
Synergistic Field Crop Pest Management Properties of Plant-Derived Essential Oils in Combination with Synthetic Pesticides and Bioactive Molecules: A Review

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Historical Background and Development of Natural Products in Agriculture
3. Sources and Chemical Composition of Plant-Derived Essential Oils
4. Pesticidal and Fungicidal Action Mechanisms of Plant-Derived Essential Oils
4.1. Mode of Action of Insecticidal Essential Oils
4.2. Mode of Action of Fungicidal Essential Oils
5. Synergistic and Hybridized Insect Pest Management Products of Botanical Essential Oils
5.1. As Homosynergistic Agents
5.2. As Enhancers of Commercial Insecticides
6. Synergistic and Hybridized Fungicidal Activity of Botanical Essential Oils
6.1. As Homosynergistic Agents
6.2. As Enhancers of Commercial Antifungal Agents
7. Novel Developments in Synergistic Insecticidal and Fungicidal Plant-Derived Essential Oils
8. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Van Dijk, M.; Morley, T.; Rau, M.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security. A review. Agron. Sustain. Dev. 2012, 33, 243–255. [Google Scholar] [CrossRef]
- Zhang, W. Global pesticide use: Profile, trend, cost/benefit and more. Proc. Int. Acad. Ecol. Environ. Sci. 2018, 8, 1–27. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.; Handa, N.; Kohli, S.; Yadav, P.; Bali, A.; Parihar, R.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Saiyed, H.N.; Sadhu, H.G.; Bhatnagar, V.K.; Dewan, A.; Venkaiah, K.; Kashyap, S.K. Cardiac toxicity following short-term exposure to methomyl in spraymen and rabbits. Hum. Exp. Toxicol. 1992, 11, 93–97. [Google Scholar] [CrossRef]
- Sanders, H.O. Toxicity of Pesticides to the Crustacean Gammarus Lacustris; Technical Papers of the Bureau of Sport Fisheries and Wildlife 1969, No. 25; US Department of Interior Fish and Wildlife Service: Washington, DC, USA, 1969. [Google Scholar]
- United Nations Sustainable Development. Goal 2: Zero Hunger. Available online: https://www.un.org/sustainabledevelopment/hunger/ (accessed on 10 June 2021).
- Lourenço, S.; Moldão-Martins, M.; Alves, V. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Pott, D.; Osorio, S.; Vallarino, J. From central to specialized metabolism: An overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front. Plant. Sci. 2019, 10, 835. [Google Scholar] [CrossRef]
- Preedy, V. Essential Oils in Food Preservation, Flavor and Safety 2016; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential oils as natural sources of fragrance compounds for cosmetics and cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, K. Investigation of volatile oil glands of Satureja hortensis L. (summer savory) and phytochemical comparison of different varieties. Int. J. Aromather. 2003, 13, 196–202. [Google Scholar] [CrossRef]
- Fahn, A. Structure and function of secretory cells. Adv. Bot. Res. 2000, 31, 37–75. [Google Scholar]
- Asgar, E.; Jalal, J.S. A review on recent research results on bio-effects of plant essential oils against major coleopteran insect pests. Toxin Rev. 2015, 34, 76–91. [Google Scholar]
- WHO. International Code of Conduct on the Distribution and Use of Pesticides: Guidelines for the Registration of Pesticides; World Health Organization: Rome, Italy, 2010. [Google Scholar]
- FAO. Manual on the Submission and Evaluation of Pesticide Residues Data for the Estimation of Maximum Residue Levels in Food and Feed; Food and Agriculture Organization: Rome, Italy, 2002. [Google Scholar]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial Agents—Myth Or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, Q.; Jacobsen, S.; Tang, Y. The impact and prospect of natural product discovery in agriculture. EMBO Rep. 2018, 19, e46824. [Google Scholar] [CrossRef]
- Boulogne, I.; Petit, P.; Ozier-Lafontaine, H.; Desfontaines, L.; Loranger-Merciris, G. Insecticidal and antifungal chemicals produced by plants: A review. Environ. Chem. Lett. 2012, 10, 325–347. [Google Scholar] [CrossRef]
- Bynum, B. Shedding new light on the story of penicillin. Lancet 2007, 369, 1991–1992. [Google Scholar] [CrossRef]
- Sparks, T.; Hahn, D.; Garizi, N. Natural products, their derivatives, mimics and synthetic equivalents: Role in agrochemical discovery. Pest Manag. Sci. 2016, 73, 700–715. [Google Scholar] [CrossRef]
- Rao, G.V.; Rupela, O.P.; Rao, V.R.; Reddy, Y.V. Role of biopesticides in crop protection: Present status and future prospects. Indian J. Plant. Prot. 2007, 35, 1–9. [Google Scholar]
- Miller, G. Living in the Environment, 12th ed.; Thomson Learning: Belmont, CA, USA, 2002. [Google Scholar]
- Maheshwari, R. Fungi: Experimental Methods in Biology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Pandey, A.K.; Sain, S.K.; Singh, P. A perspective on integrated disease management in agriculture. Bio Bull. 2016, 2, 13–29. [Google Scholar]
- Kirst, H. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Áy, Z.; Mihály, R.; Cserháti, M.; Kótai, É.; Pauk, J. The effect of high concentrations of glufosinate ammonium on the yield components of transgenic spring wheat (Triticum aestivum L.) constitutively expressing the bar gene. Sci. World J. 2012, 2012, 657945. [Google Scholar] [CrossRef]
- Hoerlein, G. Glufosinate (phosphinothricin), a natural amino acid with unexpected herbicidal properties. Rev. Environ. Contam. Toxicol. 1994, 138, 73–145. [Google Scholar] [CrossRef]
- Owen, W.; Meyer, K.; Slanec, T.; Meyer, S.; Wang, N.; Fitzpatrick, G.; Niyaz, N.; Nugent, J.; Ricks, M.; Rogers, R.; et al. Synthesis and biological activity of analogs of the antifungal antibiotic uk-2a. iii. impact of modifications to the macrocycle isobutyryl ester position. Pest Manag. Sci. 2019, 76, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Tongnuanchan, P.; Benjakul, S. Essential oils: Extraction, bioactivities, and their uses for food preservation. J. Food Sci. 2014, 79, 1231–1249. [Google Scholar] [CrossRef]
- Masango, P. Cleaner production of essential oils by steam distillation. J. Clean. Prod. 2005, 13, 833–839. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Mohamed, A.A.; El-Emary, G.A.; Ali, H.F. Influence of some citrus essential oils on cell viability, glutathione-s-transferase and lipid peroxidation in ehrlich ascites carcinoma cells. J. Am. Sci. 2010, 6, 820–826. [Google Scholar]
- Ngoh, S.; Choo, L.; Pang, F.; Huang, Y.; Kini, M.; Ho, S. Insecticidal and repellent properties of nine volatile constituents of essential oils against the american cockroach, Periplaneta americana (L.). Pestic. Sci. 1998, 54, 261–268. [Google Scholar] [CrossRef]
- Butnariu, M.; Sarac, I. Essential oils from plants. J. Biotechnol. Biomed. Sci. 2018, 1, 35–43. [Google Scholar] [CrossRef]
- Borzoui, E.; Naseri, B.; Abedi, Z.; Karimi-Pormehr, M. Lethal and sublethal effects of essential oils from Artemisia khorassanica and Vitex pseudo-negundo against Plodia interpunctella (Lepidoptera: Pyralidae). Environ. Entomol. 2016, 45, 1220–1226. [Google Scholar] [CrossRef]
- Daferera, D.; Ziogas, B.; Polissiou, M. GC-MS analysis of essential oils from some greek aromatic plants and their fungitoxicity on Penicillium digitatum. J. Agric. Food Chem. 2000, 48, 2576–2581. [Google Scholar] [CrossRef] [PubMed]
- Hermans, P.E.; Keys, T.F. Antifungal agents used for deep-seated mycotic infections. Mayo Clin. Proc. 1983, 58, 223–231. [Google Scholar] [PubMed]
- Kyle, A.; Dahl, M. Topical therapy for fungal infections. Am. J. Clin. Dermatol. 2004, 5, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Niroumand, M.C.; Farzaei, M.; Karimpour Razkenari, E.; Amin, G.; Khanavi, M.; Akbarzadeh, T.; Shams-Ardekani, M. An evidence-based review on medicinal plants used as insecticide and insect repellent in traditional iranian medicine. Iran. Red Crescent Med. J. 2016, 18, e22361. [Google Scholar]
- Kim, J.; Jung, C.; Koh, Y.; Lee, S. Molecular, biochemical and histochemical characterization of two acetylcholinesterase cDNAs from the German cockroach Blattella germanica. Insect Mol. Biol. 2006, 15, 513–522. [Google Scholar] [CrossRef]
- Pezzementi, L.; Rowland, M.; Wolfe, M.; Tsigelny, I. Inactivation of an invertebrate acetylcholinesterase by sulfhydryl reagents: The roles of two cysteines in the catalytic gorge of the enzyme. Invertebr. Neurosci. 2006, 6, 47–55. [Google Scholar] [CrossRef]
- Pang, Y.; Singh, S.; Gao, Y.; Lassiter, T.; Mishra, R.; Zhu, K.; Brimijoin, S. Selective and irreversible inhibitors of aphid acetylcholinesterases: Steps toward human-safe insecticides. PLoS ONE 2009, 4, e4349. [Google Scholar] [CrossRef]
- Polsinelli, G.; Singh, S.; Mishra, R.; Suranyi, R.; Ragsdale, D.; Pang, Y.; Brimijoin, S. Insect-specific irreversible inhibitors of acetylcholinesterase in pests including the bed bug, the eastern yellowjacket, German and American cockroaches, and the confused flour beetle. Chem. Biol. Interact. 2010, 187, 142–147. [Google Scholar] [CrossRef]
- Pang, Y.; Brimijoin, S.; Ragsdale, D.W.; Yan Zhu, K.; Suranyi, R. Novel and viable acetylcholinesterase target site for developing effective and environmentally safe insecticides. Curr. Drug Targets 2012, 13, 471–482. [Google Scholar] [CrossRef]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef]
- Gnagey, A.; Forte, M.; Rosenberry, T. Isolation and characterization of acetylcholinesterase from Drosophila. J. Biol. Chem. 1987, 262, 13290–13298. [Google Scholar] [CrossRef]
- Bourguet, D.; Roig, A.; Toutant, J.; Arpagaus, M. Analysis of molecular forms and pharmacological properties of acetylcholinesterase in several mosquito species. Neurochem. Int. 1997, 31, 65–72. [Google Scholar] [CrossRef]
- Marcel, V.; Palacios, L.; Pertuy, C.; Masson, P.; Fournier, D. Two invertebrate acetylcholinesterases show activation followed by inhibition with substrate concentration. Biochem. J. 1998, 329, 329–334. [Google Scholar] [CrossRef]
- Marrs, T.; Maynard, R. Neurotranmission systems as targets for toxicants: A review. Cell Biol. Toxicol. 2013, 29, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Re, L.; Barocci, S.; Sonnino, S.; Mencarelli, A.; Vivani, C.; Paolucci, G.; Scarpantonio, A.; Rinaldi, L.; Mosca, E. Linalool modifies the nicotinic receptor-ion channel kinetics at the mouse neuromuscular junction. Pharmacol. Res. 2000, 42, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Evans, P.D. Biogenic amines in the insect nervous system. Adv. Insect Physiol. 1980, 15, 317–473. [Google Scholar]
- Evans, P.D. Multiple receptor types for octopamine in the locust. J. Physiol. 1981, 318, 99–122. [Google Scholar] [CrossRef]
- Farooqui, T. Octopamine-mediated neuromodulation of insect senses. Neurochem. Res. 2007, 32, 1511–1529. [Google Scholar] [CrossRef]
- Ohta, H.; Ozoe, Y. Molecular signalling, pharmacology, and physiology of octopamine and tyramine receptors as potential insect pest control targets. Adv. Insect Physiol. 2014, 46, 73–166. [Google Scholar]
- Lange, A.; Orchard, I. Identified octopaminergic neurons modulate contractions of locust visceral muscle via adenosine 3′,5′-monophosphate (cyclic AMP). Brain Res. 1986, 363, 340–349. [Google Scholar] [CrossRef]
- Nathanson, J. Octopamine receptors, adenosine 3′,5′-monophosphate, and neural control of firefly flashing. Science 1979, 203, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Orchard, I.; Carlisle, J.; Loughton, B.; Gole, J.; Downer, R. In vitro studies on the effects of octopamine on locust fat body. Gen. Comp. Endocrinol. 1982, 48, 7–13. [Google Scholar] [CrossRef]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.; Cherubini, E. The GABA excitatory/inhibitory shift in brain maturation and neurological disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef]
- Sattelle, D. GABA receptors of insects. Adv. Insect Physiol. 1990, 22, 1–113. [Google Scholar]
- Sigel, E.; Steinmann, M. Structure, function, and modulation of GABAA receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef]
- Yu, H.; Xu, J.; Wang, X.; Ma, Y.; Yu, D.; Fei, D.; Zhang, S.; Wang, W. Identification of four ATP-binding cassette transporter genes in Cnaphalocrocis medinalis and their expression in response to insecticide treatment. J. Insect Sci. 2017, 17, 44. [Google Scholar] [CrossRef]
- Merzendorfer, H. Chitin synthesis inhibitors: Old molecules and new developments. Insect Sci. 2012, 20, 121–138. [Google Scholar] [CrossRef]
- Campolo, O.; Giunti, G.; Russo, A.; Palmeri, V.; Zappalà, L. Essential oils in stored product insect pest control. J. Food Qual. 2018, 2018, 6906105. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, J.; Kong, W.; Zhao, G.; Yang, M. Mechanisms of antifungal and anti-aflatoxigenic properties of essential oil derived from turmeric (Curcuma longa L.) on aspergillus flavus. Food Chem. 2017, 220, 1–8. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Singh, P.; Kedia, A.; Dubey, N. Assessment of some essential oils as food preservatives based on antifungal, antiaflatoxin, antioxidant activities and in vivo efficacy in food system. Food Res. Int. 2012, 49, 201–208. [Google Scholar] [CrossRef]
- Lang, G.; Buchbauer, G. A review on recent research results (2008–2010) on essential oils as antimicrobials and antifungals. A review. Flavour Fragr. J. 2011, 27, 13–39. [Google Scholar] [CrossRef]
- Wu, X.Z.; Cheng, A.X.; Sun, L.M.; Lou, H.X. Effect of plagiochin E, an antifungal macrocyclic bis (bibenzyl), on cell wall chitin synthesis in Candida albicans. Acta Pharmacol. Sin. 2008, 12, 78–1485. [Google Scholar] [CrossRef] [PubMed]
- Kerekes, E.; Deák, É.; Takó, M.; Tserennadmid, R.; Petkovits, T.; Vágvölgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.B.; Karuppayil, S.M. Small molecules inhibit growth, viability and ergosterol biosynthesis in Candida albicans. Springerplus 2013, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, A.; Kumar, P.; Bhatt, R.P.; Manzoor, N. Antifungal activity of Coriaria nepalensis essential oil by disrupting ergosterol biosynthesis and membrane integrity against Candida. Yeast 2011, 28, 611–617. [Google Scholar] [CrossRef]
- Freires, I.; Murata, R.; Furletti, V.; Sartoratto, A.; Alencar, S.; Figueira, G.; de Oliveira Rodrigues, J.; Duarte, M.; Rosalen, P. Coriandrum Sativum L. (coriander) essential oil: Antifungal activity and mode of action on Candida spp., and molecular targets affected in human whole-genome expression. PLoS ONE 2014, 9, e99086. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ahn, Y. Growth-inhibiting effects of cinnamomum cassia bark-derived materials on human intestinal bacteria. J. Agric. Food Chem. 1998, 46, 8–12. [Google Scholar] [CrossRef]
- Bang, K.; Lee, D.; Park, H.; Rhee, Y. Inhibition of fungal cell wall synthesizing enzymes bytrans-cinnamaldehyde. Biosci. Biotechnol. Biochem. 2000, 64, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed]
- Winans, S.C.; Bassler, B.L. Mob psychology. J. Bacteriol. 2002, 184, 873–883. [Google Scholar] [CrossRef]
- Li, Z.; Nair, S. Quorum sensing: How bacteria can coordinate activity and synchronize their response to external signals? Protein Sci. 2012, 21, 1403–1417. [Google Scholar] [CrossRef]
- Lumjiaktase, P.; Aguilar, C.; Battin, T.; Riedel, K.; Eberl, L. Construction of self-transmissible green fluorescent protein-based biosensor plasmids and their use for identification of N-acyl homoserine-producing bacteria in lake sediments. Appl. Environ. Microbiol. 2010, 76, 6119–6127. [Google Scholar] [CrossRef]
- Uroz, S.; D’Angelo-Picard, C.; Carlier, A.; Elasri, M.; Sicot, C.; Petit, A.; Oger, P.; Faure, D.; Dessaux, Y. Novel bacteria degrading N-acylhomoserine lactones and their use as quenchers of quorum-sensing-regulated functions of plant-pathogenic bacteria. Microbiology 2003, 149, 1981–1989. [Google Scholar] [CrossRef]
- Rasmussen, T.; Skindersoe, M.; Bjarnsholt, T.; Phipps, R.; Christensen, K.; Jensen, P.; Andersen, J.; Koch, B.; Larsen, T.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef]
- Kumrungsee, N.; Pluempanupat, W.; Koul, O.; Bullangpoti, V. Toxicity of essential oil compounds against diamondback moth, plutella xylostella, and their impact on detoxification enzyme activities. J. Pest Sci. 2014, 87, 721–729. [Google Scholar] [CrossRef]
- Tak, J.; Isman, M. Enhanced cuticular penetration as the mechanism for synergy of insecticidal constituents of rosemary essential oil in Trichoplusia ni. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tak, J.; Isman, M. Penetration-enhancement underlies synergy of plant essential oil terpenoids as insecticides in the cabbage looper, Trichoplusia ni. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Singh, R.; Koul, O.; Rup, P.; Jindal, J. Toxicity of some essential oil constituents and their binary mixtures against Chilo partellus (lepidoptera: Pyralidae). Int. J. Trop. Insect Sci. 2009, 29, 93–101. [Google Scholar] [CrossRef]
- Hummelbrunner, L.; Isman, M. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef]
- Liu, C.; Mishra, A.; Tan, R.; Tang, C.; Yang, H.; Shen, Y. Repellent and insecticidal activities of essential oils from Artemisia princeps and Cinnamomum camphora and their effect on seed germination of wheat and broad bean. Bioresour. Technol. 2006, 97, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, R.; Li, W.; Cheng, D.; Mao, R.; Zhang, Z. Addition of cinnamon oil improves toxicity of rotenone tospodoptera litura (lepidoptera: Noctuidae) larvae. Fla. Entomol. 2017, 100, 515–521. [Google Scholar] [CrossRef]
- Abbassy, M.; Abdelgaleil, S.; Rabie, R. Insecticidal and synergistic effects of Majorana hortensis essential oil and some of its major constituents. Entomol. Exp. Appl. 2009, 131, 225–232. [Google Scholar] [CrossRef]
- Andrés, M.; Rossa, G.; Cassel, E.; Vargas, R.; Santana, O.; Díaz, C.; González-Coloma, A. Biocidal effects of piper hispidinervum (piperaceae) essential oil and synergism among its main components. Food Chem. Toxicol. 2017, 109, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Xu, J.; Yang, Y.; Shao, Y.; Zhou, F.; Wang, J. Toxicity and synergistic effect of elsholtzia ciliata essential oil and its main components against the adult and larval stages of tribolium castaneum. Foods 2020, 9, 345. [Google Scholar] [CrossRef]
- El-Meniawi, F.A.; Ismail, S.M. Toxic and biochemical impact of certain plant essential oils on Bemisia tabaci genn. (Hom., Aleyrodidae). J. Pest Cont. Environ. Sci. 2006, 14, 18–99. [Google Scholar]
- Faraone, N.; Hillier, N.; Cutler, G. Plant essential oils synergize and antagonize toxicity of different conventional insecticides against Myzus persicae (hemiptera: Aphididae). PLoS ONE 2015, 10, e0127774. [Google Scholar] [CrossRef] [PubMed]
- Martín-López, B.; Varela, I.; Marnotes, S.; Cabaleiro, C. Use of oils combined with low doses of insecticide for the control ofMyzus persicae and PVY epidemics. Pest Manag. Sci. 2006, 62, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Cunha, J.; Carvalho, S.; Zandonadi, C.; Martins, R.; Chang, R. Ocimum basilicum essential oil combined with deltamethrin to improve the management of Spodoptera frugiperda. Ciência Agrotecnologia 2017, 41, 665–675. [Google Scholar] [CrossRef]
- Macedo Silva, S.; Cunha, J.; Zandonadi, C.; Assunção, H.; Gregorio Marques, M. Synergistic effects of binary mixtures of linalool with pyrethroids against fall armyworm. Biosci. J. 2020, 36, 228–237. [Google Scholar] [CrossRef]
- Ismail, S.M. Synergistic efficacy of plant essential oils with cypermethrin and chlorpyrifos against Spodoptera littoralis, field populations in egypt. Int. J. Adv. Biol. Biomed. Res. 2021, 9, 128–137. [Google Scholar]
- Mantzoukas, S.; Kalyvas, N.; Ntoukas, A.; Lagogiannis, I.; Farsalinos, K.; Eliopoulos, P.; Poulas, K. Combined toxicity of cannabidiol oil with three bio-pesticides against adults of Sitophilus zeamais, Rhyzopertha dominica, Prostephanus truncatus and Trogoderma granarium. Int. J. Environ. Res. Public Health 2020, 17, 6664. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Gwiazdowska, D.; Marchwińska, K.; Juś, K.; Gwiazdowski, R.; Waśkiewicz, A. Antifungal activity of selected essential oils against fusarium culmorum and f. graminearum and their secondary metabolites in wheat seeds. Arch. Microbiol. 2019, 201, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; López, D.; Anaya, L.; López, D.; Quijano, R.; Ovando, A. Chitosan composite films: Physicochemical characterization and their use as coating in papaya Maradol stored at room temperature. Emir. J. Food Agric. 2017, 29, 779–791. [Google Scholar] [CrossRef]
- Nardoni, S.; D'Ascenzi, C.; Caracciolo, I.; Mannaioni, G.; Papini, R.; Pistelli, L.; Najar, B.; Mancianti, F. Activity of selected essential oils on spoiling fungi cultured from marzolino cheese. Ann. Agric. Environ. Med. 2018, 25, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Bedoya-Serna, C.; Dacanal, G.; Fernandes, A.; Pinho, S. Antifungal activity of nanoemulsions encapsulating oregano (Origanum vulgare) essential oil: In vitro study and application in Minas Padrão cheese. Braz. J. Microbiol. 2018, 49, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Bounar, R.; Krimat, S.; Boureghda, H.; Dob, T. Chemical analyses, antioxidant and antifungal effects of oregano and thyme essential oils alone or in combination against selected Fusarium species. Int. Food Res. J. 2020, 27, 66–77. [Google Scholar]
- Yen, T.; Chang, S. Synergistic effects of cinnamaldehyde in combination with eugenol against wood decay fungi. Bioresour. Technol. 2008, 99, 232–236. [Google Scholar] [CrossRef]
- Hartati, S.; Sukamto, N.; Karyani, N.; Zulhisnain, N. Efficacy of single formula of clove, eucalyptus, neem and citronella oil against budok disease of patchouli. Bul. Penelit. Tanam. Rempah Dan Obat 2018, 28, 153. [Google Scholar] [CrossRef]
- Sukatta, U.; Haruthaithanasa, V.; Chantarapanont, W.; Dilokkunanant, U.; Suppakul, P. Antifungal activity of clove and cinnamon oil and their synergistic against postharvest decay fungi of grape in vitro. Kasetsart J. Nat. Sci. 2008, 42, 169–174. [Google Scholar]
- Yu, D.; Wang, J.; Shao, X.; Xu, F.; Wang, H. Antifungal modes of action of tea tree oil and its two characteristic components against Botrytis cinerea. J. Appl. Microbiol. 2015, 119, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The mechanism of antifungal action of essential oil from dill (Anethum graveolens L.) on aspergillus flavus. PLoS ONE 2012, 7, e30147. [Google Scholar]
- Nakahara, K.; Alzoreky, N.; Yoshihashi, T.; Nguyen, H.; Trakoontivakorn, G. Chemical composition and antifungal activity of essential oil from Cymbopogon nardus (citronella grass). Jpn. Agric. Res. Q. JARQ 2013, 37, 249–252. [Google Scholar] [CrossRef]
- Khan, M.; Ahmad, I. Antifungal activity of essential oils and their synergy with fluconazole against drug-resistant strains of Aspergillus fumigatus and Trichophyton rubrum. Appl. Microbiol. Biotechnol. 2011, 90, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Gadban, L.C.; Camiletti, B.X.; Bigatton, E.D.; Distéfano, S.G.; Lucini, E.I. Combinations of Tagetes filifolia lag. essential oil with chemical fungicides to control Colletotrichum truncatum and their effects on the biocontrol agent Trichoderma harzianum. J. Plant Prot. Res. 2020, 60, 41–50. [Google Scholar]
- Liu, X.; Yan, D.; Ouyang, C.; Yang, D.; Wang, Q.; Li, Y.; Guo, M.; Cao, A. Oils extracted from Eupatorium adenophorum leaves show potential to control Phythium myriotylum in commercially-grown ginger. PLoS ONE 2017, 12, e0176126. [Google Scholar]
- Camiletti, B.; Asensio, C.; Gadban, L.; Pecci, M.; Conles, M.; Lucini, E. Essential oils and their combinations with iprodione fungicide as potential antifungal agents against withe rot (Sclerotium cepivorum berk) in garlic (Allium sativum L.) crops. Ind. Crop. Prod. 2016, 85, 117–124. [Google Scholar] [CrossRef]
- Shin, S. Anti-aspergillus activities of plant essential oils and their combination effects with ketoconazole or amphotericin b. Arch. Pharmacal Res. 2003, 26, 389–393. [Google Scholar] [CrossRef]
- Konecka, E.; Kaznowski, A.; Grzesiek, W.; Nowicki, P.; Czarniewska, E.; Baranek, J. Synergistic interaction between carvacrol and Bacillus thuringiensis crystalline proteins against Cydia pomonella and Spodoptera exigua. BioControl 2020, 65, 447–460. [Google Scholar] [CrossRef]
- Konecka, E.; Kaznowski, A.; Tomkowiak, D. Insecticidal activity of mixtures of Bacillus thuringiensis crystals with plant oils of Sinapis alba and Azadirachta indica. Ann. Appl. Biol. 2019, 174, 364–371. [Google Scholar] [CrossRef]
- Radha, R.; Murugan, K.; Wei, H.; Amerasan, D.; Madhiyazhagan, P.; Chen, F.; Kovendan, K.; Nataraj, T.; Nareshkumar, A.; Hwang, J.; et al. Insecticidal activity of essential oils and entomopathogenic fungiagainst cowpea bruchid, Callosobruchus maculatus (f.) (insecta:coleoptera: Bruchidae). Int. J. Curr. Innov. Res. 2014, 1, 12–19. [Google Scholar]
- Yang, F.L.; Li, X.G.; Zhu, F.; Lei, C.L. Structural characterization of nanoparticles loaded with garlic essential oil and their insecticidal activity against Tribolium castaneum (Herbst) (coleoptera: Tenebrionidae). J. Agric. Food Chem. 2009, 57, 10156–10162. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.M.; Salem, N.Y.; Abdel-Aziz, N.F.; Ibrahim, S.S. Application of new nano pesticide Geranium oil loadedsolid lipid nanoparticles for control the black cutworm Agrotis ipsilon (Hub.) (Lepi., Noctuidae). EurAsian J. BioSci. 2019, 13, 1453–1461. [Google Scholar]
- Pierattini, E.; Bedini, S.; Venturi, F.; Ascrizzi, R.; Flamini, G.; Bocchino, R.; Girardi, J.; Giannotti, P.; Ferroni, G.; Conti, B. Sensory quality of essential oils and their synergistic effect with diatomaceous earth, for the control of stored grain insects. Insects 2019, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Sayed, W.; El-Bendary, H.; El-Helaly, A. Increasing the efficacy of the cotton leaf worm Spodoptera littoralis nucleopolyhedrosis virus using certain essential oils. Egypt. J. Biol. Pest Control 2020, 30, 8. [Google Scholar] [CrossRef]
- Al-alawi, M.S. Evaluation of Jordanian isolates of Beauveria bassiana (Balsamo) Vuillemin and their interaction with essential plant oils when combined for the two spotted spider mite, Tetranychus urticae koch control. Adv. Environ. Biol. 2019, 13, 4–10. [Google Scholar]
- Nasseri, M.; Golmohammadzadeh, S.; Arouiee, H.; Jaafari, M.R.; Neamati, H. Antifungal activity of Zataria multiflora essential oil-loaded solid lipid nanoparticles in-vitro condition. Iran. J. Basic Med. Sci. 2016, 19, 1231–1237. [Google Scholar] [PubMed]
- Luque-Alcaraz, A.; Cortez-Rocha, M.; Velázquez-Contreras, C.; Acosta-Silva, A.; Santacruz-Ortega, H.; Burgos-Hernández, A.; Argüelles-Monal, W.; Plascencia-Jatomea, M. Enhanced antifungal effect of chitosan/pepper tree (Schinus molle) essential oil bionanocomposites on the viability of Aspergillus parasiticus spores. J. Nanomater. 2016, 2016, 6060137. [Google Scholar] [CrossRef]
- Thanighaiarassu, R.R.; Sivamai, P.; Devika, R.; Nambikkairaj, B. Green synthesis of gold nanoparticles characterization by using plant essential oil menthapiperita and their antifungal activity against human pathogenic fungi. J. Nanomed. Nanotechnol. 2014, 5, 229. [Google Scholar] [CrossRef]
- Amir, A.; Naghmeh, M. Encapsulation of Satureja khuzestanica essential oil in chitosan nanoparticles with enhanced antifungal activity. Int. J. Nutr. Food Eng. 2017, 11, 331–336. [Google Scholar] [CrossRef]
- Kalagatur, N.; Nirmal Ghosh, O.; Sundararaj, N.; Mudili, V. Antifungal activity of chitosan nanoparticles encapsulated with Cymbopogon martinii essential oil on plant pathogenic fungi Fusarium graminearum. Front. Pharmacol. 2018, 9, 610. [Google Scholar] [CrossRef]
- Latha, M.K.V.; Lal, A.A. Efficacy of micro algae and thyme essential oil in the management of alternaria leaf spot of broccoli (Brassica oleracea var. italica). Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 297–303. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, J.; Morrell, J.J.; Freitag, C.M.; Karchesy, J.J. An integrated approach using bacillus subtilis B26 and essential oils to limit fungal discoloration of wood. BioResources 2012, 7, 3132–3141. [Google Scholar]
- Nissa, A.; Utami, R.; Sari, A.; Nursiwi, A. Combination effect of nisin and red ginger essential oil (Zingiber officinale var. rubrum) against foodborne pathogens and food spoilage microorganisms. AIP Conf. Proc. 2018, 2014, 020023. [Google Scholar] [CrossRef]
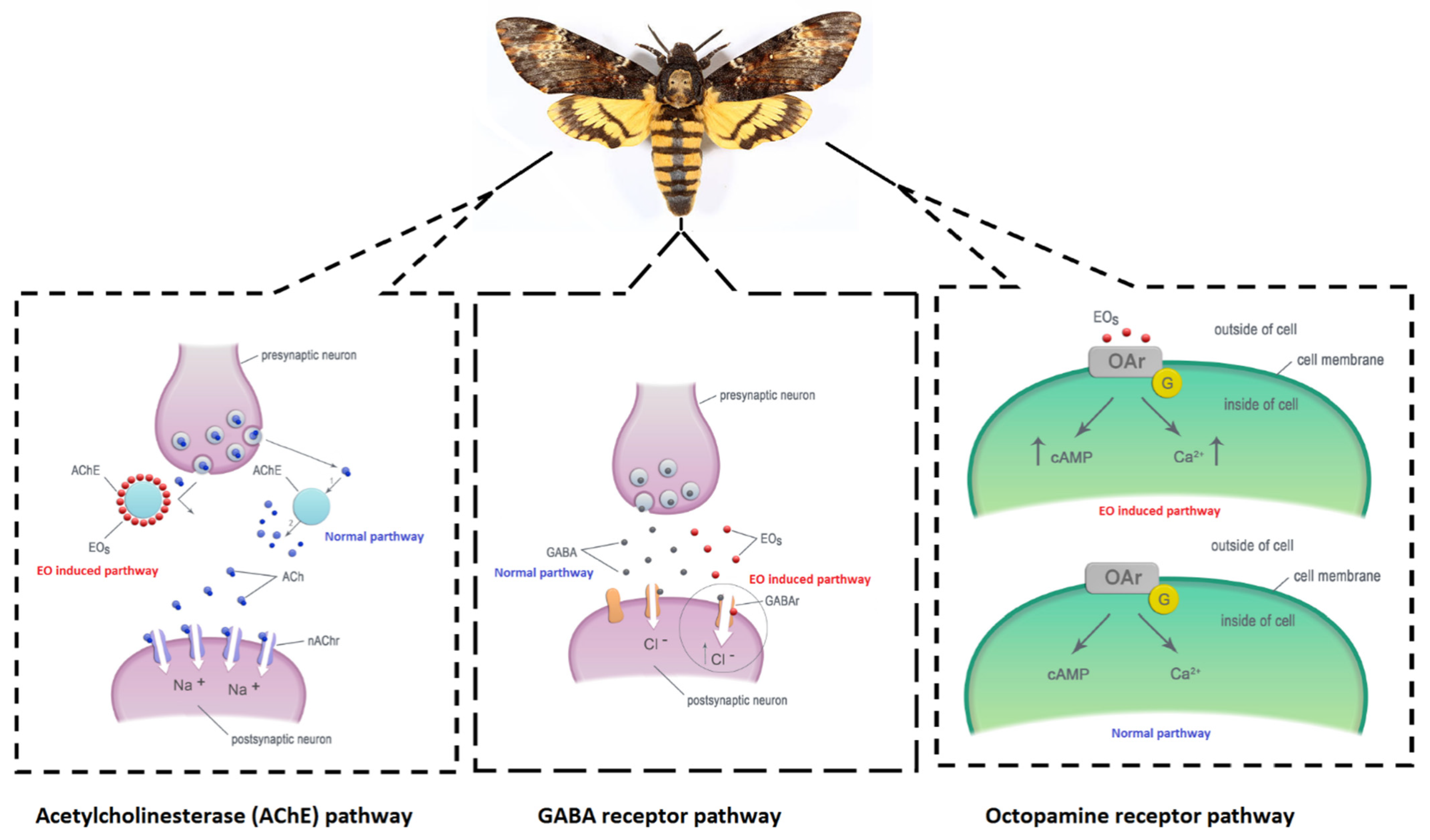
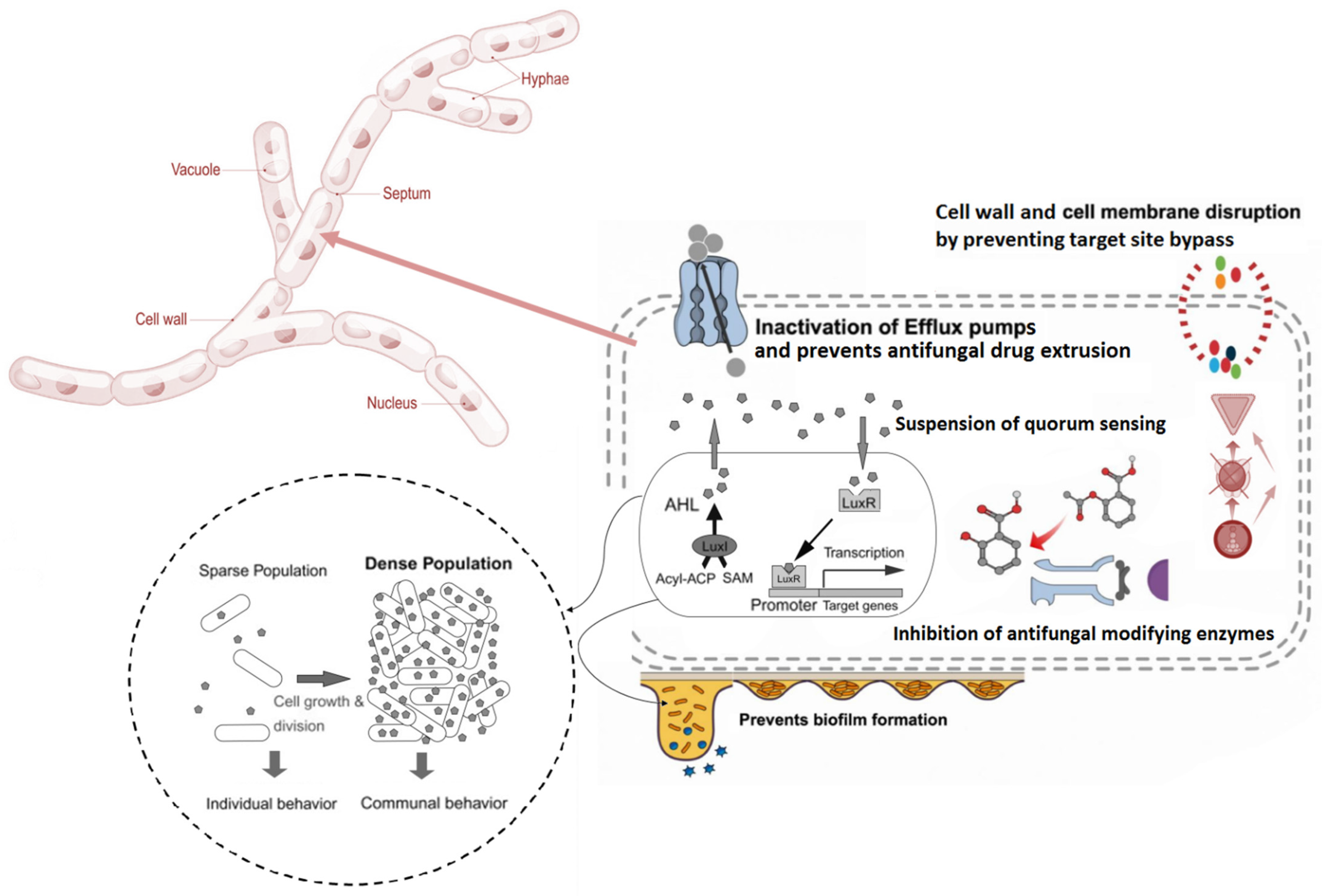
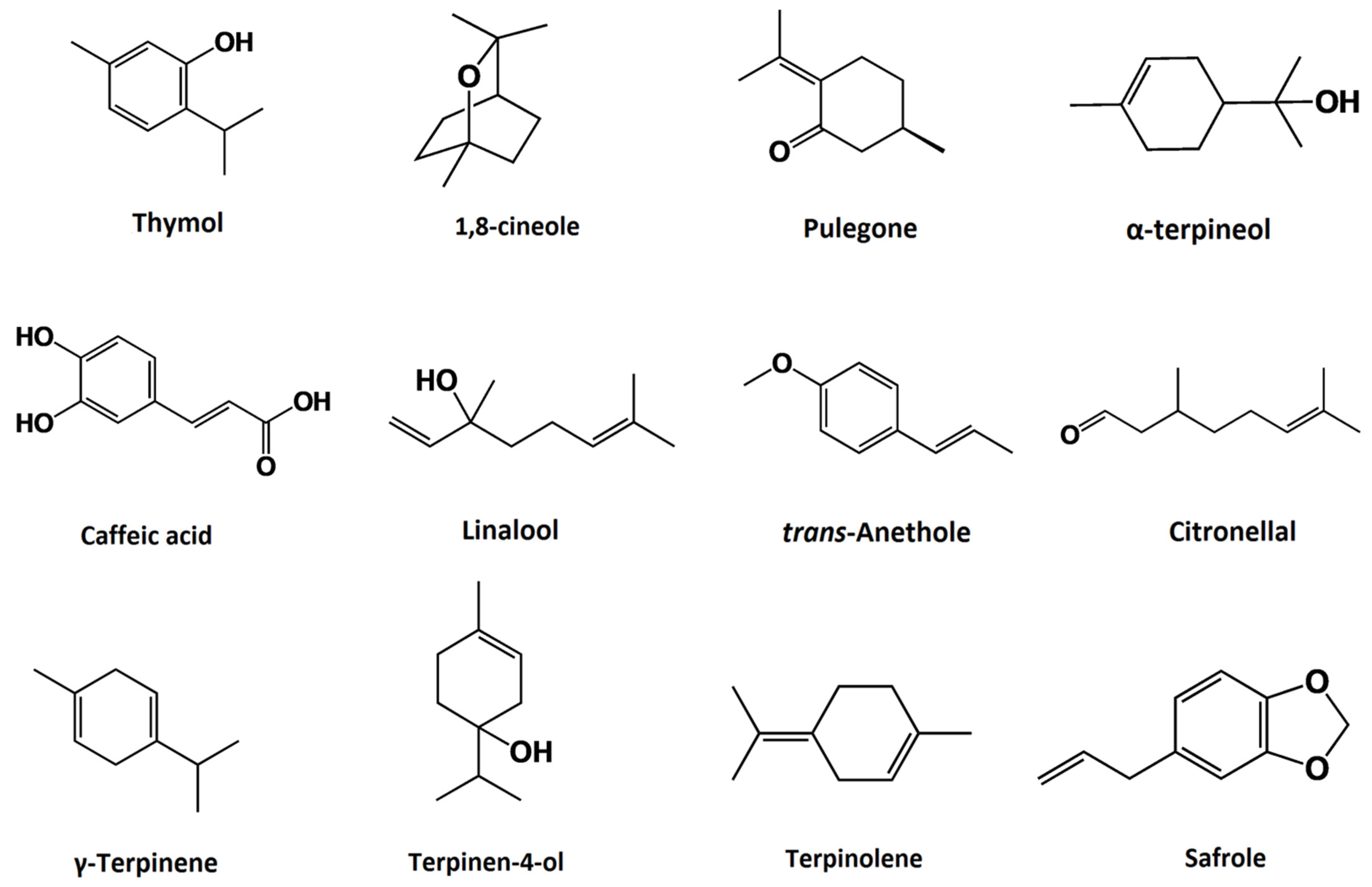
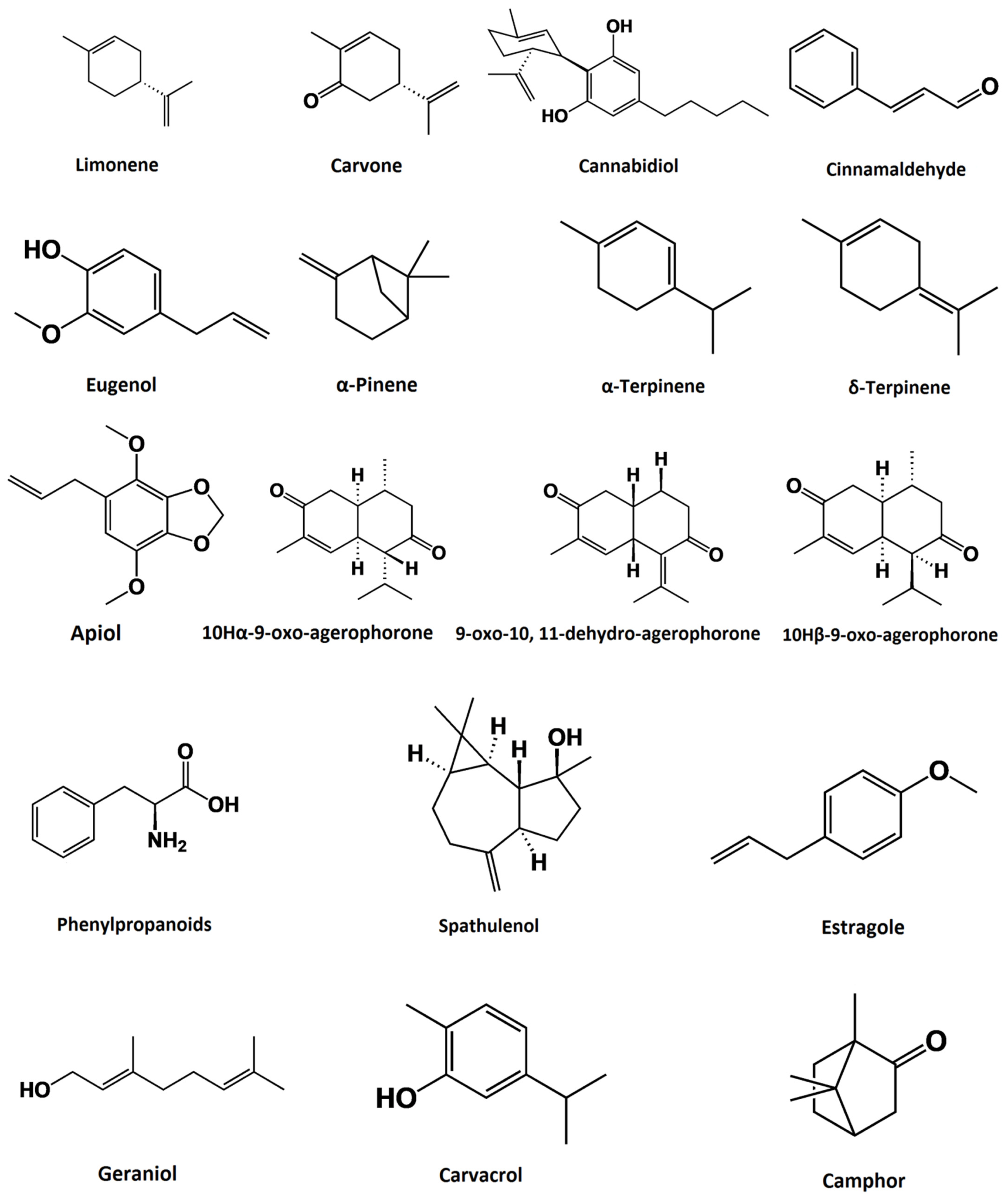

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Source | EO Compound | Synergist Used with EO Compound | Insect Pest//Fungal Pathogen | Reference |
|---|---|---|---|---|
| N/S | Cinnamaldehyde | Eugenol | L. sulphureus | [112] |
| N/S | Cinnamaldehyde | Eugenol | C. nardus | [113] |
| Melaleuca alternifolia | Terpinolene, Terpinen-4-ol, δ-Terpinene, | α-pinene, 1,8-cineole, α-terpineol | B. cinerea | [115] |
| Anathallis graveolens | Carvone, Apiol | Limonene | A. flavus | [116] |
| Cymbopogon nardus | Linalool | Citronellal | Aspergillus sp., Eurotium sp., Penicillium sp. | [117] |
| Tagetes minuta, Laurus nobilis, Tagetes filifolia | Anethole, Phenylpropanoids, Sphatulenol, Estragole | Iprodione | S. cepivorum | [121] |
| N/S | Carvacrol | Crystalline proteins of B. thuringiensis | C. pomonella, S. exigua | [123] |
| Azadirachta indica Rosmarinus officinalis | Azadirachtin Camphor | Crystalline toxins of B. thuringiensis 1,8-cineole | S. exigua, C. pomonella, D. pini T. ni | [124] [91] |
| N/S | α-terpineol | Thymol | C. partellus | [93] |
| N/S | Trans-anethole, Citronellal | α-terpineol, and thymol | S. litura | [94] |
| N/S | Cinnamon oil | Rotenone | S. litura | [96] |
| N/S | Terpinolene | Safrole | L. decemlineata, S. littoralis, R. padi, M. persicae | [98] |
| Piper hispidinervum | γ--terpinene | Terpinen-4-ol | A. fabae | [97] |
| S. littoralis | ||||
| Majorana hortensis | Cinnamaldehyde | Fluconazole | A. fumigatus | [118] |
| N/S N/S Simmodsia chinesis | 10Hα-9-oxo-agerophorone, 9-oxo-10, 11-dehydro-agerophorone, 10Hβ-9-oxo-agerophorone Jojoba oil | Mefenoxam, Mancozeb Cyhalothrin | P. myriotylum B. tabaci | [120] [100] |
| Allium sativum | Garlic oil | Diuron | ||
| Mentha piperita | Peppermint oil | Malathion | ||
| Thymus vulgaris | N/S | Imidacloprid, Spirotetramat | M. persicae | [101] |
| Lavandula angustifolia | N/S | Deltamethrin | S. frugiperda | [103] |
| N/S | Linalool, Thymol | Decis® (25CE) | S. Littoralis | [104] |
| Ocimum basilicum | Linalool | Chlorpyrifos, Cypermethrin | S. zeamais | [105] |
| Madex, Azatin, Helicovex | R. dominica | [106] | ||
| N/S | Garlic oil | P. truncatus | ||
| N/S | Cannabidiol oil | T. granarium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dassanayake, M.K.; Chong, C.H.; Khoo, T.-J.; Figiel, A.; Szumny, A.; Choo, C.M. Synergistic Field Crop Pest Management Properties of Plant-Derived Essential Oils in Combination with Synthetic Pesticides and Bioactive Molecules: A Review. Foods 2021, 10, 2016. https://doi.org/10.3390/foods10092016
Dassanayake MK, Chong CH, Khoo T-J, Figiel A, Szumny A, Choo CM. Synergistic Field Crop Pest Management Properties of Plant-Derived Essential Oils in Combination with Synthetic Pesticides and Bioactive Molecules: A Review. Foods. 2021; 10(9):2016. https://doi.org/10.3390/foods10092016
Chicago/Turabian StyleDassanayake, Mackingsley Kushan, Chien Hwa Chong, Teng-Jin Khoo, Adam Figiel, Antoni Szumny, and Chee Ming Choo. 2021. "Synergistic Field Crop Pest Management Properties of Plant-Derived Essential Oils in Combination with Synthetic Pesticides and Bioactive Molecules: A Review" Foods 10, no. 9: 2016. https://doi.org/10.3390/foods10092016