
Inhibitory Effect and Potential Mechanism of Lactobacillus plantarum YE4 against Dipeptidyl Peptidase-4


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Caco-2 Cells Culture
2.3. Determination of DPP-4 Activity in Caco-2 Cells
2.4. Preparation of Cell-Free Excretory Supernatant (CFS) and Cell-Free Intracellular Extract (CFE)
2.5. Determination of the Inhibitory Effects of CFS and CFE on DPP-4 Activity
2.6. qRT-PCR Analysis
2.7. RNA-Seq Analysis
2.8. Analysis of Active Compounds of CFE from LAB Strain Inhibiting DPP-4
2.9. Statistical Analysis
3. Results
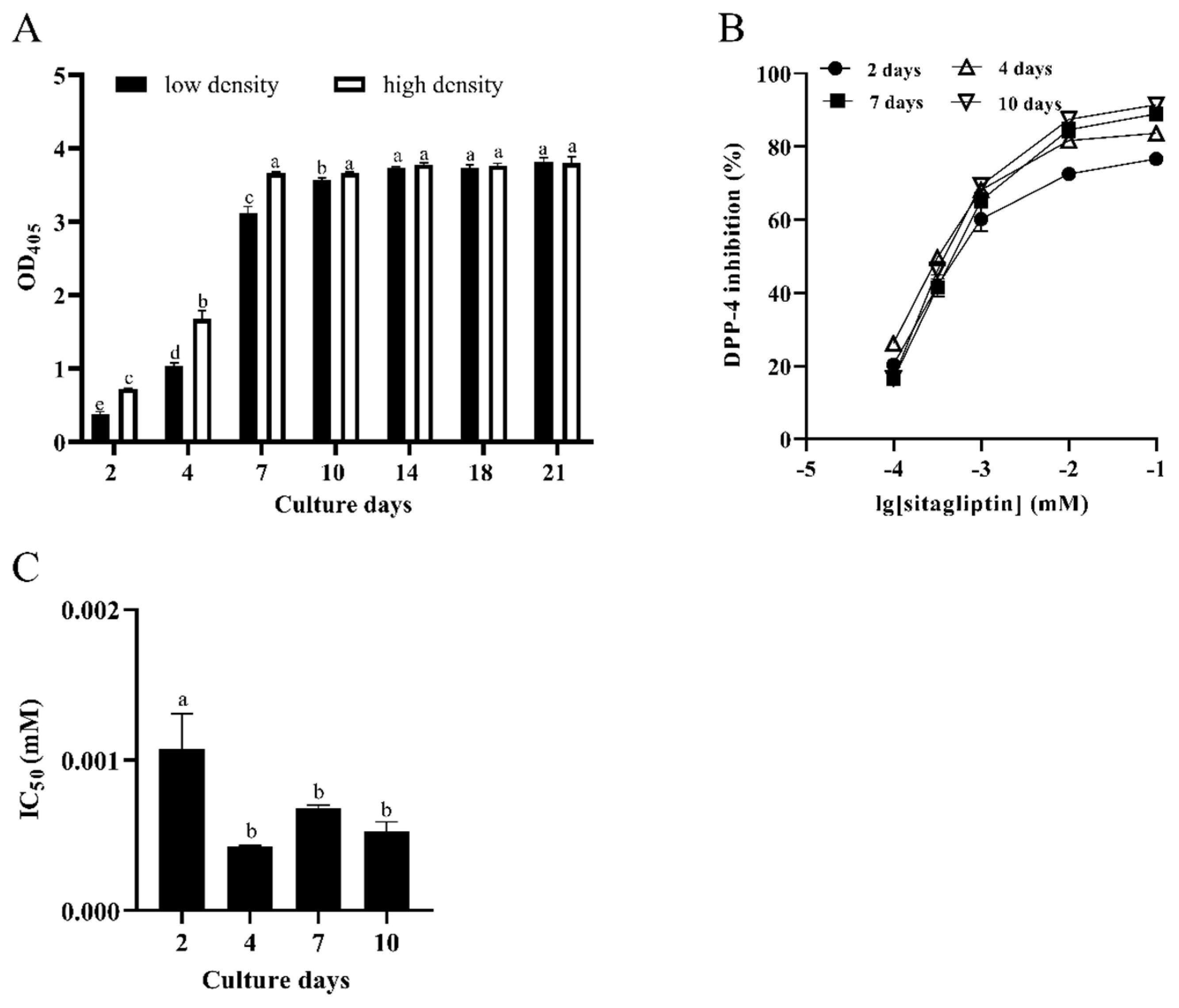
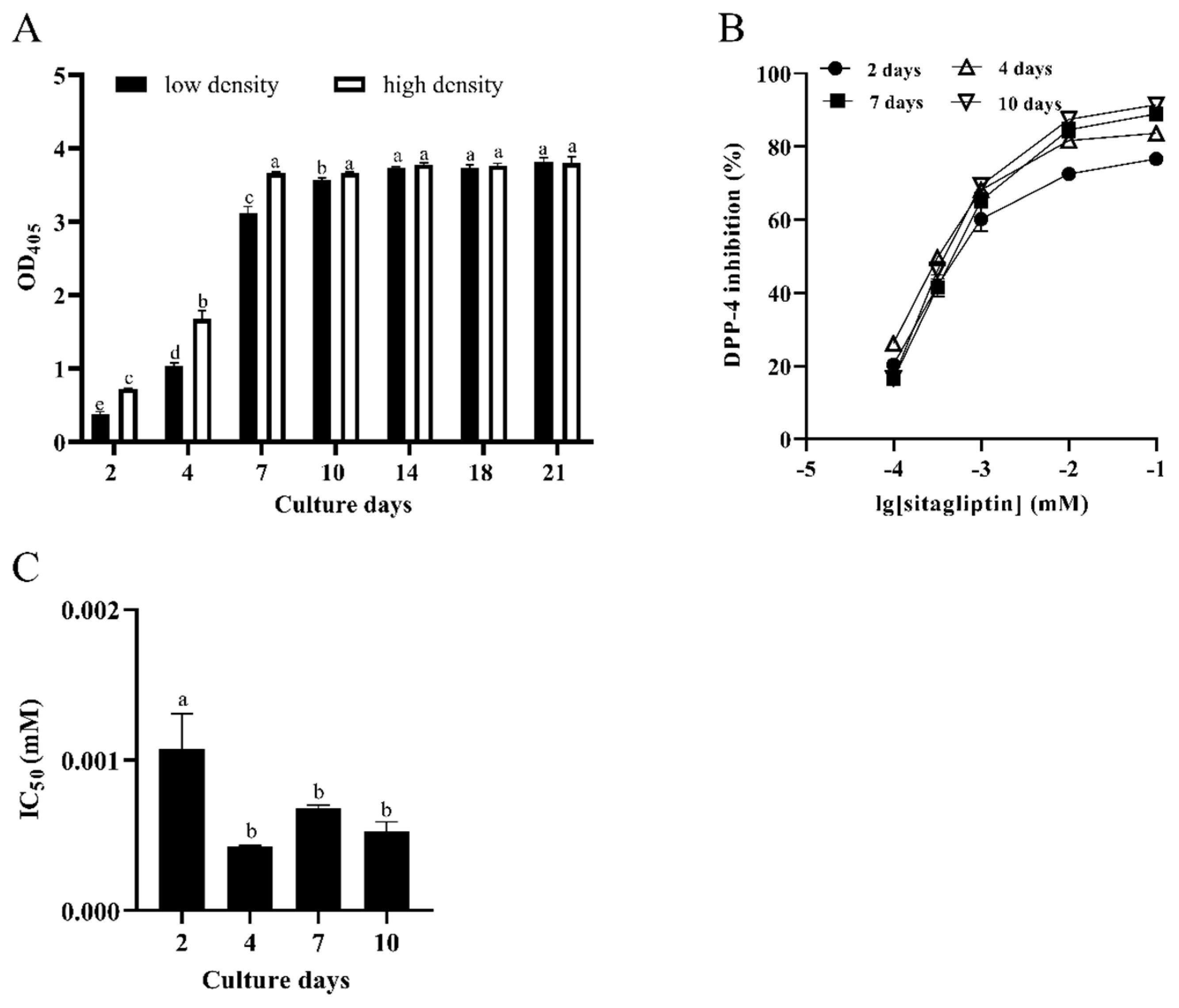
3.1. Characterization of DPP-4 Activity in Caco-2 Cells
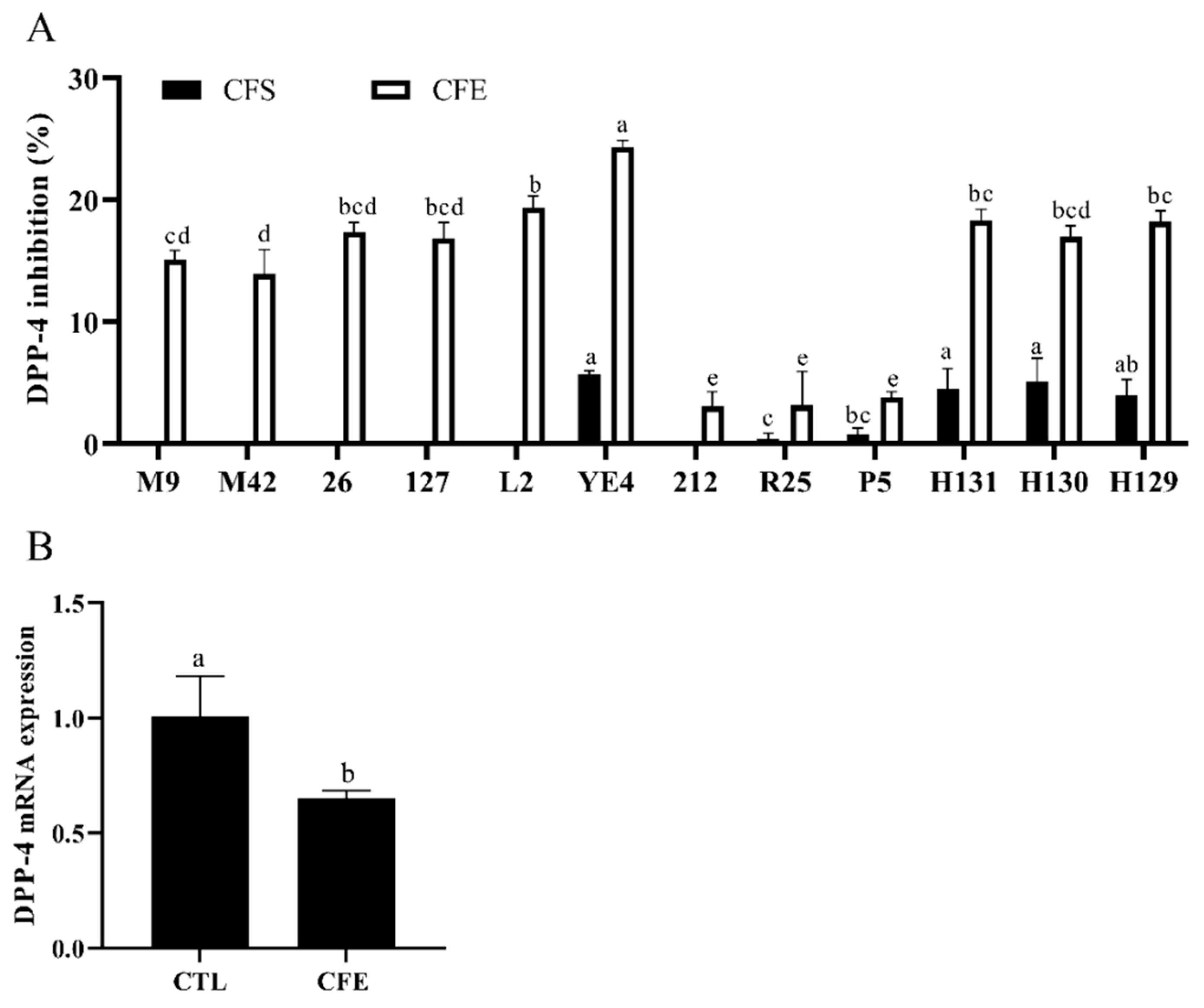
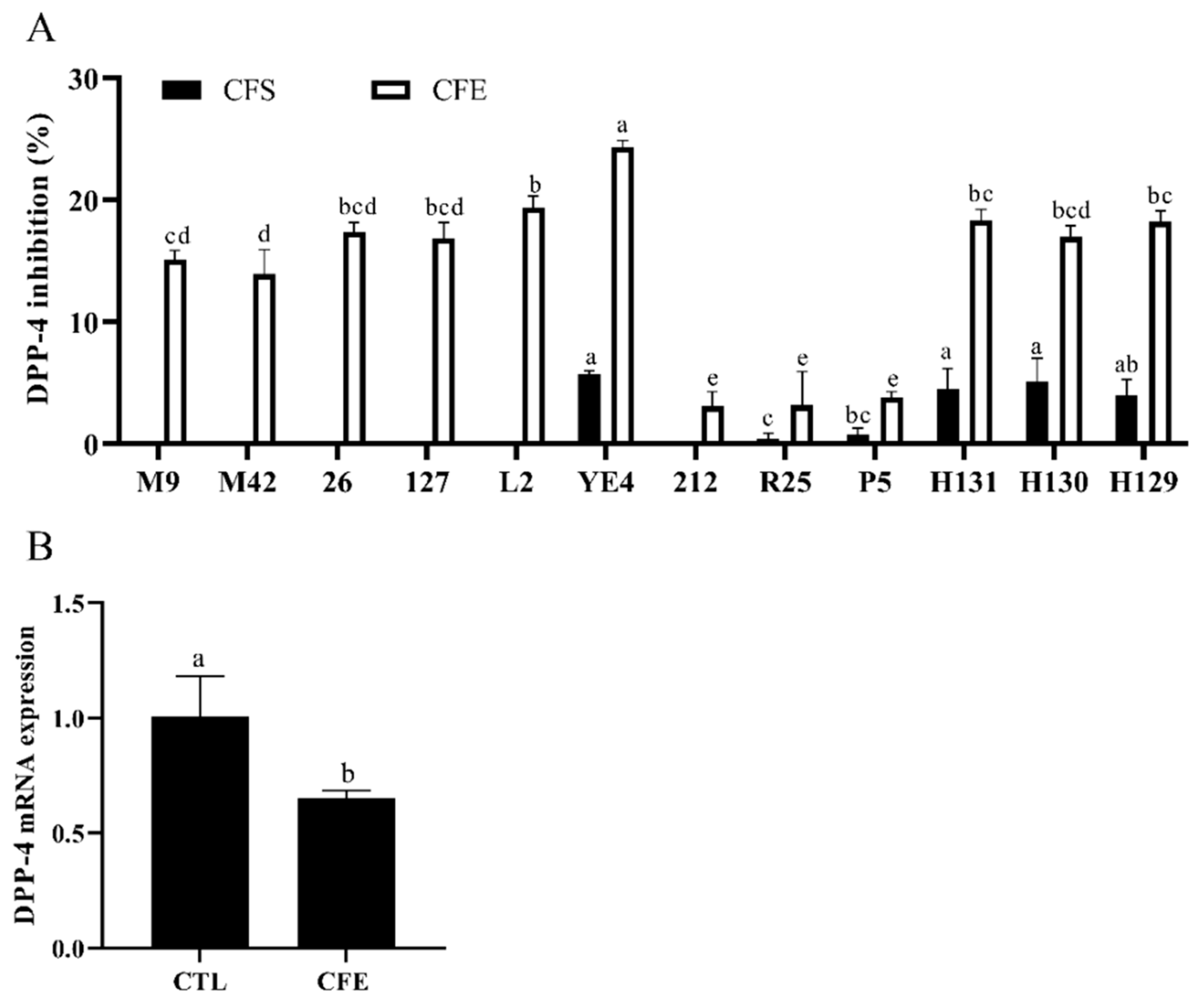
3.2. Effects of LAB Strains on DPP-4 Activity
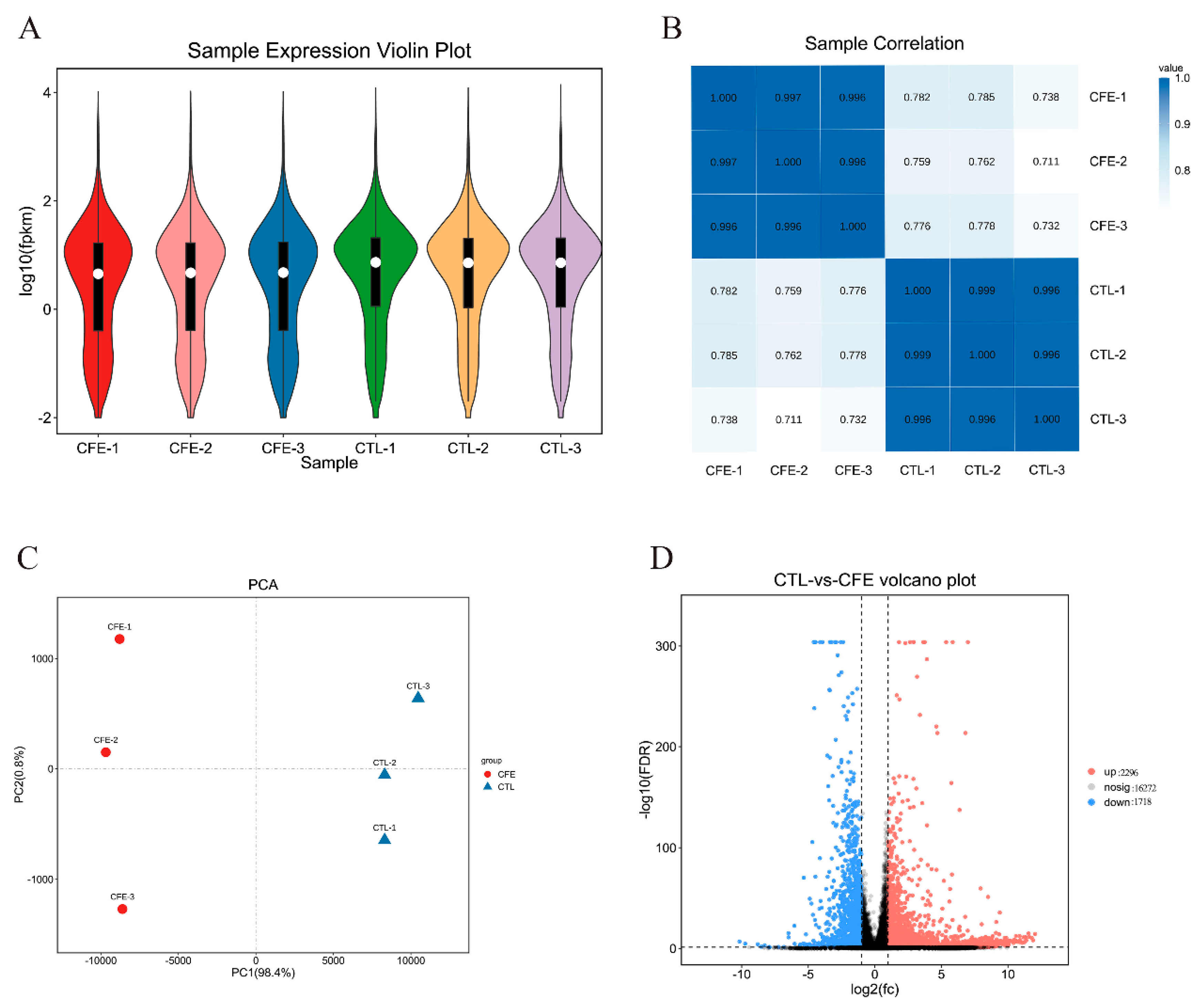
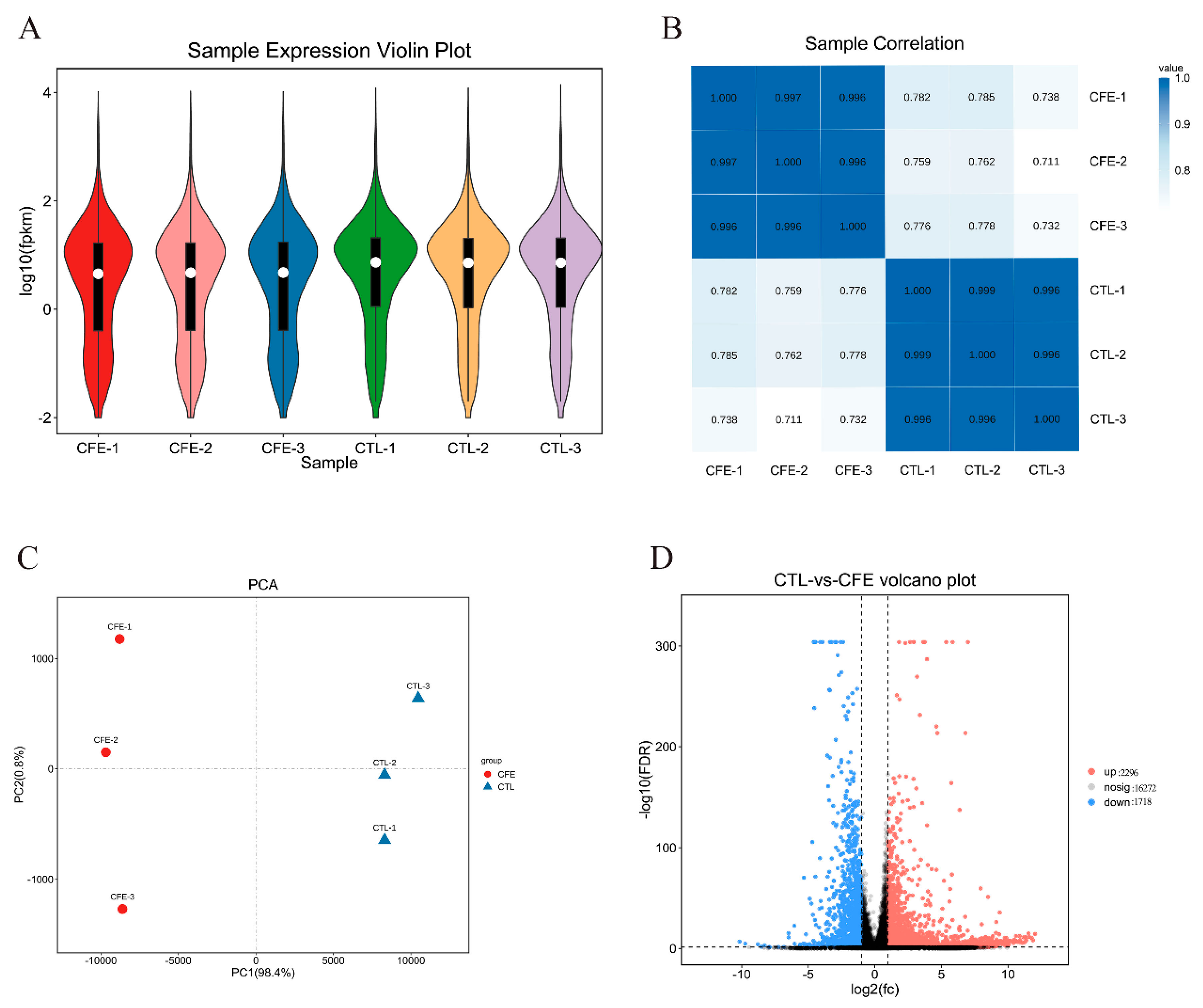
3.3. Overview of the Transcriptional Changes
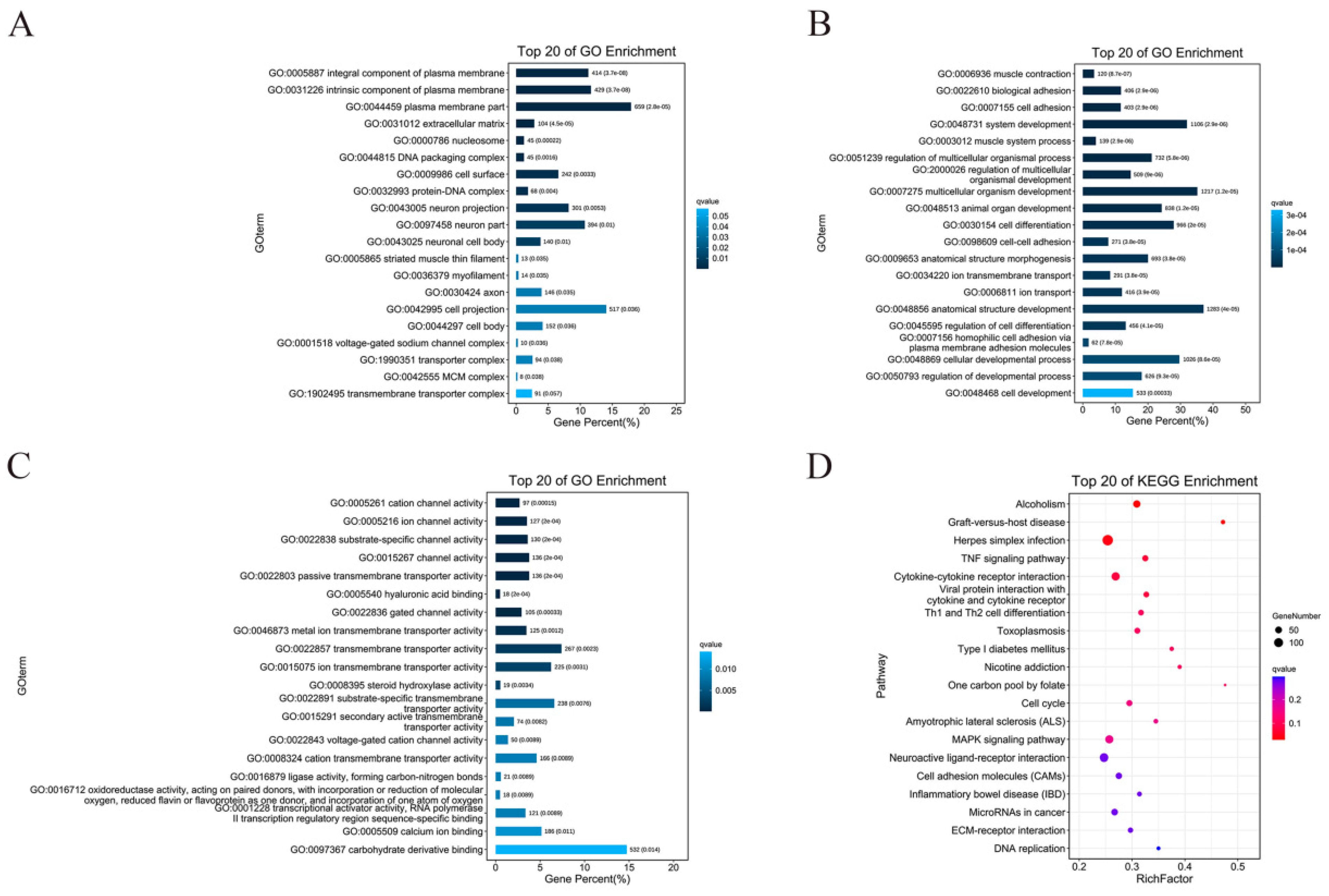
3.4. GO Enrichment and KEGG Enrichment Analysis of DEGs
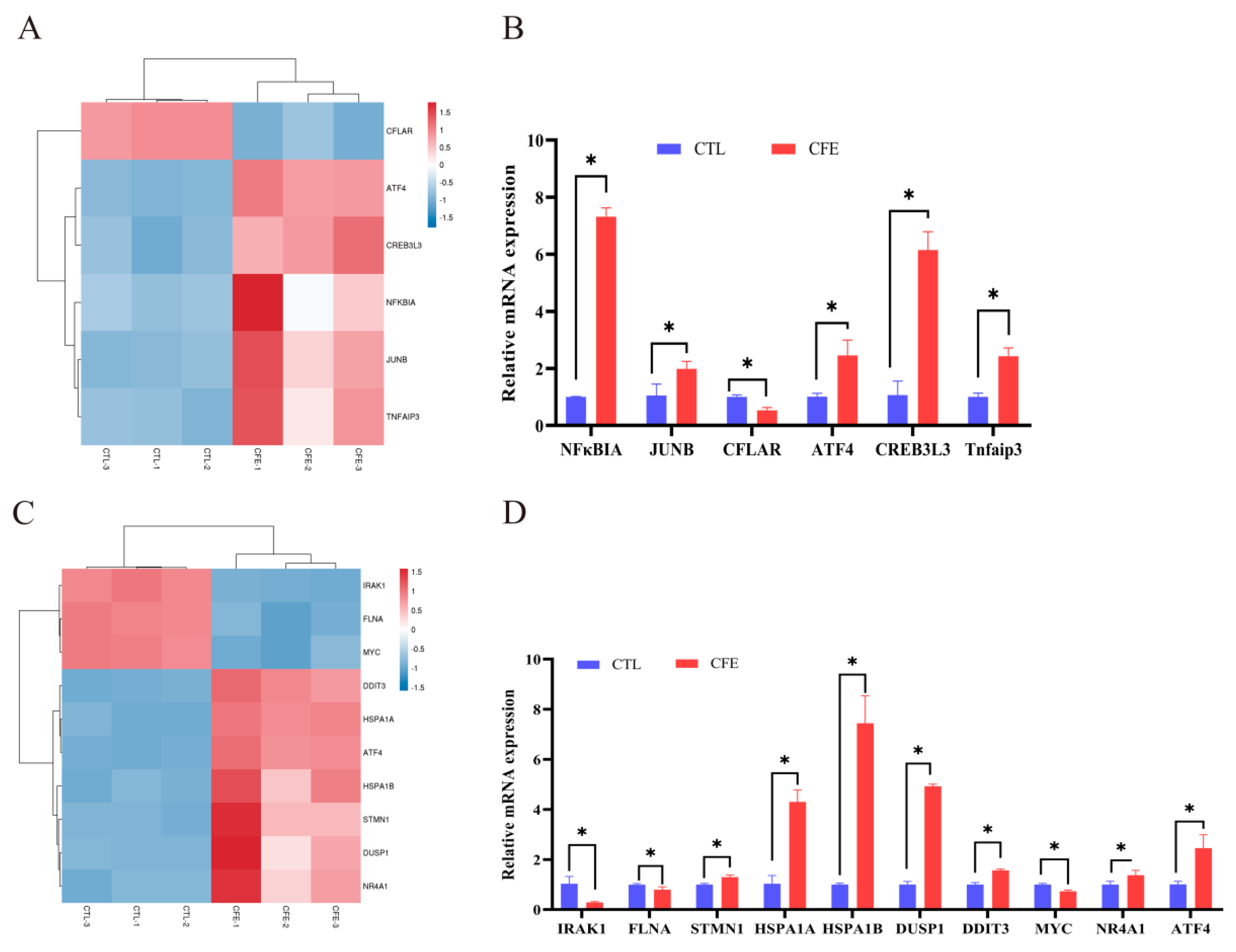
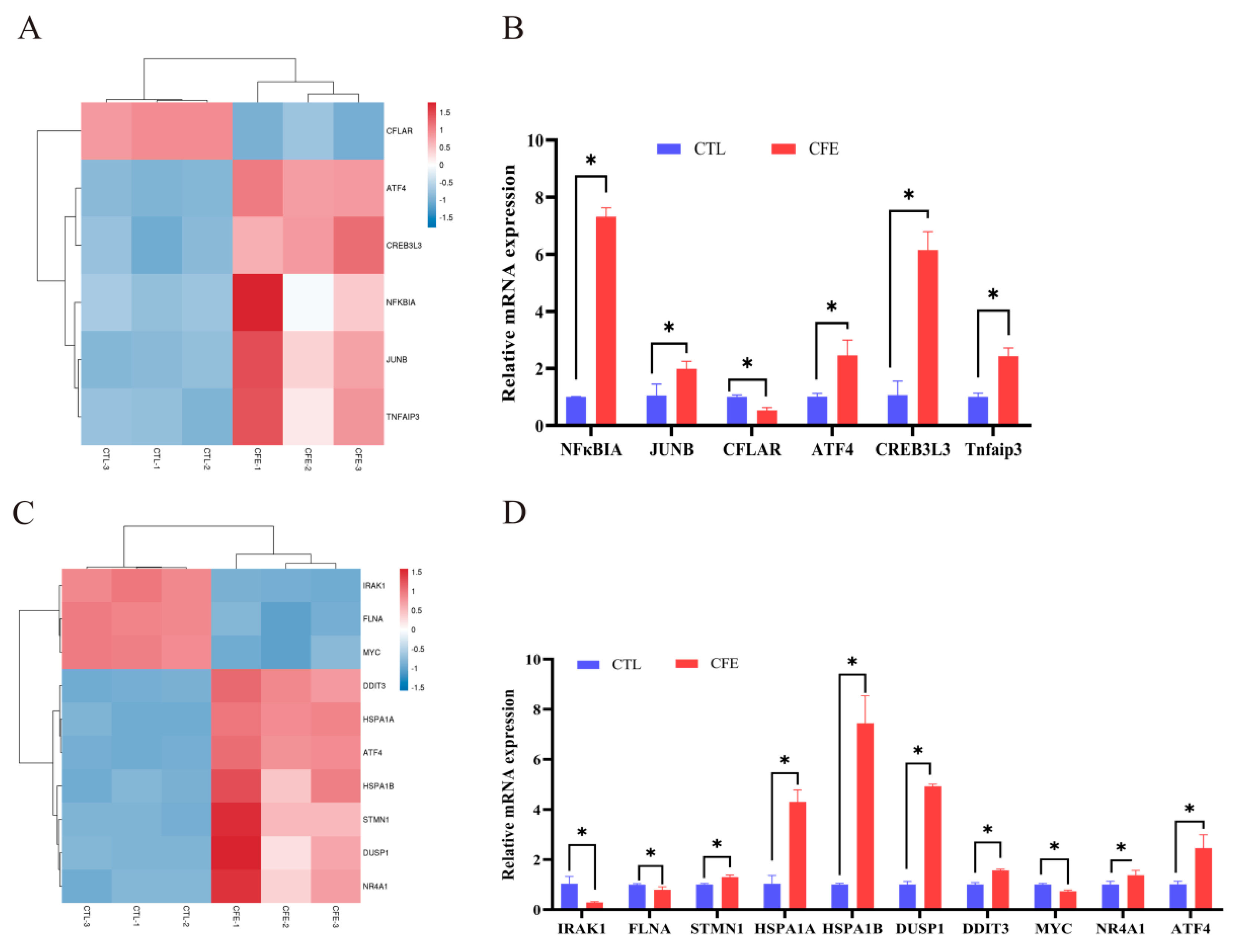
3.5. Effect of YE4-CFE on the Expression of TNF Signaling Pathway-Related Genes
3.6. Effect of YE4-CFE on the Expression of MAPK Signaling Pathway-Related Genes
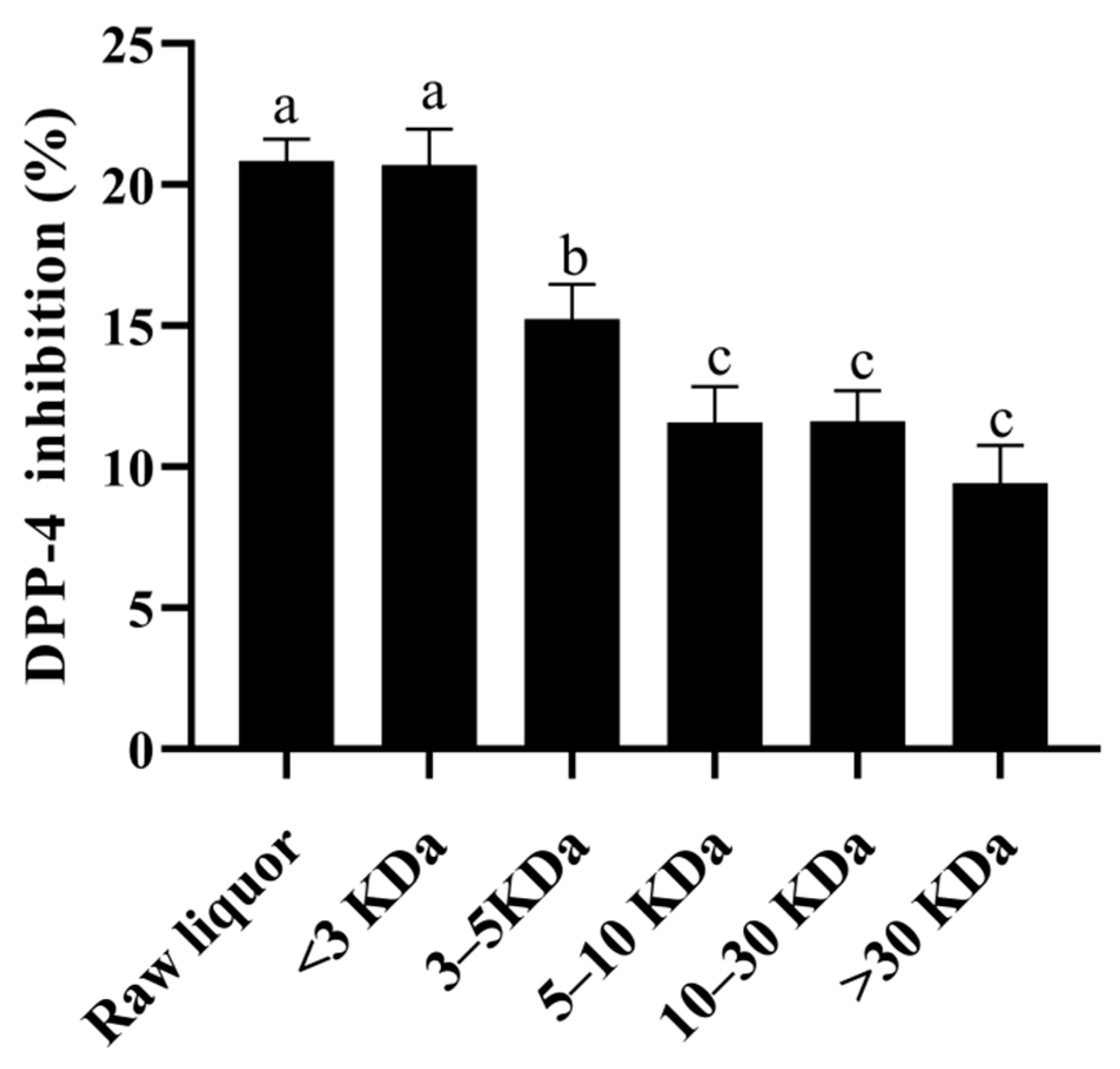
3.7. Analysis of the Effects of the Active Components of YE4-CFE against DPP-4 Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care. 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Einarson, T.R.; Annabel, A.; Craig, L.; Panton, U.H. Prevalence of cardiovascular disease in type 2 diabetes: A systematic literature review of scientific evidence from across the world in 2007–2017. Cardiovasc. Diabetol. 2018, 17, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Diabetes Federation. Advocacy Guide to the IDF Diabetes Atlas 2019. 2019. Available online: https://www.diabetesatlas.org/en/resources/ (accessed on 26 December 2020).
- Patel, B.D.; Ghate, M.D. Recent approaches to medicinal chemistry and therapeutic potential of dipeptidyl peptidase-4 (DPP-4) inhibitors. Eur. J. Med. Chem. 2014, 74, 574–605. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation-Current knowledge and future research considerations. Trends Food Sci. Technol. 2016, 54, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Luo, J.; Zuo, F.; Zhang, Y.; Ma, H.; Chen, S. Screening for potential novel probiotic Lactobacillus strains based on high dipeptidyl peptidase IV and α-glucosidase inhibitory activity. J. Funct. Foods. 2016, 20, 486–495. [Google Scholar] [CrossRef]
- Green, B.; Gault, V.; O’Harte, F.; Flatt, P. Structurally modified analogues of glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) as future antidiabetic agents. Curr. Pharm. Design. 2004, 10, 3651–3662. [Google Scholar] [CrossRef] [PubMed]
- Lucienne, J.J. Dipeptidyl peptidase IV and its inhibitors: Therapeutics for type 2 diabetes and what else? J. Med. Chem. 2014, 57, 2197–2212. [Google Scholar]
- Deacon, C.F. Dipeptidyl peptidase-4 inhibitors in the treatment of type 2 diabetes: A comparative review. Diabetes Obes. Metab. 2015, 13, 7–18. [Google Scholar] [CrossRef]
- Kushner, P.; Gorrell, M.D. DPP-4 inhibitors in type 2 diabetes: Importance of selective enzyme inhibition and implications for clinical use. J. Fam. Pract. 2010, 59, 1. [Google Scholar]
- Feng, L.; Xie, Y.; Peng, C.; Liu, Y.; Wang, H. A Novel Antidiabetic Food Produced via Solid-State Fermentation of Tartary Buckwheat using L. plantarum TK9 and L. paracasei TK1501. Food Technol. Biotechnol. 2018, 56, 373–380. [Google Scholar] [CrossRef]
- Panwar, H.; Calderwood, D.; Grant, I.R.; Grover, S.; Green, B.D. Lactobacilli possess inhibitory activity against dipeptidyl peptidase-4 (DPP-4). Ann. Microbiol. 2016, 66, 505–509. [Google Scholar] [CrossRef]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef] [Green Version]
- Harsh, P.; Hogarehalli Mallappa, R.; Virender Kumar, B.; Sunita, G. Probiotics as potential biotherapeutics in the management of type 2 diabetes-prospects and perspectives. Diabetes 2013, 29, 103–112. [Google Scholar]
- Miyoshi, M.; Ogawa, A.; Higurashi, S.; Kadooka, Y. Anti-obesity effect of Lactobacillus gasseri SBT2055 accompanied by inhibition of pro-inflammatory gene expression in the visceral adipose tissue in diet-induced obese mice. Eur. J. Nutr. 2014, 53, 599. [Google Scholar] [CrossRef]
- Panwar, H.; Calderwood, D.; Grant, I.R.; Grover, S.; Green, B.D. Lactobacillus strains isolated from infant faeces possess potent inhibitory activity against intestinal alpha- and beta-glucosidases suggesting anti-diabetic potential. Eur. J. Nutr. 2014, 53, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Yong, Z.; Lifeng, W.; Jiachao, Z.; Yunxu, L.; Qiuwen, H.; He, L.; Xiao, G.; Jianlin, G.; Heping, Z. Probiotic Lactobacillus casei Zhang ameliorates high-fructose-induced impaired glucose tolerance in hyperinsulinemia rats. Eur. J. Nutr. 2014, 53, 221–232. [Google Scholar]
- Xie, Y.; Han, X.; Wang, H.; Lu, F. Anti-diabetic Potential of Soymilk Fermented by Lactobacillus Paracasei TK1501 Isolated from Naturally Fermented Congee. Fresenius Environ. Bull. 2018, 27, 4381–4388. [Google Scholar]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum. Nutrients 2018, 10, 1082. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Huang, W.; Zhang, J.; Wang, X.; Tao, T.; Yang, L.; Zheng, Y.; Liu, S.; Yang, J.; Zhu, L.; et al. TMPRSS4 Promotes Cell Proliferation and Inhibits Apoptosis in Pancreatic Ductal Adenocarcinoma by Activating ERK1/2 Signaling Pathway. Front. Oncol. 2021, 11, 628353. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Zhao, D.; Liu, Q.; Liu, Y.; Huang, X.; Yang, J.; Zhang, L.; Li, Y. Transcriptome analysis reveals new insight of duck Tembusu virus (DTMUV)-infected DF-1 cells. Res. Vet. Sci. 2021, 137, 150–158. [Google Scholar] [CrossRef]
- Bian, X.; Zhao, Y.; Xiao, S.; Yang, H.; Han, Y.; Zhang, L. Metabolome and transcriptome analysis reveals the molecular profiles underlying the ginseng response to rusty root symptoms. BMC Plant Biol. 2021, 21, 215. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, B.; Li, M.; Liu, S.; Pan, T.; Ma, C.; Qiu, D. Melatonin-mediate acid rain stress tolerance mechanism through alteration of transcriptional factors and secondary metabolites gene expression in tomato. Ecotoxicol. Environ. Safe 2020, 200, 110720. [Google Scholar] [CrossRef]
- Yoshioka, M.; Erickson, R.H.; Matsumoto, H.; Gum, E.; Kim, Y.S. Expression of dipeptidyl aminopeptidase IV during enterocytic differentiation of human colon cancer (Caco-2) cells. Int. J. Cancer 1991, 47, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Noemi, G.; Neus, M.; Mayte, B.; Gerard, P.; Santiago, G.; Montserrat, P.; Anna, A. Grape seed-derived procyanidins decrease dipeptidyl-peptidase 4 activity and expression. J. Agric. Food Chem. 2012, 60, 9055–9061. [Google Scholar]
- Emrich, S.J.; Barbazuk, W.B.; Li, L.; Schnable, P.S. Gene discovery and annotation using LCM-454 transcriptome sequencing. Genome Res. 2007, 17, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Li, X.; Zheng, J. Transcriptome profiling using Illumina- and SMRT-based RNA-seq of hot pepper for in-depth understanding of genes involved in CMV infection. Gene 2018, 666, 123–133. [Google Scholar] [CrossRef]
- Musialik, K. The influence of chosen adipocytokines on blood pressure values in patients with metabolic syndrome. Kardiol. Pol. 2012, 70, 1237–1242. [Google Scholar]
- Srikanthan, K.; Feyh, A.; Visweshwar, H.; Shapiro, J.I.; Sodhi, K. Systematic Review of Metabolic Syndrome Biomarkers: A Panel for Early Detection, Management, and Risk Stratification in the West Virginian Population. Int. J. Med. Sci. 2016, 13, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Balakumar, M.; Prabhu, D.; Sathishkumar, C.; Prabu, P.; Rokana, N.; Kumar, R.; Raghavan, S.; Soundarajan, A.; Grover, S.; Batish, V.K.; et al. Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice. Eur. J. Nutr. 2018, 57, 279–295. [Google Scholar] [CrossRef]
- Degen, A.S.; Krynytska, I.; Kamyshnyi, A.M. Changes in the transcriptional activity of the entero-insular axis genes in streptozotocin-induced diabetes and after the administration of TNF-α non-selective blockers. Endocr. Regul. 2020, 54, 160–171. [Google Scholar] [CrossRef]
- Jia, Y.; Zai, W.; Shiqing, X.; Wenjian, Z.; Qing, F.; Honglin, L.; Liang, P.; Tingting, D.; Jinning, L. Advanced Glycation End Products Impair Glucose-Stimulated Insulin Secretion of a Pancreatic β-Cell Line INS-1-3 by Disturbance of Microtubule Cytoskeleton via p38/MAPK Activation. J. Diabetes Res. 2016, 2016, 9073037. [Google Scholar]
- Li, R.; Chinnathambi, A.; Alharbi, S.A.; Shair, O.H.M.; Veeraraghavan, V.P.; Surapaneni, K.M.; Rengarajan, T. Anti-inflammatory effects of rhaponticin on LPS-induced human endothelial cells through inhibition of MAPK/NF-κβ signaling pathways. J. Biochem. Mol. Toxic. 2021, 35, e22733. [Google Scholar] [CrossRef]
- Wronkowitz, N.; Rgens, S.W.G.; Romacho, T.; Villalobos, L.A.; Sánchez-Ferrer, C.F.; Peiró, C.; Sell, H.; Eckel, J. Soluble DPP4 induces inflammation and proliferation of human smooth muscle cells via protease-activated receptor 2. BBA-Mol. Basis Dis. 2014, 1842, 1613–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Lin, S.; Riyaphan, J.; Fu, Y.; Weng, C. Polyalthia Clerodane Diterpene Potentiates Hypoglycemia via Inhibition of Dipeptidyl Peptidase 4. Int. J. Mol. Sci. 2019, 20, 530. [Google Scholar] [CrossRef] [Green Version]
- Shih-Li, H.; Chuan-Chuan, H.; Chia-Ling, J.; Yu-Shan, T.; Kuo-Chiang, H. Porcine skin gelatin hydrolysate as a dipeptidyl peptidase IV inhibitor improves glycemic control in streptozotocin-induced diabetic rats. J. Funct. Foods 2014, 11, 235–242. [Google Scholar]
- Wang, T.Y.; Hsieh, C.H.; Hung, C.C.; Jao, C.L.; Chen, M.C.; Hsu, K.-C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm- and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Dong, J.; Liu, L.; Jia, C.; Zhu, J.; Lü, J. Isolation, Purification and Identification of Dipeptidyl Peptidase-4 Inhibitor from Lactic Acid Bacterial Metabolite. Food Sci. 2020, 41, 116–122. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | m/z | Class | Relative Content (%) |
|---|---|---|---|
| Adenine | 119.0357 | Imidazopyrimidines | 10.573 |
| Acetylcholine | 146.118 | Others | 7.479 |
| L-Phenylalanine | 166.0867 | Carboxylic acids and derivatives | 7.177 |
| Guanine | 152.0572 | Imidazopyrimidines | 6.418 |
| 2-Hydroxycinnamic acid | 165.0553 | Cinnamic acids and derivatives | 5.975 |
| L-Tyrosine | 182.0819 | Carboxylic acids and derivatives | 5.683 |
| 2′-Deoxyadenosine | 252.1099 | Purine nucleosides | 4.321 |
| Tetradecanedioic acid | 257.1758 | Fatty acyls | 2.790 |
| N-Benzylformamide | 136.0762 | Others | 2.450 |
| 2-[2-(1-isobutylcyclohexyl)-1-methylethylidene]hydrazine-1-carboxamide | 292.178 | Others | 2.301 |
| Indole-3-acrylic acid | 188.0713 | Others | 2.234 |
| N-cyclooctylurea | 171.1497 | Others | 1.982 |
| Palmitic acid | 274.2747 | Fatty acyls | 1.972 |
| Uracil | 113.0349 | Diazines | 1.628 |
| Choline | 104.1072 | Organonitrogen compounds | 1.521 |
| 3-Amino-4-methylpentanoic acid | 130.087 | Carboxylic acids and derivatives | 1.501 |
| Indole-3-lactic acid | 206.0819 | Indoles and derivatives | 1.281 |
| 4-Hexylresorcinol | 195.1235 | Others | 1.256 |
| Azelaic acid | 189.1129 | Fatty acyls | 1.054 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sha, J.; Song, J.; Huang, Y.; Zhang, Y.; Wang, H.; Zhang, Y.; Suo, H. Inhibitory Effect and Potential Mechanism of Lactobacillus plantarum YE4 against Dipeptidyl Peptidase-4. Foods 2022, 11, 80. https://doi.org/10.3390/foods11010080
Sha J, Song J, Huang Y, Zhang Y, Wang H, Zhang Y, Suo H. Inhibitory Effect and Potential Mechanism of Lactobacillus plantarum YE4 against Dipeptidyl Peptidase-4. Foods. 2022; 11(1):80. https://doi.org/10.3390/foods11010080
Chicago/Turabian StyleSha, Jia, Jiajia Song, Yechuan Huang, Yuhong Zhang, Hongwei Wang, Yu Zhang, and Huayi Suo. 2022. "Inhibitory Effect and Potential Mechanism of Lactobacillus plantarum YE4 against Dipeptidyl Peptidase-4" Foods 11, no. 1: 80. https://doi.org/10.3390/foods11010080
APA StyleSha, J., Song, J., Huang, Y., Zhang, Y., Wang, H., Zhang, Y., & Suo, H. (2022). Inhibitory Effect and Potential Mechanism of Lactobacillus plantarum YE4 against Dipeptidyl Peptidase-4. Foods, 11(1), 80. https://doi.org/10.3390/foods11010080