
Small RNAs, Degradome, and Transcriptome Sequencing Provide Insights into Papaya Fruit Ripening Regulated by 1-MCP

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. RNA Extraction and cDNA Library Preparation
2.3. RT-qPCR Verification
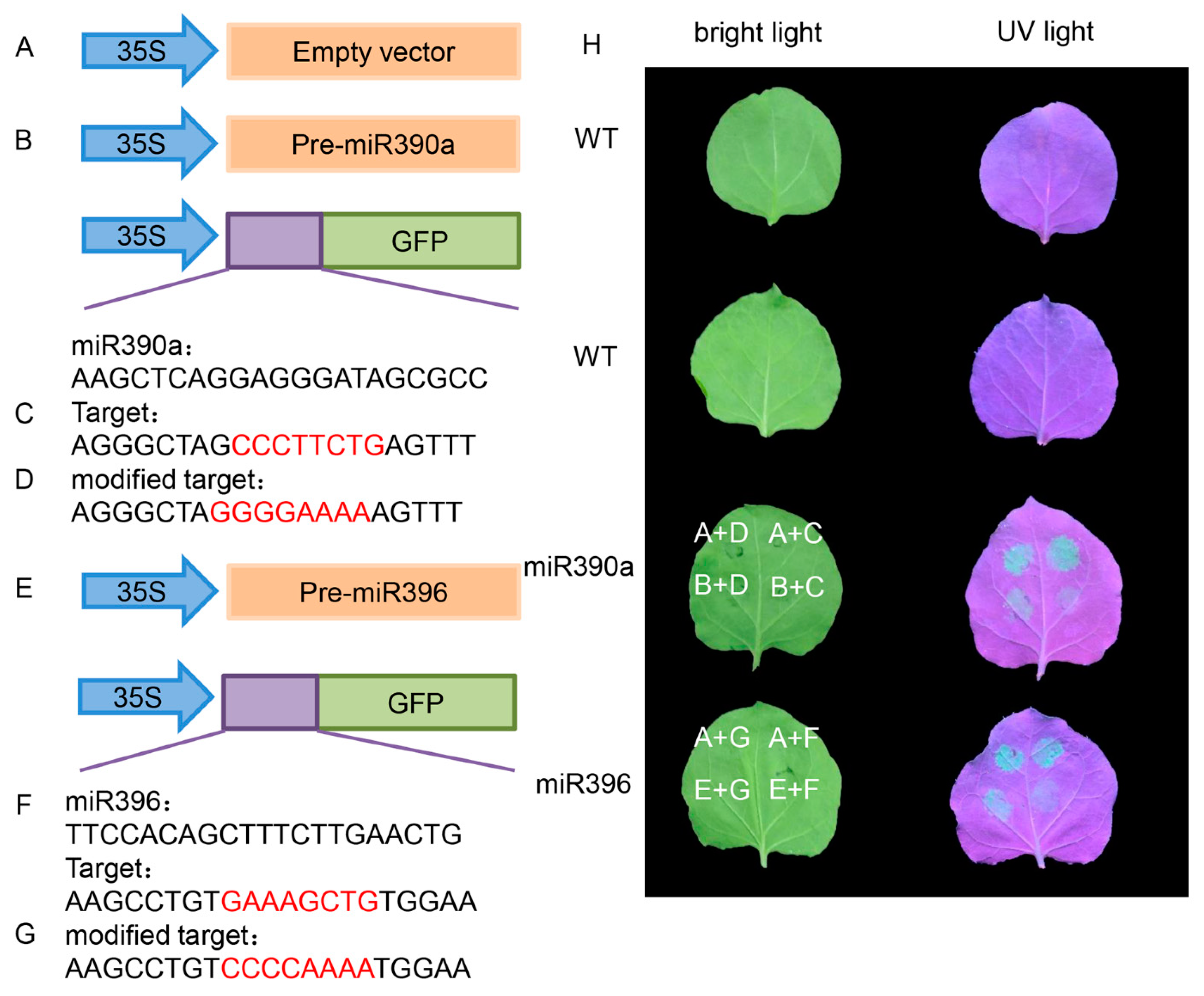
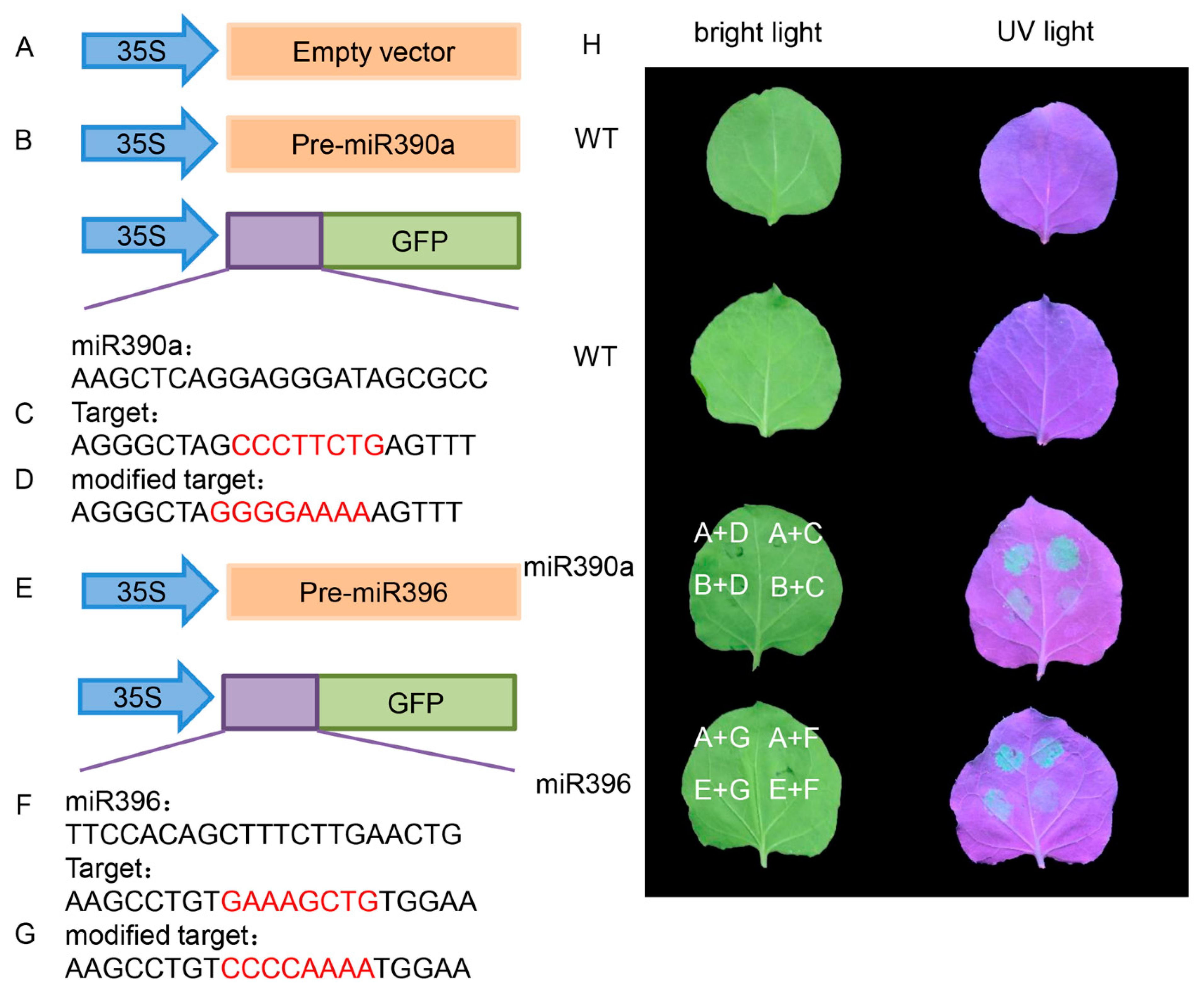
2.4. Plant Expression Vector Construction and Transformation
2.5. Statistical Analysis
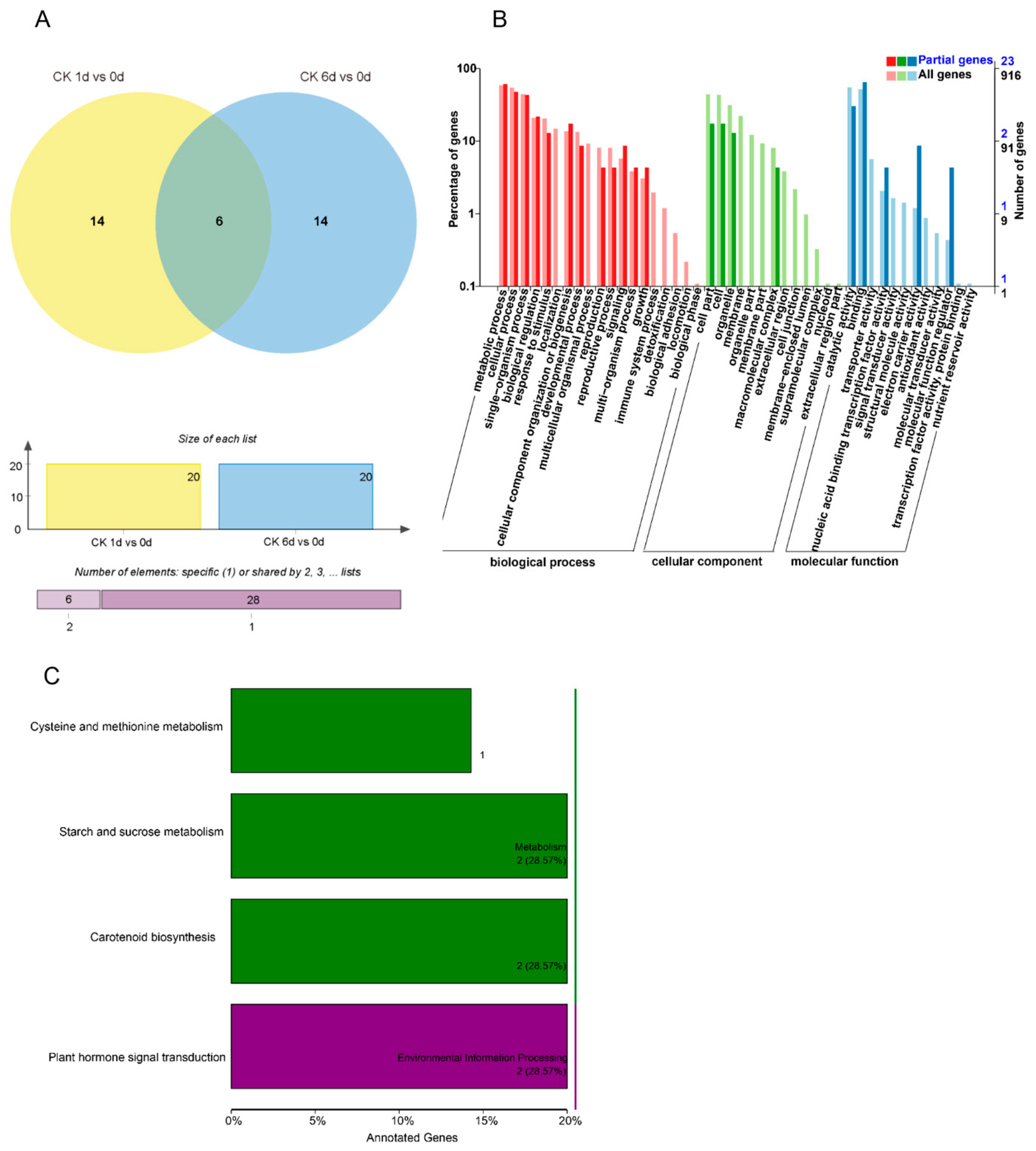
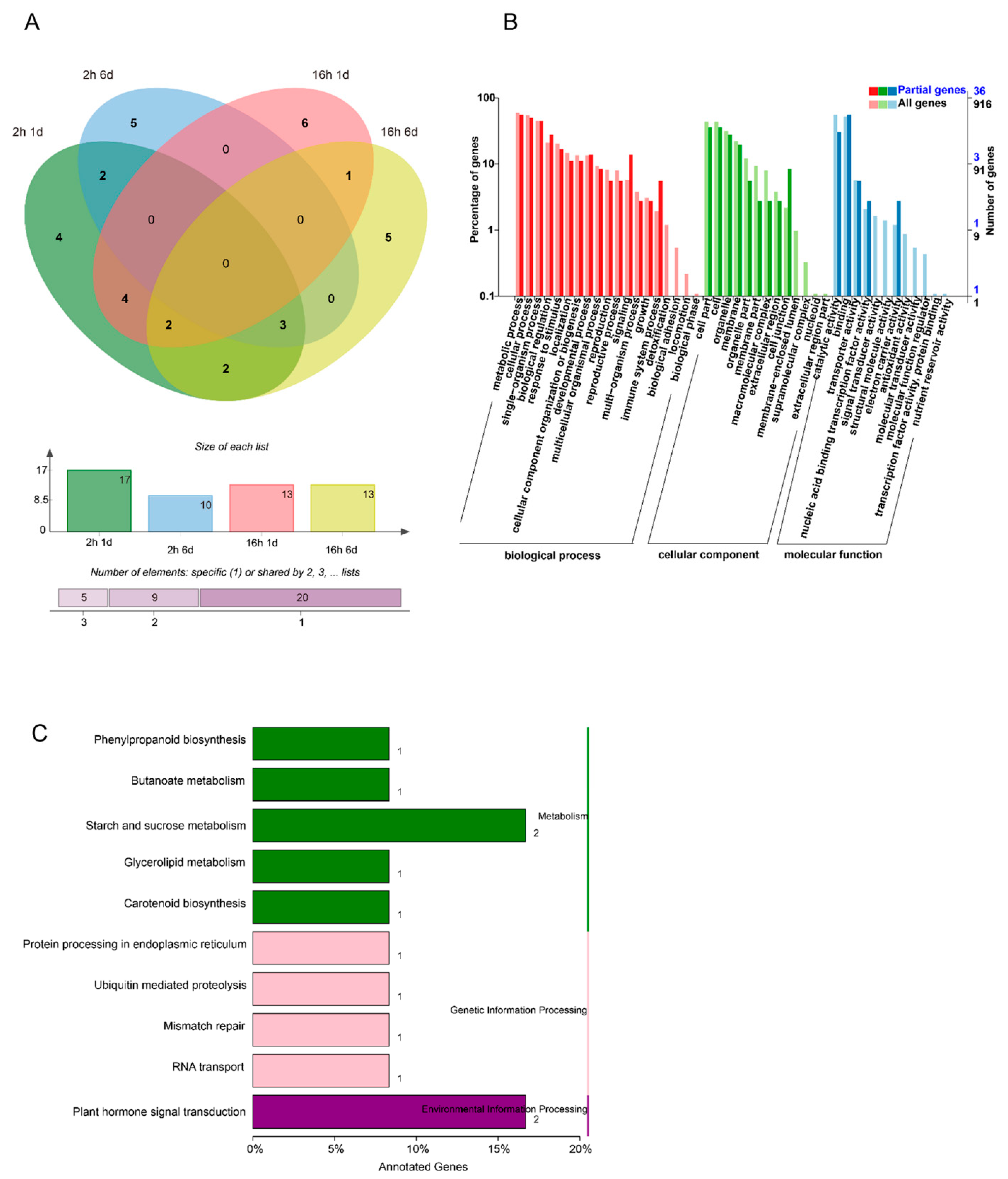
3. Results
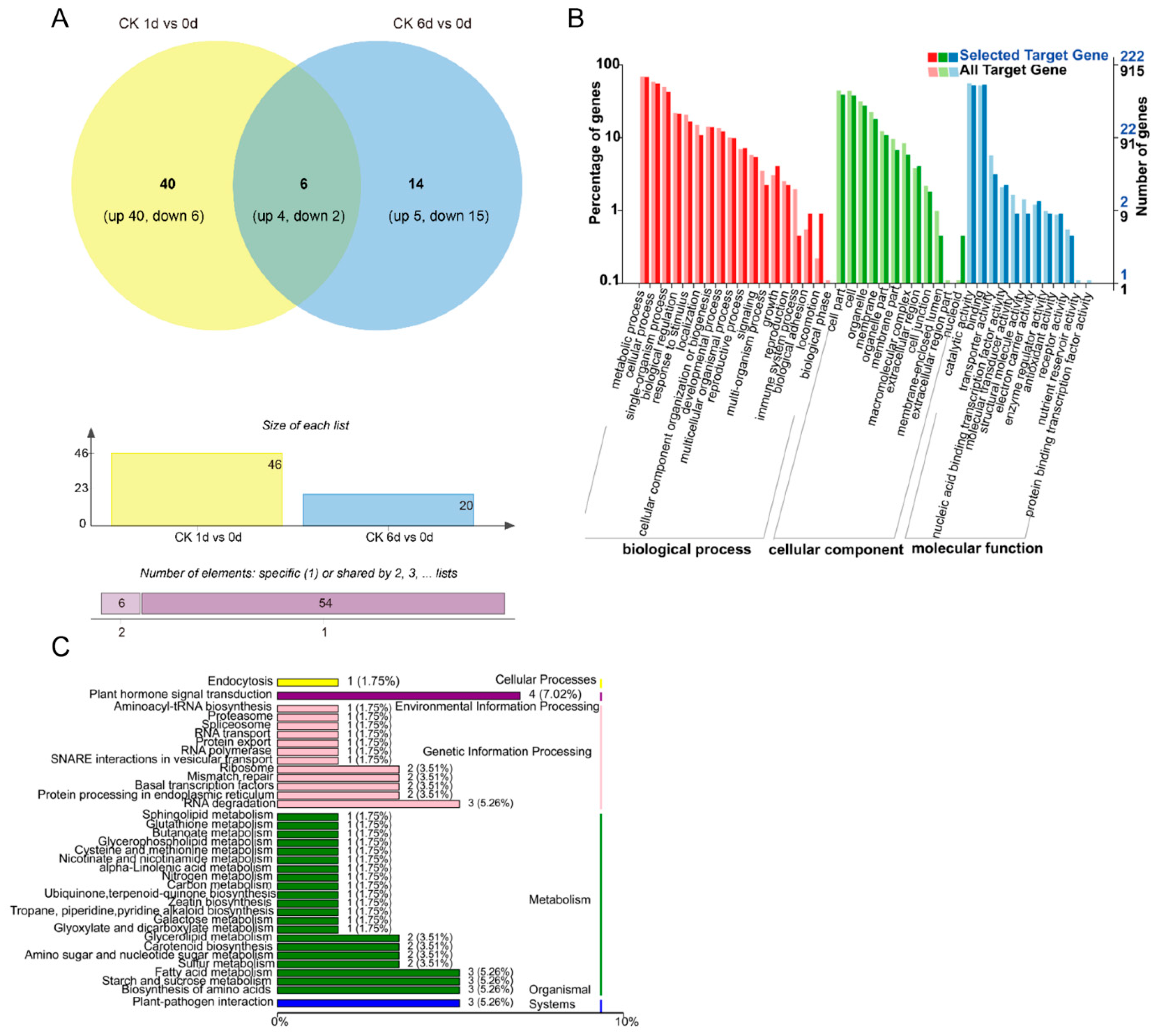
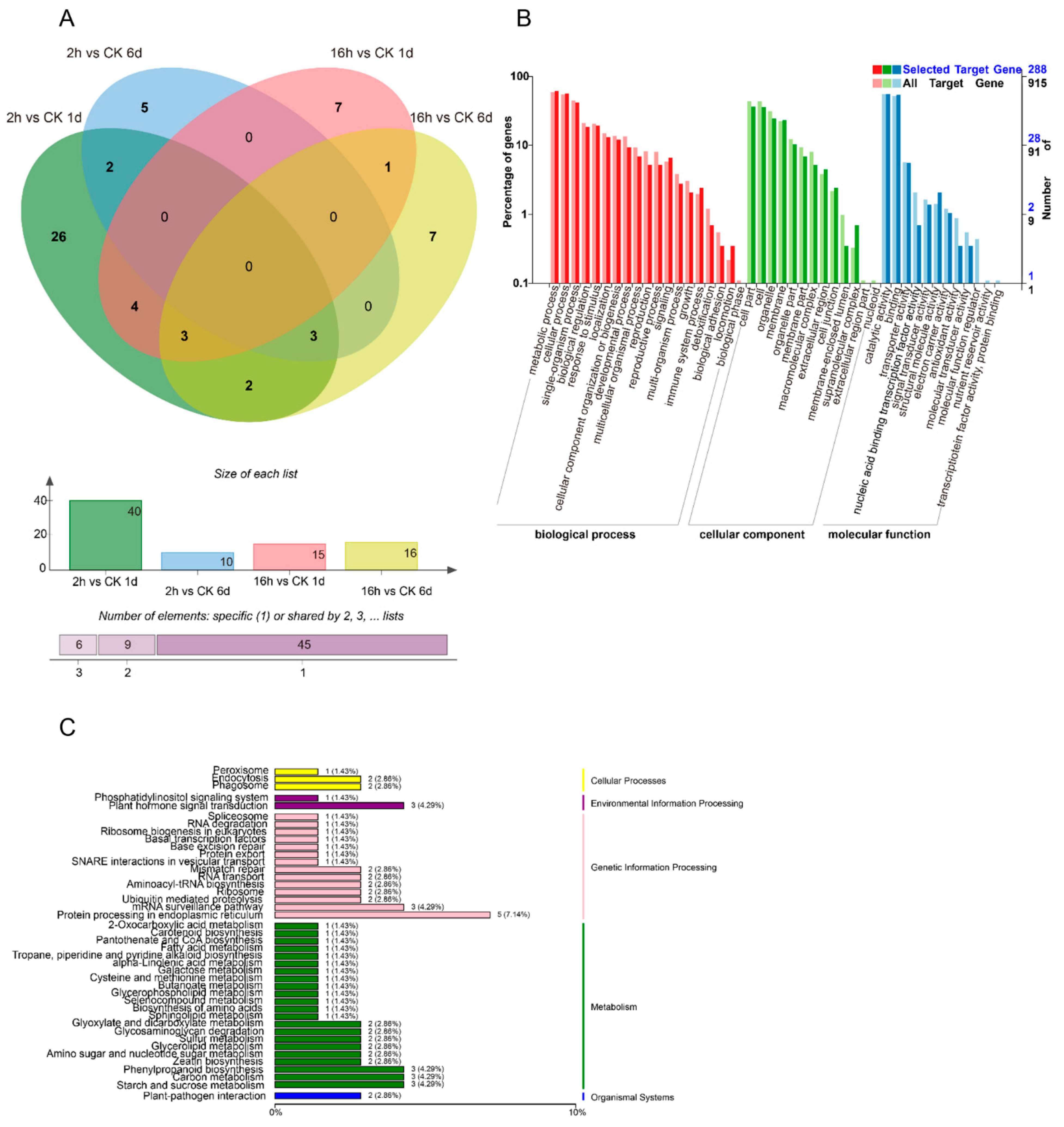
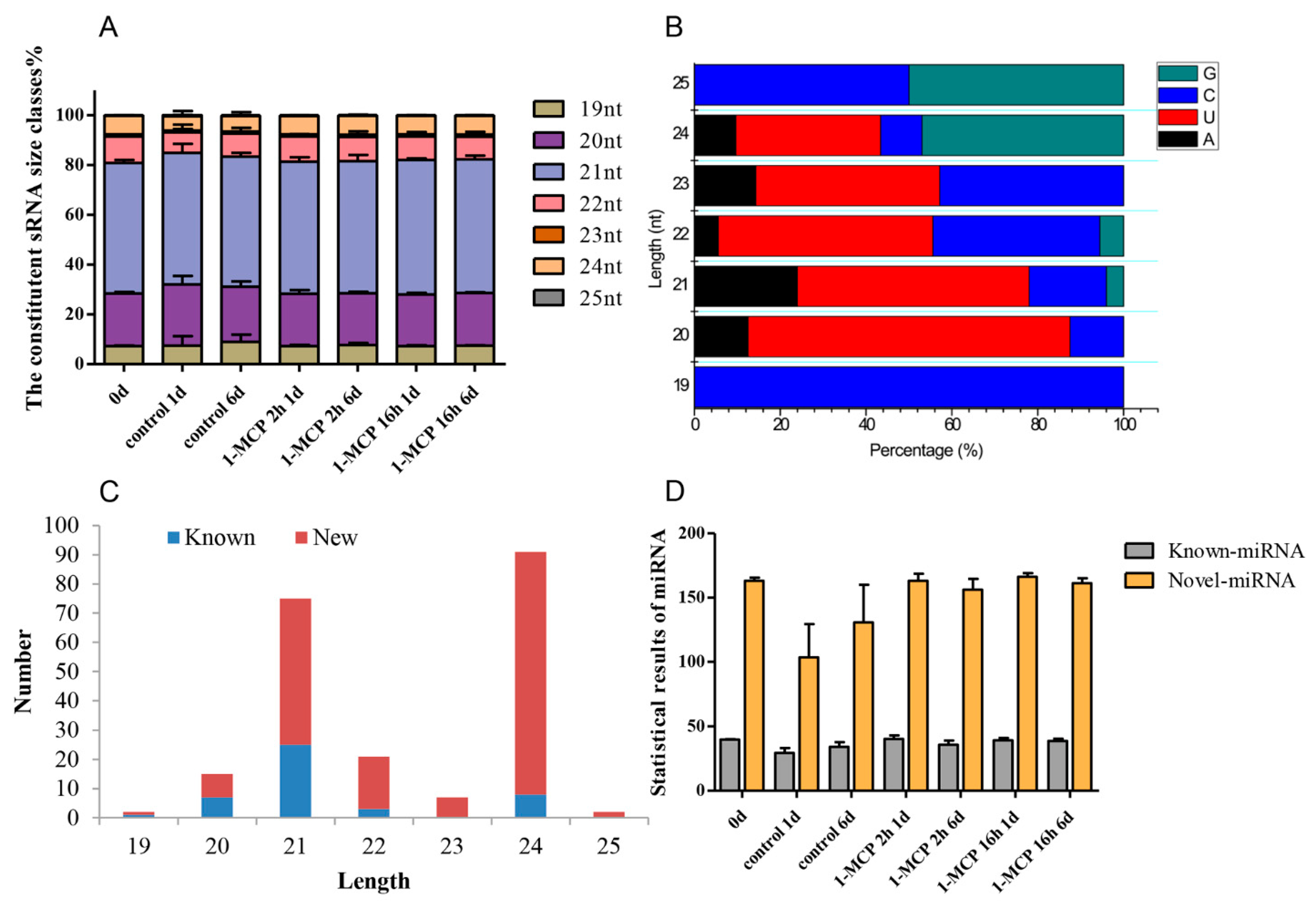
3.1. Construction and Sequencing of Small RNA Libraries
3.2. Identification of Known and Novel miRNAs
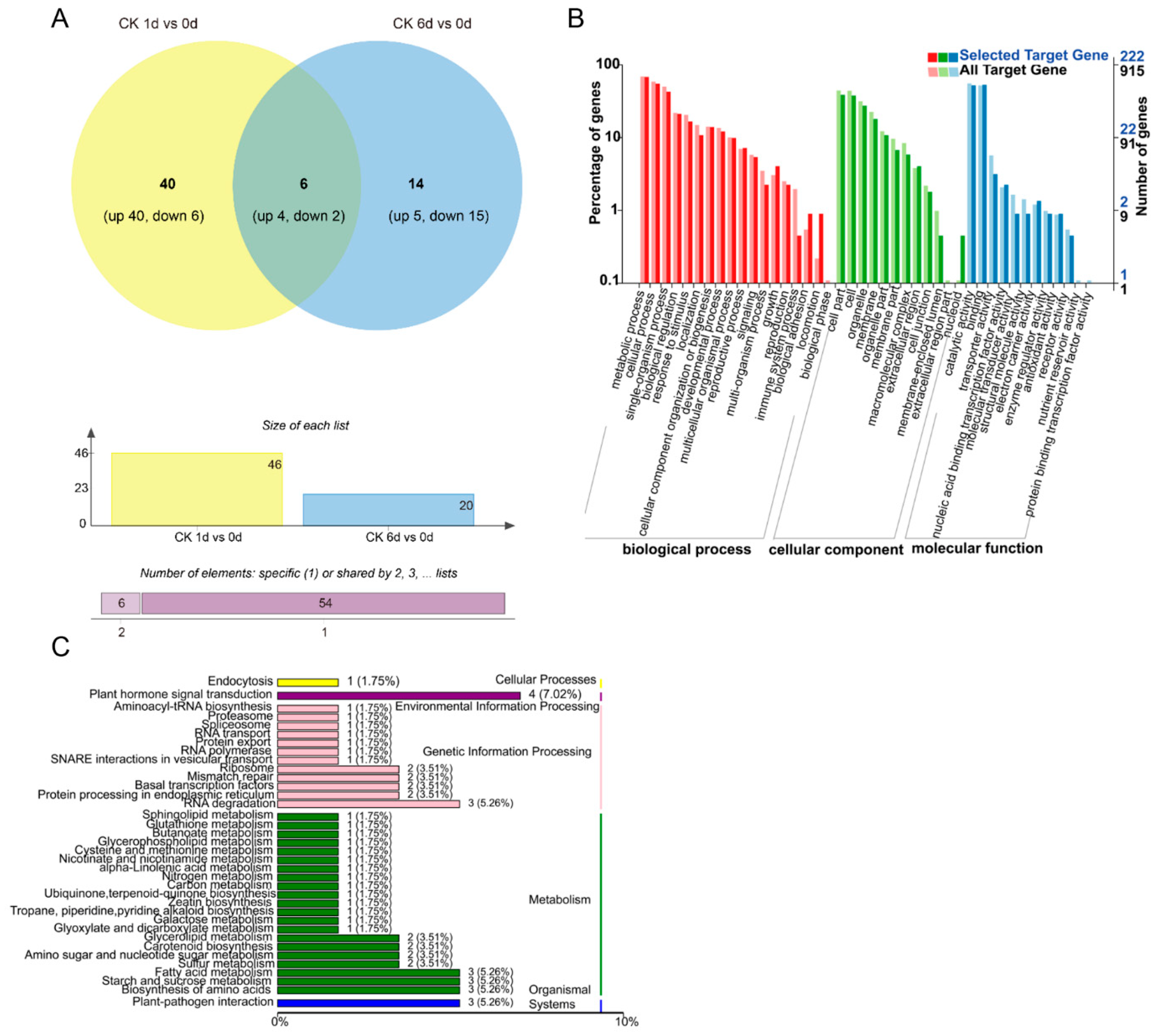
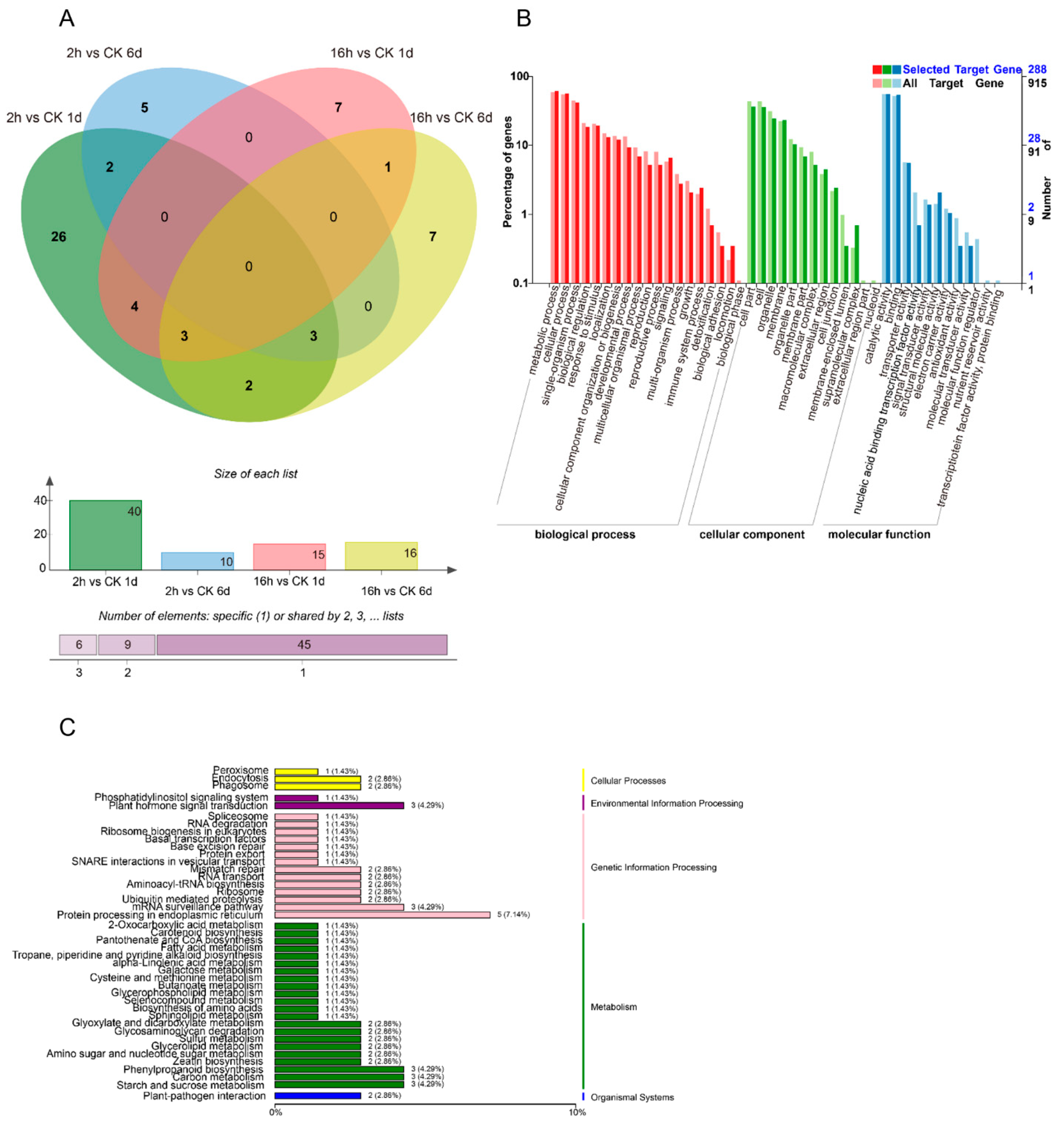
3.3. Identification of Differentially Expressed miRNAs
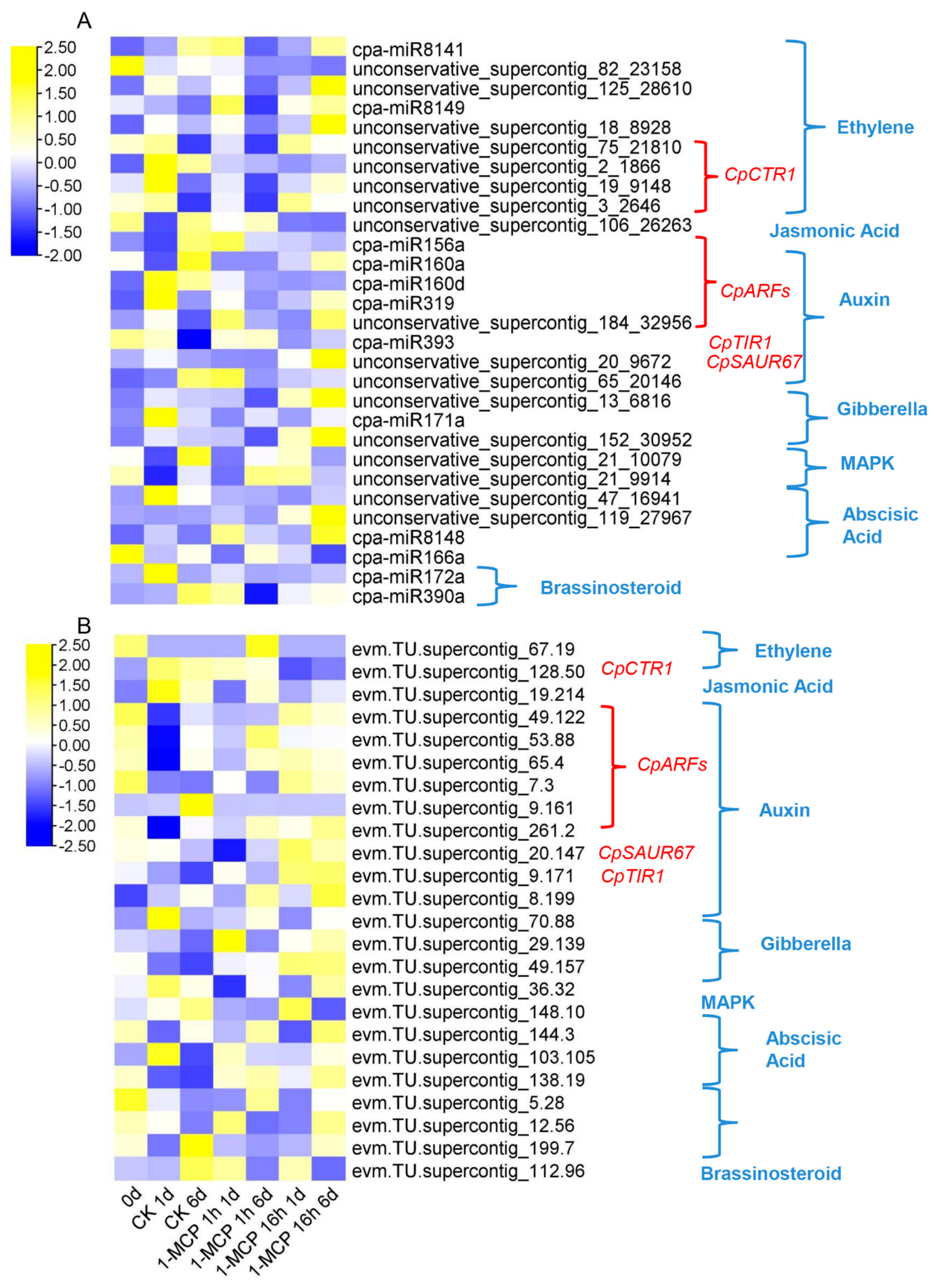
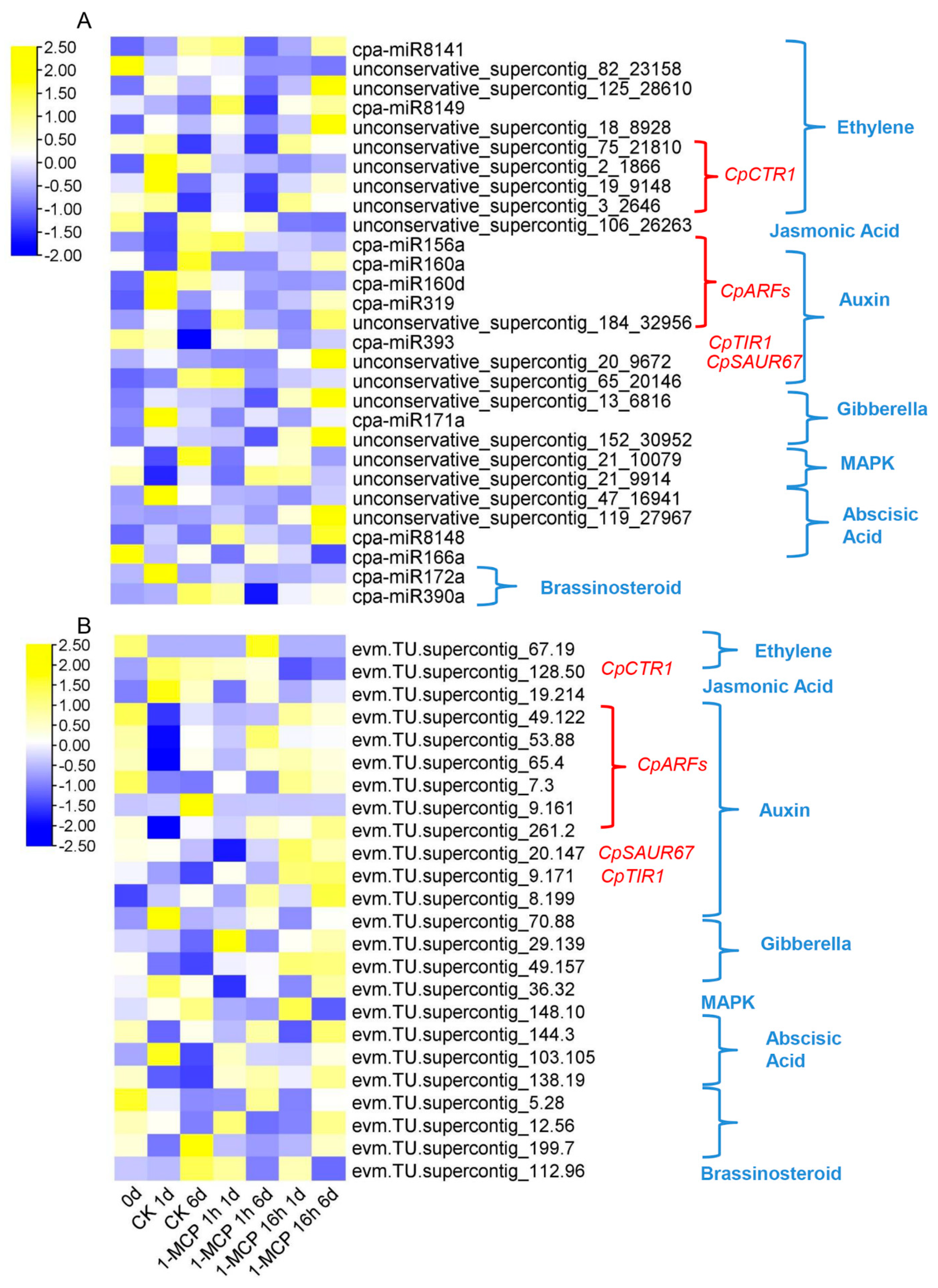
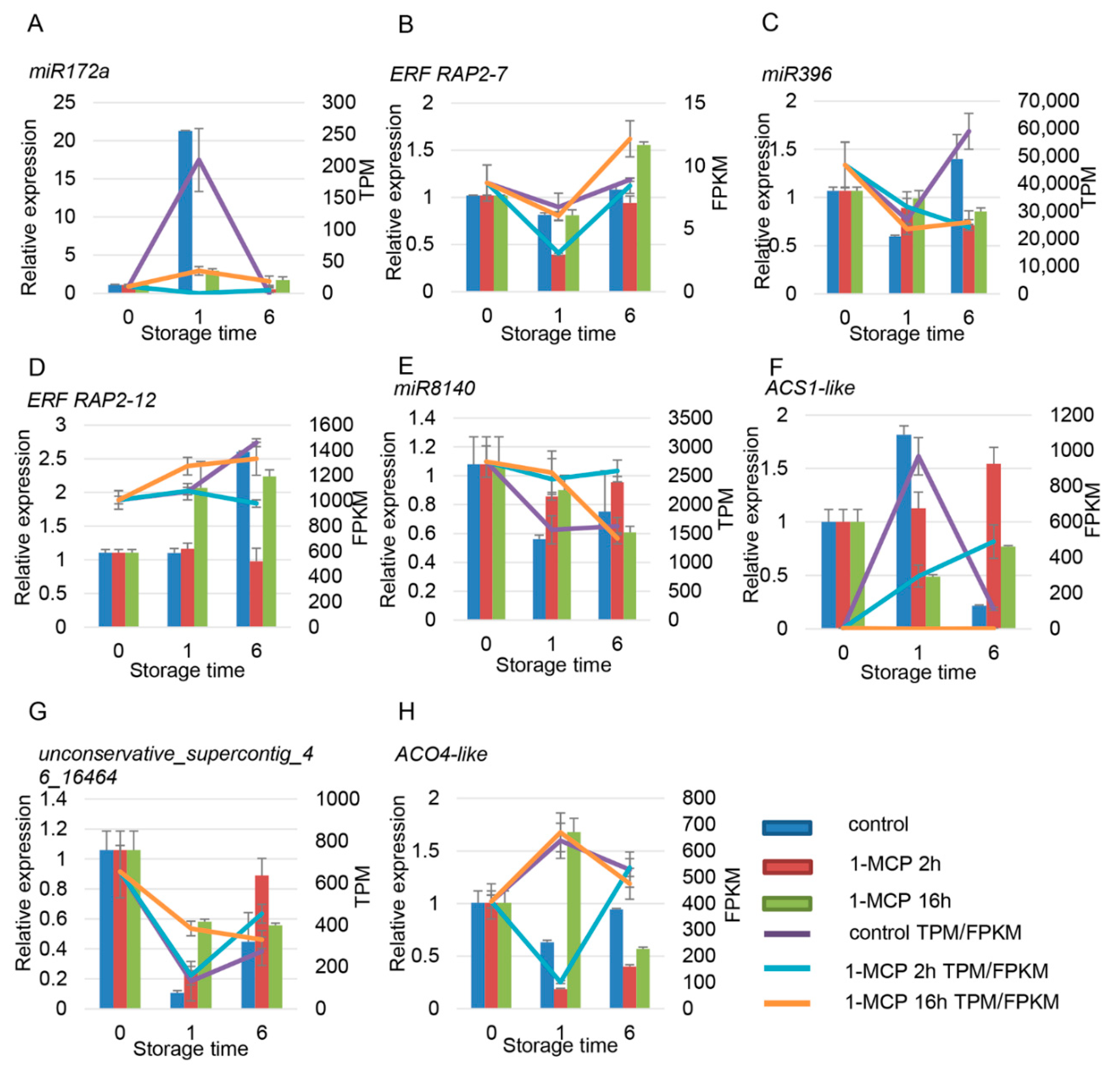
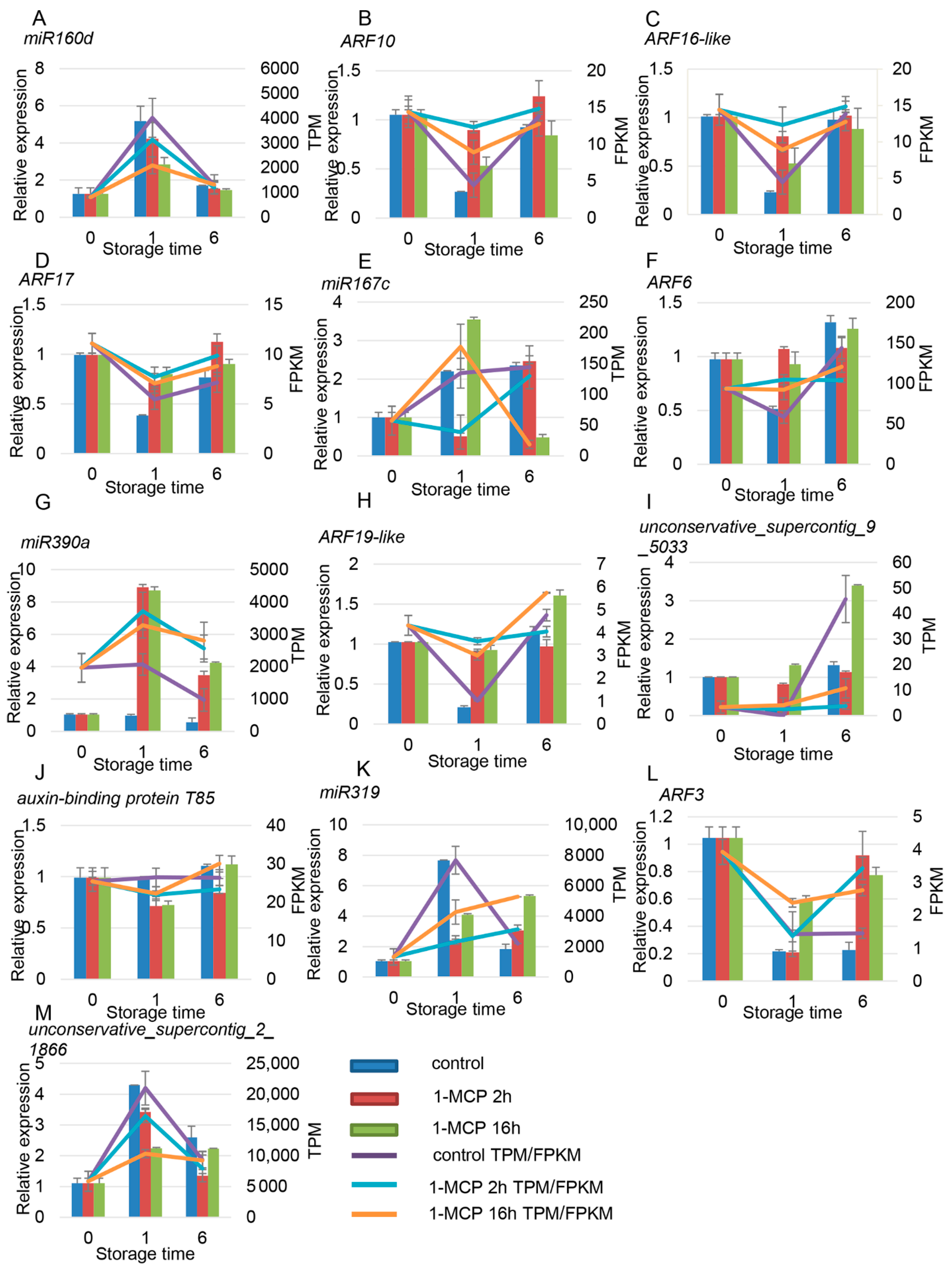
3.4. Combined Expression Analysis of miRNAs and Their Target mRNAs
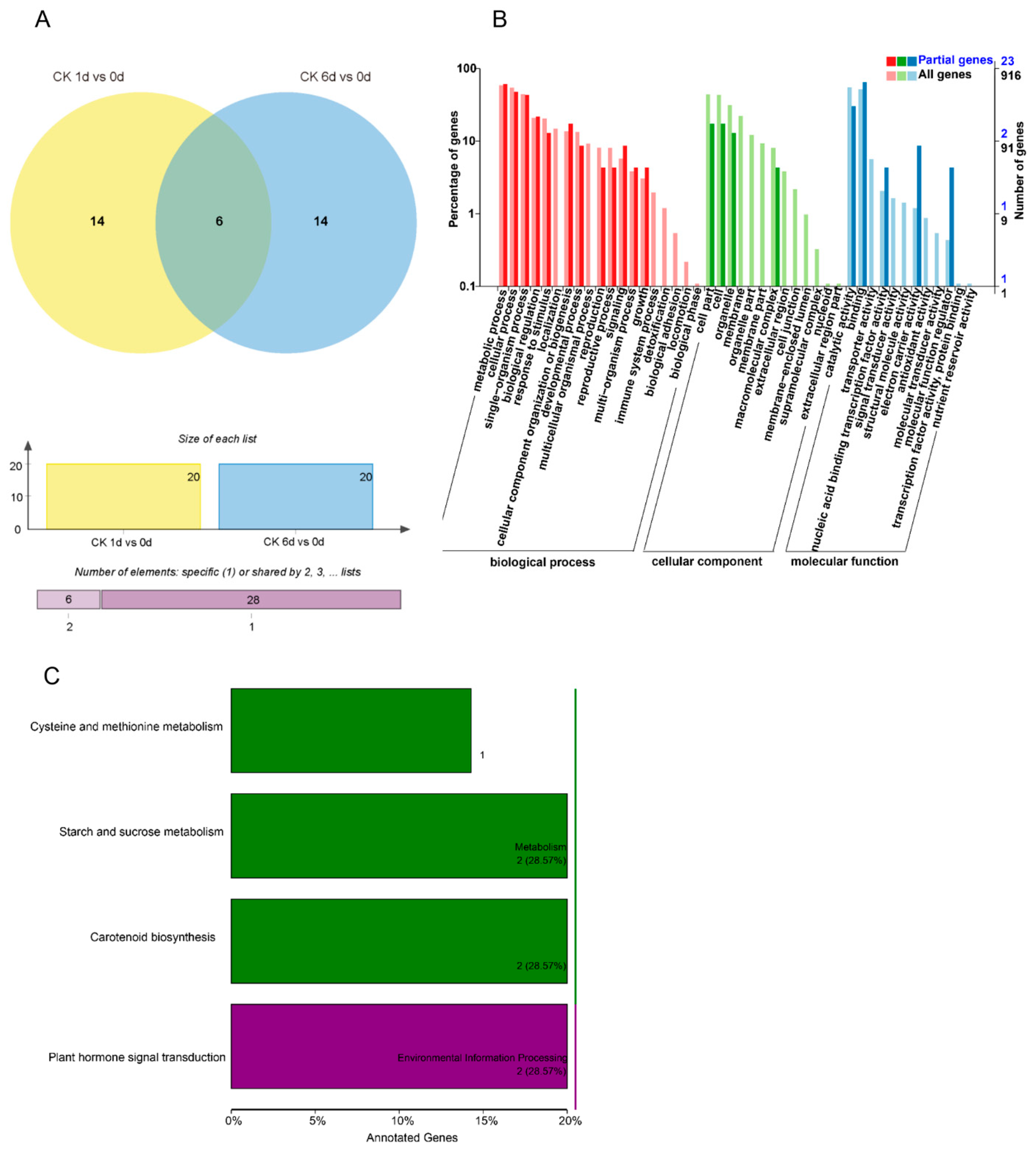
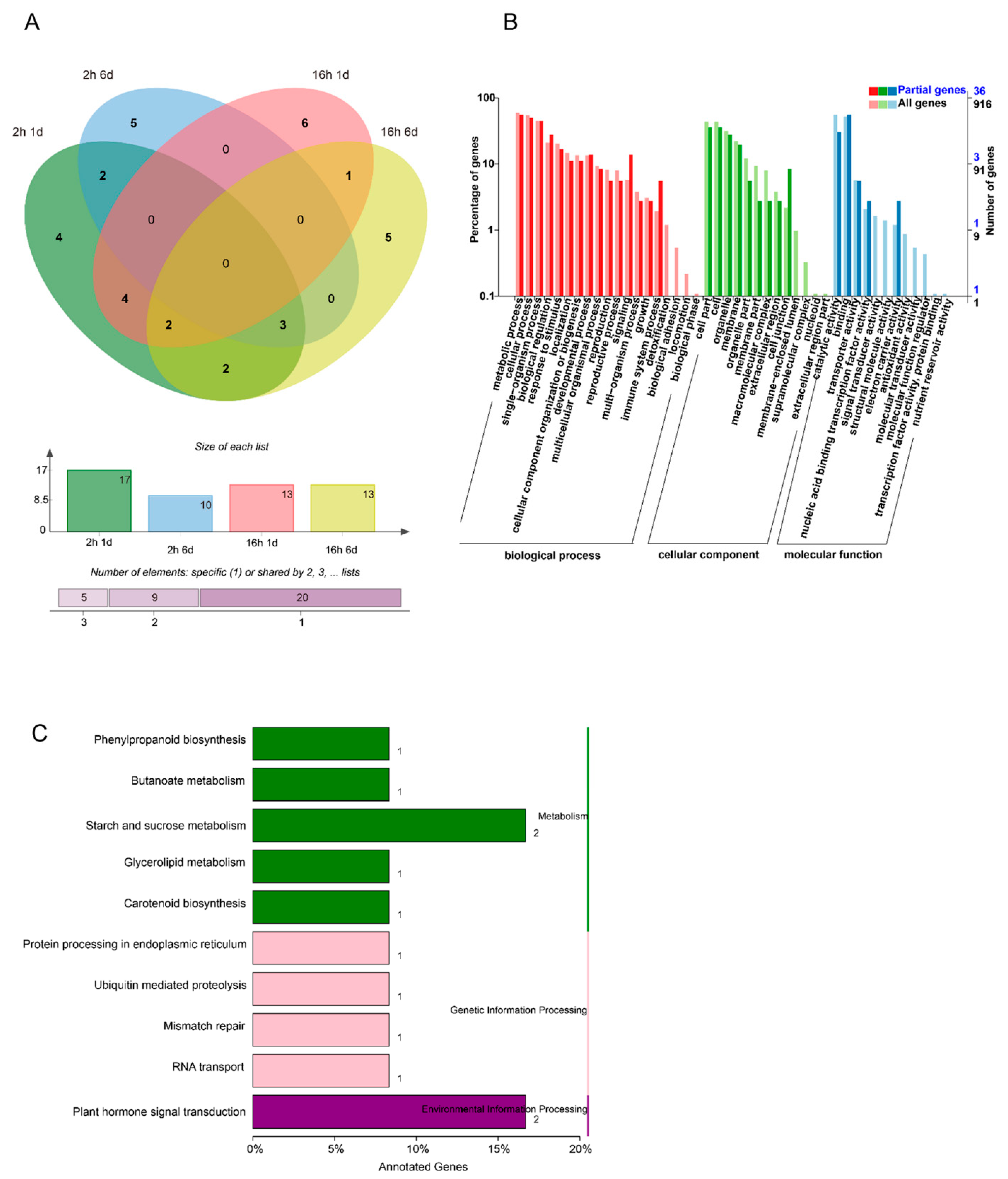
3.5. Target Gene Identification of Papaya miRNAs by Degradome Analysis
3.6. Target Gene Identification of Papaya miRNAs by Degradome Analysis
3.7. Target Gene Identification of Papaya miRNAs by Degradome Analysis
3.8. Target Gene Identification of Papaya miRNAs by Degradome Analysis
3.9. Target Gene Identification of Papaya miRNAs by Degradome Analysis
4. Discussion
4.1. Papaya miRNAs with Conserved and New Gene Targets
4.2. miRNAs Participate in Hormone Pathways during Papaya Fruit Ripening
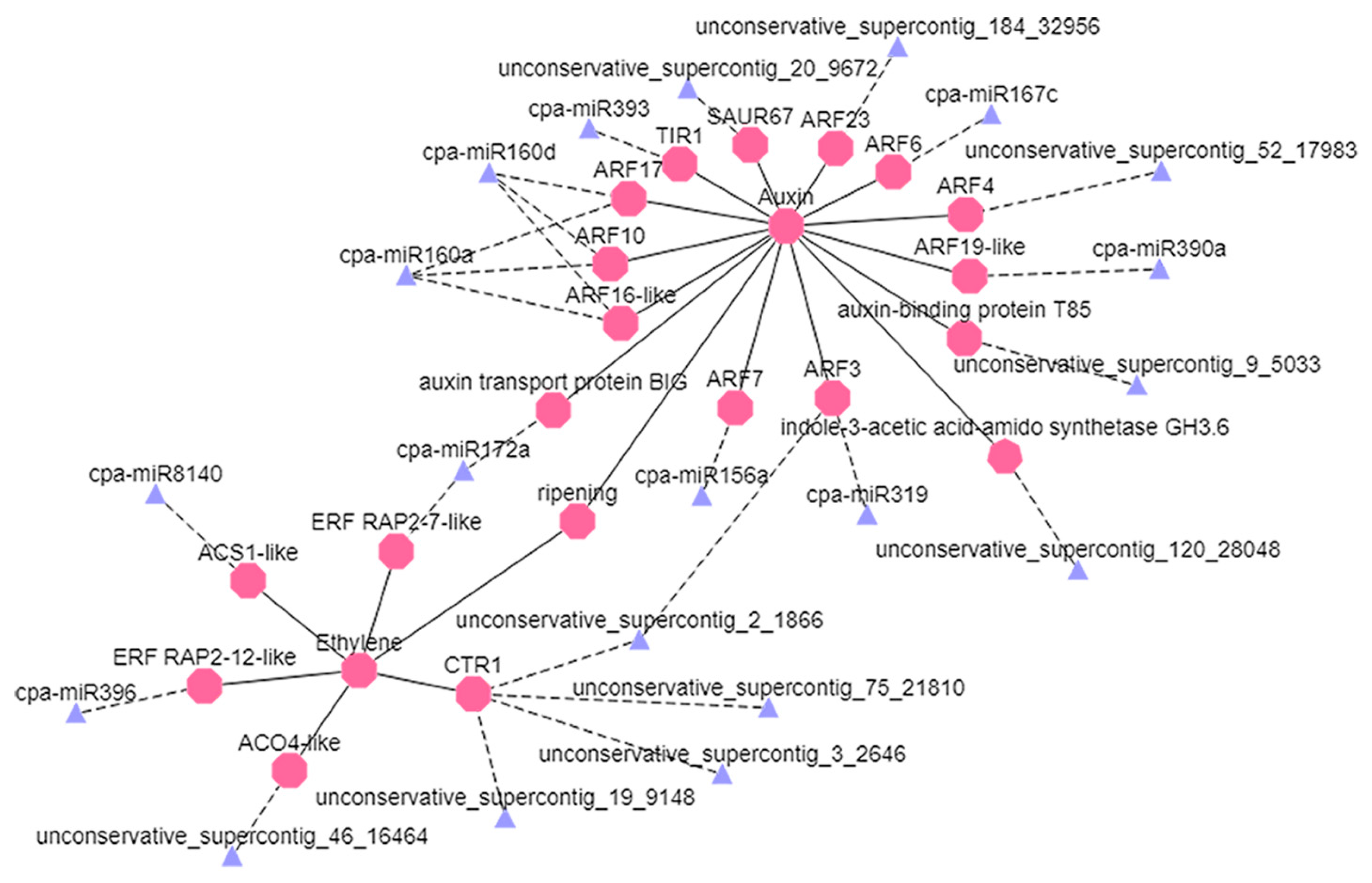
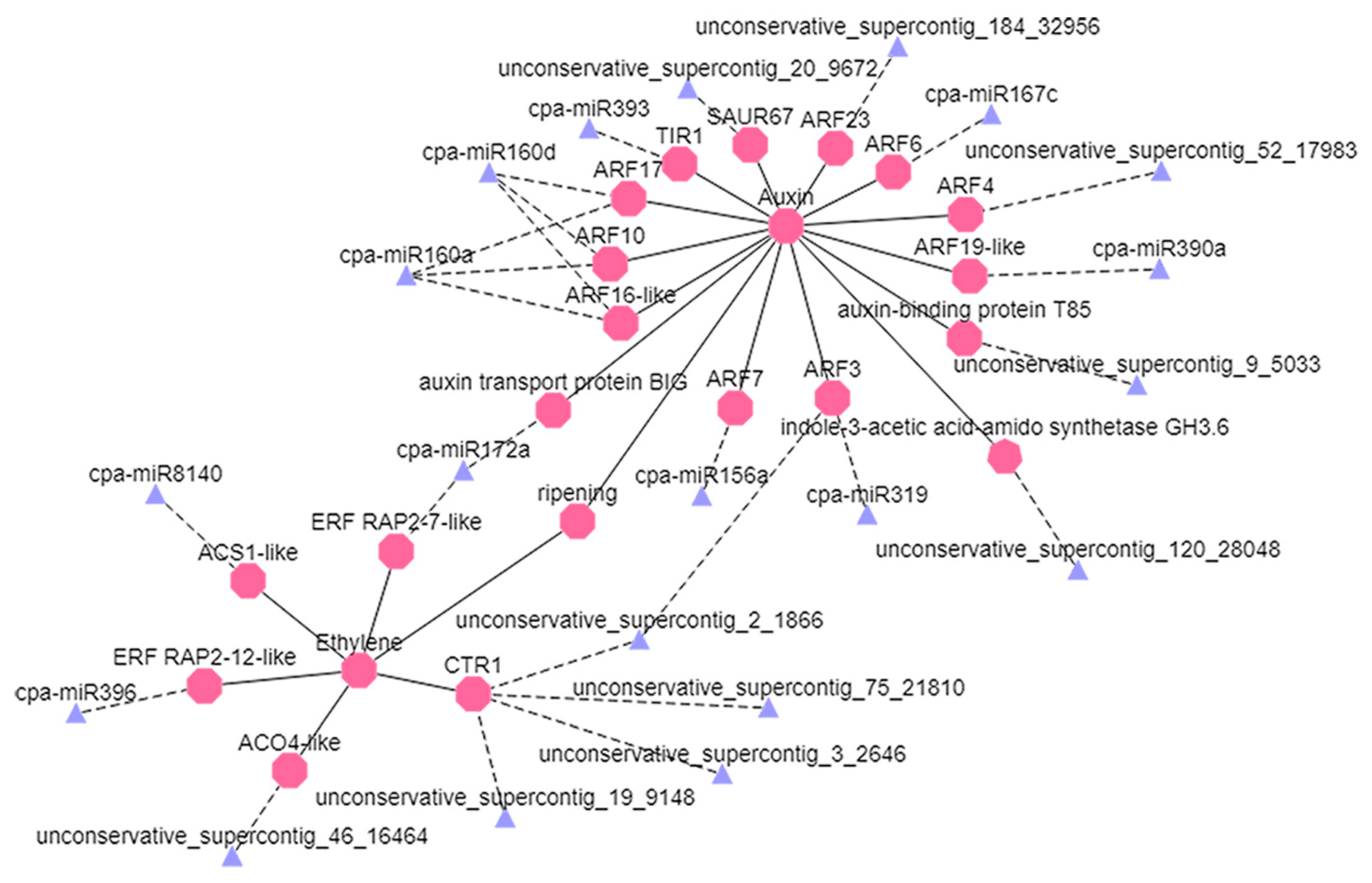
4.3. Degradation Analysis Showed That miRNAs Are Involved in Regulation of Ethylene and Auxin Signaling Pathways
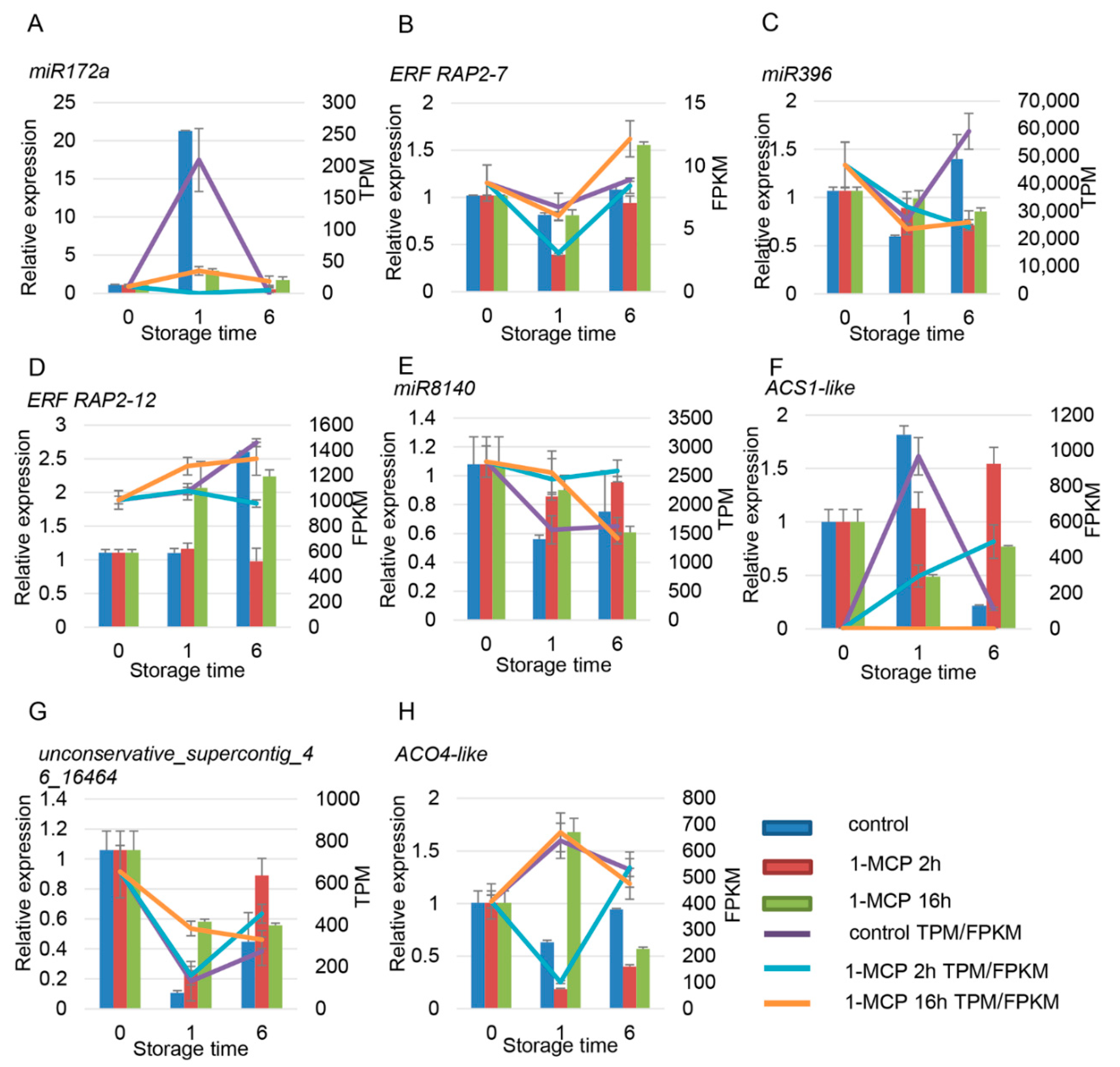
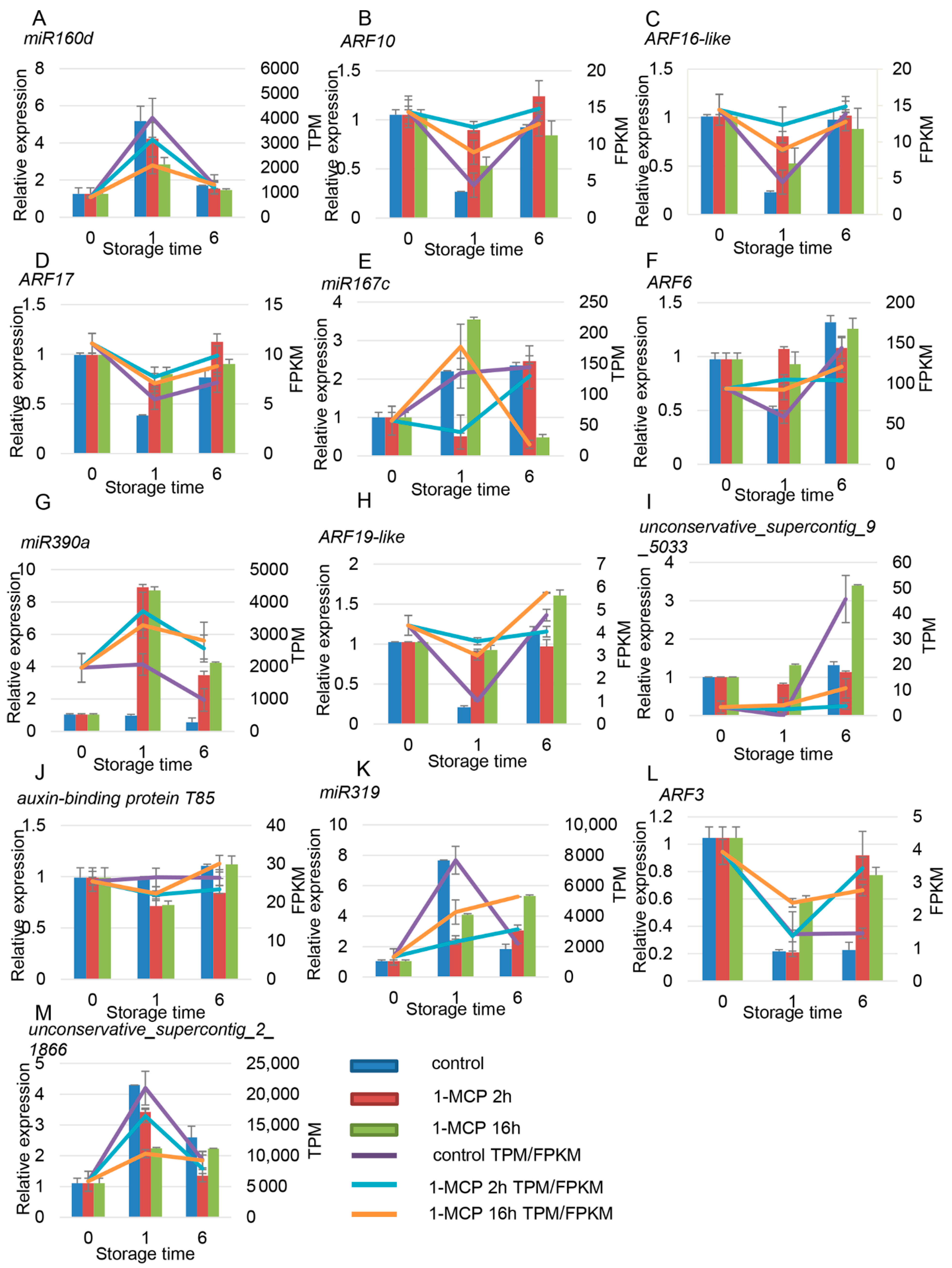
4.4. Network Regulation Diagram of miRNAs and Target Gene Verification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fabi, J.P.; Mendes, L.R.B.C.; Lajolo, F.M.; do Nascimento, J.R.O. Transcript profiling of papaya fruit reveals differentially expressed genes associated with fruit ripening. Plant Sci. 2010, 179, 225–233. [Google Scholar] [CrossRef]
- Girón-Ramírez, A.; Peña-Rodríguez, L.M.; Escalante-Erosa, F.; Fuentes, G.; Santamaría, J.M. Identification of the SHINE clade of AP2/ERF domain transcription factors genes in Carica papaya; Their gene expression and their possible role in wax accumulation and water deficit stress tolerance in a wild and a commercial papaya genotypes. Environ. Exp. Bot. 2021, 183. [Google Scholar] [CrossRef]
- Krishna, K.L.; Paridhavi, M.; Patel, J.A. Review on nutritional, medicinal and pharmacological properties of papaya (Carica papaya linn.). Indian J. Nat. Prod. Resour. 2008, 7, 364–373. [Google Scholar] [CrossRef]
- Li, X.; Zhu, X.; Mao, J.; Zou, Y.; Fu, D.; Chen, W.; Lu, W. Isolation and characterization of ethylene response factor family genes during development, ethylene regulation and stress treatments in papaya fruit. Plant Physiol. Biochem. 2013, 70, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Maringgal, B.; Hashim, N.; Tawakkal, I.S.M.A.; Mohamed, M.T.M.; Hamzah, M.H.; Ali, M.M. Effect of Kelulut honey nanoparticles coating on the changes of respiration rate, ascorbic acid, and total phenolic content of papaya (Carica papaya L.) during cold storage. Foods 2021, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.J.; Paull, R.E. Endoxylanase expressed during papaya fruit ripening: Purification, cloning and characterization. Funct. Plant Biol. 2003, 30, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Shiga, T.M.; Fabi, J.P.; Do Nascimento, J.R.O.; De Petkowicz, C.L.O.; Vriesmann, L.C.; Lajolo, F.M.; Cordenunsi, B.R. Changes in cell wall composition associated to the softening of ripening papaya: Evidence of extensive solubilization of large molecular mass galactouronides. J. Agric. Food Chem. 2009, 57, 7064–7071. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Thewes, F.R.; Anese, R.O.; Thewes, F.R.; Ludwig, V.; Klein, B.; Wagner, R.; Nora, F.R.; Rombaldi, C.V.; Brackmann, A. Dynamic controlled atmosphere (DCA) and 1-MCP: Impact on volatile esters synthesis and overall quality of ‘Galaxy’ apples. Food Packag. Shelf Life 2020, 26. [Google Scholar] [CrossRef]
- Zhu, X.; Song, Z.; Li, Q.; Li, J.; Chen, W.; Li, X. Physiological and transcriptomic analysis reveals the roles of 1-MCP in the ripening and fruit aroma quality of banana fruit (Fenjiao). Food Res. Int. 2020, 130. [Google Scholar] [CrossRef]
- Qian, C.; Ji, Z.; Zhu, Q.; Qi, X.; Li, Q.; Yin, J.; Liu, J.; Kan, J.; Zhang, M.; Jin, C.; et al. Effects of 1-mcp on proline, polyamine, and nitric oxide metabolism in postharvest peach fruit under chilling stress. Hortic. Plant J. 2021, 7, 188–196. [Google Scholar] [CrossRef]
- Du, Y.; Jin, T.; Zhao, H.; Han, C.; Sun, F.; Chen, Q.; Yue, F.; Luo, Z.; Fu, M. Synergistic inhibitory effect of 1-methylcyclopropene (1-MCP) and chlorine dioxide (ClO2) treatment on chlorophyll degradation of green pepper fruit during storage. Postharvest Biol. Technol. 2021, 171, 111363. [Google Scholar] [CrossRef]
- Ding, X.; Zhu, X.; Ye, L.; Xiao, S.; Wu, Z.; Chen, W.; Li, X. The interaction of CpEBF1 with CpMADSs is involved in cell wall degradation during papaya fruit ripening. Hortic. Res. 2019, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Hao, Y.; Fan, S.; Cai, J.; Chen, W.; Li, X.; Zhu, X. Metabolomic and transcriptomic profiling provide novel insights into fruit ripening and ripening disorder caused by 1-MCP treatments in papaya. Int. J. Mol. Sci. 2021, 22, 916. [Google Scholar] [CrossRef]
- Leticia, S.; Meza, R.; Tobaruela, E.D.C.; Pascoal, G.B.; Massaretto, I.L.; Purgatto, E. Post-harvest treatment with methyl jasmonate impacts lipid metabolism in tomato pericarp (Solanum lycopersicum L. Cv. Grape) at different ripening stages. Foods 2021, 10, 877. [Google Scholar] [CrossRef]
- Park, M.-H.; Kim, S.-J.; Lee, J.-S.; Hong, Y.-P.; Chae, S.-H.; Ku, K.-M. Carbon dioxide pretreatment and cold storage synergistically delay tomato ripening through transcriptional change in ethylene-related genes and respiration-related metabolism. Foods 2021, 10, 744. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.J.; Zhang, J.Z. Aux/IAA gene family in plants: Molecular structure, regulation, and function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Xue, J.; Li, H.; Han, S.; Jiao, J.; Rao, J. Ethylene response factors regulate ethylene biosynthesis and cell wall modification in persimmon (Diospyros kaki L.) fruit during ripening. Postharvest Biol. Technol. 2020, 168, 111255. [Google Scholar] [CrossRef]
- Fu, C.C.; Chen, H.J.; Gao, H.Y.; Wang, S.L.; Wang, N.; Jin, J.C.; Lu, Y.; Yu, Z.L.; Ma, Q.; Han, Y.C. Papaya CpMADS4 and CpNAC3 co-operatively regulate ethylene signal genes CpERF9 and CpEIL5 during fruit ripening. Postharvest Biol. Technol. 2021, 175, 111485. [Google Scholar] [CrossRef]
- Wu, Q.; Tao, X.; Ai, X.; Luo, Z.; Mao, L.; Ying, T.; Li, L. Effect of exogenous auxin on aroma volatiles of cherry tomato (Solanum lycopersicum L.) fruit during postharvest ripening. Postharvest Biol. Technol. 2018, 146, 108–116. [Google Scholar] [CrossRef]
- Khaksar, G.; Sirikantaramas, S. Auxin Response Factor 2A is part of the regulatory network mediating fruit ripening through auxin-ethylene crosstalk in durian. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Abreu, P.M.V.; Gaspar, C.G.; Buss, D.S.; Ventura, J.A.; Ferreira, P.C.G.; Fernandes, P.M.B. Carica papaya microRNAs are responsive to Papaya meleira virus infection. PLoS ONE 2014, 9, e103401. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Lei, M.; Li, Z.; Jing, Y.; Hu, H.; Zhu, S.; Xu, L. Small RNA sequencing revealed various microRNAs involved in ethylene-triggered flowering process in Aechmea fasciata. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Wu, J.; Zhao, X.; Hao, M.; Geng, S.; Yan, J.; Jiang, X.; Zhang, L.; Wu, J.; et al. mRNA and small RNA transcriptomes reveal insights into dynamic homoeolog regulation of allopolyploid heterosis in nascent hexaploid wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [Green Version]
- Pagano, L.; Rossi, R.; Paesano, L.; Marmiroli, N.; Marmiroli, M. miRNA regulation and stress adaptation in plants. Environ. Exp. Bot. 2021, 184, 104369. [Google Scholar] [CrossRef]
- Das, B.; Sen, A.; Roy, S.; Banerjee, O.; Bhattacharya, S. miRNAs: Tiny super-soldiers shaping the life of rice plants for facing “stress”-ful times. Plant Gene 2021, 26, 100281. [Google Scholar] [CrossRef]
- Li, K.; Liu, Z.; Xing, L.; Wei, Y.; Mao, J.; Meng, Y.; Bao, L.; Han, M.; Zhao, C.; Zhang, D. miRNAs associated with auxin signaling, stress response, and cellular activities mediate adventitious root formation in apple rootstocks. Plant Physiol. Biochem. 2019, 139, 66–81. [Google Scholar] [CrossRef]
- Zhang, T.; Li, W.; Xie, R.; Xu, L.; Zhou, Y.; Li, H.; Yuan, C.; Zheng, X.; Xiao, L.; Liu, K. CpARF2 and CpEIL1 interact to mediate auxin–ethylene interaction and regulate fruit ripening in papaya. Plant J. 2020, 103, 1318–1337. [Google Scholar] [CrossRef] [PubMed]
- Gidhi, A.; Kumar, M.; Mukhopadhyay, K. The auxin response factor gene family in wheat (Triticum aestivum L.): Genome-wide identification, characterization and expression analyses in response to leaf rust. South Afr. J. Bot. 2020, 000. [Google Scholar] [CrossRef]
- Dan, M.; Huang, M.; Liao, F.; Qin, R.; Liang, X.; Zhang, E.; Huang, M.; Huang, Z.; He, Q. Identification of ethylene responsive mirnas and their targets from newly harvested banana fruits using high-throughput sequencing. J. Agric. Food Chem. 2018, 66, 10628–10639. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Ye, L.; Ding, X.; Gao, Q.; Xiao, S.; Tan, Q.; Huang, J.; Chen, W.; Li, X. Transcriptomic analysis reveals key factors in fruit ripening and rubbery texture caused by 1-MCP in papaya. BMC Plant Biol. 2019, 19, 1–22. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Chen, D.; Wu, X.; Huang, D.; Chen, L.; Li, L.; Deng, X.; Xu, Q. Genome-wide comparison of microRNAs and their targeted transcripts among leaf, flower and fruit of sweet orange. BMC Genom. 2014, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Li, X.; Chen, W.; Chen, J.; Lu, W.; Chen, L.; Fu, D. Evaluation of new reference genes in papaya for accurate transcript normalization under different experimental conditions. PLoS ONE 2012, 7, e44405. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, H.; Wall, M.M.; Yang, J. Roles of transcription factor SQUAMOSA promoter binding protein-like gene family in papaya (Carica papaya) development and ripening. Genomics 2020, 112, 2734–2747. [Google Scholar] [CrossRef]
- Wang, R.; Fang, Y.N.; Wu, X.M.; Qing, M.; Li, C.C.; Xie, K.D.; Deng, X.X.; Guo, W.W. The miR399-CsUBC24 module regulates reproductive development and male fertility in citrus. Plant Physiol. 2020, 183, 1681–1695. [Google Scholar] [CrossRef] [PubMed]
- Aryal, R.; Yang, X.; Yu, Q.; Sunkar, R.; Li, L.; Ming, R. Asymmetric purine-pyrimidine distribution in cellular small RNA population of papaya. BMC Genom. 2012, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.M.; Wu, M.C.; Wu, C.L.; Chang, H.M. Pectinesterase and polygalacturonase activities and textural properties of rubbery papaya (Carica papaya Linn.). J. Food Sci. 2003, 68, 1590–1594. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, L.; Hao, Y.; Xiao, S.; Wu, Z.; Chen, W.; Li, X.; Zhu, X. Genome-wide identification and expression analyses of the calmodulin and calmodulin-like proteins reveal their involvement in stress response and fruit ripening in papaya. Postharvest Biol. Technol. 2018, 143, 13–27. [Google Scholar] [CrossRef]
- Façanha, R.V.; Spricigo, P.C.; Purgatto, E.; Jacomino, A.P. Combined application of ethylene and 1-methylcyclopropene on ripening and volatile compound production of “Golden” papaya. Postharvest Biol. Technol. 2019, 151, 160–169. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Gao, L.; Zhu, B.; Ju, Z.; Luo, Y.; Zuo, J. Parsing the regulatory network between small RNAs and target genes in ethylene pathway in tomato. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Sun, F.; Chen, N.; Sun, G.; Wang, C.Y.; Wu, D.X. MiR396 regulatory network and its expression during grain development in wheat. Protoplasma 2021, 258, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Du, C.; Gao, H.; Liu, S.; Sun, W.; Lu, H.; Kang, J.; Xie, Y.; Ma, D.; Wang, C. Identification of microRNAs in developing wheat grain that are potentially involved in regulating grain characteristics and the response to nitrogen levels. BMC Plant Biol. 2020, 20, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Akdogan, G.; Tufekci, E.D.; Uranbey, S.; Unver, T. miRNA-based drought regulation in wheat. Funct. Integr. Genom. 2016, 16, 221–233. [Google Scholar] [CrossRef]
- Ragupathy, R.; Ravichandran, S.; Mahdi, M.S.R.; Huang, D.; Reimer, E.; Domaratzki, M.; Cloutier, S. Deep sequencing of wheat sRNA transcriptome reveals distinct temporal expression pattern of miRNAs in response to heat, light and UV. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Zhang, R.; Zhang, S.; Li, Y.; Gao, J.; Han, X.; Chen, M.; Wang, J.; Li, W.; Li, G. Response of microRNAs to cold treatment in the young spikes of common wheat. BMC Genom. 2017, 18, 212. [Google Scholar] [CrossRef] [Green Version]
- Luan, Y.; Cui, J.; Li, J.; Jiang, N.; Liu, P.; Meng, J. Effective enhancement of resistance to Phytophthora infestans by overexpression of miR172a and b in Solanum lycopersicum. Planta 2018, 247, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Grierson, D.; Courtney, L.T.; Wang, Y.; Gao, L.; Zhao, X.; Zhu, B.; Luo, Y.; Wang, Q.; Giovannoni, J.J. Relationships between genome methylation, levels of non-coding RNAs, mRNAs and metabolites in ripening tomato fruit. Plant J. 2020, 103, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Zhu, B.; Fu, D.; Zhu, Y.; Ma, Y.; Chi, L.; Ju, Z.; Wang, Y.; Zhai, B.; Luo, Y. Sculpting the maturation, softening and ethylene pathway: The influences of microRNAs on tomato fruits. BMC Genom. 2012, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Yue, R.; Sun, T.; Zhang, L.; Xu, L.; Tie, S.; Wang, H.; Yang, Y. Genome-wide identification and expression analysis of auxin response factor gene family in Medicago truncatula. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Yuan, C.; Feng, S.; Zhong, S.; Li, H.; Zhong, J.; Shen, C.; Liu, J. Genome-wide analysis and characterization of Aux/IAA family genes related to fruit ripening in papaya (Carica papaya L.). BMC Genom. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Z.; Qiao, M.; Xiang, F. miR393 inhibits in vitro shoot regeneration in Arabidopsis thaliana via repressing TIR1. Plant Sci. 2018, 266, 1–8. [Google Scholar] [CrossRef]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis Auxin Response Factor17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azahar, I.; Ghosh, S.; Adhikari, A.; Adhikari, S.; Roy, D.; Shaw, A.K.; Singh, K.; Hossain, Z. Comparative analysis of maize root sRNA transcriptome unveils the regulatory roles of miRNAs in submergence stress response mechanism. Environ. Exp. Bot. 2020, 171, 103924. [Google Scholar] [CrossRef]
- Jin, L.; Yarra, R.; Zhou, L.; Zhao, Z.; Cao, H. miRNAs as key regulators via targeting the phytohormone signaling pathways during somatic embryogenesis of plants. 3 Biotech 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | miR_Regulate | Predicted Target Gene ID | Predicted Target Gene Name | |
|---|---|---|---|---|
| CK 1 d vs. 0 d | cpa-miR319 | Down | evm.TU.supercontig_7.3 | Carica papaya auxin response factor 3 |
| cpa-miR396 | Up | evm.TU.supercontig_48.26 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | |
| cpa-miR396 | Up | evm.TU.supercontig_481.1 | Carica papaya ethylene-responsive transcription factor RAP2-12-like | |
| cpa-miR8140 | Up | evm.TU.supercontig_1322.1 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | |
| CK 6 d vs. 0 d | cpa-miR160d | Down | evm.TU.supercontig_49.122 | Carica papaya auxin response factor 17 |
| cpa-miR160d | Down | evm.TU.supercontig_53.88 | Carica papaya auxin response factor 16-like | |
| cpa-miR160d | Down | evm.TU.supercontig_65.4 | Carica papaya auxin response factor 10 | |
| 2 h 6 d vs. CK 6 d | cpa-miR390a | Up | evm.TU.supercontig_261.2 | Carica papaya auxin response factor 19-like |
| 16 h 1 d vs. CK 1 d | cpa-miR172a | Up | C.papaya_newGene_850 | Carica papaya auxin transport protein BIG |
| cpa-miR172a | Up | evm.TU.supercontig_1.271 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | |
| cpa-miR172a | Up | evm.TU.supercontig_114.55 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | |
| 16 h 6 d vs. CK 6 d | cpa-miR160d | Up | evm.TU.supercontig_49.122 | Carica papaya auxin response factor 17 |
| cpa-miR160d | Up | evm.TU.supercontig_53.88 | Carica papaya auxin response factor 16-like | |
| cpa-miR160d | Up | evm.TU.supercontig_65.4 | Carica papaya auxin response factor 10 | |
| unconservative_supercontig_2_1866 | Up | evm.TU.supercontig_7.3 | Carica papaya auxin response factor 3 | |
| 2h 6 d vs. 1 d | cpa-miR172a | Up | C.papaya_newGene_850 | Carica papaya auxin transport protein BIG |
| cpa-miR172a | Up | evm.TU.supercontig_1.271 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | |
| cpa-miR172a | Up | evm.TU.supercontig_114.55 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | |
| cpa-miR396 | Down | evm.TU.supercontig_48.26 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | |
| cpa-miR396 | Down | evm.TU.supercontig_481.1 | Carica papaya ethylene-responsive transcription factor RAP2-12-like | |
| 16 h 6 d vs. 1 d | cpa-miR167c | Up | evm.TU.supercontig_17.52 | Carica papaya auxin response factor 6 |
| cpa-miR8140 | Up | evm.TU.supercontig_1322.1 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like |
| miRNA | miR_Regulate | Target Gene ID | Target Gene Name | Target Gene Regulation | |
|---|---|---|---|---|---|
| CK 1 d vs. 0 d | cpa-miR396 | Up | evm.TU.supercontig_48.26 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | Down |
| cpa-miR396 | Up | evm.TU.supercontig_481.1 | Carica papaya ethylene-responsive transcription factor RAP2-12-like | Down | |
| cpa-miR8140 | Up | evm.TU.supercontig_1322.1 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | Down | |
| unconservative_supercontig_120_28048 | Up | evm.TU.contig_32826 | Carica papaya indole-3-acetic acid-amido synthetase GH3.6 | Down | |
| unconservative_supercontig_46_16464 | Up | evm.TU.supercontig_2.209 | Carica papaya 1-aminocyclopropane-1-carboxylate oxidase homolog 4-like | Down | |
| cpa-miR319 | Down | evm.TU.supercontig_7.3 | Carica papaya auxin response factor 3 | Up | |
| CK 6 d vs. 0 d | unconservative_supercontig_46_16464 | Up | evm.TU.supercontig_2.209 | Carica papaya 1-aminocyclopropane-1-carboxylate oxidase homolog 4-like | Down |
| cpa-miR160d | Down | evm.TU.supercontig_49.122 | Carica papaya auxin response factor 17 | Up | |
| cpa-miR160d | Down | evm.TU.supercontig_53.88 | Carica papaya auxin response factor 16-like | Up | |
| cpa-miR160d | Down | evm.TU.supercontig_65.4 | Carica papaya auxin response factor 10 | Up | |
| unconservative_supercontig_2_1866 | Down | evm.TU.supercontig_7.3 | Carica papaya auxin response factor 3 | Up | |
| 2h 6 d vs. CK 6 d | cpa-miR390a | Up | evm.TU.supercontig_261.2 | Carica papaya auxin response factor 19-like | Down |
| 16 h 1 d vs. CK 1 d | cpa-miR172a | Up | C.papaya_newGene_850 | Carica papaya auxin transport protein BIG | Down |
| cpa-miR172a | Up | evm.TU.supercontig_1.271 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | Down | |
| cpa-miR172a | Up | evm.TU.supercontig_114.55 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | Down | |
| cpa-miR172a | Up | evm.TU.supercontig_139.43 | Carica papaya ethylene-responsive transcription factor RAP2-7 | Down | |
| unconservative_supercontig_120_28048 | Down | evm.TU.contig_32826 | Carica papaya indole-3-acetic acid-amido synthetase GH3.6 | Up | |
| unconservative_supercontig_52_17983 | Up | evm.TU.supercontig_83.80 | Carica papaya ethylene-responsive transcription factor 4 | Down | |
| 16 h 6 d vs. CK 6 d | cpa-miR160d | Up | evm.TU.supercontig_49.122 | Carica papaya auxin response factor 17 | Down |
| cpa-miR160d | Up | evm.TU.supercontig_53.88 | Carica papaya auxin response factor 16-like | Down | |
| cpa-miR160d | Up | evm.TU.supercontig_65.4 | Carica papaya auxin response factor 10 | Down | |
| unconservative_supercontig_2_1866 | Up | evm.TU.supercontig_7.3 | Carica papaya auxin response factor 3 | Down | |
| 2 h 6 d vs. 1 d | cpa-miR172a | Up | C.papaya_newGene_850 | Carica papaya auxin transport protein BIG | Down |
| cpa-miR172a | Up | evm.TU.supercontig_1.271 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | Down | |
| cpa-miR172a | Up | evm.TU.supercontig_114.55 | Carica papaya ethylene-responsive transcription factor RAP2-7-like | Down | |
| cpa-miR172a | Up | evm.TU.supercontig_139.43 | Carica papaya ethylene-responsive transcription factor RAP2-7 | Down | |
| cpa-miR396 | Down | evm.TU.supercontig_48.26 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | Up | |
| cpa-miR396 | Down | evm.TU.supercontig_481.1 | Carica papaya ethylene-responsive transcription factor RAP2-12-like | Up | |
| unconservative_supercontig_9_5033 | Down | evm.TU.supercontig_9.126 | Carica papaya auxin-binding protein T85 | Up | |
| 16 h 6 d vs. 1 d | cpa-miR167c | Up | evm.TU.supercontig_17.52 | Carica papaya auxin response factor 6 | Down |
| cpa-miR8140 | Up | evm.TU.supercontig_1322.1 | Carica papaya 1-aminocyclopropane-1-carboxylate synthase 1-like | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, J.; Wu, Z.; Hao, Y.; Liu, Y.; Song, Z.; Chen, W.; Li, X.; Zhu, X. Small RNAs, Degradome, and Transcriptome Sequencing Provide Insights into Papaya Fruit Ripening Regulated by 1-MCP. Foods 2021, 10, 1643. https://doi.org/10.3390/foods10071643
Cai J, Wu Z, Hao Y, Liu Y, Song Z, Chen W, Li X, Zhu X. Small RNAs, Degradome, and Transcriptome Sequencing Provide Insights into Papaya Fruit Ripening Regulated by 1-MCP. Foods. 2021; 10(7):1643. https://doi.org/10.3390/foods10071643
Chicago/Turabian StyleCai, Jiahui, Ziling Wu, Yanwei Hao, Yuanlong Liu, Zunyang Song, Weixin Chen, Xueping Li, and Xiaoyang Zhu. 2021. "Small RNAs, Degradome, and Transcriptome Sequencing Provide Insights into Papaya Fruit Ripening Regulated by 1-MCP" Foods 10, no. 7: 1643. https://doi.org/10.3390/foods10071643
APA StyleCai, J., Wu, Z., Hao, Y., Liu, Y., Song, Z., Chen, W., Li, X., & Zhu, X. (2021). Small RNAs, Degradome, and Transcriptome Sequencing Provide Insights into Papaya Fruit Ripening Regulated by 1-MCP. Foods, 10(7), 1643. https://doi.org/10.3390/foods10071643