1), 81. https://doi.org/10.3390/foods11010081