
Microsatellite Markers in Olives (Olea europaea L.): Utility in the Cataloging of Germplasm, Food Authenticity and Traceability Studies



Abstract
1. Introduction
2. The Olive Germplasm
3. Microsatellites in Olives
3.1. Development and Available SSRs
3.2. SSR Protocols for Cultivar Genotyping
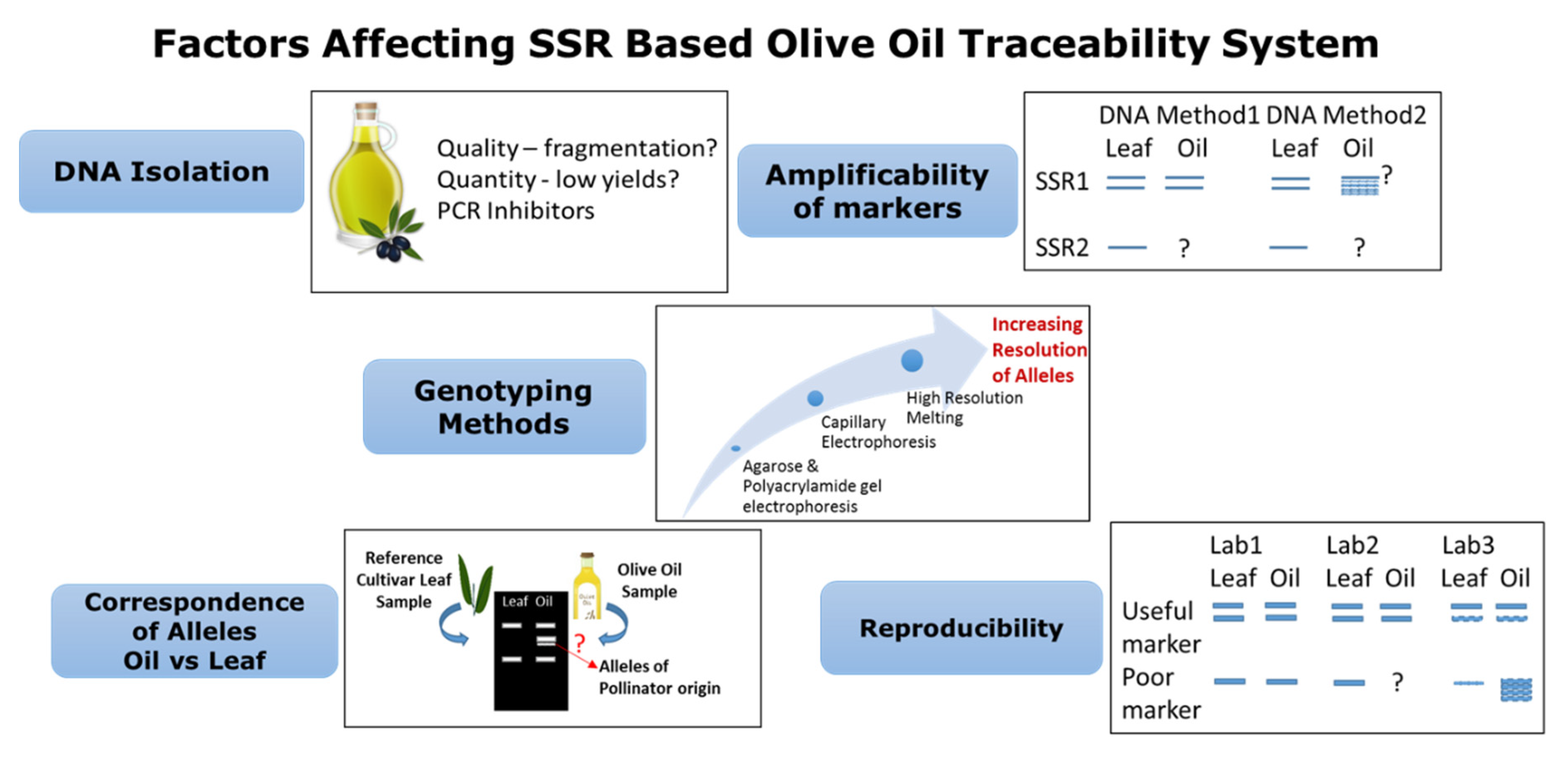
3.3. Genotyping Methods
3.4. SSR Databases
4. Applications of SSRs: Cataloging of Olive Germplasm, Food Authenticity and Traceability Studies
4.1. Cataloging Olive Germplasm
4.1.1. Cataloging of Germplasm Banks
4.1.2. Local Cultivars and Centennial Trees
4.1.3. Characterization of Wild Olive Populations
4.2. Agri-Food Traceability: Olive Oil and Table Olives
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Jiménez, F.; Ruano, J.; Perez-Martinez, P.; Lopez-Segura, F.; Lopez-Miranda, J. The influence of olive oil on human health: Not a question of fat alone. Mol. Nutr. Food Res. 2007, 51, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Cicerale, S.; Lucas, L.; Keast, R. Biological activities of phenolic compounds present in virgin olive oil. Int. J. Mol. Sci. 2010, 11, 458–479. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Hu, F.B.; Martínez-González, M.A.; Fitó, M.; Bulló, M.; Estruch, R.; Ros, E.; Corella, D.; Recondo, J.; Gómez-Gracia, E.; et al. Olive oil intake and risk of cardiovascular disease and mortality in the PREDIMED Study. BMC Med. 2014, 12, 78. [Google Scholar] [CrossRef]
- Besnard, G.; Bakkali, A.E.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef]
- Barranco, D.; Trujillo, I.; Rallo, P. Are “Oblonga” and “Frantoio” olives the same cultivar? HortScience 2000, 35, 1323–1325. [Google Scholar] [CrossRef]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian olive cultivars: Comparative analysis of molecular and phenotypic data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef]
- Bartolini, G.; Cerreti, S. Olive Germplasm (Olea europaea L.) Cultivars, Synonyms, Cultivation Area, Collections, Descriptors. Available online: www.oleadb.it/olivodb.html (accessed on 10 February 2021).
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Gupta, P.K.; Varshney, R.K. The development and use of microsatellite markers for genetic analysis and plant breeding with emphasis on bread wheat. Euphytica 2000, 113, 163–185. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Morgante, M.; Olivieri, A.M. PCR-amplified microsatellites as markers in plant genetics. Plant J. 1993. [Google Scholar] [CrossRef]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C. de F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef]
- Belaj, A.; Satovic, Z.; Cipriani, G.; Baldoni, L.; Testolin, R.; Rallo, L.; Trujillo, I. Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and of their effectiveness in establishing genetic relationships in olive. Theor. Appl. Genet. 2003, 107, 736–744. [Google Scholar] [CrossRef]
- De la Rosa, R.; Angiolillo, A.; Guerrero, C.; Pellegrini, M.; Rallo, L.; Besnard, G.; Bervillé, A.; Martin, A.; Baldoni, L. A first linkage map of olive (Olea europaea L.) cultivars using RAPD, AFLP, RFLP and SSR markers. Theor. Appl. Genet. 2003, 106, 1273–1282. [Google Scholar] [CrossRef]
- Bandelj, D.; Jakše, J.; Javornik, B. Assessment of genetic variability of olive varieties by microsatellite and AFLP markers. Euphytica 2004, 136, 93–102. [Google Scholar] [CrossRef]
- Wu, S.; Collins, G.; Sedgley, M. A molecular linkage map of olive (Olea europaea L.) based on RAPD, microsatellite, and SCAR markers. Genome 2004, 47, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, C.; Simeone, R.; Pasqualone, A.; Ferrara, E.; Blanco, A. Genetic relationships and cultivar identification among 112 olive accessions using AFLP and SSR markers. J. Hortic. Sci. Biotechnol. 2005, 80, 105–110. [Google Scholar] [CrossRef]
- Ganino, T.; Beghè, D.; Valenti, S.; Nisi, R.; Fabbri, A. RAPD and SSR markers for characterization and identification of ancient cultivars of Olea europaea L. in the Emilia region, Northern Italy. Genet. Resour. Crop Evol. 2007, 54, 1531–1540. [Google Scholar] [CrossRef]
- Gomes, S.; Martins-Lopes, P.; Lima-Brito, J.; Meirinhos, J.; Lopes, J.; Martins, A.; Guedes-Pinto, H. Evidence for clonal variation in “Verdeal-Transmontana” olive using RAPD, ISSR and SSR markers. J. Hortic. Sci. Biotechnol. 2008, 83, 395–400. [Google Scholar] [CrossRef]
- Khadari, B.; El Aabidine, A.Z.; Grout, C.; Ben Sadok, I.; Doligez, A.; Moutier, N.; Santoni, S.; Costes, E. A Genetic Linkage Map of Olive Based on Amplified Fragment Length Polymorphism, Intersimple Sequence Repeat and Simple Sequence Repeat Markers. J. Am. Soc. Hortic. Sci. 2010, 135, 548–555. [Google Scholar] [CrossRef]
- Linos, A.; Nikoloudakis, N.; Katsiotis, A.; Hagidimitriou, M. Genetic structure of the Greek olive germplasm revealed by RAPD, ISSR and SSR markers. Sci. Hortic. 2014, 175, 33–43. [Google Scholar] [CrossRef]
- Kaya, H.B.; Cetin, O.; Kaya, H.; Sahin, M.; Sefer, F.; Kahraman, A.; Tanyolac, B. SNP Discovery by Illumina-Based Transcriptome Sequencing of the Olive and the Genetic Characterization of Turkish Olive Genotypes Revealed by AFLP, SSR and SNP Markers. PLoS ONE 2013, 8, e93146. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.B.; Cetin, O.; Kaya, H.S.; Sahin, M.; Sefer, F.; Tanyolac, B. Association Mapping in Turkish Olive Cultivars Revealed Significant Markers Related to Some Important Agronomic Traits. Biochem. Genet. 2016, 54, 506–533. [Google Scholar] [CrossRef]
- Angiolillo, A.; Mencuccini, M.; Baldoni, L. Olive genetic diversity assessed using amplified fragment length polymorphisms. Theor. Appl. Genet. 1999, 98, 411–421. [Google Scholar] [CrossRef]
- Lumaret, R.; Ouazzani, N.; Michaud, H.; Vivier, G.; Deguilloux, M.-F.; Di Giusto, F. Allozyme variation of oleaster populations (wild olive tree) (Olea europaea L.) in the Mediterranean Basin. Heredity 2004, 92, 343–351. [Google Scholar] [CrossRef]
- Zohary, D.; Spiegel-Roy, P. Beginnings of Fruit Growing in the Old World. Science 1975, 187, 319–327. [Google Scholar] [CrossRef]
- Lavee, S. ¿Por qué la necesidad de nuevas variedades de olivos? Fruticultura 1994, 62, 29–37. [Google Scholar]
- Van Hintum, T.J.L.; Brown, A.H.D.; Spillane, C.; Hodgkin, T. Core Collections of Plant Genetic Resources; IPGRI Technical Bulletin No. 3; International Plant Genetic Resources Institute: Rome, Italy, 2000; ISBN 92-9043-454-6. [Google Scholar]
- Bartolini, G.; Prevost, G.; Messeri, C.; Carignani, G. Olive Germplasm: Cultivars and World-Wide Collections; FAO: Rome, Italy, 1998. [Google Scholar]
- Rallo, L.; Barranco, D.; Díez, C.M.; Rallo, P.; Suárez, M.P.; Trapero, C.; Pliego-Alfaro, F. Strategies for Olive (Olea europaea L.) Breeding: Cultivated Genetic Resources and Crossbreeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 3, pp. 535–600. ISBN 978-3-319-91944-7. [Google Scholar]
- Baldoni, L.; Belaj, A. Olive. In Oil Crops; Vollmann, J., Rajcan, I., Eds.; Springer: New York, NY, USA, 2009; pp. 397–421. ISBN 978-0-387-77594-4. [Google Scholar]
- Belaj, A. Germplasm bank: WOGBC-IFAPA (ESP046). In Proceedings of the International Olive Council (IOC) Network of Germplasm Banks and Phytosanitary Management II Workshop True Healthy Olive Cultivars, Videoconference, 9–11 December 2020. [Google Scholar]
- Morello, P. Germplasm Bank WOGBC-UCO Spain. In Proceedings of the International Olive Council (IOC) Network of Germplasm Banks and Phytosanitary Management II Workshop True Healthy Olive Cultivars, Videoconference, 9–11 December 2020. [Google Scholar]
- Haouane, H.; El Bakkali, A.; Moukhli, A.; Tollon, C.; Santoni, S.; Oukabli, A.; El Modafar, C.; Khadari, B. Genetic structure and core collection of the World Olive Germplasm Bank of Marrakech: Towards the optimised management and use of Mediterranean olive genetic resources. Genetica 2011, 139, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Veral, G.M. Germplasm bank: Izmir World Olive and Collection Turkish National Olive Collection. In Proceedings of the International Olive Council (IOC) Network of Germplasm Banks and Phytosanitary Management II Workshop True Healthy Olive Cultivars, Videoconference, 9–11 December 2020. [Google Scholar]
- Rallo, P.; Dorado, G.; Martín, A. Development of simple sequence repeats (SSRs) in olive tree (Olea europaea L.). Theor. Appl. Genet. 2000, 101, 984–989. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendonça, D.; Dos Santos, M.R.; Da Câmara Machado, M.L.; Da Câmara Machado, A. Identification of microsatellite loci in olive (Olea europaea) and their characaterization in Italian and Iberian olive trees. Mol. Ecol. 2000, 9, 1171–1173. [Google Scholar] [CrossRef] [PubMed]
- Carriero, F.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). TAG Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europaea L.) are suitable for individual fingerprinting and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- De La Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellites in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Díaz, A.; De La Rosa, R.; Martín, A.; Rallo, P. Development, characterization and inheritance of new microsatellites in olive (Olea europaea L.) and evaluation of their usefulness in cultivar identification and genetic relationship studies. Tree Genet. Genomes 2006, 2, 165–175. [Google Scholar] [CrossRef]
- Gil, F.S.; Busconi, M.; Da Câmara Machado, A.; Fogher, C. Development and characterization of microsatellite loci from Olea europaea. Mol. Ecol. Notes 2006, 6, 1275–1277. [Google Scholar] [CrossRef]
- Munoz-Merida, A.; Gonzalez-Plaza, J.J.; Canada, A.; Blanco, A.M.; Garcia-Lopez, M.d.C.; Rodriguez, J.M.; Pedrola, L.; Sicardo, M.D.; Hernandez, M.L.; De la Rosa, R.; et al. De Novo Assembly and Functional Annotation of the Olive (Olea europaea) Transcriptome. DNA Res. 2013, 20, 93–108. [Google Scholar] [CrossRef]
- De la Rosa, R.; Belaj, A.; Munoz-Merida, A.; Trelles, O.; Ortiz-Martin, I.; Gonzalez-Plaza, J.J.; Valpuesta, V.; Beuzon, C.R. Development of EST-derived SSR Markers with Long-core Repeat in Olive and Their Use for Paternity Testing. J. Am. Soc. Hort. Sci. 2013, 138, 290–296. [Google Scholar] [CrossRef]
- Adawy, S.S.; Mokhtar, M.M.; Alsamman, A.M.; Sakr, M.M. Development of Annotated EST-SSR Database in Olive (Olea europaea). Int. J. Sci. Res. 2015, 14, 1063–1073. [Google Scholar]
- Alagna, F.; D’Agostino, N.; Torchia, L.; Servili, M.; Rao, R.; Pietrella, M.; Giuliano, G.; Chiusano, M.L.; Baldoni, L.; Perrotta, G. Comparative 454 pyrosequencing of transcripts from two olive genotypes during fruit development. BMC Genom. 2009. [Google Scholar] [CrossRef]
- Alagna, F.; Cirilli, M.; Galla, G.; Carbone, F.; Daddiego, L.; Facella, P.; Lopez, L.; Colao, C.; Mariotti, R.; Cultrera, N.; et al. Transcript analysis and regulative events during flower development in olive (Olea europaea L.). PLoS ONE 2016. [Google Scholar] [CrossRef]
- Corrado, G.; Alagna, F.; Rocco, M.; Renzone, G.; Varricchio, P.; Coppola, V.; Coppola, M.; Garonna, A.; Baldoni, L.; Scaloni, A.; et al. Molecular interactions between the olive and the fruit fly Bactrocera oleae. BMC Plant Biol. 2012, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, R.; Cultrera, N.G.M.; Mousavi, S.; Baglivo, F.; Rossi, M.; Albertini, E.; Alagna, F.; Carbone, F.; Perrotta, G.; Baldoni, L. Development, evaluation, and validation of new EST-SSR markers in olive (Olea europaea L.). Tree Genet. Genomes 2016, 12, 120. [Google Scholar] [CrossRef]
- Arbeiter, A.B.; Hladnik, M.; Jakše, J.; Bandelj, D.; Arbeiter, A.B.; Hladnik, M.; Jakše, J.; Bandelj, D. Identification and validation of novel EST-SSR markers in olives. Sci. Agric. 2017, 74, 215–225. [Google Scholar] [CrossRef]
- Resetic, T.; Stajner, N.; Bandelj, D.; Javornik, B.; Jakse, J. Validation of candidate reference genes in RT-qPCR studies of developing olive fruit and expression analysis of four genes involved in fatty acids metabolism. Mol. Breed. 2013, 32, 211–222. [Google Scholar] [CrossRef]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Comparative assessment of genetic diversity in Albanian olive (Olea europaea L.) using SSRs from anonymous and transcribed genomic regions. Tree Genet. Genomes 2018, 14, 53. [Google Scholar] [CrossRef]
- Li, D.; Long, C.; Pang, X.; Ning, D.; Wu, T.; Dong, M.; Han, X.; Guo, H. The newly developed genomic-SSR markers uncover the genetic characteristics and relationships of olive accessions. PeerJ 2020, 8, e8573. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Rodríguez, M.V.; Beuzon, C.; González-Plaza, J.J.; Fernández-Ocaña, A.M. Identification of an olive (Olea europaea L.) core collection with a new set of SSR markers. Genet. Resour. Crop Evol. 2021, 68, 117–133. [Google Scholar] [CrossRef]
- Rallo, P.; Tenzer, I.; Gessler, C.; Baldoni, L.; Dorado, G.; Martín, A. Transferability of olive microsatellite loci across the genus Olea. Theor. Appl. Genet. 2003, 107, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Doveri, S.; Sabino Gil, F.; Díaz, A.; Reale, S.; Busconi, M.; da Câmara Machado, A.; Martín, A.; Fogher, C.; Donini, P.; Lee, D. Standardization of a set of microsatellite markers for use in cultivar identification studies in olive (Olea europaea L.). Sci. Hortic. 2008, 116, 367–373. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Ricciolini, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellite markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Aksehirli-Pakyurek, M.; Koubouris, G.C.; Petrakis, P.V.; Hepaksoy, S.; Metzidakis, I.T.; Yalcinkaya, E.; Doulis, A.G. Cultivated and Wild Olives in Crete, Greece—Genetic Diversity and Relationships with Major Turkish Cultivars Revealed by SSR Markers. Plant Mol. Biol. Report. 2017, 35, 575–585. [Google Scholar] [CrossRef]
- Bandelj, D.; Jakse, J.; Javornik, B. DNA fingerprinting of olive varieties by microsatellite markers. Food Technol. Biotechnol. 2002, 40, 185–190. [Google Scholar]
- Mackay, J.F.; Wright, C.D.; Bonfiglioli, R.G. A new approach to varietal identification in plants by microsatellite high resolution melting analysis: Application to the verification of grapevine and olive cultivars. Plant Methods 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Xanthopoulou, A.; Ganopoulos, I.; Koubouris, G.; Tsaftaris, A.; Sergendani, C.; Kalivas, A.; Madesis, P. Microsatellite high-resolution melting (SSR-HRM) analysis for genotyping and molecular characterization of an Olea europaea germplasm collection. Plant Genet. Resour. 2014, 12, 273–277. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Argiriou, A.; Tsaftaris, A. Microsatellite high resolution melting (SSR-HRM) analysis for authenticity testing of protected designation of origin (PDO) sweet cherry products. Food Control 2011, 22, 532–541. [Google Scholar] [CrossRef]
- Distefano, G.; Caruso, M.; La Malfa, S.; Gentile, A.; Wu, S.B. High Resolution Melting Analysis Is a More Sensitive and Effective Alternative to Gel-Based Platforms in Analysis of SSR—An Example in Citrus. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Poljuha, D.; Sladonja, B.; Šetić, E.; Milotić, A.; Bandelj, D.; Jakše, J.; Javornik, B. DNA fingerprinting of olive varieties in Istria (Croatia) by microsatellite markers. Sci. Hortic. 2008, 115, 223–230. [Google Scholar] [CrossRef]
- Carmona, R.; Zafra, A.; Seoane, P.; Castro, A.J.; Guerrero-Fernández, D.; Castillo-Castillo, T.; Medina-García, A.; Cánovas, F.M.; Aldana-Montes, J.F.; Navas-Delgado, I.; et al. ReprOlive: A database with linked data for the olive tree (Olea europaea L.) reproductive transcriptome. Front. Plant Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Ben Ayed, R.; Ben Hassen, H.; Ennouri, K.; Ben Marzoug, R.; Rebai, A. OGDD (Olive Genetic Diversity Database): A microsatellite markers’ genotypes database of worldwide olive trees for cultivar identification and virgin olive oil traceability. Database 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, I.; Barranco, D.; Cabello, A.; Gordon, P.; Morello, C.; Díez, M.; Rallo, L. Proposal of a Guide for the cataloging, sanitation and management of the Germplasm Banks of the IOC Network (UCOLIVO). In Proceedings of the International Seminar. The IOC Network of Germplasm Banks and the True Healthy Olive Cultivars Project, Córdoba, Spain, 21–24 October 2019. [Google Scholar]
- Barranco, D.; Cimato, A.; Fiorino, P.; Rallo, L.; Touzani, A.; Castañeda, C.; Serafini, F.; Trujillo, I. World Catalogue of Olive Cultivars; International Olive Oil Council: Madrid, Spain, 2000. [Google Scholar]
- Rallo, L.; Barranco, D.; Castro-García, S.; Connor, D.J.; Gómez del Campo, M.; Rallo, P. High-Density Olive Plantations. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; Volume 41, pp. 303–384. [Google Scholar]
- Fernández i Martí, A.; Font i Forcada, C.; Socias i Company, R.; Rubio-Cabetas, M.J. Genetic relationships and population structure of local olive tree accessions from Northeastern Spain revealed by SSR markers. Acta Physiol. Plant. 2015, 37. [Google Scholar] [CrossRef]
- Sakar, E.; Unver, H.; Bakir, M.; Ulas, M.; Sakar, Z.M. Genetic Relationships Among Olive (Olea europaea L.) Cultivars Native to Turkey. Biochem. Genet. 2016, 54, 348–359. [Google Scholar] [CrossRef]
- Sakar, E.; Unver, H.; Ulas, M.; Lazovic, B.; Ercisli, S. Genetic Diversity and Relationships among Local Olive (Olea europeaea L.) Genotypes from Gaziantep Province and Notable Cultivars in Turkey, Based on SSR Markers. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 557–562. [Google Scholar] [CrossRef]
- Unver, H.; Sakar, E.; Ulas, M.; Ercisli, S.; Ak, B. Molecular characterization of indigenous olive genotypes based on SSR analysis. Genetika 2016, 48, 1017–1025. [Google Scholar] [CrossRef]
- Hmmam, I.; Mariotti, R.; Ruperti, B.; Cultrera, N.; Baldoni, L.; Barcaccia, G. Venetian olive (Olea europaea) germplasm: Disclosing the genetic identity of locally grown cultivars suited for typical extra virgin oil productions. Genet. Resour. Crop Evol. 2018, 65, 1733–1750. [Google Scholar] [CrossRef]
- Reboredo-Rodríguez, P.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J.; Trujillo, I. Genotypic and phenotypic identification of olive cultivars from north-western Spain and characterization of their extra virgin olive oils in terms of fatty acid composition and minor compounds. Sci. Hortic. 2018, 232, 269–279. [Google Scholar] [CrossRef]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Rotondi, A.; Ganino, T.; Beghè, D.; Di Virgilio, N.; Morrone, L.; Fabbri, A.; Neri, L. Genetic and landscape characterization of ancient autochthonous olive trees in northern Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2018, 152, 1067–1074. [Google Scholar] [CrossRef]
- Ninot, A.; Howad, W.; Aranzana, M.J.; Senar, R.; Romero, A.; Mariotti, R.; Baldoni, L.; Belaj, A. Survey of over 4, 500 monumental olive trees preserved on-farm in the northeast Iberian Peninsula, their genotyping and characterization. Sci. Hortic. 2018, 231, 253–264. [Google Scholar] [CrossRef]
- Erre, P.; Chessa, I.; Muñoz-Diez, C.; Belaj, A.; Rallo, L.; Trujillo, I. Genetic diversity and relationships between wild and cultivated olives (Olea europaea L.) in Sardinia as assessed by SSR markers. Genet. Resour. Crop Evol. 2010, 57, 41–54. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Porceddu, A.; Barranco, D.; Satovic, Z. Genetic diversity and population structure of wild olives from the north-western Mediterranean assessed by SSR markers. Ann. Bot. 2007, 100, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Belaj, A.; León, L.; Satovic, Z.; De la Rosa, R. Variability of wild olives (Olea europaea subsp. europaea var. sylvestris) analyzed by agro-morphological traits and SSR markers. Sci. Hortic. 2011, 129, 561–569. [Google Scholar] [CrossRef]
- Breton, C.; Tersac, M.; Bervillé, A. Genetic diversity and gene flow between the wild olive (oleaster, Olea europaea L.) and the olive: Several Plio-Pleistocene refuge zones in the Mediterranean basin suggested by simple sequence repeats analysis. J. Biogeogr. 2006, 33, 1916–1928. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Ben El Hadj, S.; El Gazzah, M.; Bervillé, A. Are olive cultivars distinguishable from oleaster trees based on morphology of drupes and pits, oil composition and microsatellite polymorphisms? Acta Bot. Gall. 2008, 155, 531–545. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Ben El Hadj, S.; El Gazzah, M.; Bervillé, A. Differences between native and introduced olive cultivars as revealed by morphology of drupes, oil composition and SSR polymorphisms: A case study in Tunisia. Sci. Hortic. 2008, 116, 280–290. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Hadj, S.B.E.; Gazzah, M.E.; Berville, A. Genetic Relationships between Cultivated and Wild Olive Trees (Olea europaea L. var. europaea and var. sylvestris) Based on Nuclear and Chloroplast SSR Markers. Nat. Resour. 2010, 1, 95–103. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Satovic, Z.; Barranco, D. Genetic diversity and relationships of wild and cultivated olives at regional level in Spain. Sci. Hortic. 2010, 124, 323–330. [Google Scholar] [CrossRef]
- Yoruk, B.; Taskin, V. Genetic diversity and relationships of wild and cultivated olives in Turkey. Plant Syst. Evol. 2014, 300, 1247–1258. [Google Scholar] [CrossRef]
- Boucheffa, S.; Miazzi, M.M.; di Rienzo, V.; Mangini, G.; Fanelli, V.; Tamendjari, A.; Pignone, D.; Montemurro, C. The coexistence of oleaster and traditional varieties affects genetic diversity and population structure in Algerian olive (Olea europaea) germplasm. Genet. Resour. Crop Evol. 2017, 64, 379–390. [Google Scholar] [CrossRef]
- Chiappetta, A.; Muto, A.; Muzzalupo, R.; Muzzalupo, I. New rapid procedure for genetic characterization of Italian wild olive (Olea europaea) and traceability of virgin olive oils by means of SSR markers. Sci. Hortic. 2017, 226, 42–49. [Google Scholar] [CrossRef]
- Belaj, A.; Veral, M.G.; Sikaoui, H.; Moukhli, A.; Khadari, B.; Mariotti, R.; Baldoni, L. Olive Genetic Resources. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 27–54. [Google Scholar]
- Omrani-Sabbaghi, A.; Shahriari, M.; Falahati-Anbaran, M.; Mohammadi, S.A.; Nankali, A.; Mardi, M.; Ghareyazie, B. Microsatellite markers based assessment of genetic diversity in Iranian olive (Olea europaea L.) collections. Sci. Hortic. 2007, 112, 439–447. [Google Scholar] [CrossRef]
- Stambuk, S.; Sutlović, D.; Bakarić, P.; Petricević, S.; Andelinović, S. Forensic botany: Potential usefulness of microsatellite-based genotyping of Croatian olive (Olea europaea L.) in forensic casework. Croat. Med. J. 2007, 48, 556–562. [Google Scholar]
- Cantini, C.; Cimato, A.; Autino, A.; Redi, A.; Cresti, M. Assessment of the Tuscan Olive Germplasm by Microsatellite Markers Reveals Genetic Identities and Different Discrimination Capacity among and within Cultivars. J. Am. Soc. Hortic. Sci. 2008, 133, 598–604. [Google Scholar] [CrossRef]
- Ganino, T.; Beghè, D.; Rotondi, A.; Fabbri, A. Genetic resources of Olea europaea L. in the Bologna province (Italy): SSR analysis and identification of local germplasm. Adv. Hortic. Sci. 2008, 22, 149–155. [Google Scholar] [CrossRef]
- Rekik, I.; Salimonti, A.; Grati Kamoun, N.; Muzzalupo, I.; Lepais, O.; Gerber, S.; Perri, E.; Rebai, A. Characterization and Identification of Tunisian Olive Tree Varieties by Microsatellite Markers. HortScience 2008, 43, 1371–1376. [Google Scholar] [CrossRef]
- Ipek, A.; Barut, E.; Gulen, H.; Oz, A.T.; Tangu, N.A.; Ipek, M. SSR analysis demonstrates that olive production in the southern Marmara region in Turkey uses a single genotype. Genet. Mol. Res. 2009, 8, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, Z.; Hosseini-Mazinani, M.; Trujillo, I.; Belaj, A. Study of intracultivar variation among main Iranian olive cultivars using SSR markers. Acta Biol. Szeged. 2009, 53, 27–32. [Google Scholar]
- Corrado, G.; Imperato, A.; la Mura, M.; Perri, E.; Rao, R. Genetic diversity among olive varieties of southern italy and the traceability of olive oil using SSR markers. J. Hortic. Sci. Biotechnol. 2011, 86, 461–466. [Google Scholar] [CrossRef]
- Ercisli, S.; Ipek, A.; Barut, E. SSR Marker-Based DNA Fingerprinting and Cultivar Identification of Olives (Olea europaea). Biochem. Genet. 2011, 49, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.U.; Mailer, R.J.; Belaj, A.; De La Rosa, R.; Raman, H. Microsatellite marker-based identification of mother plants for the reliable propagation of olive (Olea europaea L.) cultivars in Australia. J. Hortic. Sci. Biotechnol. 2012, 87, 647–653. [Google Scholar] [CrossRef]
- El Bakkali, A.; Haouane, H.; Moukhli, A.; Costes, E.; van Damme, P.; Khadari, B. Construction of Core Collections Suitable for Association Mapping to Optimize Use of Mediterranean Olive (Olea europaea L.) Genetic Resources. PLoS ONE 2013, 8, e61265. [Google Scholar] [CrossRef]
- Abdessemed, S.; Muzzalupo, I.; Benbouza, H. Assessment of genetic diversity among Algerian olive (Olea europaea L.) cultivars using SSR marker. Sci. Hortic. 2015, 192, 10–20. [Google Scholar] [CrossRef]
- Beghè, D.; Molano, J.F.G.; Fabbri, A.; Ganino, T. Olive biodiversity in Colombia. A molecular study of local germplasm. Sci. Hortic. 2015, 189, 122131. [Google Scholar] [CrossRef]
- Casadei, E.; Valli, E.; Panni, F.; Donarski, J.; Farrús Gubern, J.; Lucci, P.; Conte, L.; Lacoste, F.; Maquet, A.; Brereton, P.; et al. Emerging trends in olive oil fraud and possible countermeasures. Food Control 2021, 124, 107902. [Google Scholar] [CrossRef]
- Kiritsakis, A.; Christie, W.W. Analysis of Edible Oils. In Handbook of Olive Oil; Harwood, J., Aparicio, R., Eds.; Springer US: Boston, MA, USA, 2000; pp. 129–158. ISBN 978-1-4757-5371-4. [Google Scholar]
- Downey, G.; Boussion, J. Authentication of Coffee Bean Variety by Near-infrared Reflectance Spectroscopy of Dried Extract. J. Sci. Food Agric. 1996, 71, 41–49. [Google Scholar] [CrossRef]
- Melchiade, D.; Foroni, I.; Corrado, G.; Santangelo, I.; Rao, R. Authentication of the ‘Annurca’ apple in agro-food chain by ampli-fication of microsatellite loci. Food Biotechnol. 2007, 21, 33–43. [Google Scholar] [CrossRef]
- Mouly, P.P.; Gaydou, E.M.; Faure, R.; Estienne, J.M. Blood Orange Juice Authentication Using Cinnamic Acid Derivatives. Variety Differentiations Associated with Flavanone Glycoside Content. J. Agric. Food Chem. 1997, 45, 373–377. [Google Scholar] [CrossRef]
- Montealegre, C.; Marina Alegre, M.L.; García-Ruiz, C. Traceability Markers to the Botanical Origin in Olive Oils. J. Agric. Food Chem. 2010, 58, 28–38. [Google Scholar] [CrossRef]
- Bajoub, A.; Bendini, A.; Fernández-Gutiérrez, A.; Carrasco-Pancorbo, A. Olive oil authentication: A comparative analysis of regulatory frameworks with especial emphasis on quality and authenticity indices, and recent analytical techniques developed for their assessment. A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 832–857. [Google Scholar] [CrossRef] [PubMed]
- Lukić, I.; Ros, A.D.; Guella, G.; Camin, F.; Masuero, D.; Mulinacci, N.; Vrhovsek, U.; Mattivi, F. Lipid profiling and stable isotopic data analysis for differentiation of extra virgin olive oils based on their origin. Molecules 2020, 25, 4. [Google Scholar] [CrossRef]
- Jiménez-Morillo, N.T.; Palma, V.; Garcia, R.; Dias, C.B.; Cabrita, M.J. Combination of Stable Isotope Analysis and Chemometrics to Discriminate Geoclimatically and Temporally the Virgin Olive Oils from Three Mediterranean Countries. Foods 2020, 9, 1855. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ayed, R.; Kamoun-Grati, N.; Rebai, A. An overview of the authentication of olive tree and oil. Compr. Rev. Food Sci. Food Saf. 2013, 12, 218–227. [Google Scholar] [CrossRef]
- Costa, J.; Mafra, I.; Oliveira, M.B.P.P. Advances in vegetable oil authentication by DNA-based markers. Trends Food Sci. Technol. 2012, 26, 43–55. [Google Scholar] [CrossRef]
- Breton, C.; Claux, D.; Metton, I.; Skorski, G.; Bervillé, A. Comparative Study of Methods for DNA Preparation from Olive Oil Samples to Identify Cultivar SSR Alleles in Commercial Oil Samples: Possible Forensic Applications. J. Agric. Food Chem. 2004, 52, 531–537. [Google Scholar] [CrossRef]
- Testolin, R.; Lain, O. DNA Extraction from Olive Oil and PCR Amplification of Microsatellite Markers. J. Food Sci. 2005, 70, C108–C112. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Pellegrino, M.; Perri, E. Detection of DNA in virgin olive oils extracted from destoned fruits. Eur. Food Res. Technol. 2007, 224, 469–475. [Google Scholar] [CrossRef]
- Ben Ayed, R.; Grati-Kamoun, N.; Moreau, F.; Rebaï, A. Comparative study of microsatellite profiles of DNA from oil and leaves of two Tunisian olive cultivars. Eur. Food Res. Technol. 2009, 229, 757–762. [Google Scholar] [CrossRef]
- Raieta, K.; Muccillo, L.; Colantuoni, V. A novel reliable method of DNA extraction from olive oil suitable for molecular traceability. Food Chem. 2015, 172, 596–602. [Google Scholar] [CrossRef]
- Piarulli, L.; Savoia, M.A.; Taranto, F.; D’Agostino, N.; Sardaro, R.; Girone, S.; Gadaleta, S.; Fucili, V.; De Giovanni, C.; Montemurro, C.; et al. A Robust DNA Isolation Protocol from Filtered Commercial Olive Oil for PCR-Based Fingerprinting. Foods 2019, 8, 462. [Google Scholar] [CrossRef]
- Consolandi, C.; Palmieri, L.; Severgnini, M.; Maestri, E.; Marmiroli, N.; Agrimonti, C.; Baldoni, L.; Donini, P.; De Bellis, G.; Castiglioni, B. A procedure for olive oil traceability and authenticity: DNA extraction, multiplex PCR and LDR-universal array analysis. Eur. Food Res. Technol. 2008, 227, 1429–1438. [Google Scholar] [CrossRef]
- Carvalho, J.; Puertas, G.; Gaspar, J.; Azinheiro, S.; Diéguez, L.; Garrido-Maestu, A.; Vázquez, M.; Barros-Velázquez, J.; Cardoso, S.; Prado, M. Highly efficient DNA extraction and purification from olive oil on a washable and reusable miniaturized device. Anal. Chim. Acta 2018, 1020, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; Montemurro, C.; Caponio, F.; Blanco, A. Identification of Virgin Olive Oil from Different Cultivars by Analysis of DNA Microsatellites. J. Agric. Food Chem. 2004, 52, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; Montemurro, C.; Summo, C.; Sabetta, W.; Caponio, F.; Blanco, A. Effectiveness of Microsatellite DNA Markers in Checking the Identity of Protected Designation of Origin Extra Virgin Olive Oil. J. Agric. Food Chem. 2007, 55, 3857–3862. [Google Scholar] [CrossRef]
- Martins-Lopes, P.; Gomes, S.; Santos, E.; Guedes-Pinto, H. DNA Markers for Portuguese Olive Oil Fingerprinting. J. Agric. Food Chem. 2008, 56, 11786–11791. [Google Scholar] [CrossRef] [PubMed]
- Alba, V.; Sabetta, W.; Blanco, A.; Pasqualone, A.; Montemurro, C. Microsatellite markers to identify specific alleles in DNA extracted from monovarietal virgin olive oils. Eur. Food Res. Technol. 2009, 229, 375–382. [Google Scholar] [CrossRef]
- Vietina, M.; Agrimonti, C.; Marmiroli, M.; Bonas, U.; Marmiroli, N. Applicability of SSR markers to the traceability of monovarietal olive oils. J. Sci. Food Agric. 2011, 91, 1381–1391. [Google Scholar] [CrossRef]
- Ben-Ayed, R.; Grati-Kamoun, N.; Sans-Grout, C.; Moreau, F.; Rebai, A. Characterization and authenticity of virgin olive oil (Olea europaea L.) cultivars by microsatellite markers. Eur. Food Res. Technol. 2012, 234, 263–271. [Google Scholar] [CrossRef]
- Pasqualone, A.; Di Rienzo, V.; Nasti, R.; Blanco, A.; Gomes, T.; Montemurro, C. Traceability of Italian Protected Designation of Origin (PDO) table olives by means of microsatellite molecular markers. J. Agric. Food Chem. 2013, 61, 3068–3073. [Google Scholar] [CrossRef]
- Pérez-Jiménez, M.; Besnard, G.; Dorado, G.; Hernandez, P. Varietal tracing of virgin olive oils based on plastid DNA variation profiling. PLoS ONE 2013, 8, e70507. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Pisani, F.; Greco, F.; Chiappetta, A. Direct DNA amplification from virgin olive oil for traceability and authenticity. Eur. Food Res. Technol. 2015, 241, 151–155. [Google Scholar] [CrossRef]
- Montemurro, C.; Miazzi, M.M.; Pasqualone, A.; Fanelli, V.; Sabetta, W.; Di Rienzo, V. Traceability of PDO Olive Oil “Terra di Bari” Using High Resolution Melting. J. Chem. 2015, 2015, 1–7. [Google Scholar] [CrossRef]
- Pasqualone, A.; Rienzo, V.D.; Miazzi, M.M.; Fanelli, V.; Caponio, F.; Montemurro, C. High resolution melting analysis of DNA microsatellites in olive pastes and virgin olive oils obtained by talc addition. Eur. J. Lipid Sci. Technol. 2015, 117, 2044–2048. [Google Scholar] [CrossRef]
- Gomes, S.; Breia, R.; Carvalho, T.; Carnide, V.; Martins-Lopes, P. Microsatellite High-Resolution Melting (SSR-HRM) to Track Olive Genotypes: From Field to Olive Oil. J. Food Sci. 2018, 83, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; Di Rienzo, V.; Sabetta, W.; Fanelli, V.; Summo, C.; Paradiso, V.M.; Montemurro, C.; Caponio, F. Chemical and Molecular Characterization of Crude Oil Obtained by Olive-Pomace Recentrifugation. J. Chem. 2016, 2016, 1–7. [Google Scholar] [CrossRef]
- Rotondi, A.; Beghè, D.; Fabbri, A.; Ganino, T. Olive oil traceability by means of chemical and sensory analyses: A comparison with SSR biomolecular profiles. Food Chem. 2011, 129, 1825–1831. [Google Scholar] [CrossRef]
- Crawford, L.M.; Carrasquilla-Garcia, N.; Cook, D.; Wang, S.C. Analysis of Microsatellites (SSRs) in Processed Olives as a Means of Cultivar Traceability and Authentication. J. Agric. Food Chem. 2020, 68, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Crawford, L.M.; Janovick, J.L.; Carrasquilla-Garcia, N.; Hatzakis, E.; Wang, S.C. Comparison of DNA analysis, targeted metabolite profiling, and non-targeted NMR fingerprinting for differentiating cultivars of processed olives. Food Control 2020, 114, 107264. [Google Scholar] [CrossRef]
- Chedid, E.; Rizou, M.; Kalaitzis, P. Application of high resolution melting combined with DNA-based markers for quantitative analysis of olive oil authenticity and adulteration. Food Chem. X 2020, 6, 100082. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reference | Naming of SSR Loci (Prefixes) | Type of SSR | No. of Polymorphic SSRs Reported | No. of Cultivars Used in Characterization of SSRs | Na | Ho | He |
|---|---|---|---|---|---|---|---|
| [37] | IAS-oli | Genomic | 05 | 46 | 3–9 | - | 0.460–0.710 |
| [38] | DCA | Genomic | 15 | 47 | 4–15 | 0.283–0.979 | 0.357–0.859 |
| [41] | EMO | Genomic | 13 | 23 | 6–9 | 0.391–0.913 | 0.620–0.811 |
| [39] | GAPU | Genomic | 10 | 20 | 3–9 | - | - |
| [40] | UDO99 | Genomic | 28 | 13 | 1–5 | - | 0.000–0.770 |
| [43] | ssrOeIGP | Genomic | 12 | 33 | 2–14 | 0.188–0.813 | 0.417–0.895 |
| [42] | IAS-oli | Genomic | 12 | 51 | 1–13 | ||
| [45] | OLEAGEN-H | EST-SSR | 08 | 15 | 2–7 | 0.380–1.000 | 0.490–0.850 |
| [46] | Oe-ESSR | EST-SSR | 1801; 25 of these used | 09 | - | - | - |
| [51] | OeUP | EST-SSR | 46 | 24 | 2–8 | 0.042–1.000 | 0.042–0.869 |
| [50] | OLEST | EST-SSR | 26 | 32 | 2–10 | 0.219–0.813 | 0.195–0.839 |
| [53] | SNB, SiBi | EST-SSR | - | - | - | 0.357–0.932 | 0.294–0.790 |
| [54] | BFU | EST-SSR | 21 | 53 | 3–10 | 0.140–0.910 | 0.520–0.810 |
| [55] | Olea | EST-SSR | 08 | 36 | 4–7 | 0.350–0.710 | 0.540–0.750 |
| Objective | Cultivars or Accessions | Region | No. of SSRs | Ho | He | Na | PIC | Reference | Key Remarks |
|---|---|---|---|---|---|---|---|---|---|
| Characterization and Identification of Olive Cultivars | 19 | Slovenia, Italy, France, Spain | 14 (DCA-1,3,4,5,7,8,9,10, 11, 13, 14, 15, 16, 17) | 0.263–1.000 | NA | 3–12 | NA | [60] | Identification key of 19 olive varieties |
| Characterization and Identification of Olive Cultivars | 87 | Iran | 16 (DCA 18, 17, 16, 15, 14, 13, 11, 10, GAPU101, 103A, 89, 71B, 72, 90) | NA | NA | NA | 0.620–0.950 | [92] | Intra-cultivar variation, cultivar denominations and origin investigated using SSRs |
| Characterization of Autochthonous Olives | 44 | Croatia | 16 (UDO-08, 12, 19, 24, 28, 31, 39, 43, DCA3, 8, 9, 10, 14, 16, EMO2, 3) | 0.273–0.932 | 0.499–0.910 | 5–20 | NA | [93] | SSR-based varietal discrimination achieved |
| Germplasm Characterization | 154 | Tuscany | 12 (UDO-04, 06, 09, 11, 12, 17, 19, 24, 27, 31, SIU06, 08) | 0.278–0.722 | 0.428–0.855 | 3–10 | NA | [94] | Homonyms and synonyms detected |
| Database Development and Cultivar Identification | 17 | Mediterranean Basin | 08 (DCA3, 4, 8, 9, 11, 13, 14, 15) | NA | NA | 3–12 | NA | [57] | Standardization of SSR set for olive cultivar studies |
| Characterization and Identification of Olive Cultivars | 18 | Bologna, Italy | 17 (DCA3,4,5,7,9,13,14,15,16,17,18, EMO90, GAPU59,101,103, UDO-24, 43) | 0.500–1.000 | 0.431–0.841 | 4–10 | NA | [95] | Synonyms identified and diversity in germplasm revealed |
| Characterization of Olive Cultivars | 20 | Tunisia | 10 (GAPU59, 71A, 71B, 103A, UDO-03, 12, 28, 39, DCA9, 18) | 0.300–0.950 | 0.562–0.801 | 3–6 | NA | [96] | Cultivars broadly grouped by their end use and phenotypes |
| Characterization of Olive Cultivars | 38 | Southern Marmara region, Turkey | 10 (GAPU103A, 101, UDO-06, 07, 09, 11, 12, 14, 15, 35) | NA | NA | 2–5 | NA | [97] | Cultivar Gemlik revealed as major olive cultivar in the region |
| Characterization of Olive Cultivars | 51 | Iran | 13 (DCA3,9,16,18,11,15, UDO-43, 11, 19, 24, GAPU59, 71B, 101) | 0.000–1.000 | 0.000–0.800 | 1–8 | 0.000–0.750 | [98] | Synonyms, homonyms and intra-cultivar polymorphisms detected |
| Characterization and Diversity Analysis | 47 | Southern Italy | 6 (DCA3, 4, 16, 17, UDO-31, GAPU 47) | 0.700–0.890 | NA | 11–17 | 0.830–0.870 | [99] | Genetic distinctness of accessions from Campania region established |
| Characterization of Olive cultivars | 10 | Turkey | 7 (DCA-4, 9, 11, 16, 17, GAPU-89, UDO-14) | NA | NA | 3–6 | NA | [100] | Misnamings among cultivars identified |
| Cultivar Identification Using SSR | 53 | Australia | 7 (9, 3, 16, 18, 5, EMO90, 30) | 0.490–0.980 | 0.480–0.840 | 7–12 | 0.467–0.813 | [101] | Samples grouped into distinct genotypes |
| Characterization of Olive Germplasm | 561 | Marrakech (OWGB collection) | 17 (DCA1, 3, 4, 5, 8, 9, 11, 14, 15, 18, UDO-36, GAPU59, 71A, 71B, EMO03, 90,PA(ATT)2*) | 0.490–0.928 | 0.454–0.876 | 5–32 | 0.403–0.864 | [102] | Construction of two core collections |
| Cultivar Characterization and Diversity Analysis | 26 | Algeria | 11 (DCA9, 18, GAPU59, 71A, 71B, 101, 103A, UDO-12, 43, 28, 39) | 0.135–0.889 | 0.070–0.510 | 6–21 | NA | [103] | SSR genotyping allowed unambiguous identification of all the cultivars |
| Cultivar characterization and Diversity Analysis | 39 | Colombia | 10 (DCA3, 5, 9, 16, 17, 18, UDO-43, GAPU101, 103A, EMO90) | 0.250–1.000 | 0.312–0.909 | 3–15 | 0.282–0.876 | [104] | Synonyms, homonyms identified and 19 genetic profiles discriminated |
| Sample Type | SSR Markers Used | References |
|---|---|---|
| Oil samples of different origins | EMO series SSR primers | [116] |
| VOO (virgin olive oil) from 10 Italian olive cultivars | DCA4, 15,17; GAPU71, 89, 101; UDO03 | [124] |
| Samples of filtered and unfiltered VOO of cv. Carolea | UDO08, 09, 12, 24, 39, 043 and respective shortened internal primers | [117] |
| VOO of cv. Ogliarola salentina cultivar | GAPU59, 71A 103A and UDO01, 03, 39 | [118] |
| Samples of Collina di Brindisi PDO oil (four unfiltered and two filtered oils); samples of constituent cultivars used for preparation of the PDO mix | UDO09, 19, 25, 35, 044 and GAPU89, 101 | [125] |
| 11 monovarietal olive oil samples from Portuguese cultivars; 12 commercial olive oils | DCA1, 3, 5, and 9 along with 11 RAPD and 08 ISSR primers | [126] |
| Oil from 2 Tunisian olive cultivars: Chemlali and Chetoui | DCA1, 3; GAPU59, 71A, 71B and UDO12 | [119] |
| Monovarietal oils from 7 Italian cultivars (Coratina, Picholine, Toscanina, Cima di Melfi, Frantoio, Leccino and Cellina di Nardo) | DCA3, 4, 7, 14, 15, 18; GAPU103; EMO90; EMOL and UDO43 | [127] |
| Monovarietal extra virgin olive oil (EVOO) from cultivars Pisciottana, Frantoio and Leccino and their 1:1 mixtures | DCA 3, 4, 16, 17; UDO31 and GAPU 47 | [99] |
| 21 monovarietal olive oils from 16 cultivars | EMO30, 90; DCA5, 8, 17, 18; GAPU71B, 89; UDO09; and Shortened DCA14 and EMO30 | [128] |
| VOO from 22 cultivars | DCA1, 3, 4; GAPU59, 71A, 71B; UDO12, 09 | [129] |
| 03 Italian PDO table olives and 07 highly diffused cultivars of table olives | 16 SSR markers of DCA, GAPU, EMO and UDO99 series. | [130] |
| 14 monovarietal and commercial olive oils | 09 cpDNA markers | [131] |
| 3 VOO samples (Frantoio, Italian PDO Terre di Bari and other) | GAPU59, 71A, 71B, 103A; UDO01, 03, 12, 28, 39 and DCA9, 18 | [132] |
| “Terra di Bari” PDO EVOO; 9 Apulia region cultivars; experimental mixtures of oils | 17 SSR markers of DCA, GAPU, EMO and UDO99 series; HRM analysis of DCA18 | [133] |
| Oil from cv. Coratina | SSR-HRM analysis of DCA3, 16, 18; GAPU103A | [134] |
| 10 monovarietal olive oils from different Portuguese PDO regions; 2 commercial EVOO olive oils | HRM analysis of 15 SSR from DCA and UDO99 series | [135] |
| VOO and crude olive pomace oil of cv. Coratina | HRM analysis DCA4, 9 and 14 | [136] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, S.; Carvalho, J.; Trujillo, I.; Prado, M. Microsatellite Markers in Olives (Olea europaea L.): Utility in the Cataloging of Germplasm, Food Authenticity and Traceability Studies. Foods 2021, 10, 1907. https://doi.org/10.3390/foods10081907
Yadav S, Carvalho J, Trujillo I, Prado M. Microsatellite Markers in Olives (Olea europaea L.): Utility in the Cataloging of Germplasm, Food Authenticity and Traceability Studies. Foods. 2021; 10(8):1907. https://doi.org/10.3390/foods10081907
Chicago/Turabian StyleYadav, Shambhavi, Joana Carvalho, Isabel Trujillo, and Marta Prado. 2021. "Microsatellite Markers in Olives (Olea europaea L.): Utility in the Cataloging of Germplasm, Food Authenticity and Traceability Studies" Foods 10, no. 8: 1907. https://doi.org/10.3390/foods10081907
APA StyleYadav, S., Carvalho, J., Trujillo, I., & Prado, M. (2021). Microsatellite Markers in Olives (Olea europaea L.): Utility in the Cataloging of Germplasm, Food Authenticity and Traceability Studies. Foods, 10(8), 1907. https://doi.org/10.3390/foods10081907