
4-Hydroxyderricin Promotes Apoptosis and Cell Cycle Arrest through Regulating PI3K/AKT/mTOR Pathway in Hepatocellular Cells

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Cell Culture
2.2. Cell Counting Kit-8 (CCK-8) Assay
2.3. Wound Healing Assay
2.4. Transwell Assay
2.5. Cell-Cycle Analysis
2.6. Apoptosis Analysis
2.7. TUNEL Assay
2.8. Immunofluorescence Staining
2.9. Western Blotting Analysis
2.10. Statistical Analysis
3. Results
3.1. Subsection
3.1.1. 4-HD Inhibited the Proliferation and Metastasis of HepG2 and Huh7
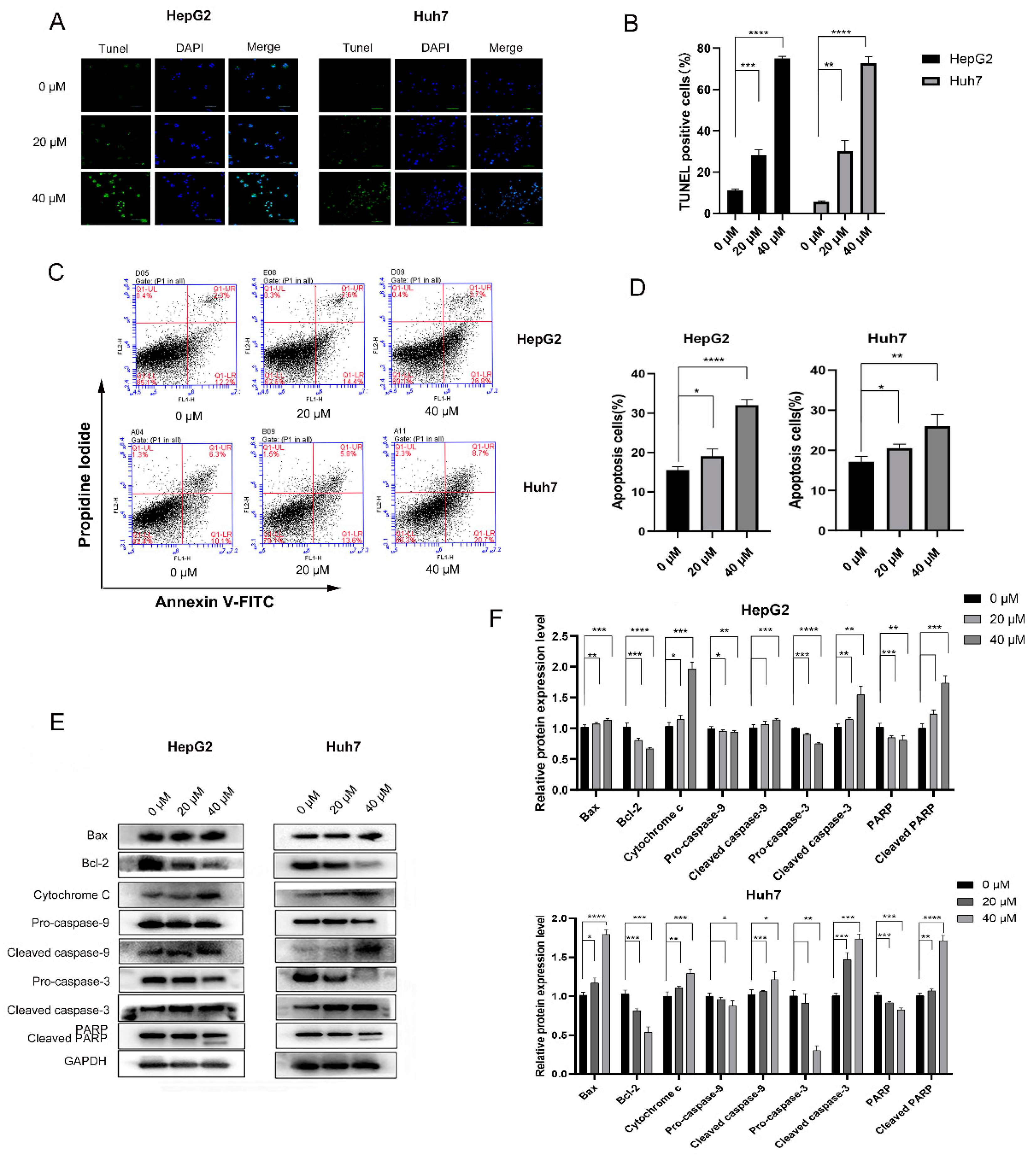
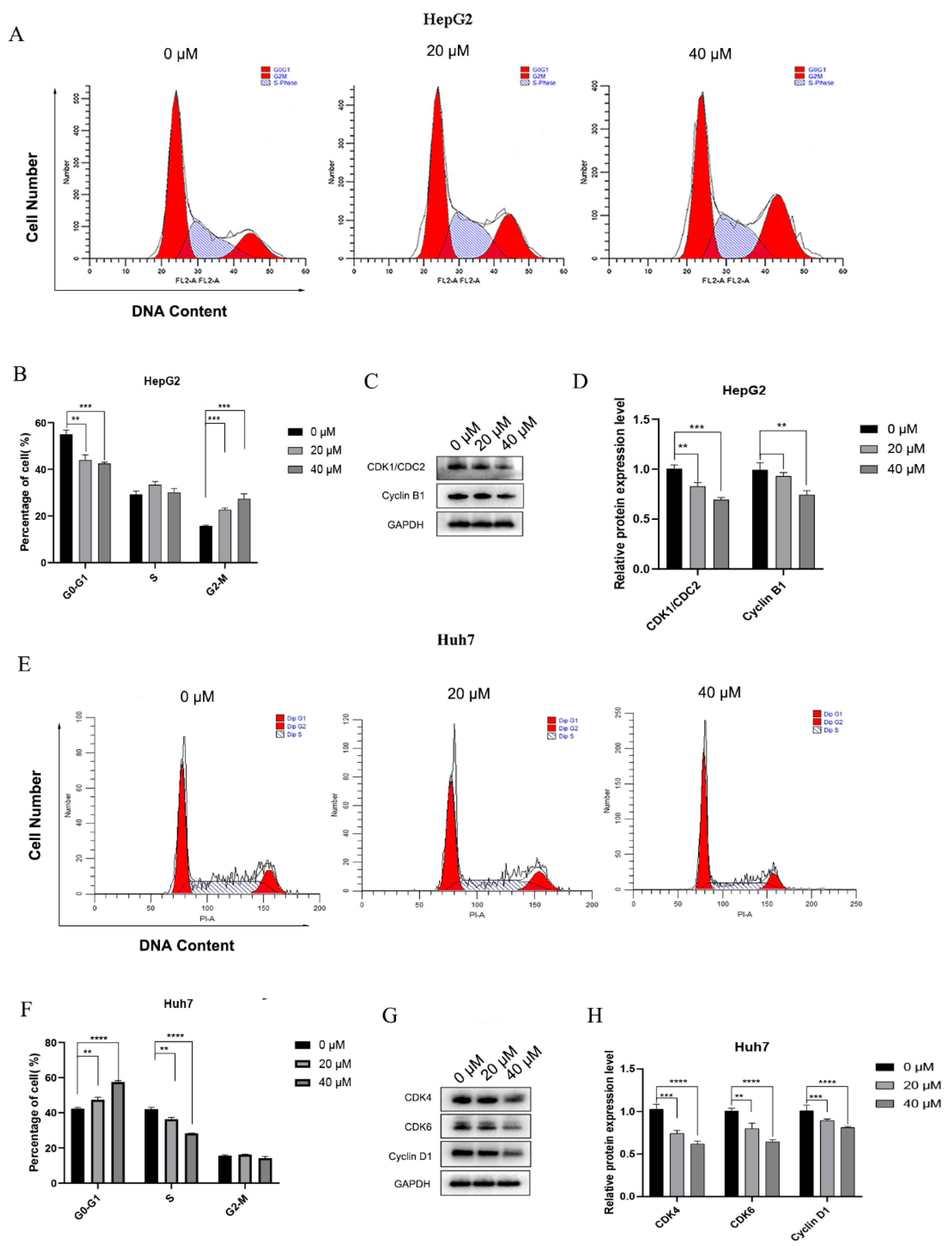
3.1.2. 4-HD Induced Apoptosis and Cell Cycle Arrest in HepG2 and Huh7 Cells
4-HD Induced Apoptosis in HepG2 and Huh7 Cells
4-HD Induced Cell Cycle Arrest in HCC Cells
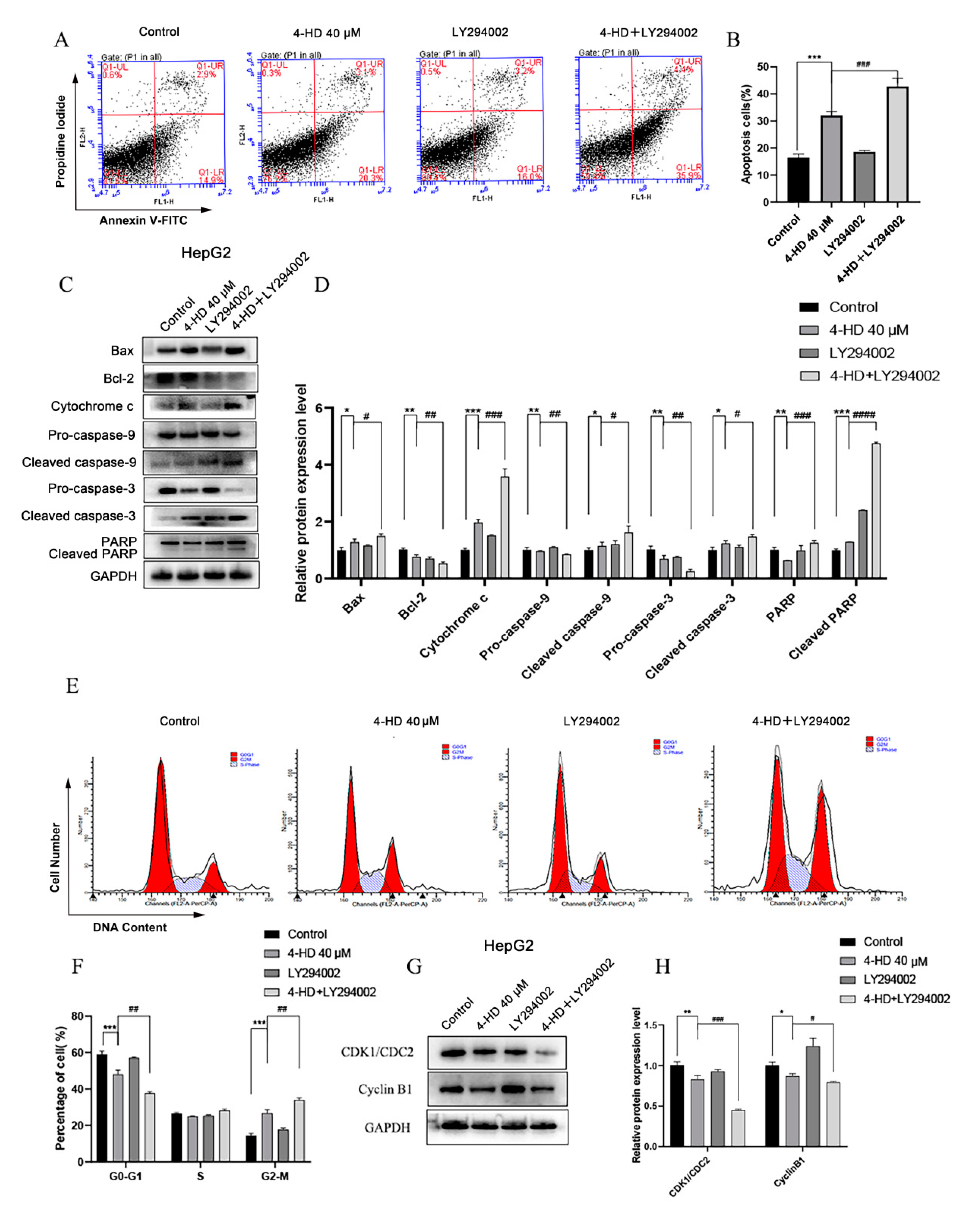
3.1.3. 4-HD Regulated the PI3K/AKT/mTOR Pathway
3.1.4. 4-HD Induced Apoptosis and Cycle Arrest of HCC Cells by Regulating the PI3K/AKT/mTOR Pathway
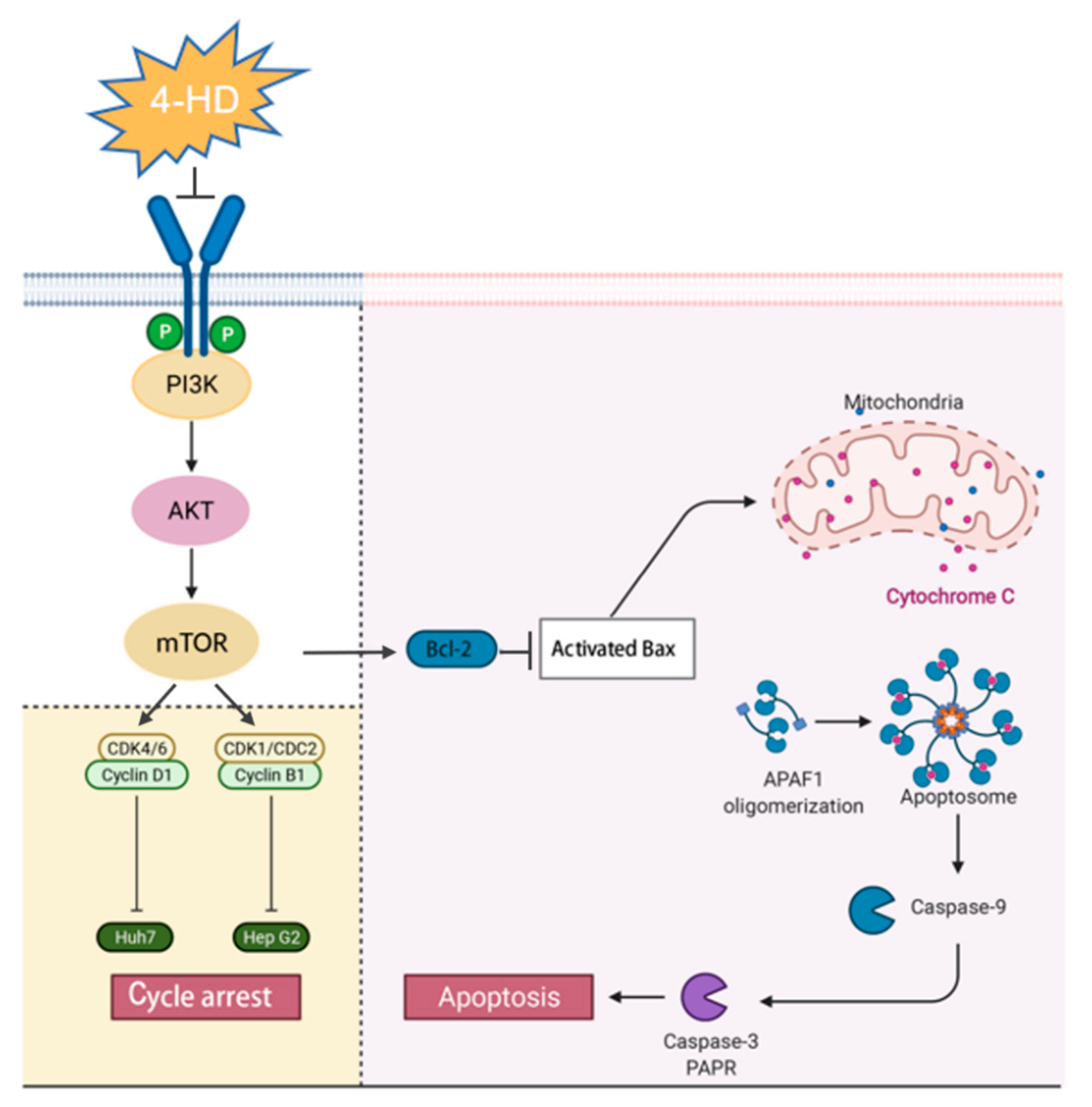
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ji, X.; Wei, X.; Qian, J.; Mo, X.; Kai, G.; An, F.; Lu, Y. 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone induced apoptosis and G1 cell cycle arrest through PI3K/AKT pathway in BEL-7402/5-FU cells. Food Chem. Toxicol. 2019, 131, 110533. [Google Scholar] [CrossRef]
- Tang, A.; Hallouch, O.; Chernyak, V.; Kamaya, A.; Sirlin, C.B. Epidemiology of hepatocellular carcinoma: Target population for surveillance and diagnosis. Abdom. Radiol. 2018, 43, 13–25. [Google Scholar] [CrossRef]
- Brunetti, O.; Gnoni, A.; Licchetta, A.; Longo, V.; Calabrese, A.; Argentiero, A.; Delcuratolo, S.; Solimando, A.G.; Casadei-Gardini, A.; Silvestris, N. Predictive and prognostic factors in HCC patients treated with sorafenib. Medicina 2019, 55, 707. [Google Scholar] [CrossRef]
- Díez, D.V.; Zubiaur, A.G.; Montalvo, S.M. Reactive perforating collagenosis: A rare side effect associated with sorafenib. Rev. Esp. Enferm. Dig. 2020, 112, 960–961. [Google Scholar] [CrossRef]
- Méndez-Blanco, C.; Fondevila, F.; Palomo, A.G.; González-Gallego, J.; Mauriz, J.L. Sorafenib resistance in hepatocarcinoma: Role of hypoxia-inducible factors. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Ma, L.; Liu, W.; Xu, A.; Ji, Q.; Ma, Y.; Tai, Y.; Wang, Y.; Shen, C.; Liu, Y.; Wang, T.; et al. Activator of thyroid and retinoid receptor increases sorafenib resistance in hepatocellular carcinoma by facilitating the Warburg effect. Cancer Sci. 2020, 111, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Sahin, I.D.; Christodoulou, M.S.; Guzelcan, E.A.; Koyas, A.; Karaca, C.; Passarella, D.; Cetin-Atalay, R. A small library of chalcones induce liver cancer cell death through Akt phosphorylation inhibition. Sci. Rep. 2020, 10, 11814. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.-Q.; Yao, Y.; Bao, Y.-L.; Song, Z.-B.; Yang, C.; Gao, X.-L.; Zhang, W.-J.; Sun, L.-G.; Yu, C.-L.; Huang, Y.-X.; et al. Juglanthraquinone C induces intracellular ROS increase and apoptosis by activating the Akt/Foxo signal pathway in HCC cells. Oxid. Med. Cell. Longev. 2016, 2016, 4941623. [Google Scholar] [CrossRef]
- He, Q.; Montalbano, J.; Corcoran, C.; Jin, W.; Huang, Y.; Sheikh, M.S. Effect of Bax deficiency on death receptor 5 and mitochondrial pathways during endoplasmic reticulum calcium pool depletion-induced apoptosis. Oncogene 2003, 22, 2674–2679. [Google Scholar] [CrossRef][Green Version]
- Reuter, S.; Eifes, S.; Dicato, M.; Aggarwal, B.B.; Diederich, M. Modulation of anti-apoptotic and survival pathways by curcumin as a strategy to induce apoptosis in cancer cells. Biochem. Pharmacol. 2008, 76, 1340–1351. [Google Scholar] [CrossRef]
- Zhang, Y.-S.; Ma, Y.-L.; Thakur, K.; Hussain, S.S.; Wang, J.; Zhang, Q.; Zhang, J.-G.; Wei, Z.-J. Molecular mechanism and inhibitory targets of dioscin in HepG2 cells. Food Chem. Toxicol. 2018, 120, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Asghar, U.; Witkiewicz, A.K.; Turner, N.C.; Knudsen, E.S. The history and future of targeting cyclin-dependent kinases in cancer therapy. Nat. Rev. Drug Discov. 2015, 14, 130–146. [Google Scholar] [CrossRef]
- Chen, J. The Cell-Cycle Arrest and apoptotic functions of p53 in tumor initiation and progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef]
- Barbieri, F.; Cagnoli, M.; Ragni, N.; Pedullà, F.; Foglia, G.; Alama, A. Expression of cyclin D1 correlates with malignancy in human ovarian tumours. Br. J. Cancer 1997, 75, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhao, Y.; Hu, W.; Zhang, Y.; Wu, X.; Lu, J.; Li, M.; Li, W.; Wu, W.; Wang, J.; et al. m(6)A RNA modification modulates PI3K/Akt/mTOR signal pathway in gastrointestinal cancer. Theranostics 2020, 10, 9528–9543. [Google Scholar] [CrossRef]
- Miricescu, D.; Totan, A.; Stanescu-Spinu, I.-I.; Badoiu, S.C.; Stefani, C.; Greabu, M. PI3K/AKT/mTOR signaling pathway in breast cancer: From molecular landscape to clinical aspects. Int. J. Mol. Sci. 2020, 22, 173. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin inhibits migration and invasion of non-small cell lung cancer cells through up-regulation of miR-206 and suppression of PI3K/AKT/mTOR signaling pathway. Acta Pharm. 2020, 70, 399–409. [Google Scholar] [CrossRef]
- Nepstad, I.; Hatfield, K.J.; Grønningsæter, I.S.; Reikvam, H. The PI3K-Akt-mTOR signaling pathway in human acute myeloid leukemia (AML) cells. Int. J. Mol. Sci. 2020, 21, 2907. [Google Scholar] [CrossRef]
- Wang, W.; Dong, X.; Liu, Y.; Ni, B.; Sai, N.; You, L.; Sun, M.; Yao, Y.; Qu, C.; Yin, X.; et al. Itraconazole exerts anti-liver cancer potential through the Wnt, PI3K/AKT/mTOR, and ROS pathways. Biomed. Pharmacother. 2020, 131, 110661. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Zhang, F.; Zhang, Y.-S.; Thakur, K.; Zhang, J.-G.; Liu, Y.; Kan, H.; Wei, Z.-J. Mechanism of juglone-induced cell cycle arrest and apoptosis in Ishikawa human endometrial cancer cells. J. Agric. Food Chem. 2019, 67, 7378–7389. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.D.; Gao, G.D.; Zhang, K.; Song, Y.W.; Li, H.B. Puerarin 6″-O-xyloside suppressed HCC via regulating proliferation, stemness, and apoptosis with inhibited PI3K/AKT/mTOR. Cancer Med. 2020, 9, 6399–6410. [Google Scholar] [CrossRef]
- Wu, T.; Dong, X.; Yu, D.; Shen, Z.; Yu, J.; Yan, S. Natural product pectolinarigenin inhibits proliferation, induces apoptosis, and causes G2/M phase arrest of HCC via PI3K/AKT/mTOR/ERK signaling pathway. Onco. Targets Ther. 2018, 11, 8633–8642. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Goto, T.; Ikutani, R.; Lin, S.; Takahashi, N.; Takahashi, H.; Jheng, H.-F.; Yu, R.; Taniguchi, M.; Baba, K.; et al. Xanthoangelol and 4-hydroxyderrcin suppress obesity-induced inflammatory responses. Obesity 2016, 24, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhang, X.; Jiang, Y.; Su, Q.; Li, Q.; Li, Z. Autophagy: Mechanisms and therapeutic potential of flavonoids in cancer. Biomolecules 2021, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, Y.; Pang, X.; Hua, P.; Gao, X.; Li, Q.; Li, Z. Simultaneous optimization of ultrasound-assisted extraction for flavonoids and antioxidant activity of Angelica keiskei using response surface methodology (RSM). Molecules 2019, 24, 3461. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, X.; Liu, X.; Wang, Z. Fabrication of Pickering emulsion based on particles combining pectin and zein: Effects of pectin methylation. Carbohydr. Polym. 2021, 256, 117515. [Google Scholar] [CrossRef]
- Pang, X.; Gao, X.; Liu, F.; Jiang, Y.; Wang, M.; Li, Q.; Li, Z. Xanthoangelol modulates Caspase-1-dependent pyroptotic death among hepatocellular carcinoma cells with high expression of GSDMD. J. Funct. Foods 2021, 84, 104577. [Google Scholar] [CrossRef]
- Inamori, Y.; Baba, K.; Tsujibo, H.; Taniguchi, M.; Nakata, K.; Kozawa, M. Antibacterial activity of two chalcones, xanthoangelol and 4-hydroxyderricin, isolated from the root of Angelica keiskei Koidzumi. Chem. Pharm. Bull. 1991, 39, 1604–1605. [Google Scholar] [CrossRef]
- Enoki, T.; Ohnogi, H.; Nagamine, K.; Kudo, Y.; Sugiyama, K.; Tanabe, M.; Kobayashi, E.; Sagawa, H.; Kato, I. Antidiabetic activities of chalcones isolated from a Japanese Herb, Angelica keiskei. J. Agric. Food Chem. 2007, 55, 6013–6017. [Google Scholar] [CrossRef]
- Aulifa, D.L.; Adnyana, I.K.; Levita, J.; Sukrasno, S. 4-Hydroxyderricin isolated from the sap of Angelica keiskei Koidzumi: Evaluation of its inhibitory activity towards dipeptidyl peptidase-IV. Sci. Pharm. 2019, 87, 30. [Google Scholar] [CrossRef]
- Ogawa, H.; Ohno, M.; Baba, K. Hypotensive and lipid regulatory actions of 4-hydroxyderricin, a chalcone from Angelica keiskei, in stroke-prone spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2005, 32, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.S.; Sawada, K.; Yamamoto, N.; Ashida, H. 4-Hydroxyderricin and xanthoangelol from Ashitaba (Angelica keiskei) suppress differentiation of preadiopocytes to adipocytes via AMPK and MAPK pathways. Mol. Nutr. Food Res. 2013, 57, 1729–1740. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Goto, T.; Yamakuni, K.; Takahashi, H.; Takahashi, N.; Jheng, H.-F.; Nomura, W.; Taniguchi, M.; Baba, K.; Murakami, S.; et al. 4-Hydroxyderricin, as a PPARγ agonist, promotes adipogenesis, adiponectin secretion, and glucose uptake in 3T3-L1 cells. Lipids 2016, 51, 787–795. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Samukawa, Y.; Yamashita, Y.; Ashida, H. 4-Hydroxyderricin and xanthoangelol isolated from Angelica keiskei prevent dexamethasone-induced muscle loss. Food Funct. 2020, 11, 5498–5512. [Google Scholar] [CrossRef]
- Kweon, M.; Lee, H.; Park, C.; Choi, Y.H.; Ryu, J.-H. A chalcone from ashitaba (Angelica keiskei) stimulates myoblast differentiation and inhibits dexamethasone-induced muscle atrophy. Nutrients 2019, 11, 2419. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Taniguchi, M.; Baba, K.; Kimura, Y. Antitumor and antimetastatic actions of xanthoangelol and 4-hydroxyderricin isolated from Angelica keiskei roots through the inhibited activation and differentiation of M2 macrophages. Phytomedicine 2015, 22, 759–767. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Q.; Fredimoses, M.; Gao, G.; Wang, K.; Chen, H.; Wang, T.; Oi, N.; Zykova, T.A.; Reddy, K.; et al. The ashitaba (Angelica keiskei) chalcones 4-hydroxyderricin and xanthoangelol suppress melanomagenesis by targeting BRAF and PI3K. Cancer Prev. Res. 2018, 11, 607–620. [Google Scholar] [CrossRef]
- Cai, J.; Luo, S.; Lv, X.; Deng, Y.; Huang, H.; Zhao, B.; Zhang, Q.; Li, G. Formulation of injectable glycyrrhizic acid-hydroxycamptothecin micelles as new generation of DNA topoisomerase I inhibitor for enhanced antitumor activity. Int. J. Pharmaceut. 2019, 571, 118693. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, X.; Wei, Y.; Liu, Q.; Jiang, Y.; Zhao, L.; Ulaah, S. Isolation, purification and the anti-hypertensive effect of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from Ruditapes philippinarum fermented with Bacillus natto. Food Funct. 2018, 9, 5230–5237. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Tian, Y.; Randell, E.; Zhou, H.; Sun, G. Unfavorable associations between serum trimethylamine N-oxide and L-carnitine levels with components of metabolic syndrome in the newfoundland population. Front. Endocrinol. 2019, 10, 168. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Y.; Wang, L.; Jin, Q.; Pan, M. Nobiletin selectively inhibits oral cancer cell growth by promoting apoptosis and DNA damage in vitro. Oral Surg. Oral Med. O. 2020, 130, 419–427. [Google Scholar] [CrossRef]
- Reddy, D.; Kumavath, R.; Ghosh, P.; Barh, D. Lanatoside C induces G2/M cell cycle arrest and suppresses cancer cell growth by attenuating MAPK, Wnt, JAK-STAT, and PI3K/AKT/mTOR signaling pathways. Biomolecules 2019, 9, 792. [Google Scholar] [CrossRef]
- Bertin-Ciftci, J.; Barré, B.; Le Pen, J.; Maillet, L.; Couriaud, C.; Juin, P.; Braun, F. pRb/E2F-1-mediated caspase-dependent induction of Noxa amplifies the apoptotic effects of the Bcl-2/Bcl-xL inhibitor ABT-737. Cell Death Differ. 2013, 20, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, Y.; Zhao, L. Tambulin targets histone deacetylase 1 inhibiting cell growth and inducing apoptosis in human lung squamous cell carcinoma. Front. Pharmacol. 2020, 11, 1188. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Heng, H.; Yu, H.; Dan, M.; Jie, C.; Zeng, Y.; Ning, H.; Liu, Z.; Wang, Z.; Lin, W. Anticancer effects of 10-hydroxycamptothecin induce apoptosis of human osteosarcoma through activating caspase-3, p53 and cytochrome c pathways. Oncol. Lett. 2017, 15, 2459–2464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, J.; Nussinov, R.; Ma, B. Release of cytochrome C from Bax pores at the mitochondrial membrane. Sci. Rep. 2017, 7, 2635. [Google Scholar] [CrossRef] [PubMed]
- Akihisa, T.; Kikuchi, T.; Nagai, H.; Ishii, K.; Tabata, K.; Suzuki, T. 4-Hydroxyderricin from Angelica keiskei roots induces caspase-dependent apoptotic cell death in HL60 human leukemia cells. J. Oleo Sci. 2011, 60, 71–77. [Google Scholar] [CrossRef]
- Jia, X.; Han, C.; Chen, J. Effect of tea on preneoplastic lesions and cell cycle regulators in rat liver. Cancer Epidem. Biomar. 2003, 11, 1663–1667. [Google Scholar] [CrossRef]
- Wu, G.-S.; Lu, J.-J.; Guo, J.-J.; Li, Y.-B.; Tan, W.; Dang, Y.-Y.; Zhong, Z.-F.; Xu, Z.-T.; Chen, X.-P.; Wang, Y.-T. Ganoderic acid DM, a natural triterpenoid, induces DNA damage, G1 cell cycle arrest and apoptosis in human breast cancer cells. Fitoterapia 2012, 83, 408–414. [Google Scholar] [CrossRef]
- Mari, A.; Mani, G.; Nagabhishek, S.N.; Balaraman, G.; Subramanian, N.; Mirza, F.B.; Sundaram, J.; Thiruvengadam, D. Carvacrol promotes cell cycle arrest and apoptosis through PI3K/AKT signaling pathway in MCF-7 breast cancer cells. Chin. J. Integr. Med. 2020, 1–8. [Google Scholar] [CrossRef]
- Arber, N.; Doki, Y.; Han, E.K.; Sgambato, A.; Zhou, P.; Kim, N.H.; Delohery, T.; Klein, M.G.; Holt, P.; Weinstein, I.B. Antisense to cyclin D1 inhibits the growth and tumorigenicity of human colon cancer cells. Cancer Res. 1997, 57, 1569–1574. [Google Scholar] [PubMed]
- Zhou, P.; Jiang, W.; Zhang, Y.J.; Kahn, S.M.; Schieren, I.; Santella, R.M.; Weinsteir, I.B. Antisense to cyclin D1 inhibits growth and reverses the transformed phenotype of human esophageal cancer cells. Oncogene 1995, 11, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Park, S.; Song, G.; Lim, W. Inhibitory effects of osthole on human breast cancer cell progression via induction of cell cycle arrest, mitochondrial dysfunction, and ER stress. Nutrients 2019, 11, 2777. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.-C.; Zhong, Q.-L.; Pang, W.-Q.; Dian, M.-J.; Li, J.; Han, L.-X.; Zhao, W.-T.; Zhang, X.-L.; Xiao, S.-J.; Xiao, D.; et al. MST4 inhibits human hepatocellular carcinoma cell proliferation and induces cell cycle arrest via suppression of PI3K/AKT pathway. J. Cancer 2020, 11, 5106–5117. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, M.; Lin, X.; Wang, Y.; He, X. A steroidal saponin isolated from Allium chinense simultaneously induces apoptosis and autophagy by modulating the PI3K/Akt/mTOR signaling pathway in human gastric adenocarcinoma. Steroids 2020, 161, 108672. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Yu, R. Lycopene inhibits epithelial–mesenchymal transition and promotes apoptosis in oral cancer via PI3K/AKT/m-TOR signal pathway. Drug Des. Devel. Ther. 2020, 14, 2461–2471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, M.; Guan, W. Naturally occurring aesculetin coumarin exerts antiproliferative effects in gastric cancer cells mediated via apoptotic cell death, cell cycle arrest and targeting PI3K/AKT/M-TOR signaling pathway. Acta Biochim. Pol. 2021, 68, 109–113. [Google Scholar] [CrossRef]
- Yu, J.-Z.; Ying, Y.; Liu, Y.; Sun, C.-B.; Dai, C.; Zhao, S.; Tian, S.-Z.; Peng, J.; Han, N.-P.; Yuan, J.-L.; et al. Antifibrotic action of Yifei Sanjie formula enhanced autophagy via PI3K-AKT-mTOR signaling pathway in mouse model of pulmonary fibrosis. Biomed. Pharmacother. 2019, 118, 109293. [Google Scholar] [CrossRef]
- Khan, K.; Quispe, C.; Javed, Z.; Iqbal, M.J.; Sadia, H.; Raza, S.; Irshad, A.; Salehi, B.; Reiner, Z.; Sharifi-Rad, J. Resveratrol, curcumin, paclitaxel and miRNAs mediated regulation of PI3K/Akt/mTOR pathway: Go four better to treat bladder cancer. Cancer Cell Int. 2020, 20, 560. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Jiang, Y.; Xu, Q.; Liu, F.; Pang, X.; Wang, M.; Li, Q.; Li, Z. 4-Hydroxyderricin Promotes Apoptosis and Cell Cycle Arrest through Regulating PI3K/AKT/mTOR Pathway in Hepatocellular Cells. Foods 2021, 10, 2036. https://doi.org/10.3390/foods10092036
Gao X, Jiang Y, Xu Q, Liu F, Pang X, Wang M, Li Q, Li Z. 4-Hydroxyderricin Promotes Apoptosis and Cell Cycle Arrest through Regulating PI3K/AKT/mTOR Pathway in Hepatocellular Cells. Foods. 2021; 10(9):2036. https://doi.org/10.3390/foods10092036
Chicago/Turabian StyleGao, Xiang, Yuhuan Jiang, Qi Xu, Feng Liu, Xuening Pang, Mingji Wang, Qun Li, and Zichao Li. 2021. "4-Hydroxyderricin Promotes Apoptosis and Cell Cycle Arrest through Regulating PI3K/AKT/mTOR Pathway in Hepatocellular Cells" Foods 10, no. 9: 2036. https://doi.org/10.3390/foods10092036
APA StyleGao, X., Jiang, Y., Xu, Q., Liu, F., Pang, X., Wang, M., Li, Q., & Li, Z. (2021). 4-Hydroxyderricin Promotes Apoptosis and Cell Cycle Arrest through Regulating PI3K/AKT/mTOR Pathway in Hepatocellular Cells. Foods, 10(9), 2036. https://doi.org/10.3390/foods10092036