
Effect of Dietary Seaweed Supplementation in Cows on Milk Macrominerals, Trace Elements and Heavy Metal Concentrations

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Analysis of Milk and Feed for Chemical Composition
2.3. Quantification of Mineral Concentrations in Milk and Feed
2.4. Statistical Analysis
3. Results
3.1. Animal and Diet Parameters
3.2. Milk Yield, Basic Composition, and Efficiency
3.3. Milk Mineral Concentrations
3.4. Estimated Mineral Transfer Efficiencies from Feed to Milk
4. Discussion
4.1. Effect of Seaweed Supplementation on Milk Yield, Basic Composition and Efficiency Parameters
4.2. Effect of Seaweed on Milk Mineral Concentrations and Estimated Mineral Transfer Efficiencies from Feed to Milk
4.2.1. Trace Elements
4.2.2. Heavy Metals
4.3. Nutritional Implications of Milk from Seaweed-Fed Cows for Consumers (I, Cu, Se, As)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roque, B.M.; Brooke, C.G.; Ladau, J.; Polley, T.; Marsh, L.J.; Najafi, N.; Pandey, P.; Singh, L.; Kinley, R.; Salwen, J.K.; et al. Effect of the macroalgae Asparagopsis taxiformis on methane production and rumen microbiome assemblage. Anim. Microbiome 2019, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from Seaweeds: An Ocean of Opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed. Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Gegg, P.; Wells, V. UK Macro-Algae Biofuels: A Strategic Management Review and Future Research Agenda. J. Mar. Sci. Eng. 2017, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Ferdouse, F.L.H.; Susan, L.; Smith, R.; Murúa, P.; Yang, Z. The Global Status of Seaweed Production, Trade and Utilization; Food and Agriculture Organization of the UN: Rome, Italy, 2018; p. 120. [Google Scholar]
- Saker, K.E.; Fike, J.H.; Veit, H.; Ward, D.L. Brown seaweed- (TascoTM) treated conserved forage enhances antioxidant status and immune function in heat-stressed wether lambs. J. Anim. Physiol. Anim. Nutr. 2004, 88, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Braden, K.W.; Blanton, J.R., Jr.; Allen, V.G.; Pond, K.R.; Miller, M.F. Ascophyllum nodosum supplementation: A preharvest intervention for reducing Escherichia coli O157:H7 and Salmonella spp. in feedlot steers. J. Food Prot. 2004, 67, 1824–1828. [Google Scholar] [CrossRef]
- Antaya, N.T.; Ghelichkhan, M.; Pereira, A.B.D.; Soder, K.J.; Brito, A.F. Production, milk iodine, and nutrient utilization in Jersey cows supplemented with the brown seaweed Ascophyllum nodosum (kelp meal) during the grazing season. J. Dairy Sci. 2019, 102, 8040–8058. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; Robertson, R.C.; Stanton, C.; O’Doherty, J.V.; Kerry, J.P. Influence of level and duration of feeding polysaccharide (laminarin and fucoidan) extracts from brown seaweed (Laminaria digitata) on quality indices of fresh pork. Meat Sci. 2015, 99, 132–141. [Google Scholar] [CrossRef]
- de la Moneda, A.; Carro, M.D.; Weisbjerg, M.R.; Roleda, M.Y.; Lind, V.; Novoa-Garrido, M.; Molina-Alcaide, E. Variability and Potential of Seaweeds as Ingredients of Ruminant Diets: An In Vitro Study. Animals 2019, 9, 851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belanche, A.; Jones, E.; Parveen, I.; Newbold, C.J. A Metagenomics Approach to Evaluate the Impact of Dietary Supplementation with Ascophyllum nodosum or Laminaria digitata on Rumen Function in Rusitec Fermenters. Front. Microbiol. 2016, 7, 299. [Google Scholar] [CrossRef]
- Buschmann, A.; Camus, C.; Infante Rosselot, J.; Neori, A.; Israel, A.; Hernández-González, M.; Pereda, S.; Gomez Pinchetti, J.L.; Golberg, A.; Tadmor Shalev, N.; et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Ugarte, R.; Sharp, G. Management and production of the brown algae Ascophyllum nodosum in the Canadian maritimes. J. Appl. Phycol. 2012, 24, 409–416. [Google Scholar] [CrossRef]
- Capuzzo, E.; McKie, T. Seaweed in the UK and aborad—Status, Products, Limitations, Gaps and Cefas Role; Centre for Environment, Fisheries & Aquacultural Science: Suffolk, UK, 2016. [Google Scholar]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Borges, D.; Araujo, R.; Azevedo, I.; Pinto, I.S. Sustainable management of economically valuable seaweed stocks at the limits of their range of distribution: Ascophyllum nodosum (Phaeophyceae) and its southernmost population in Europe. J. Appl. Phycol. 2020, 32, 1365–1375. [Google Scholar] [CrossRef]
- Edwards, M.; Watson, L. Aquaculture Explained: Cultivating Laminaria Digitata; BIM Bord Iascaigh Mhara, Irish Sea Fisheries Board: Dublin, Ireland, 2015. [Google Scholar]
- Davoult, D.; Engel, C.; Arzel, P.; Knoch, D.; Laurans, M. Environmental factors and commercial harvesting: Exploring possible links behind the decline of the kelp Laminaria digitata in Brittany, France. Cah. Biol. Mar. 2011, 52, 429–435. [Google Scholar]
- Brantsæter, A.L.; Knutsen, H.; Johansen, N.; Nyheim, K.; Erlund, I.; Meltzer, H.; Henjum, S. Inadequate Iodine Intake in Population Groups Defined by Age, Life Stage and Vegetarian Dietary Practice in a Norwegian Convenience Sample. Nutrients 2018, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatzia, M.; Christaki, E.; Bonos, E.; Karatzias, C.; Florou-Paneri, P. The influence of dietary Ascophyllum nodosum on haematologic parameters of dairy cows. Ital. J. Anim. Sci. 2012, 11, e31. [Google Scholar] [CrossRef]
- Rayman, M.; Bath, S. The new emergence of iodine deficiency in the UK: Consequences for child neurodevelopment. Ann. Clin. Biochem. 2015, 52, 705–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey-Crespo, F.; López-Alonso, M.; Miranda, M. The use of seaweed from the Galician coast as a mineral supplement in organic dairy cattle. Animal 2014, 8, 580–586. [Google Scholar] [CrossRef]
- Zhou, M.; Hünerberg, M.; Chen, Y.; Reuter, T.; McAllister, T.A.; Evans, F.; Critchley, A.T.; Guan, L.L. Air-dried brown seaweed, Ascophyllum nodosum, alters the rumen microbiome in a manner that changes rumen fermentation profiles and lowers the prevalence of foodborne pathogens. mSphere 2018, 3, e17–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from Macroalgae Origin: Health Benefits and Risks for Consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L. A Review of the Nutrient Composition of Selected Edible Seaweeds; Nova Science Publishers Inc.: Athens, GA, USA, 2011; pp. 15–47. [Google Scholar]
- Smith, D.G.; Young, E.G. The combined amino acids in several species of marine algae. J. Biol. Chem. 1955, 217, 845–853. [Google Scholar] [CrossRef]
- Gaillard, C.; Bhatti, H.S.; Novoa-Garrido, M.; Lind, V.; Roleda, M.Y.; Weisbjerg, M.R. Amino acid profiles of nine seaweed species and their in situ degradability in dairy cows. Anim. Feed. Sci. Technol. 2018, 241, 210–222. [Google Scholar] [CrossRef]
- Górska-Warsewicz, H.; Rejman, K.; Laskowski, W.; Czeczotko, M. Milk and Dairy Products and Their Nutritional Contribution to the Average Polish Diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef] [Green Version]
- Haug, A.; Høstmark, A.T.; Harstad, O.M. Bovine milk in human nutrition--a review. Lipids Health Dis. 2007, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Thorning, T.; Raben, A.; Tholstrup, T.; Soedamah-Muthu, S.; Givens, I.; Astrup, A. Milk and dairy products: Good or bad for human health? An assessment of the totality of scientific evidence. Food Nutr. Res. 2016, 60, 32527. [Google Scholar] [CrossRef] [Green Version]
- Antaya, N.T.; Soder, K.J.; Kraft, J.; Whitehouse, N.L.; Guindon, N.E.; Erickson, P.S.; Conroy, A.B.; Brito, A.F. Incremental amounts of Ascophyllum nodosum meal do not improve animal performance but do increase milk iodine output in early lactation dairy cows fed high-forage diets. J. Dairy Sci. 2015, 98, 1991–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besada, V.; Andrade, J.; Schultze, F.; Gonzalez, J. Heavy metals in edible seaweeds commercialised for human consumption. J. Mar. Syst. 2009, 75, 305–313. [Google Scholar] [CrossRef]
- Pomin, V.H. Seaweed: Ecology, Nutrient Composition, and Medicinal Uses; Nova Science Publishers, Inc.: New York, NY, USA, 2011; p. 248. [Google Scholar]
- Smith, J.L.; Summers, G.; Wong, R. Nutrient and heavy metal content of edible seaweeds in New Zealand. N. Z. J. Crop. Hortic. Sci. 2010, 38, 19–28. [Google Scholar] [CrossRef]
- Commision Regulation (EC). Consolidated text: Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union L 2006, 187, 11–15. [Google Scholar]
- EFSA. Dietary exposure to inorganic arsenic in the European population. EFSA J. 2014, 12. [Google Scholar] [CrossRef]
- Ronan, J.M.; Stengel, D.B.; Raab, A.; Feldmann, J.; O’Hea, L.; Bralatei, E.; McGovern, E. High proportions of inorganic arsenic in Laminaria digitata but not in Ascophyllum nodosum samples from Ireland. Chemosphere 2017, 186, 17–23. [Google Scholar] [CrossRef]
- Pétursdóttir, Á.H.; Gunnlaugsdóttir, H. Selective and fast screening method for inorganic arsenic in seaweed using hydride generation inductively coupled plasma mass spectrometry (HG-ICPMS). Microchem. J. 2019, 144, 45–50. [Google Scholar] [CrossRef]
- El-Said, G.; El-Sikaily, A. Chemical composition of some seaweed from Mediterranean Sea coast, Egypt. Environ. Monit. Assess. 2012, 185, 6089–6099. [Google Scholar] [CrossRef] [Green Version]
- Mišurcová, L. Chemical Composition of Seaweeds. In Handbook of Marine Macroalgae; TBU Publications: Zlín, Czech Republic, 2012; pp. 171–192. [Google Scholar]
- Mwalugha, H.; Wakibia, J.; Kenji, G.; Mwasaru, M. Chemical Composition of Common Seaweeds from the Kenya Coast. J. Food Res. 2015, 4, 28. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EU) 2015/186 of 6 February 2015 Amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council as Regards Maximum Levels for Arsenic, Fluorine, Lead, Mercury, Endosulfan and Ambrosia Seeds Text with EEA Relevance; European Commission: Brussels, Belgium, 2015. [Google Scholar]
- Butler, G.; Nielsen, J.H.; Slots, T.; Seal, C.; Eyre, M.D.; Sanderson, R.; Leifert, C. Fatty acid and fat-soluble antioxidant concentrations in milk from high- and low-input conventional and organic systems: Seasonal variation. J. Sci. Food Agric. 2008, 88, 1431–1441. [Google Scholar] [CrossRef]
- Sjaunja, L.O.; Junkkarinen, L.; Pedersen, J.; Setala, J. A Nordic proposal for an energy corrected milk (ECM) formula. In Proceedings of the 27th Session International Committee for Recording and Productivity of Milk Animals, Paris, France, 2–6 July 1990; pp. 1156–1157. [Google Scholar]
- NordVal International. Trace Elements—As, Cd, Hg, Pb and Other Elements: Determination by ICP-MS after Pressure Digestion; NMKL: Tamil Nadu, India, 2007; p. 186. [Google Scholar]
- Payling, L.M.; Juniper, D.T.; Drake, C.; Rymer, C.; Givens, D.I. Effect of milk type and processing on iodine concentration of organic and conventional winter milk at retail: Implications for nutrition. Food Chem. 2015, 178, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Minitab (Version 18). Minitab 18: Session Commands; Minitab. Inc.: State College, PA, USA, 2019; Available online: https://support.minitab.com/en-us/minitab/18/session_commands.pdf (accessed on 17 March 2021).
- Hong, Z.S.; Kim, E.J.; Jin, Y.C.; Lee, J.S.; Choi, Y.J.; Lee, H.G. Effects of Supplementing Brown Seaweed By-products in the Diet of Holstein Cows during Transition on Ruminal Fermentation, Growth Performance and Endocrine Responses. Asian Aust. J. Anim. Sci. 2015, 28, 1296–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves Lopez, C.; Serio, A.; Rossi, C.; Mazzarrino, G.; Marchetti, S.; Castellani, F.; Grotta, L.; Fiorentino, F.P.; Paparella, A.; Martino, G. Effect of diet supplementation with Ascophyllum nodosum on cow milk composition and microbiota. J. Dairy Sci. 2016, 99, 6285–6297. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Nielsen, J.H.; Larsen, M.K.; Slots, T.; Steinshamn, H.; Butler, G. Effect of Feeding Intensity and Milking System on Nutritionally Relevant Milk Components in Dairy Farming Systems in the North East of England. J. Agric. Food Chem. 2012, 60, 7270–7281. [Google Scholar] [CrossRef]
- Stergiadis, S.; Bieber, A.; Franceschin, E.; Isensee, A.; Eyre, M.D.; Maurer, V.; Chatzidimitriou, E.; Cozzi, G.; Bapst, B.; Stewart, G.; et al. Impact of US Brown Swiss genetics on milk quality from low-input herds in Switzerland: Interactions with grazing intake and pasture type. Food Chem. 2015, 175, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Chopra, R.; Rai, S.; Verma, M.; Mohanta, R. Nutritional Evaluation of Seaweed on Nutrient Digestibility, Nitrogen Balance, Milk Production and Composition in Sahiwal Cows. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 87, 437–443. [Google Scholar] [CrossRef]
- Suttle, N. Mineral Nutrition of Livestock, 4th ed.; Cabi: Wallingford, UK, 2010; pp. 1–547. [Google Scholar]
- Sorge, U.S.; Henriksen, M.; Bastan, A.; Cremers, N.; Olsen, K.; Crooker, B.A. Short communication: Iodine concentrations in serum, milk, and tears after feeding Ascophyllum nodosum to dairy cows—A pilot study. J. Dairy Sci. 2016, 99, 8472–8476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Sun, F.; Jiang, L.; Hua, D.; Wang, Y.; Nan, X.; Zhao, Y.; Xiong, B. Effects of Partial Replacment of Dietary Forage Using Kelp Powder (Thallus laminariae) on Ruminal Fermentation and Lactation Performances of Dairy Cows. Animals 2019, 9, 852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöne, F.; Leiterer, M.; Lebzien, P.; Bemmann, D.; Spolders, M.; Flachowsky, G. Iodine concentration of milk in a dose–response study with dairy cows and implications for consumer iodine intake. J. Trace Elem. Med. Biol. 2009, 23, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Flachowsky, G.; Franke, K.; Meyer, U.; Leiterer, M.; Schöne, F. Influencing factors on iodine content of cow milk. Eur. J. Nutr. 2013, 53, 351–365. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the safety and efficacy of iodine compounds (E2) as feed additives for all species: Calcium iodate anhydrous and potassium iodide, based on a dossier submitted by HELM AG. EFSA J. 2013, 11, 3101. [Google Scholar] [CrossRef]
- NRC. Mineral Tolerance of Animals; The National Academies Press: Washington, DC, USA, 2005. [Google Scholar]
- Arriagada, A.A.; Albornoz, E.; Opazo, M.C.; Becerra, A.; Vidal, G.; Fardella, C.; Michea, L.; Carrasco, N.; Simon, F.; Elorza, A.A.; et al. Excess Iodide Induces an Acute Inhibition of the Sodium/Iodide Symporter in Thyroid Male Rat Cells by Increasing Reactive Oxygen Species. Endocrinology 2015, 156, 1540–1551. [Google Scholar] [CrossRef] [Green Version]
- Ammerman, C.B.; Miller, S.M. Selenium in Ruminant Nutrition: A Review1. J. Dairy Sci. 1975, 58, 1561–1577. [Google Scholar] [CrossRef]
- Grace, N.D.; Lee, J.; Mills, R.A.; Death, A.F. Influence of Se status on milk Se concentrations in dairy cows. N. Z. J. Agric. Res. 1997, 40, 75–78. [Google Scholar] [CrossRef]
- Galbraith, M.; Vorachek, W.; Estill, C.; Whanger, P.; Bobe, G.; Davis, T.; Hall, J. Rumen Microorganisms Decrease Bioavailability of Inorganic Selenium Supplements. Biol. Trace Elem. Res. 2016, 171, 338–343. [Google Scholar] [CrossRef]
- Netto, A.S.; Zanetti, M.A.; Correa, L.B.; Del Claro, G.R.; Salles, M.S.V.; Vilela, F.G. Effects of dietary selenium, sulphur and copper levels on selenium concentration in the serum and liver of lamb. Asian Aust. J. Anim. Sci. 2014, 27, 1082–1087. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Akhtara, S.; Amjada, A.; Naeema, I.; Tariqa, M. Comparative Assessment of Arsenic Contamination in Raw Milk, Infant Formulas and Breast Milk. J. Dairy Vet. Sci. 2019, 13, 555851. [Google Scholar]
- Cubadda, F.; Jackson, B.P.; Cottingham, K.L.; Van Horne, Y.O.; Kurzius-Spencer, M. Human exposure to dietary inorganic arsenic and other arsenic species: State of knowledge, gaps and uncertainties. Sci. Total Environ. 2017, 579, 1228–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bampidis, V.; Nistor, E.; Nitas, D. Arsenic, cadmium, lead and mercury as undesirable substances in animal feeds. Scientific Pap. Anim. Sci. Biotechnol. 2013, 46, 17–22. [Google Scholar]
- Scholz-Ahrens, K.E.; Ahrens, F.; Barth, C.A. Nutritional and health attributes of milk and milk imitations. Eur. J. Nutr. 2020, 59, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Iceland, Department of Health. Ráðlagðir Dagskammtar (RDS) af Ýmsum Vítamínum; Department of Health: Reykjavík, Iceland, 2013.
- WHO. Iodine Deficiency in Europe: A Continuing Public Health Problem; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Adalsteinsdottir, S.; Tryggvadottir, E.; Hrolfsdottir, L.; Halldorsson, T.; Birgisdottir, B.; Hreidarsdottir, I.; Hardardottir, H.; Arohonka, P.; Erlund, I.; Gunnarsdottir, I. Insufficient iodine status in pregnant women as consequence of dietary changes. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Faludi, G.; Beauclercq, S.; Pitt, J.; Desnica, N.; Petursdottir, A.; Newton, E.E.; Angelidis, A.; Givens, I.; Juniper, D.; et al. Macromineral and Trace Element Concentrations and Their Seasonal Variation in Milk from Organic and Conventional Dairy Herds; Elsevier: Amsterdam, The Netherlands, 2021; p. 129865. [Google Scholar]
- Dahl, L.; Opsahl, J.; Meltzer, H.; Julshamn, K. Iodine concentration in Norwegian milk and dairy products. Br. J. Nutr. 2003, 90, 679–685. [Google Scholar] [CrossRef]
- O’Brien, B.; Gleeson, D.E.; Jordan, K. Iodine concentrations in milk. Ir. J. Agric. Food Res. 2013, 209–216. [Google Scholar]
- Hansen, K. Iceland’s Raging Rivers. Available online: https://earthobservatory.nasa.gov/images/145408/icelands-raging-rivers (accessed on 28 June 2021).
- Nyström, H.F.; Brantsæter, A.L.; Erlund, I.; Gunnarsdottir, I.; Hulthén, L.; Laurberg, P.; Mattisson, I.; Rasmussen, L.B.; Virtanen, S.; Meltzer, H.M. Iodine status in the Nordic countries—Past and present. Food Nutr. Res. 2016, 60, 31969. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.; Orth, B.; Reiser, R.; Bürge, D.; Lehto, N.; Almond, P.; Gaw, S.; Thomson, B.; Lilburne, L.; Robinson, B. Environmental Parameters Affecting the Concentration of Iodine in New Zealand Pasture. J. Environ. Qual. 2019, 48, 1517–1523. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.A.; Bruhn, J.C.; Osland, R.B. Factors Affecting Iodine Concentration of Milk of Individual Cows. J. Dairy Sci. 1983, 66, 997–1002. [Google Scholar] [CrossRef]
- FAOSTAT. Food Supply-Livestock and Fish Primary Equivalent; FAOSTAT: Rome, Itlay, 2013. [Google Scholar]
- Leung, A.M.; Braverman, L.E. Consequences of excess iodine. Nat. Rev. Endocrinol. 2014, 10, 136–142. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Tolerable Upper Intake Levels for Vitamins and Minerals; EFSA: Parma, Italy, 2006. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Hughes, M.F.; Beck, B.D.; Chen, Y.; Lewis, A.S.; Thomas, D.J. Arsenic exposure and toxicology: A historical perspective. Toxicol. Sci. 2011, 123, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Ravenscroft, P.; Brammer, H.; Richards, K. Arsenic Pollution: A Global Synthesis; Wiley-Blackwell: Chichester, UK, 2009; pp. 387–454. [Google Scholar] [CrossRef]
- Grandjean, A. Water Requirements, Impinging Factors, and Recommended Intakes. Nutr. Drink. Water 2009. Available online: https://aloyoun.com.sa/documentation/nutrientsindw.pdf#page=34 (accessed on 28 June 2021).
- Jackson, B.P.; Taylor, V.F.; Punshon, T.; Cottingham, K.L. Arsenic concentration and speciation in infant formulas and first foods. Pure Appl. Chem. 2012, 84, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Silage | Concentrate 1 | |||||||
|---|---|---|---|---|---|---|---|---|
| Chemical Composition (g/kg Dry Matter) | Mean | SD | Min | Max | Mean | SD | Min | Max |
| Dry Matter (g/kg fresh) | 301 | 7.0 | 290 | 309 | 895 | 1.2 | 894 | 897 |
| Ash | 70 | 2.4 | 67 | 74 | 89 | 2.7 | 83 | 93 |
| NCDG 2 | 768 | 12.2 | 750 | 780 | NM 3 | NM 3 | NM 3 | NM 3 |
| Crude Protein | 166 | 11.6 | 143 | 178 | 213 | 3.3 | 203 | 220 |
| Neutral Detergent Fiber | 501 | 15.5 | 483 | 525 | 118 | 6.6 | 103 | 129 |
| Acid Detergent Fiber | 301 | 13.6 | 285 | 328 | 564 | 46.0 | 501 | 647 |
| Single Cell Protein | 111 | 5.8 | 101 | 118 | NM 3 | NM 3 | NM 3 | NM 3 |
| Indigestible Neutral Detergent Fiber | 87 | 12.1 | 78 | 113 | NM 3 | NM 3 | NM 3 | NM 3 |
| Sugar | 47 | 9.9 | 30 | 62 | NM 3 | NM 3 | NM 3 | NM 3 |
| Fat | 59 | 4.7 | 54 | 65 | 26 | 1.2 | 24 | 28 |
| Ammonia | 0.7 | 0.14 | 0.5 | 0.9 | NM 3 | NM 3 | NM 3 | NM 3 |
| Starch | NM 3 | NM 3 | NM 3 | NM 3 | 276 | 9.4 | 252 | 286 |
| Water-soluble Carbohydrates | NM 3 | NM 3 | NM 3 | NM 3 | 136 | 12.2 | 115 | 160 |
| Silage | Concentrate 1 | Concentrate with Seaweed 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Minerals (mg/kg Dry Matter) | Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max |
| Aluminum (Al) | 552 | 698.8 | 164 | 2107 | 510 | 166.1 | 408 | 701.6 | 419 | 50.3 | 383 | 454 |
| Arsenic (As) | 0.07 | 0.045 | 0.04 | 0.17 | 0.77 | 0.526 | 0.41 | 1.373 | 1.06 | 0.106 | 0.98 | 1.13 |
| Cadmium (Cd) | 0.02 | 0.005 | 0.01 | 0.02 | 0.08 | 0.022 | 0.05 | 0.097 | 0.11 | 0.004 | 0.11 | 0.12 |
| Calcium (Ca) | 4373 | 484.8 | 3859 | 5220 | 14,945 | 342.8 | 14,632 | 15,311 | 14,979 | 446.1 | 14,663 | 15,294 |
| Chromium (Cr) | 37 | 40.7 | 12 | 128 | 26.1 | 8.0 | 19.2 | 34.78 | 22 | 3.0 | 20 | 24 |
| Cobalt (Co) | 0.66 | 0.461 | 0.37 | 1.68 | 3.60 | 0.785 | 2.82 | 4.393 | 2.67 | 0.184 | 2.54 | 2.80 |
| Copper (Cu) | 11 | 1.4 | 9 | 13 | 71 | 10.2 | 60 | 80.88 | 57 | 3.8 | 55 | 60 |
| Iodine (I) | 0.22 | 0.138 | 0.14 | 0.53 | 4.2 | 0.33 | 3.6 | 4.499 | 18 | 6.2 | 8 | 26 |
| Iron (Fe) | 1468 | 1317.2 | 685 | 4392 | 917 | 260.9 | 723 | 1212.0 | 760 | 50.1 | 725 | 795 |
| Lead (Pb) | 0.09 | 0.081 | 0.04 | 0.27 | 0.37 | 0.084 | 0.29 | 0.460 | 0.20 | 0.000 | 0.20 | 0.20 |
| Magnesium (Mg) | 1939 | 182.1 | 1757 | 2260 | 4936 | 250.8 | 4783 | 5225 | 4670 | 166.4 | 4552 | 4788 |
| Manganese (Mn) | 73 | 11.6 | 54 | 89 | 181 | 36.8 | 145 | 218.0 | 193 | 28.1 | 173 | 213 |
| Mercury (Hg) | 0.01 | 0.000 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.003 | 0.0035 | 0.000 | 0.005 |
| Molybdenum (Mo) | 0.67 | 0.366 | 0.37 | 1.46 | 2.2 | 0.39 | 1.8 | 2.517 | 1.94 | 0.209 | 1.80 | 2.09 |
| Nickel (Ni) | 13 | 9.6 | 6 | 34 | 11.9 | 2.31 | 9.3 | 13.72 | 11.3 | 1.06 | 10.5 | 12.0 |
| Phosphorus (P) | 3091 | 632.2 | 2635 | 4429 | 7208 | 257.1 | 5912 | 7372 | 7005 | 416.3 | 6710 | 7299 |
| Potassium (K) | 20,399 | 2102.5 | 17,581 | 24,024 | 11,991 | 739.6 | 11,254 | 12,733 | 12,814 | 935.2 | 12,152 | 13,475 |
| Selenium (Se) | 0.17 | 0.055 | 0.10 | 0.28 | 0.94 | 0.052 | 0.89 | 0.993 | 0.98 | 0.039 | 0.96 | 0.01 |
| Sodium (Na) | 1067 | 220.6 | 730 | 1331 | 3279 | 157.6 | 3111 | 3424 | 3082 | 184.4 | 2951 | 3212 |
| Tin (Sn) | 0.09 | 0.038 | 0.06 | 0.17 | 0.12 | 0.017 | 0.10 | 0.130 | 0.13 | 0.004 | 0.13 | 0.14 |
| Zinc (Zn) | 43 | 5.1 | 36 | 51 | 153 | 29.9 | 126 | 185.2 | 130 | 13.7 | 120 | 139 |
| Diet | ANOVA p-Values 1 | ||||||
|---|---|---|---|---|---|---|---|
| Parameters | CON n = 66 | LSW n = 78 | HSW n = 78 | SE | Diet | Week | Diet × Week |
| Animal Data | |||||||
| Parity | 2.0 | 2.1 | 2.3 | 0.15 | |||
| Lactation weeks | 20.0 | 24.0 | 21.9 | 1.79 | |||
| Bodyweight (kg) | 445 | 446 | 450 | 5.0 | |||
| Animal Diet | |||||||
| Dry Matter Intake 2 (kg/d) | 14.3 | 14.3 | 14.5 | 0.08 | 0.075 | 0.041 | 0.943 |
| Forage:concentrate | 44.7 | 45.4 | 44.8 | 1.80 | 0.942 | <0.001 | 0.793 |
| Silage Intake (kg DM/d) | 6.35 | 5.43 | 6.47 | 0.254 | 0.946 | <0.001 | 0.776 |
| Concentrate Intake (kg DM/d) | 8.01 | 7.85 | 8.02 | 0.251 | 0.872 | <0.001 | 0.844 |
| Seaweed Intake (g DM/d) | 0.00 c | 12.8 b | 50.2 a | 0.004 | <0.001 | <0.001 | <0.001 |
| Milk Production | |||||||
| Yield (kg/d) | 25.3 | 24.9 | 26.5 | 0.60 | 0.097 | 0.041 | 0.943 |
| ECMY 3 (kg/d) | 27.0 | 25.9 | 27.1 | 0.70 | 0.399 | 0.133 | 0.775 |
| Milk Composition | |||||||
| Fat (g/100 g) | 4.56 | 4.46 | 4.35 | 0.082 | 0.157 | 0.843 | 0.878 |
| Protein (g/100 g) | 3.33 a | 3.27 a | 3.20 b | 0.027 | 0.004 | <0.001 | 0.632 |
| Casein (g/100 g) | 2.43 a | 2.39 a | 2.33 b | 0.234 | 0.006 | <0.001 | 0.694 |
| Lactose (g/100 g) | 4.58 | 4.62 | 4.63 | 0.030 | 0.517 | <0.001 | 0.767 |
| Whey Protein (g/100 g) | 0.90 | 0.88 | 0.87 | 0.008 | 0.111 | <0.001 | 0.627 |
| Urea (mmol/L) | 6.37 | 6.13 | 3.08 | 0.139 | 0.283 | <0.001 | 0.109 |
| Free Fatty Acids (mmol/L) | 0.80 | 0.87 | 0.91 | 0.038 | 0.144 | <0.001 | 0.929 |
| Fat:Protein | 1.37 | 1.37 | 1.36 | 0.028 | 0.931 | 0.198 | 0.579 |
| Somatic Cell Count (×103/mL) | 181 | 206 | 193 | 65.1 | 0.965 | 0.255 | 0.699 |
| Efficiency (g/kg DMI) | |||||||
| Feed Efficiency | 1753 | 1730 | 1807 | 29.6 | 0.134 | 0.022 | 0.899 |
| Fat Efficiency | 80.4 | 76.8 | 77.8 | 2.15 | 0.461 | 0.335 | 0.637 |
| Protein Efficiency | 58.3 | 56.2 | 57.3 | 0.86 | 0.191 | 0.244 | 0.894 |
| Diet | ANOVA p-Values 1 | ||||||
|---|---|---|---|---|---|---|---|
| Minerals | CON n = 66 | LSW n = 78 | HSW n = 78 | SE | Diet | Week | Diet × Week |
| Macrominerals (mg/kg) | |||||||
| Calcium (Ca) | 1129 | 1076 | 1053 | 29.7 | 0.192 | <0.001 | 0.797 |
| Magnesium (Mg) | 110.4 | 103.0 | 99.2 | 4.30 | 0.179 | 0.021 | 0.481 |
| Phosphorus (P) | 881.8 | 866.8 | 851.0 | 26.72 | 0.708 | <0.001 | 0.892 |
| Potassium (K) | 1471 | 1433 | 1423 | 40.2 | 0.661 | <0.001 | 0.711 |
| Sodium (Na) | 432.9 | 435.2 | 403.0 | 20.31 | 0.422 | 0.033 | 0.525 |
| Essential Trace Elements (μg/kg) | |||||||
| Copper (Cu) | 47.3 a | 40.9 ab | 35.7 b | 3.05 | 0.034 | <0.001 | 0.364 |
| Iron (Fe) | 223.9 | 224.1 | 223.9 | 9.72 | 1.000 | 0.020 | 0.337 |
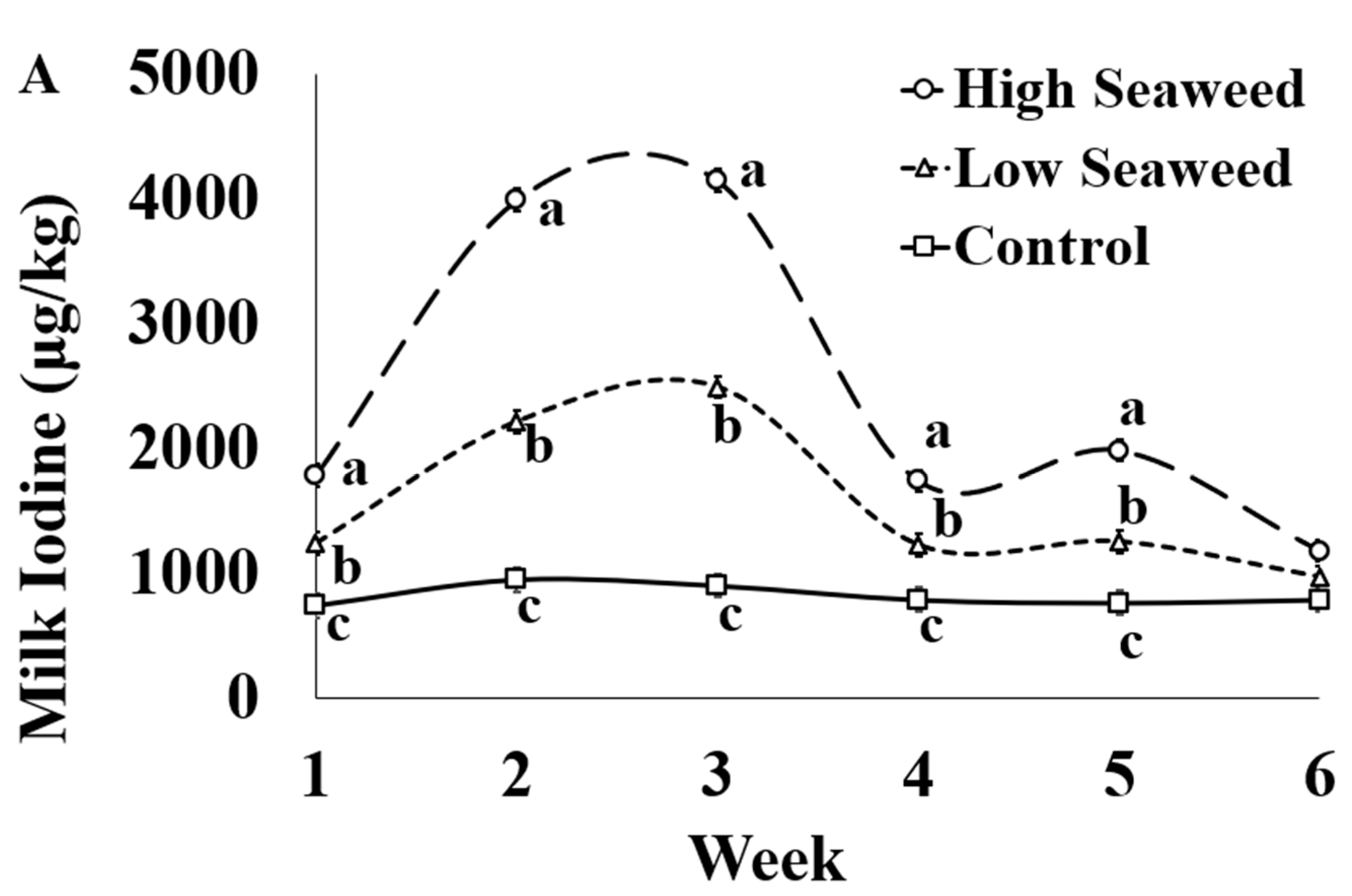
| Iodine (I) | 821.5 c | 1565.3 b | 2470.8 a | 60.98 | <0.001 | <0.001 | <0.001 |
| Manganese (Mn) | 27.5 | 28.4 | 27.4 | 1.06 | 0.717 | 0.009 | 0.173 |
| Molybdenum (Mo) | 52.5 | 51.9 | 49.4 | 1.62 | 0.346 | <0.001 | 0.296 |
| Nickel (Ni) | 2.49 | 1.60 | 1.40 | 0.440 | 0.182 | <0.001 | 0.105 |
| Selenium (Se) | 23.2 a | 21.8 b | 20.1 c | 0.50 | <0.001 | <0.001 | 0.987 |
| Zinc (Zn) | 4720 | 4683 | 4406 | 125.5 | 0.137 | <0.001 | 0.842 |
| Non-Essential Trace Elements (μg/kg) | |||||||
| Aluminum (Al) | 63.7 | 57.3 | 60.1 | 4.53 | 0.577 | <0.001 | 0.202 |
| Cobalt (Co) | 0.52 | 0.48 | 0.43 | 0.029 | 0.088 | <0.001 | 0.140 |
| Heavy Metals (μg/kg) | |||||||
| Arsenic (As) | 0.455 b | 0.483 b | 0.622 a | 0.0416 | 0.013 | <0.001 | 0.102 |
| Diet | ANOVA p-Values 1 | ||||||
|---|---|---|---|---|---|---|---|
| Minerals (g in Milk/100 g Ingested) | CON n = 66 | LSW n = 78 | HSW n = 78 | SE | Diet | Week | Diet × Week |
| Macrominerals | |||||||
| Calcium (Ca) | 19.7 | 18.4 | 18.0 | 0.67 | 0.170 | <0.001 | 0.830 |
| Magnesium (Mg) | 8.7 | 7.9 | 7.6 | 0.51 | 0.294 | <0.001 | 0.846 |
| Phosphorus (P) | 29.2 | 28.1 | 27.8 | 0.99 | 0.590 | <0.001 | 0.944 |
| Potassium (K) | 16.2 | 16.1 | 15.8 | 0.72 | 0.912 | <0.001 | 0.834 |
| Sodium (Na) | 33.3 | 32.4 | 29.9 | 1.74 | 0.341 | 0.031 | 0.488 |
| Essential Trace Elements | |||||||
| Copper (Cu) | 0.20 a | 0.17 a,b | 0.15 b | 0.145 | 0.042 | <0.001 | 0.308 |
| Iron (Fe) | 0.04 | 0.04 | 0.04 | 0.002 | 0.638 | <0.001 | 0.364 |
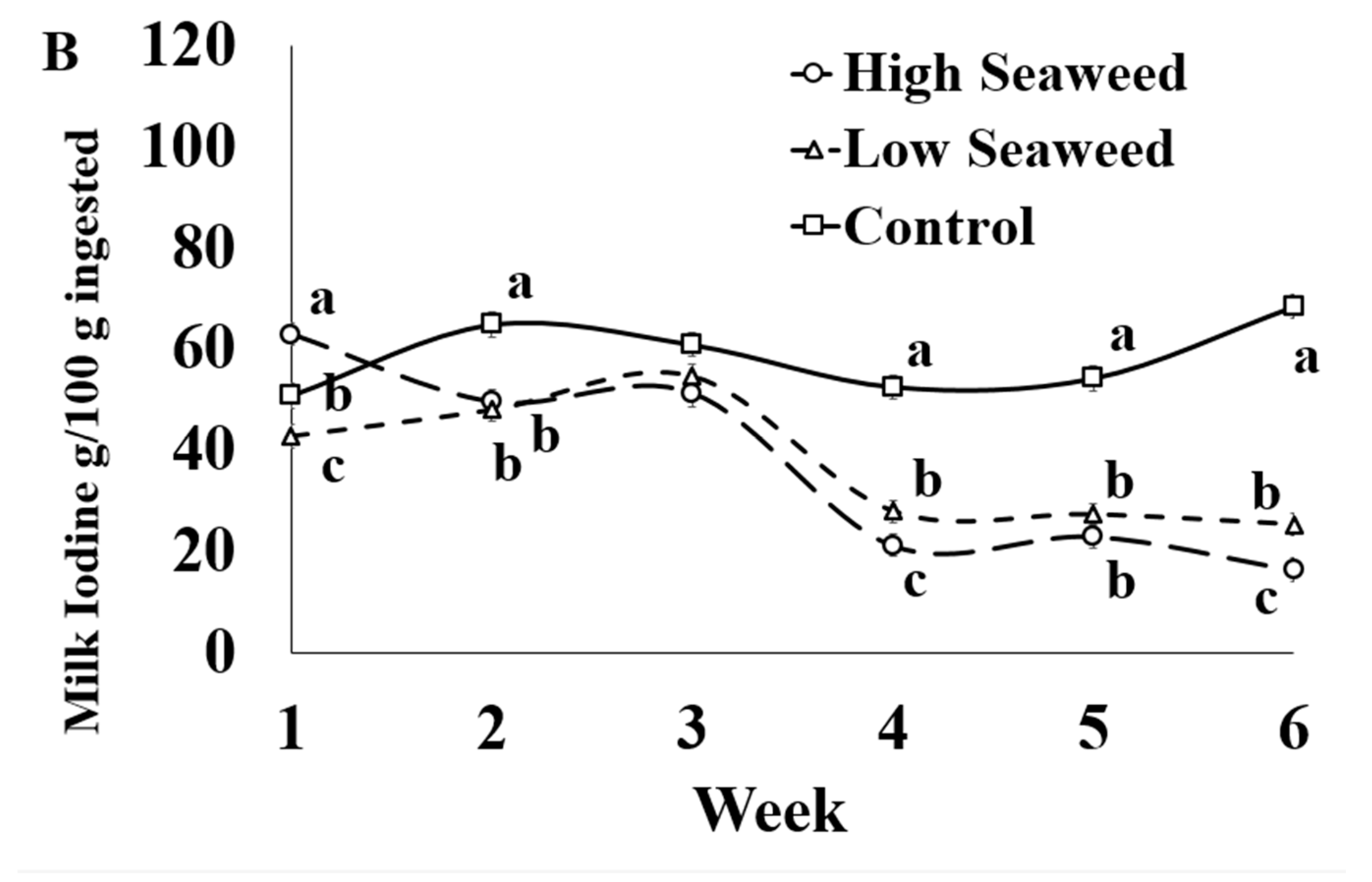
| Iodine (I) | 58.7 a | 37.7 b | 37.5 b | 1.70 | <0.001 | <0.001 | <0.001 |
| Manganese (Mn) | 0.04 | 0.04 | 0.04 | 0.002 | 0.950 | <0.001 | 0.279 |
| Molybdenum (Mo) | 6.8 | 6.6 | 6.3 | 0.23 | 0.378 | <0.001 | 0.436 |
| Nickel (Ni) | 0.05 | 0.03 | 0.02 | 0.011 | 0.215 | <0.001 | 0.087 |
| Selenium (Se) | 7.2 a | 6.5 b | 6.2 b | 0.22 | 0.007 | <0.001 | 0.961 |
| Zinc (Zn) | 8.5 | 8.4 | 7.9 | 0.23 | 0.128 | <0.001 | 0.690 |
| Non-Essential Trace Elements | |||||||
| Aluminum (Al) | 0.03 | 0.02 | 0.03 | 0.002 | 0.563 | <0.001 | 0.252 |
| Cobalt (Co) | 0.044 a | 0.039 a,b | 0.035 b | 0.0025 | 0.037 | <0.001 | 0.128 |
| Heavy metals | |||||||
| Arsenic (As) | 0.22 | 0.20 | 0.19 | 0.018 | 0.679 | <0.001 | 0.252 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Newton, E.E.; Pétursdóttir, Á.H.; Ríkharðsson, G.; Beaumal, C.; Desnica, N.; Giannakopoulou, K.; Juniper, D.; Ray, P.; Stergiadis, S. Effect of Dietary Seaweed Supplementation in Cows on Milk Macrominerals, Trace Elements and Heavy Metal Concentrations. Foods 2021, 10, 1526. https://doi.org/10.3390/foods10071526
Newton EE, Pétursdóttir ÁH, Ríkharðsson G, Beaumal C, Desnica N, Giannakopoulou K, Juniper D, Ray P, Stergiadis S. Effect of Dietary Seaweed Supplementation in Cows on Milk Macrominerals, Trace Elements and Heavy Metal Concentrations. Foods. 2021; 10(7):1526. https://doi.org/10.3390/foods10071526
Chicago/Turabian StyleNewton, Eric E., Ásta H. Pétursdóttir, Gunnar Ríkharðsson, Corentin Beaumal, Natasa Desnica, Konstantina Giannakopoulou, Darren Juniper, Partha Ray, and Sokratis Stergiadis. 2021. "Effect of Dietary Seaweed Supplementation in Cows on Milk Macrominerals, Trace Elements and Heavy Metal Concentrations" Foods 10, no. 7: 1526. https://doi.org/10.3390/foods10071526
APA StyleNewton, E. E., Pétursdóttir, Á. H., Ríkharðsson, G., Beaumal, C., Desnica, N., Giannakopoulou, K., Juniper, D., Ray, P., & Stergiadis, S. (2021). Effect of Dietary Seaweed Supplementation in Cows on Milk Macrominerals, Trace Elements and Heavy Metal Concentrations. Foods, 10(7), 1526. https://doi.org/10.3390/foods10071526