
Dynamic Modeling of Carnobacterium maltaromaticum CNCM I-3298 Growth and Metabolite Production and Model-Based Process Optimization

 and
and 
Abstract
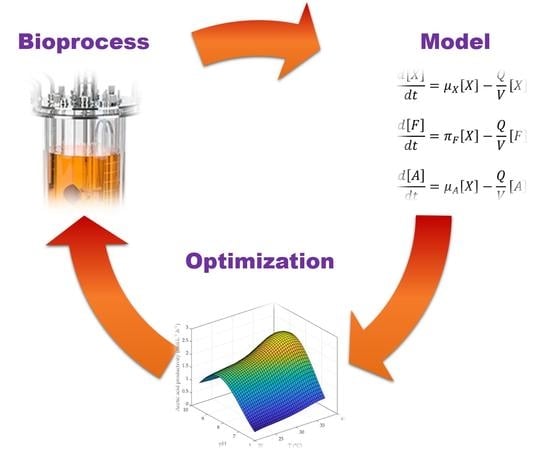
1. Introduction
2. Materials and Methods
2.1. Fermentation
2.1.1. Culture Medium and Bacterial Strain
2.1.2. Bioreactor and Parameter Control
2.2. Kinetic Measurements
2.2.1. Cell Growth
2.2.2. Total Acid Production
2.2.3. Substrate Consumption and Metabolite Production
3. Dynamic Model
3.1. Model Formulation
3.2. Model Parameter Identification
3.3. Response Surface Model for Parameter Dependence on Fermentation Conditions
4. Results and Discussion
4.1. Model Parameter Identification
4.2. Response Surface Model for Parameter Dependence on Fermentation Conditions
4.3. Model Validation
4.4. Model-Based Optimization of Fermentation Operating Conditions for Industrial Use
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
| A | Acetic acid |
| E | Ethanol |
| F | Formic acid |
| L | Lactic acid |
| S | Carbon substrate (trehalose) |
| X | Biomass |
| EA | Enzymatic activation factor |
| C (mol·L−1) | Concentration (in the calculation of errors and the definition of the inhibition factors) |
| [i] (mol·L−1) | Concentration of species i (substrate, metabolite, biomass) in the culture medium (in the system of differential equations) |
| Im | Production inhibition factor of metabolite m |
| IX | Biomass growth inhibition factor |
| KIm (mol·L−1) | Concentration for 50% production rate inhibition of metabolite m |
| KIX (molC·L−1) | Concentration for 50% growth rate inhibition of biomass |
| KSm (mol·L−1) | Concentration of production rate saturation of metabolite m |
| KSX (molC·L−1) | Concentration of biomass growth rate saturation |
| molC | Carbon-mol of biomass |
| n | Shape factor of the growth inhibition function |
| p | Shape factor of the production inhibition function |
| pH | Potential of hydrogen |
| Q (L·h−1) | Rate of base addition for pH control |
| R | Shape factor of the enzymatic activation function |
| T (K) | Temperature |
| TTI | Time–temperature indicator |
| RMSE | Root-mean square error |
| RME | Relative mean error |
| SE | Standard error |
| t (h) | Time |
| tlag (h) | Lag time |
| V (L) | Culture medium volume |
| Yi/S (mol·mol−1) | Yield of product i on substrate S |
| YX/S (molC·mol−1) | Biomass yield on substrate S |
| μX (h−1) | Specific growth rate |
| μmax,X (h−1) | Maximal specific growth rate |
| πm (h−1) | Specific production rate of metabolite m |
| πmax,m (h−1) | Maximum specific production rate of metabolite m |
Appendix A
Appendix A.1. Model Parameter Identification

| RMSE (Mol L−1) | RME (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X | S | F | A | L | E | A Total | X | S | F | A | L | E | A Total | |
| F01 | 0.008 | 0.006 | 0.010 | 0.007 | 0.022 | 0.007 | 0.033 | 4 | 4 | 7 | 13 | 6 | 10 | 7 |
| F02 | 0.005 | 0.007 | 0.006 | 0.003 | 0.004 | 0.003 | 0.005 | 4 | 16 | 8 | 8 | 8 | 7 | 3 |
| F03 | 0.005 | 0.006 | 0.003 | 0.001 | 0.003 | 0.001 | 0.006 | 4 | 9 | 3 | 2 | 3 | 2 | 1 |
| F04 | 0.006 | 0.006 | 0.003 | 0.001 | 0.005 | 0.002 | 0.005 | 7 | 13 | 5 | 5 | 6 | 6 | 3 |
| F05 | 0.008 | 0.005 | 0.014 | 0.007 | 0.027 | 0.007 | 0.034 | 4 | 4 | 9 | 11 | 10 | 9 | 8 |
| F06 | 0.006 | 0.004 | 0.012 | 0.005 | 0.024 | 0.006 | 0.012 | 5 | 2 | 7 | 7 | 8 | 8 | 3 |
| F07 | 0.006 | 0.007 | 0.011 | 0.006 | 0.009 | 0.006 | 0.007 | 4 | 5 | 14 | 33 | 4 | 11 | 2 |
| F08 | 0.006 | 0.007 | 0.011 | 0.005 | 0.030 | 0.004 | 0.003 | 4 | 5 | 17 | 23 | 8 | 7 | 1 |
| F09 | 0.004 | 0.005 | 0.002 | 0.001 | 0.002 | 0.002 | 0.003 | 7 | 17 | 6 | 7 | 5 | 6 | 2 |
| F10 | 0.005 | 0.009 | 0.006 | 0.004 | 0.028 | 0.004 | 0.026 | 3 | 7 | 4 | 7 | 9 | 5 | 6 |
| F11 | 0.003 | 0.018 | 0.009 | 0.005 | 0.011 | 0.006 | 0.032 | 3 | 27 | 9 | 10 | 9 | 11 | 6 |
| F12 | 0.005 | 0.013 | 0.002 | 0.001 | 0.003 | 0.001 | 0.004 | 13 | 61 | 3 | 4 | 8 | 2 | 2 |
| F13 | 0.002 | 0.006 | 0.012 | 0.005 | 0.024 | 0.005 | 0.007 | 2 | 4 | 7 | 6 | 6 | 5 | 2 |
| F14 | 0.003 | 0.003 | 0.009 | 0.005 | 0.012 | 0.006 | 0.015 | 4 | 3 | 16 | 20 | 5 | 12 | 4 |
| F15 | 0.009 | 0.005 | 0.009 | 0.006 | 0.023 | 0.005 | 0.011 | 5 | 4 | 7 | 13 | 8 | 8 | 2 |
| F16 | 0.003 | 0.001 | 0.001 | 0.001 | 0.008 | 0.001 | 0.014 | 2 | 1 | 2 | 4 | 6 | 2 | 3 |
| Mean | 0.005 | 0.007 | 0.008 | 0.004 | 0.015 | 0.004 | 0.014 | 5 | 11 | 8 | 11 | 7 | 7 | 4 |
| Variable | Coefficient | µmaxX (h−1) | πmaxF (h−1) | πmaxA (h−1) | πmaxL (h−1) | πmaxE (h−1) | KIX (Mol L−1) | KIm (Mol L−1) |
|---|---|---|---|---|---|---|---|---|
| Constant | Value (β0) | −1.38 × 101 | −1.64 × 101 | −1.74 × 101 | −1.21 × 101 | −1.12 × 101 | −8.61 × 10−1 | 1.19 × 101 |
| Standard error | 0.05 × 101 | 0.07 × 101 | 0.09 × 101 | 0.09 × 101 | 0.07 × 101 | 0.05 × 10−1 | 0.03 × 101 | |
| T | Value (β1) | 2.63 × 10−1 | 2.66 × 10−1 | 2.71 × 10−1 | 1.82 × 10−1 | 1.27 × 10−1 | −1.87 × 10−1 | −5.80 × 10−1 |
| Standard error | 0.13 × 10−1 | 0.26 × 10−1 | 0.28 × 10−1 | 0.23 × 10−1 | 0.14 × 10−1 | 0.05 × 10−1 | 0.17 × 10−1 | |
| pH | Value (β2) | 2.42 × 100 | 2.89 × 100 | 3.01 × 100 | 2.27 × 100 | 2.02 × 100 | 6.65 × 10−1 | −1.27 × 100 |
| Standard error | 0.11 × 100 | 0.14 × 100 | 0.19 × 100 | 0.16 × 100 | 0.13 × 100 | 0.45 × 10−1 | 0.16 × 100 | |
| T² | Value (β3) | −4.26 × 10−3 | −3.45 × 10−3 | −3.26 × 10−3 | −2.05 × 10−3 | −1.92 × 10−3 | 3.97 × 10−3 | 7.95 × 10−3 |
| Standard error | 0.13 × 10−3 | 0.32 × 10−3 | 0.33 × 10−3 | 0.81 × 10−3 | 0.22 × 10−3 | 0.09 × 10−3 | 0.32 × 10−3 | |
| pH² | Value (β4) | −1.46 × 10−1 | −1.63 × 10−1 | −1.66 × 10−1 | −1.34 × 10−1 | −1.22 × 10−1 | −4.03 × 10−2 | 4.35 × 10−2 |
| Standard error | 0.06 × 10−1 | 0.10 × 10−1 | 0.12 × 10−1 | 0.10 × 10−1 | 0.09 × 10−1 | 0.31 × 10−2 | 0.55 × 10−2 | |
| T·pH | Value (β5) | −3.53 × 10−3 | −7.58 × 10−3 | −9.17 × 10−3 | −7.22 × 10−3 | −4.68 × 10−3 | −3.54 × 10−3 | 1.86 × 10−2 |
| Standard error | 0.11 × 10−3 | 0.97 × 10−3 | 1.17 × 10−3 | 0.91 × 10−3 | 0.05 × 10−3 | 0.06 × 10−3 | 0.01 × 10−2 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Appendix A.2. Model Validation
| Fermentation | RMSE (Mol L−1) | RME (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X | S | F | A | L | E | At | X | S | F | A | L | E | At | ||
| Calibration | F01 | 0.010 | 0.010 | 0.021 | 0.010 | 0.041 | 0.014 | 0.033 | 5 | 6 | 14 | 20 | 10 | 18 | 7 |
| F03 | 0.021 | 0.009 | 0.008 | 0.005 | 0.008 | 0.003 | 0.012 | 15 | 14 | 7 | 9 | 7 | 5 | 4 | |
| F04 | 0.005 | 0.010 | 0.009 | 0.006 | 0.007 | 0.005 | 0.015 | 5 | 26 | 18 | 23 | 9 | 17 | 9 | |
| F07 | 0.012 | 0.009 | 0.010 | 0.006 | 0.015 | 0.006 | 0.009 | 9 | 8 | 13 | 31 | 5 | 10 | 3 | |
| F12 | 0.003 | 0.011 | 0.008 | 0.005 | 0.003 | 0.004 | 0.009 | 8 | 53 | 13 | 16 | 8 | 11 | 6 | |
| F13 | 0.010 | 0.007 | 0.006 | 0.003 | 0.036 | 0.003 | 0.011 | 7 | 5 | 4 | 4 | 8 | 4 | 2 | |
| F14 | 0.015 | 0.002 | 0.013 | 0.007 | 0.004 | 0.009 | 0.012 | 20 | 2 | 23 | 28 | 2 | 16 | 3 | |
| F16 | 0.011 | 0.023 | 0.033 | 0.014 | 0.064 | 0.018 | 0.085 | 9 | 33 | 50 | 45 | 48 | 38 | 18 | |
| Mean | 0.011 | 0.010 | 0.014 | 0.007 | 0.022 | 0.008 | 0.023 | 10 | 18 | 18 | 22 | 12 | 15 | 7 | |
| Validation | F02 | 0.005 | 0.003 | 0.017 | 0.009 | 0.008 | 0.009 | 0.007 | 5 | 8 | 20 | 22 | 15 | 21 | 4 |
| F05 | 0.011 | 0.008 | 0.007 | 0.006 | 0.045 | 0.005 | 0.016 | 7 | 7 | 4 | 8 | 13 | 8 | 4 | |
| F06 | 0.008 | 0.009 | 0.013 | 0.005 | 0.044 | 0.007 | 0.032 | 6 | 5 | 7 | 6 | 11 | 9 | 6 | |
| F08 | 0.009 | 0.010 | 0.012 | 0.006 | 0.026 | 0.008 | 0.003 | 7 | 8 | 19 | 25 | 8 | 14 | 1 | |
| F09 | 0.011 | 0.006 | 0.005 | 0.004 | 0.018 | 0.004 | 0.009 | 19 | 22 | 15 | 24 | 35 | 15 | 7 | |
| F10 | 0.021 | 0.014 | 0.025 | 0.011 | 0.063 | 0.014 | 0.044 | 11 | 10 | 17 | 18 | 20 | 20 | 10 | |
| F11 | 0.018 | 0.013 | 0.007 | 0.003 | 0.017 | 0.004 | 0.050 | 16 | 20 | 7 | 6 | 12 | 8 | 9 | |
| F15 | 0.028 | 0.023 | 0.026 | 0.010 | 0.047 | 0.014 | 0.036 | 15 | 20 | 22 | 21 | 17 | 21 | 7 | |
| Mean | 0.014 | 0.011 | 0.014 | 0.007 | 0.033 | 0.008 | 0.025 | 11 | 13 | 14 | 16 | 16 | 15 | 6 | |


References
- Edima, H.; Cailliez-Grimal, C.; Revol-Junelles, A.-M.; Tonti, L.; Linder, M.; Millière, J.B. A selective enumeration medium for Carnobacterium maltaromaticum. J. Microbiol. Methods 2007, 68, 516–521. [Google Scholar] [CrossRef]
- Afzal, M.; Jacquet, T.; Delaunay, S.; Borges, F.; Millière, J.B.; Revol-Junelles, A.M.; Cailliez-Grimal, C. Carnobacterium maltaromaticum: Identification, isolation tools, ecology and technological aspects in dairy products. Food Microbiol. 2010, 27, 573–579. [Google Scholar] [CrossRef]
- Cailliez-Grimal, C.; Edima, H.; Revol-Junelles, A.-M.; Millière, J.-B. Carnobacterium maltaromaticum: The Only Carnobacterium Species in French Ripened Soft Cheeses as Revealed by Polymerase Chain Reaction Detection. J. Dairy Sci. 2007, 90, 1133–1138. [Google Scholar] [CrossRef]
- Edima, H.; Cailliez-Grimal, C.; Revol-Junelles, A.M.; Rondags, E.; Millière, J.B. Impact of pH and Temperature on the Acidifying Activity of Carnobacterium Maltaromaticum. J. Dairy Sci. 2008, 91, 3806–3813. [Google Scholar] [CrossRef]
- Taoukis, P.; Labuza, T. Applicability of Time-Temperature Indicators as Shelf Life Monitors of Food Products. J. Food Sci. 1989, 54, 783–788. [Google Scholar] [CrossRef]
- Ellouze, M.; Pichaud, M.; Bonaiti, C.; Coroller, L.; Couvert, O.; Thuault, D.; Vaillant, R. Modelling pH evolution and lactic acid production in the growth medium of a lactic acid bacterium: Application to set a biological TTI. Int. J. Food Microbiol. 2008, 128, 101–107. [Google Scholar] [CrossRef]
- Girardeau, A.; Puentes, C.; Keravec, S.; Peteuil, P.; Trelea, I.C.; Fonseca, F. Influence of culture conditions on the technological properties of Carnobacterium maltaromaticum CNCM I-3298 starters. J. Appl. Microbiol. 2019, 126, 1468–1479. [Google Scholar] [PubMed]
- Borch, E.; Molin, G. The aerobic growth and product formation of Lactobacillus, Leuconostoc, Brochothrix, and Carnobacterium in batch cultures. Appl. Microbiol. Biotechnol. 1989, 30, 81–88. [Google Scholar] [CrossRef]
- dos Reis, F.B.; de Souza, V.M.; Thomaz, M.R.S.; Fernandes, L.P.; de Oliveira, W.P.; de Martinis, E.C.P. Use of Carnobacterium maltaromaticum cultures and hydroalcoholic extract of Lippia sidoides Cham. Against Listeria monocytogenes in fish model systems. Int. J. Food Microbiol. 2011, 146, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.I.; Delaunay, S.; Paris, C.; Borges, F.; Revol-Junelles, A.M.; Cailliez-Grimal, C. Identification of metabolic pathways involved in the biosynthesis of flavor compound 3-methylbutanal from leucine catabolism by Carnobacterium maltaromaticum LMA 28. Int. J. Food Microbiol. 2012, 157, 332–339. [Google Scholar] [CrossRef]
- Afzal, M.I.; Boulahya, K.A.; Paris, C.; Delaunay, S.; Cailliez-Grimal, C. Effect of oxygen on the biosynthesis of flavor compound 3-methylbutanal from leucine catabolism during batch culture in Carnobacterium maltaromaticum LMA 28. J. Dairy Sci. 2013, 96, 352–359. [Google Scholar] [CrossRef]
- Rahman, A.; Gleinser, M.; Lanhers, M.-C.; Riedel, C.U.; Foligne, B.; Hanse, M.; Yen, F.T.; Klouj, A.; Afzal, M.I.; Back, A.; et al. Adaptation of the lactic acid bacterium Carnobacterium maltaromaticum LMA 28 to the mammalian gastrointestinal tract: From survival in mice to interactions with human cells. Int. Dairy J. 2014, 34, 93–99. [Google Scholar]
- Rahman, A.; El Kheir, S.M.; Back, A.; Mangavel, C.; Revoljunelles, A.-M.; Borges, F. Repeat-based bequence typing of Carnobacterium maltaromaticum. Int. J. Food Microbiol. 2016, 226, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Badoni, M.; Gänzle, M.; Yang, X. Growth of Carnobacterium spp. isolated from chilled vacuum-packaged meat under relevant acidic conditions. Int. J. Food Microbiol. 2018, 286, 120–127. [Google Scholar]
- De Bruyn, I.N.; Louw, A.I.; Visser, L.; Holzapfel, W.H. Lactobacillus divergens is a homofermentative organism. Syst. Appl. Microbiol. 1987, 9, 173–175. [Google Scholar] [CrossRef]
- De Bruyn, I.N.; Holzapfel, W.H.; Visser, L.; Louw, A.I. Glucose metabolism by Lactobacillus divergens. J. Gen. Microbiol. 1988, 134, 2103–2109. [Google Scholar] [CrossRef]
- Leisner, J.J.; Laursen, B.G.; Prévost, H.; Drider, D.; Dalgaard, P. Carnobacterium: Positive and negative effects in the environment and in foods. FEMS Microbiol. Rev. 2007, 31, 592–613. [Google Scholar] [CrossRef]
- Schillinger, U.; Holzapfel, W.H. The genus Carnobacterium. In The Genera of Lactic Acid Bacteria. The Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer: Boston, MA, USA, 1995; Volume 2, pp. 307–326. [Google Scholar]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Ward, B. Bacterial energy metabolism. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.W., Liu, D., Schwartzman, J., Sussman, M., Poxton, I., Eds.; Elsevier: Waltham, MA, USA, 2015; pp. 201–233. [Google Scholar]
- Laursen, B.G.; Leisner, J.J.; Dalgaard, P. Carnobacterium Species: Effect of Metabolic Activity and Interaction with Brochothrix thermosphacta on Sensory Characteristics of Modified Atmosphere Packed Shrimp. J. Agric. Food Chem. 2006, 54, 3604–3611. [Google Scholar] [CrossRef]
- Zhang, P.; Gänzle, M.; Yang, X. Complementary Antibacterial Effects of Bacteriocins and Organic Acids as Revealed by Comparative Analysis of Carnobacterium spp. from Meat. Appl. Environ. Microbiol. 2019, 85, e01227-19. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Rådström, P. Beta-Glucose 1-phosphate-interconverting enzymes in maltose- and trehalose-fermenting lactic acid bacteria. Environ. Microbiol. 2002, 4, 81–88. [Google Scholar] [CrossRef]
- Levisauskas, D.; Galvanauskas, V.; Henrich, S.; Wilhelm, K.; Volk, N.; Lübbert, A. Model-based optimization of viral capsid protein production in fed-batch culture of recombinant Escherichia coli. Bioprocess. Biosyst. Eng. 2003, 25, 255–262. [Google Scholar] [CrossRef]
- Antelo, L.; Passot, S.; Fonseca, F.; Trelea, I.C.; Alonso, A.A. Towards optimal operation conditions of freeze-drying processes via a multi-level approach. Dry. Technol. 2012, 30, 1432–1448. [Google Scholar] [CrossRef]
- Kiparissides, A.; Pistikopoulos, E.N.; Mantalaris, A. On the model-based optimization of secreting mammalian cell (GS-NS0) cultures. Biotechnol. Bioeng. 2015, 112, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Balsa-Canto, E.; Alonso, A.A.; Arias-Méndez, A.; Garcia, M.; López-Núñez, A.; Mosquera-Fernández, M.; Vázquez, C.; Vilas, C. Modeling and optimization techniques with applications in food processes, bio-processes and bio-systems. In Numerical Simulation in Physics and Engineering; Higueras, I., Roldán, T., Torrens, J., Eds.; Springer: Cham, Switzerland, 2016; Volume 9, pp. 187–216. [Google Scholar]
- Sanchez-Castaneda, A.; Athès, V.; Moussa, M.; Lopez-Miranda, J.; Paez-Lerma, J.; Soto-Cruz, N.; Trelea, I.-C. Modeling of isoamyl acetate production by fermentation with Pichia fermentans in an aerated system coupled to in situ extraction. Process. Biochem. 2018, 65, 11–20. [Google Scholar] [CrossRef]
- Sinner, P.; Kager, J.; Daume, S.; Herwig, C. Model-based analysis and optimisation of a continuous Corynebacterium glutamicum Bioprocess Utilizing Lignocellulosic Waste. IFAC-Pap. 2019, 52, 181–186. [Google Scholar] [CrossRef]
- Battley, E.H. Growth-Reaction Equations for Saccharomyces cerevisiae. Physiol. Plant. 1960, 13, 192–203. [Google Scholar] [CrossRef]
- Bastin, G.; Dochain, D. On-Line Estimation and Adaptive Control of Bioreactors; Elsevier: Amsterdam, The Netherlands, 1990; pp. 1–82. [Google Scholar]
- Bailey, J.E. Mathematical modeling and analysis in biochemical engineering: Past accomplishments and future opportunities. Biotechnol. Prog. 1998, 14, 8–20. [Google Scholar] [CrossRef]
- Monod, J. Recherches sur la Croissance des Cultures Bactériennes; Hermann & Cie: Paris, France, 1942. [Google Scholar]
- Levenspiel, O. The monod equation: A revisit and a generalization to product inhibition situations. Biotechnol. Bioeng. 1980, 22, 1671–1687. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Aiba, S.; Shoda, M. Reassessment of the product inhibition in alcohol fermentation. J. Ferment. Technol. 1969, 47, 790–803. [Google Scholar]
- Claret, C.; Bories, A.; Soucaille, P. Inhibitory effect of dihydroxyacetone on Gluconobacter oxydans: Kinetic aspects and expression by mathematical equations. J. Ind. Microbiol. 1993, 11, 105–112. [Google Scholar] [CrossRef]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef]
- Baranyi, J. Comparison of Stochastic and Deterministic Concepts of Bacterial Lag. J. Theor. Biol. 1998, 192, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, M.; Rice, C.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.; Alston, M.; Michael, F.; Stringer, M.; Betts, R.; et al. Lag Phase Is a Distinct Growth Phase that Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. J. Bacteriol. 2012, 194, 686–701. [Google Scholar] [CrossRef]
- Bouville, M. Fermentation kinetics including product and substrate inhibitions plus biomass death: A mathematical analysis. Biotechnol. Lett. 2007, 29, 737–741. [Google Scholar] [CrossRef][Green Version]
- Seber, G.; Wild, C. Nonlinear Regression, 2nd ed.; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Owens, J.D.; Legan, J.D. Determination of the Monod substrate saturation constant for microbial growth. FEMS Microbiol. Rev. 1987, 3, 419–432. [Google Scholar] [CrossRef]
- Brock, T.D.; Madigan, M.T. Biology of Microorganisms, 6th ed.; Prentice Hall: London, UK, 1991; p. 353. [Google Scholar]
- Rodríguez, J.; Lema, J.M.; Kleerebezem, R. Energy-based models for environmental biotechnology. Trends Biotechnol. 2008, 26, 366–374. [Google Scholar] [CrossRef]
- Bellgardt, K.H. Bioprocess Models. In Bioreaction Engineering: Modeling and Control; Schügerl, K., Bellgardt, K.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 44–105. [Google Scholar]
- González-Cabaleiro, R.; Lema, J.M.; Rodríguez, J. Metabolic Energy-Based Modelling Explains Product Yielding in Anaerobic Mixed Culture Fermentations. PLoS ONE 2015, 10, e0126739. [Google Scholar] [CrossRef]







| Parameter | Constant Value |
|---|---|
| YX/S (molC.mol−1) | 6.9 |
| YF/S (mol.mol−1) | 5.6 |
| YA/S (mol.mol−1) | 3.8 |
| YL/S (mol.mol−1) | 7.0 |
| YE/S (mol.mol−1) | 4.7 |
| KSX (mol L−1) | 0.001 |
| KSm (mol L−1) | 0.001 |
| n | 3 |
| p | 1 |
| r (h−1) | 0.8 |
| Fermentation | µmaxX (h−1) | πmaxF (h−1) | πmaxA (h−1) | πmaxL (h−1) | πmaxE (h−1) | KIX (Mol.L−1) | KIm (Mol.L−1) | |
|---|---|---|---|---|---|---|---|---|
| F01 | Value | 0.224 | 0.152 | 0.064 | 0.215 | 0.078 | 0.117 | 0.069 |
| Standard error | 0.003 | 0.003 | 0.003 | 0.005 | 0.001 | 0.004 | 0.003 | |
| F02 | Value | 0.096 | 0.122 | 0.064 | 0.053 | 0.064 | 0.143 | 0.142 |
| Standard error | 0.021 | 0.012 | 0.007 | 0.006 | 0.006 | 0.083 | 0.036 | |
| F03 | Value | 0.164 | 0.137 | 0.064 | 0.131 | 0.072 | 0.099 | 0.102 |
| Standard error | 0.012 | 0.009 | 0.004 | 0.009 | 0.005 | 0.006 | 0.013 | |
| F04 | Value | 0.078 | 0.063 | 0.031 | 0.072 | 0.034 | 0.144 | 0.092 |
| Standard error | 0.012 | 0.004 | 0.002 | 0.004 | 0.002 | 0.047 | 0.016 | |
| F05 | Value | 0.130 | 0.130 | 0.060 | 0.196 | 0.070 | 0.160 | 0.100 |
| Standard error | 0.005 | 0.002 | 0.002 | 0.004 | 0.001 | 0.004 | 0.004 | |
| F06 | Value | 0.094 | 0.129 | 0.063 | 0.159 | 0.063 | 0.163 | 0.092 |
| Standard error | 0.001 | 0.002 | 0.001 | 0.003 | 0.001 | 0.003 | 0.002 | |
| F07 | Value | 0.074 | 0.028 | 0.010 | 0.089 | 0.022 | 0.122 | 0.139 |
| Standard error | 0.002 | 0.001 | 0.001 | 0.004 | 0.002 | 0.005 | 0.004 | |
| F08 | Value | 0.060 | 0.055 | 0.024 | 0.126 | 0.048 | 0.149 | 0.060 |
| Standard error | 0.002 | 0.002 | 0.000 | 0.003 | 0.002 | 0.001 | 0.002 | |
| F09 | Value | 0.071 | 0.097 | 0.048 | 0.114 | 0.071 | 0.108 | 0.044 |
| Standard error | 0.009 | 0.007 | 0.004 | 0.010 | 0.006 | 0.021 | 0.007 | |
| F10 | Value | 0.220 | 0.230 | 0.100 | 0.300 | 0.120 | 0.193 | 0.091 |
| Standard error | 0.051 | 0.013 | 0.007 | 0.022 | 0.007 | 0.044 | 0.015 | |
| F11 | Value | 0.121 | 0.127 | 0.060 | 0.116 | 0.066 | 0.179 | 0.133 |
| Standard error | 0.004 | 0.007 | 0.004 | 0.005 | 0.003 | 0.006 | 0.003 | |
| F12 | Value | 0.132 | 0.155 | 0.082 | 0.076 | 0.086 | 0.043 | 0.059 |
| Standard error | 0.010 | 0.026 | 0.003 | 0.002 | 0.006 | 0.002 | 0.007 | |
| F13 | Value | 0.147 | 0.162 | 0.077 | 0.219 | 0.092 | 0.164 | 0.106 |
| Standard error | 0.006 | 0.006 | 0.002 | 0.008 | 0.004 | 0.009 | 0.003 | |
| F14 | Value | 0.047 | 0.045 | 0.020 | 0.112 | 0.032 | 0.140 | 0.130 |
| Standard error | 0.005 | 0.004 | 0.001 | 0.006 | 0.003 | 0.016 | 0.007 | |
| F15 | Value | 0.160 | 0.180 | 0.080 | 0.330 | 0.110 | 0.260 | 0.170 |
| Standard error | 0.056 | 0.013 | 0.008 | 0.030 | 0.010 | 0.022 | 0.010 | |
| F16 | Value | 0.110 | 0.110 | 0.050 | 0.200 | 0.070 | 0.320 | 0.280 |
| Standard error | 0.003 | 0.004 | 0.002 | 0.007 | 0.003 | 0.051 | 0.104 | |
| A. Target | T (°C) | pH | Final Concentrations (mmol L−1) | Final Productivities (mmol L−1.h−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X | F | A | L | E | AT | X | F | A | L | E | AT | ||||
| Biomass | Bconc.↑ | 20.0 | 7.8 | 227 | 129 | 56 | 177 | 73 | 363 | 6.56 | 3.73 | 1.61 | 5.11 | 2.09 | 10.46 |
| Formic acid | Fconc.↑ | 28.0 | 9.5 | 133 | 176 | 89 | 123 | 94 | 387 | 1.68 | 2.20 | 1.12 | 1.54 | 1.18 | 4.86 |
| Acetic acid | Aconc.↑ | 28.0 | 9.5 | 133 | 176 | 89 | 123 | 94 | 387 | 1.68 | 2.20 | 1.12 | 1.54 | 1.18 | 4.86 |
| Lactic acid | Lconc.↑ | 37.0 | 6.0 | 180 | 95 | 42 | 296 | 72 | 433 | 1.68 | 2.20 | 1.12 | 1.54 | 1.18 | 4.86 |
| Ethanol | Econc.↑ | 37.0 | 9.5 | 118 | 147 | 77 | 128 | 139 | 352 | 2.35 | 2.90 | 1.54 | 2.54 | 2.76 | 6.98 |
| Ethanol | Econc.↓ | 27.0 | 7.6 | 217 | 143 | 62 | 166 | 68 | 372 | 6.93 | 4.57 | 1.98 | 5.31 | 2.16 | 11.86 |
| Total acids | ATconc.↑ | 37.0 | 6.0 | 180 | 95 | 42 | 296 | 72 | 433 | 3.33 | 1.76 | 0.77 | 5.48 | 1.33 | 8.01 |
| Total acids | ATconc.↓ | 37.0 | 9.5 | 118 | 147 | 77 | 128 | 139 | 352 | 2.35 | 2.90 | 1.54 | 2.54 | 2.76 | 6.98 |
| Biomass | Bprod.↑ | 33.5 | 7.5 | 199 | 139 | 64 | 178 | 73 | 382 | 7.49 | 5.25 | 2.41 | 6.71 | 2.77 | 14.38 |
| Formic acid | Fprod.↑ | 34.5 | 8.0 | 178 | 148 | 71 | 163 | 82 | 381 | 6.77 | 5.61 | 2.70 | 6.18 | 3.12 | 14.48 |
| Acetic acid | Aprod.↑ | 35.0 | 8.1 | 172 | 148 | 72 | 161 | 85 | 381 | 6.49 | 5.58 | 2.71 | 6.06 | 3.21 | 14.35 |
| Lactic acid | Lprod.↑ | 37.0 | 7.1 | 188 | 123 | 58 | 215 | 78 | 395 | 6.60 | 4.31 | 2.04 | 7.54 | 2.73 | 13.89 |
| Ethanol | Eprod.↑ | 37.0 | 8.3 | 158 | 144 | 73 | 162 | 97 | 378 | 5.57 | 5.08 | 2.56 | 5.70 | 3.43 | 13.34 |
| Ethanol | Eprod.↓ | 28.0 | 6.0 | 175 | 108 | 41 | 278 | 78 | 427 | 2.14 | 1.32 | 0.50 | 3.38 | 0.95 | 5.20 |
| Total acids | ATprod.↑ | 35.5 | 7.7 | 184 | 139 | 67 | 177 | 81 | 384 | 7.16 | 5.41 | 2.59 | 6.90 | 3.13 | 14.90 |
| Total acids | ATprod.↓ | 25.0 | 9.5 | 146 | 173 | 88 | 126 | 87 | 386 | 1.71 | 2.03 | 1.03 | 1.48 | 1.02 | 4.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puentes, C.; Girardeau, A.; Passot, S.; Fonseca, F.; Trelea, I.-C. Dynamic Modeling of Carnobacterium maltaromaticum CNCM I-3298 Growth and Metabolite Production and Model-Based Process Optimization. Foods 2021, 10, 1922. https://doi.org/10.3390/foods10081922
Puentes C, Girardeau A, Passot S, Fonseca F, Trelea I-C. Dynamic Modeling of Carnobacterium maltaromaticum CNCM I-3298 Growth and Metabolite Production and Model-Based Process Optimization. Foods. 2021; 10(8):1922. https://doi.org/10.3390/foods10081922
Chicago/Turabian StylePuentes, Cristian, Amélie Girardeau, Stephanie Passot, Fernanda Fonseca, and Ioan-Cristian Trelea. 2021. "Dynamic Modeling of Carnobacterium maltaromaticum CNCM I-3298 Growth and Metabolite Production and Model-Based Process Optimization" Foods 10, no. 8: 1922. https://doi.org/10.3390/foods10081922
APA StylePuentes, C., Girardeau, A., Passot, S., Fonseca, F., & Trelea, I.-C. (2021). Dynamic Modeling of Carnobacterium maltaromaticum CNCM I-3298 Growth and Metabolite Production and Model-Based Process Optimization. Foods, 10(8), 1922. https://doi.org/10.3390/foods10081922