Abstract
Antiviral treatments inhibiting Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) replication may represent a strategy complementary to vaccination to fight the ongoing Coronavirus disease 19 (COVID-19) pandemic. Molecules or extracts inhibiting the SARS-CoV-2 chymotripsin-like protease (3CLPro) could contribute to reducing or suppressing SARS-CoV-2 replication. Using a targeted approach, we identified 17 plant products that are included in current and traditional cuisines as promising inhibitors of SARS-CoV-2 3CLPro activity. Methanolic extracts were evaluated in vitro for inhibition of SARS-CoV-2 3CLPro activity using a quenched fluorescence resonance energy transfer (FRET) assay. Extracts from turmeric (Curcuma longa) rhizomes, mustard (Brassica nigra) seeds, and wall rocket (Diplotaxis erucoides subsp. erucoides) at 500 µg mL−1 displayed significant inhibition of the 3CLPro activity, resulting in residual protease activities of 0.0%, 9.4%, and 14.9%, respectively. Using different extract concentrations, an IC50 value of 15.74 µg mL−1 was calculated for turmeric extract. Commercial curcumin inhibited the 3CLPro activity, but did not fully account for the inhibitory effect of turmeric rhizomes extracts, suggesting that other components of the turmeric extract must also play a main role in inhibiting the 3CLPro activity. Sinigrin, a major glucosinolate present in mustard seeds and wall rocket, did not have relevant 3CLPro inhibitory activity; however, its hydrolysis product allyl isothiocyanate had an IC50 value of 41.43 µg mL−1. The current study identifies plant extracts and molecules that can be of interest in the search for treatments against COVID-19, acting as a basis for future chemical, in vivo, and clinical trials.
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a predominantly airborne transmitted coronavirus first identified in December 2019, which is the causal agent of Coronavirus disease 19 (COVID-19) [1]. Rapid spread of SARS-CoV-2 has caused a severe pandemic, resulting so far in a death toll of almost 3 million lives worldwide [2] and a disruption of many aspects of human daily life. SARS-CoV-2 is an enveloped, simple-stranded, positive-sense RNA virus with a length of 29.9 kb and a diameter of about 65–125 nm [3]. Since its identification, the genome of multiple strains of SARS-CoV-2 has been sequenced [4,5,6] and its life cycle has been thoroughly studied [3].
Many vaccine projects have been developed and are underway to fight COVID-19 [7]. Several of the first-generation vaccines already in use in vaccination campaigns have shown high levels of efficacy, albeit the level of protection is not complete and seems to be strain-dependent. In this way, the recent appearance of new highly infectious SARS-CoV-2 strains might reduce the efficacy of first-generation vaccines [8]. Moreover, vaccines are still in short supply and, in many countries, vaccination against SARS-CoV-2 is neither compulsory nor indicated for some specific groups, such as children, or people with certain medical conditions [9,10]. Therefore, the development of therapies complementary to vaccination is desirable for a comprehensive fight against COVID-19.
The development of drugs for the treatment of COVID-19 patients so far has not resulted in highly efficient or curative drug therapies. Despite the initial consideration of some pre-existent drugs as potential antiviral against SARS-CoV-2, no proven effective treatments have been found so far [11]. Among the proteins encoded by the SARS-CoV-2 RNA, the chymotrypsin-like protease (3CLPro), also known as main protease (MPro), is required for the proteolytic processing of the viral polyproteins during the maturation step, thus becoming an essential tool for the successful replication of the virus [12]. In this sense, the fundamental role of the 3CLPro in viral replication makes this enzyme an attractive target for the development of drugs inhibiting the virus replication. A similar research strategy was followed in the past against the phylogenetically related SARS-CoV [13], which caused the SARS-CoV 2002–2004 outbreak, and led to the screening of a large variety of molecules and natural products for their inhibition of the SARS-CoV 3CLPro activity [14]. In addition to single molecules, some plant extracts also inhibited the SARS-CoV 3CLPro activity in vitro [15,16].
Some natural compounds with potential against SARS-CoV-2 have been identified based on the work with SARS-CoV and related viruses and through in silico molecular docking studies [17,18,19,20,21]. The recent work of Zhou and Huang [22] reviews the current findings regarding natural compounds that might be considered for their potential antiviral activity against SARS-CoV-2, mostly based on previous experience with other coronaviruses. Some compounds found in edible and/or medicinal plant products were identified in these studies as potential natural compounds targeting the SARS-CoV-2 3CLPro, including glucosinolates, flavonoids, and other phenolic compounds. Phenolic compounds have been extensively studied for their antioxidant capacity acting as free radical scavengers tested in vitro and using in vivo models [23]. Thus, diets rich in fruits and vegetables containing high levels of these compounds are associated with a reduction in the risk of chronic inflammatory processes, thus lowering the risk of developing some degenerative and cardiovascular diseases and several types of cancer, and increasing the overall health status [24,25,26]. In addition, glucosinolates and their hydrolytic products are also bioactive molecules related to the maintenance of a good health status. The most studied derivative, sulforaphane, proved to induce detoxification phase II enzymes [27] and has been studied as an anticarcinogen likely able to be effective in all stages of cancer [28], to reduce the risk of developing type 2 diabetes [29], as well as to ameliorate several neurodegenerative diseases [30]. Other glucosinolates are promising as natural compounds by exerting antimicrobial activities [31,32].
However, the evaluation of the antiviral capacity of these groups of natural compounds is more limited. Based on this consideration, the current work aimed at the targeted testing for inhibitory capacity against the 3CLPro activity of plant extracts selected for being rich in one or more of the aforementioned chemical families. The materials selected constitute common food products in many cultures and are of easy access or, alternatively, grow profusely in many regions and are included in traditional cuisines, as is the case of wall rocket in the Mediterranean region. A cell-free cleavage assay using a fluorogenic substrate was used, as this technique has been used successfully to measure the protease activity of the related SARS-CoV 3CLPro [33,34,35]. Testing plant extracts can be considered a first approach in the search for natural compounds with antiviral activity, or even represent a basis for the development of prophylactic or therapeutic plant extracts against COVID-19. In addition, given the proven safety for human consumption of the plants from which extracts are obtained, their potential use against COVID-19 might be immediate and easily accessible.
2. Materials and Methods
2.1. Plant Material
A total of 17 plant foods and derived products were selected for the current study (Figure S1). The materials were identified by the authors and acquired in a local market at Valencia (Spain) and included citrus fruit peels (sweet orange (Citrus sinensis), lemon (C. limon), lime (C. aurantiifolia), and grapefruit (Citrus paradisi)), seasoning and aromatic herbs (celery leaves and celery stalks (Apium graveolens var. dulce), parsley (Petroselinum crispum), dill (Anethum graveolens), sweet chamomile (Marticaria chamomilla), and dried oregano (Origanum vulgare)), bulbs and rhizomes (red onion (Allium cepa) and turmeric (Curcuma longa)), a succulent plant (aloe vera (Aloe barbadensis)), and cruciferous condiments (brown mustard seeds (Brassica nigra), horseradish (Armoracia rusticana), and commercial wasabi powder (Tokyo-Ya, S.A., Japan)). The ingredients of wasabi powder were horseradish, mustard, vitamin C, acidulant E334, wasabi aroma, and colorants E102 and E133. Dried oregano and sweet chamomile were acquired from Naturcid S.L. (Monforte del Cid, Spain), and mustard seeds were acquired from Diplan S.A. (Fuenlabrada, Spain). In addition, edible baby-leaves of wall rocket or wild rucola (Diplotaxis erucoides subsp. erucoides) were included in the study owing to their accumulation of sinigrin, which derives in allyl isothiocyanate [36,37]. Wall rocket is an herbaceous plant broadly distributed and included in the traditional Mediterranean cuisine and, for the current study, it was grown in the field as described in Guijarro-Real et al. [36]. The selection of these plant materials (Table 1) was done according to their phytochemical composition as reported in the literature, based on the recent review work by Zhou and Huang [22].

Table 1.
Selected phytochemical compounds reported as potential natural inhibitors of SARS-CoV-2 3CLPro activity and naturally present in the plant materials used in the current study.
Approximately, 100 g to 200 g of material was obtained for each sample. For citrus fruits, the peels (including the flavedo and albedo) were manually separated from the flesh and used because of the higher concentrations of flavonoids in the former [38]. Celery was separated into leaves and stalks and processed separately. Turmeric and horseradish were peeled, and peels were discarded. Aloe vera leaves were separated into the outer skin and the inner parenchyma, and the former was discarded. Except for dried oregano, all materials were freeze-dried. Dried materials were ground into powder with a coffee grinder and stored at −80 °C.
2.2. Preparation of Methanolic Extracts
Phytochemical compounds were extracted from samples with 80% aqueous methanol, which has been proven as an appropriate solvent for the extraction of phenolic compounds [63,64] and glucosinolates [65]. For each material, four samples of 100 mg were extracted with 1 mL of 80% methanol upon sonication for 30 min, with intermediate stirring each 5 min to facilitate the extraction process. After a centrifugation step (5 min at 13,000 rpm), the plant material was re-extracted by adding 1 mL of 80% methanol. The recovered extracts of four independent replicates were combined and filtered through a 0.2 µm PTFE filter. Then, 5 mL of the combined extract for each material was dried under nitrogen and resuspended for a stock solution in 100% dimethyl sulfoxide (DMSO) to a final concentration of 50 mg mL−1. Extracts were kept at −80 °C until analysis.
2.3. FRET Assay for the Inhibition of the 3CL Protease Activity (SARS-CoV-2 3CLPro)
The extracts were evaluated as inhibitors of SARS-CoV-2 3CLPro by means of an in vitro quenched fluorescence resonance energy transfer (FRET) assay using a fluorogenic substrate to measure the residual activity. For the study, the MBP-tagged 3CL Protease (SARS-CoV-2) Assay Kit (BPS Bioscience, San Diego, CA, USA) was used according to the manufacturer’s instructions. Briefly, the plant-based extracts were conveniently diluted in the 3CLPro assay buffer to an extract concentration of 2.5 mg mL−1. For determination of the 3CLPro activity, 5 µL of the diluted plant extracts was pre-incubated with 150 ng of the 3CLPro for 30 min. Subsequently, the fluorogenic substrate was added to a final concentration of 50 µM and the reaction was incubated for 4 h in the darkness in the presence of 1 mM 1,4-dithio-D,L-threitol (DTT). The final concentration of the plant extracts during the reaction was 500 µg mL−1 in a reaction volume of 25 µL. The fluorescence intensity was recorded at 470 nm after excitation at 360 nm. A positive control was included to measure the maximum activity of the protease in the absence of potential inhibitors. Moreover, an inhibition control was included by pre-incubating 150 ng of the 3CLPro with 5 µL of the inhibitor GC376 (250 µM) supplied by the manufacturer. The reaction was evaluated in four replicates for each extract.
The extracts showing significant inhibition of the 3CLPro activity (<20% of residual activity compared with the positive control) were selected for the determination of the correspondent IC50. The protocol followed was the same as described above, although in this case, seven concentrations of the plant-based extracts, 5.0–10–25–50–100–200–500 µg mL−1, were evaluated. Additionally, commercial standards of curcumin (>94% curcuminoids with >80% curcumin, CAS Number 458-37-7), sinigrin (CAS Number 3952-98-5), and allyl isothiocyanate (CAS Number 57-06-7) (Sigma-Aldrich, Saint Louis, MO, USA) were also resuspended in DMSO and tested as 3CLPro activity inhibitors at different concentrations. As for the first screening, the inhibitory capacity of the plant extracts and the commercial standards was tested using four replicates for each concentration considered.
2.4. Data Analysis
The fluorescence intensity measured indicated the residual activity of the protease after the co-incubation with extracts. The residual activities were provided in percentage against the maximum activity obtained in the absence of potential inhibitory compounds or extracts (100%). For each material and concentration tested, the average value and corresponding standard error were calculated (n = 4) using the statistics software Statgraphics Centurion XVII (Statpoint Technologies, Warrenton, VA, USA), and the significance of differences among materials was tested by one-way factorial analysis of variance. Mean separation was performed with a Student–Newman–Keuls test at a signification level of p < 0.05. The dose–response curves and the relative IC50 for extracts and standards showing significant inhibition were obtained using the GraphPad Prism 9.0.1 software (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. Screening of Plant-Based Extracts for Inhibition of SARS-CoV-2 3CLPro Activity
The inhibition of SARS-CoV-2 3CLPro activity by the 17 extracts was tested at a final concentration of 500 µg mL−1. The extracts of lime peel and chamomile produced signal interferences and were discarded from the analysis. It was noted, however, that the co-incubation of the 3CLPro with the chamomile extracts produced a reduction of the protease activity even if a signal interference was observed (Figure S2).
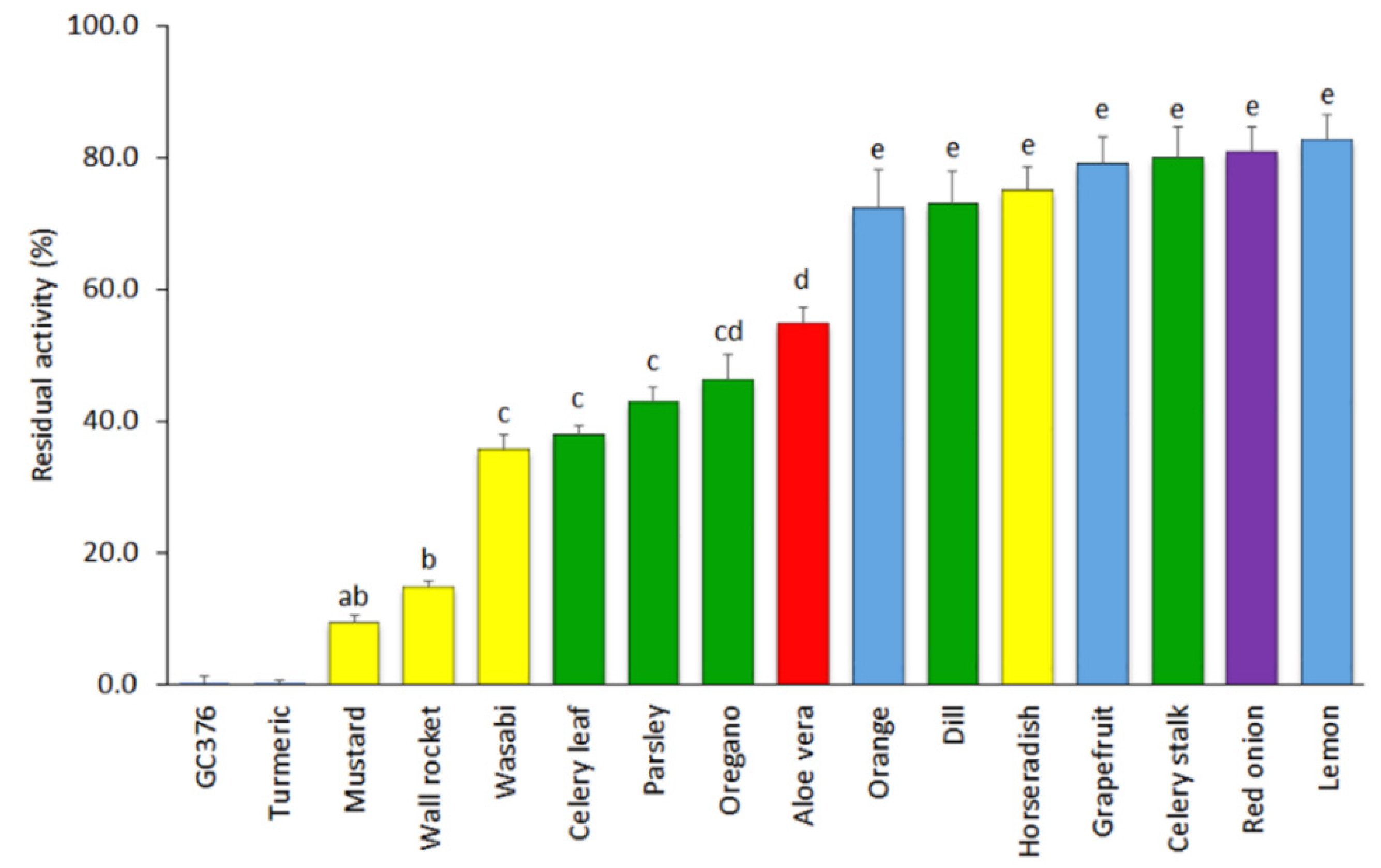
The results of the inhibitory activity of SARS-CoV-2 3CLPro of the remaining 15 extracts tested at a concentration of 500 µg mL−1 are displayed in Figure 1. Seven of the materials showed low inhibitory capacity and led to average residual protease activities over 70%. This group included extracts of grapefruit, lemon, and orange fruit peels (72.4–82.8% of mean residual activities); red onion (80.9%); celery stalk (79.9%); horseradish (75.0%); and dill (73.1%). Five other materials displayed intermediate inhibitory capacity (35–55%), including extracts of celery leaves (38.0%), parsley (42.8%), and oregano (46.3%) herbs; aloe vera leaves (54.8%); and the wasabi powder (35.8%). Finally, three extracts were found to show high inhibitory capacity and included mustard seeds (9.4%), wall rocket (14.9%), and turmeric, the latter showing a complete inhibitory capacity (0.0% of 3CLPro residual activity) at the concentration of 500 µg mL−1.
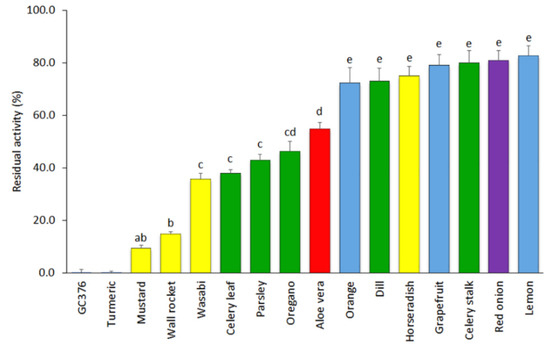
Figure 1.
Mean values and standard errors (SEs) for the residual activity of SARS-CoV-2 3CLPro in the presence of the fifteen plant-based extracts at a concentration of 500 µg mL−1 and the inhibitor control GC376 at a concentration of 50 µM (n = 4). Extracts included were obtained from citrus fruit peels (blue), seasoning and aromatic herbs (green), bulbs and rhizomes (purple; includes turmeric), a succulent plant (red), and cruciferous herbs and condiments (yellow). Both the turmeric extract and the inhibitor GC376 produced a complete inhibition of the 3CLPro. Different letters indicate significant differences in the 3CLPro residual activity according to a Student–Newman–Keuls test at p < 0.05.
3.2. Determination of the IC50 Values of Extracts with High SARS-CoV-2 3CLPro Activity Inhibition
According to the initial screening at 500 µg mL−1, the mustard seeds, wall rocket leaves, and turmeric powder extracts were selected owing to their high SARS-CoV-2 3CLPro activity inhibitory potential. For these materials, the dose-dependent inhibitory curves were obtained and the corresponding IC50 values were calculated by testing the plant extracts at eight final concentrations ranging between 5.0 and 500 µg mL−1.
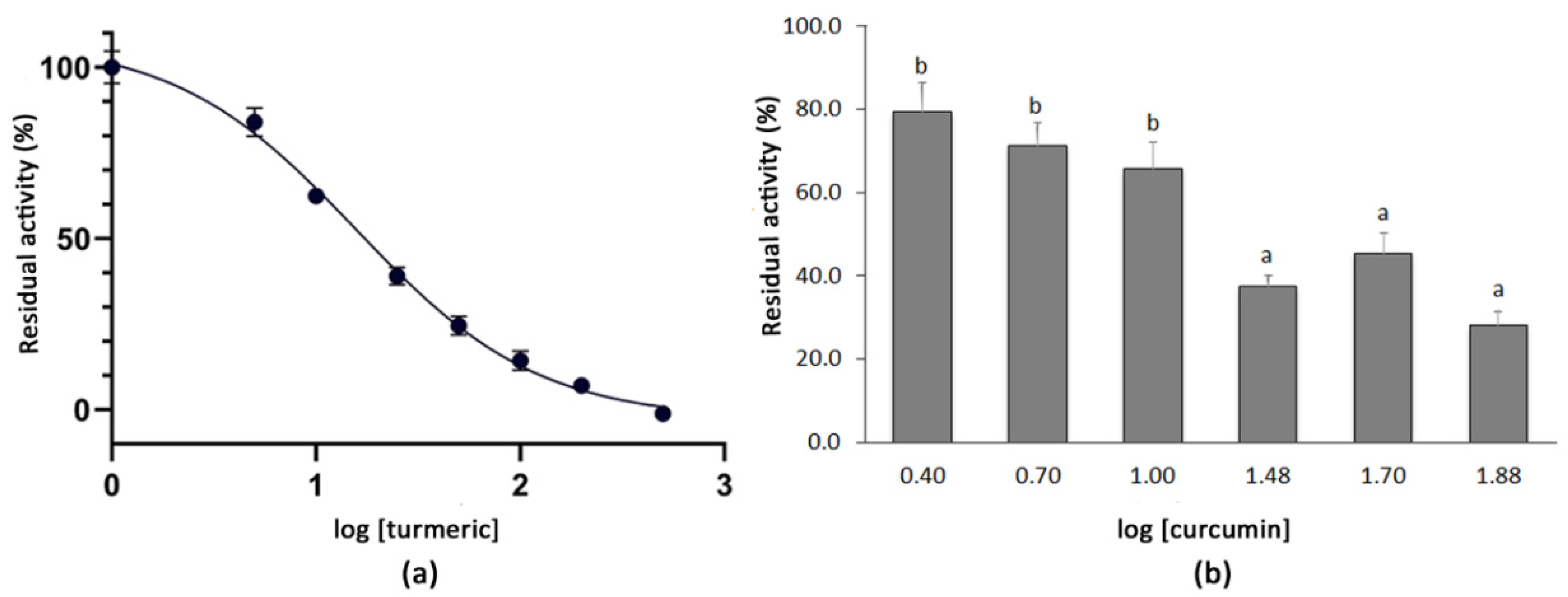
The dose-dependent inhibition curve for the turmeric powder was obtained by testing the residual activity of the SARS-CoV-2 3CLPro after the co-incubation with the plant extracts at the final concentrations of 5.0–10–25–50–100–200–500 µg mL−1. The logarithmic dose–response curve was fitted to a sigmoidal curve with an r2 value of 0.976 and an IC50 for turmeric of 15.74 µg mL−1 was determined. The highest concentrations tested, i.e., from 100 µg mL−1 (log [turmeric] = 2.0) to 500 µg mL−1 (log [turmeric] = 2.7), resulted in a very high inhibition of the SARS-CoV-2 3CLPro activity (Figure 2). In order to test whether the curcumin present in the turmeric material accounted for such inhibition, an additional analysis using commercial curcumin (>94% curcuminoids, >80% curcumin) was conducted at concentrations of 2.5–75 µg mL−1 (log [curcumin] = 0.4–1.9). Higher concentrations of the compound could not be tested because of its insolubility in the reaction buffer. As expected, the co-incubation of 3CLPro with curcumin also showed a positive effect on the protease inhibition. Thus, concentrations over 30 µg mL−1 (log [curcumin] = 1.5) produced a residual protease activity below 50%, which decreased to 28.1% when the compound was tested at 75 µg mL−1 (log [curcumin] = 1.9) (Figure 2).
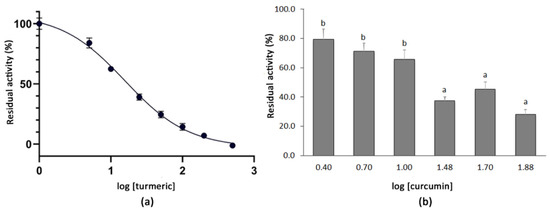
Figure 2.
(a) Dose–response curve for the methanolic extract of turmeric powder. Each point represents the average value and SE for the residual activity of the 3CLPro (%) at extract concentrations between 5.0 and 500 µg mL−1 (n = 4). The 100% of residual activity considers the activity of the 3CLPro when no inhibitor is added. (b) Mean residual activity and SE of 3CLPro (%) determined after the co-incubation with five different concentrations of commercial curcumin (2.5–75 µg mL−1) (n = 4). Different letters indicate significant differences in the 3CLPro residual activity according to a Student–Newman–Keuls test at p < 0.05.
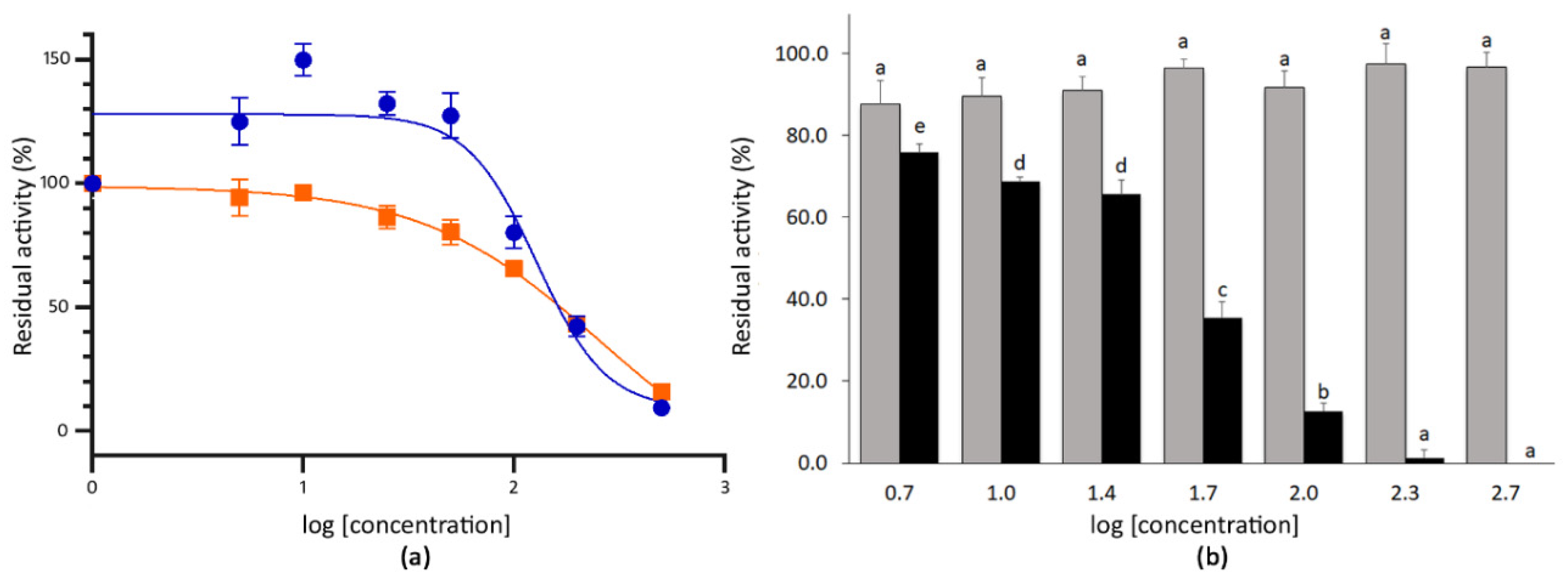
The extracts of mustard seeds showed strong inhibitory capacity at the highest doses tested. Thus, the co-incubation with 200 µg mL−1 (log (mustard) = 2.3) and 500 µg mL−1 (log (mustard) = 2.7) of the extract reduced the activity of SARS-CoV-2 3CLPro to 42.1% and 9.4%, respectively (Figure 3). By contrast, the co-incubation with 100 µg mL−1 (log (mustard) = 2.0) of mustard seeds extract only inhibited such activity by 19.9% (residual activity = 80.1%). The IC50 for the mustard seeds extract was 128.1 µg mL−1 (r2 value 0.865) (Figure 3). In the case of wall rocket extracts, the co-incubation produced residual activities of the 3CLPro ranging between 94.3% and 15.8% for 5.0 µg mL−1 (log (wall rocket) = 0.7) and 500 µg mL−1 (log (wall rocket) = 2.7) of extract, respectively. The co-incubation with 200 µg mL−1 of extract produced an inhibitory response of 56.8% compared with the absence of extract. For this material, the IC50 estimated from the dose–response curve was 257.4 µg mL−1 (r2 value 0.937) (Figure 3).
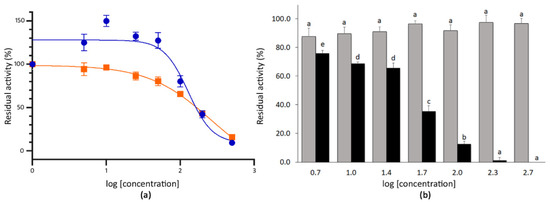
Figure 3.
(a) Dose–response curve for the methanolic extracts of mustard seeds (blue circles) and wall rocket leaves (orange squares). Each point represents the average value and SE for the residual activity of the 3CLPro (%) at extract concentrations between 5.0 and 500 µg mL−1 (n = 4). The 100% of residual activity is the activity of the 3CLPro when no inhibitor is added. (b) Mean residual activity and SE of 3CLPro (%) determined after the co-incubation with 5.0–500 µg mL−1 of sinigrin (grey) and allyl isothiocyanate (black) standards (n = 4). Different letters indicate significant differences in the 3CLPro residual activity for sinigrin or allyl isothiocyanate treatments according to two separate Student–Newman–Keuls tests at p < 0.05.
Given that both mustard and wall rocket were selected for containing glucosinolates, particularly sinigrin, both sinigrin and its hydrolytic derivative were evaluated for their SARS-CoV-2 3CLPro inhibition activity. Allyl isothiocyanate is naturally released in plants from its precursor owing to the activity of the endogenous plant myrosinases [66]. The co-incubation of the 3CLPro with the purified sinigrin did not result in a significant reduction of the protease activity even at the highest concentration tested, 500 µg mL−1. By contrast, the co-incubation with allyl isothiocyanate at 500 µg mL−1 (log (allyl isothiocyanate) = 2.7) resulted in a complete inhibition of the 3CLPro activity. Therefore, the dose–response curve was obtained for allyl isothiocyanate. The response fitted to a sigmoidal curve (r2 value 0.943) with an estimated IC50 of 41.43 µg mL−1 allyl isothiocyanate standard (Figure 3).
4. Discussion
The recent COVID-19 disease pandemic caused by the SARS-CoV-2 coronavirus has resulted in a global health crisis. Although other coronaviruses were responsible for previous outbreaks in the last two decades [67], the fast spread of SARS-CoV-2 has demonstrated the threat to human health of new coronaviruses. One year after the disease was declared as a pandemic [68], COVID-19 is still causing thousands of daily deaths and no effective treatments apart from preventive vaccination are reported for fighting the infection process. Obtaining effective treatments against SARS-CoV-2 is urgently needed because, in combination with vaccines, they may become a first line therapy not only for current SARS-CoV-2 strains, but also for new ones.
The 3CLPro is considered as one of the most promising targets for the development of antiviral drugs against coronaviruses [69]. The three-dimensional structure of SARS-CoV-2 3CLPro was unraveled by Zhang et al. [12]. The enzyme has an identity percentage of 96% with SARS-CoV 3CLPro, thus it has been inferred that both proteases have very similar specificities [70,71]. These works suggest that molecules and extracts that displayed inhibitory activity against SARS-CoV 3CLPro might be also of interest to inhibit the activity of SARS-CoV-2 3CLPro. In this context, our work used a targeted approach based on previous information on inhibitors SARS-CoV 3CLPro [22]. For the current study, plant-based extracts potentially promising for the inhibition of SARS-CoV-2 3CLPro activity were tested. Previous studies with plant-based extracts have been conducted in the past to test their inhibitory capacity against SARS-CoV 3CLPro, with promising results [15,16,72].
The first screening with plant extracts at 500 µg mL−1 revealed that the extracts of the three citrus fruits peels produced a low reduction of the protease activity. Flavonoids present in citrus fruits have been suggested as potential antivirals against coronaviruses and in therapeutics fighting the inflammation process that can derive in more severe symptoms of the viral disease [73,74]. Recent works based on computational molecular docking pointed to the flavanone hesperidin, which is the glycosylated form of hesperetin and the main form in which this flavonoid is found in citrus fruits, as one natural compound displaying high capacity for binding to the SARS-CoV-2 3CLPro [71,75]. However, our results showed that citrus fruit peels, which contain high concentrations of hesperidin [44], had a low in vitro inhibitory activity against SARS-CoV-2 3CLPro.
Interestingly, extracts of turmeric powder, mustard seeds, and wall rocket leaves produced the highest reduction in SARS-CoV-2 3CLPro activity. Turmeric rhizomes are rich in the flavonoid curcumin and its derivatives, natural compounds that have been also considered in molecular docking studies for their potential inhibitory capacity against SARS-CoV-2 3CLPro [75,76,77]. According to these studies, curcumin and its derivatives can be considered as natural compounds with potential as 3CLPro inhibitors, although the same works showed that other flavonoids might have higher potential inhibitory activity according to the docking results. In concordance, our FRET assay revealed a high inhibitory capacity of the turmeric rhizomes extracts against the 3CLPro activity. The turmeric extracts were, in fact, the most effective among all the plant-based extracts evaluated in the inhibitory activity. In this respect, other plant-based extracts were evaluated in the past against SARS-CoV 3CLPro activity and showed lower inhibitory capacity. Ryu et al. [72] found that ethanolic extracts of Torreya nucifera tested at 100 µg mL−1 resulted in 62% of activity inhibition against SARS-CoV 3CLPro, while in our case, the same concentration of turmeric extract resulted in a reduction of 86% for SARS-CoV-2 3CLPro activity. In another study, an IC50 was determined for SARS-CoV 3CLPro activity that ranged between 25 and 70 µg mL−1 for black and Puer tea extracts [15], while for the turmeric extracts, we found an IC50 of 1.6- to 4.5-fold lower than these against SARS-CoV-2 3CLPro.
The concentration of curcumin in turmeric powder is generally around 3% or lower [78], while the concentration of other curcuminoids is even lower [79]. However, commercial curcumin (>80% curcumin) even at concentrations of 75 µg mL−1, which is several-fold the expected concentration of curcumin in the turmeric extract at 500 µg mL−1, did not cause a complete inhibition of SARS-CoV-2 3CLPro activity. This result suggests that other components of turmeric or their synergetic interaction may have a major contribution to the inhibition of SARS-CoV-2 3CLPro activity. In this sense, turmeric rhizomes also contain the curcuminoid demethoxycurcumin [56], which has also been described as a potential inhibitor against SARS-CoV-2 3CLPro, even showing greater affinity than curcumin for SARS-CoV-2 3CLPro in the docking analysis of Khaerunnisa et al. [77]. Furthermore, turmeric powder also contains sesquiterpenoids [79] and quercetin derivatives [80], the latter also being considered as a potential inhibitor of SARS-CoV-2 3CLPro [77,81]. Thus, the inhibitor capacity of turmeric extracts might be the result of a synergistic activity of several compounds. Overall, our results indicate that extracts of turmeric are a strong candidate for being tested for inhibiting the in vivo replication of SARS-CoV-2. Moreover, considering these results, further studies should be addressed to characterize the methanolic extracts of turmeric powder and to individually test the compounds present.
On the other hand, two cruciferous extracts (from mustard seeds and wall rocket) that are rich in sinigrin [36,57,58,61] were found to exert a strong inhibition of SARS-CoV-2 3CLPro activity. Sinigrin is the precursor glucosinolate of allyl isothiocyanate, a reactive molecule displaying antimicrobial [82,83] and anticarcinogenic properties [84,85,86]. By contrast, references to the potential of either sinigrin or allyl isothiocyanate as antivirals are more limited [16]. These latter authors found that sinigrin had good potential as an inhibitor against SARS-CoV 3CLPro. In our case, we found that sinigrin has a low effect as in vitro inhibitor of SARS-CoV-2 3CLPro, while its derivative allyl isothiocyanate is a powerful inhibitor of SARS-CoV-2 3CLPro activity. In this way, isothiocyanate metabolites might be responsible, at least in part, for the antiviral properties of these cruciferous plants extracts, which is probably related to their chemical structure, i.e., organosulfur molecules acting as H2S donors [87]. In fact, Blanchard et al. [14] previously reported a good response of sulfur-containing compounds against SARS-CoV 3CLPro activity owing to their capacity of forming covalent adducts between their electrophilic group and a nucleophile side chain of the protease. Therefore, our results suggest that the sinigrin derivative allyl isothiocyanate is a good candidate molecule for being tested in vivo for inhibiting SARS-CoV-2 3CLPro activity. However, the high cytotoxicity of allyl isothiocyanate may preclude its practical use [83], which would depend on the concentration required. In this sense, further analysis of the methanolic extracts for mustard and wall rocket would increase the knowledge regarding the concentration at which these compounds are found, and the possible presence of other metabolites also exerting potential inhibitory activities.
This work is a first targeted approach for the evaluation of plant extracts and containing natural compounds for the development of prophylaxis, adjuvant therapies, and drug treatments aimed at inhibiting the activity of SARS-CoV-2 3CLPro. Extracts of turmeric have been identified as a candidate plant extract for reducing SARS-CoV-2 3CLPro activity, which could eventually affect the viral replication. In addition, other plant extracts, such as those of cruciferous plants containing sinigrin, which, after consumption, is degraded to allyl isothiocyanate, might be considered as well for their inhibition of SARS-CoV-2 3CLPro activity. The current study provides information that can help in the search for treatments against COVID-19, acting as a basis for future chemical, in vivo, and clinical trials. Thus, further studies following the results of this work should be addressed: (1) to chemically characterize the plant extracts with high potential inhibitory activity and to re-evaluate this capacity after the fractionation of the extracts, thus allowing the identification of the biomolecules responsible of such activity and possible synergistic effects; and (2) to evaluate this potential inhibition in cell-based studies where the virus, host cell, and plant extract interact, also determining the toxicity limit prior to conducting other pre-clinical and clinical trials.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/foods10071503/s1, Figure S1: Materials used for the analysis, from left to right and top to bottom: orange peel, lemon peel, lime peel, grapefruit peel, celery leaves, celery stalk, parsley leaves, dill leaves, dried oregano leaves, red onion bulb, turmeric rhizomes, aloe leaf, chamomile, mustard seeds, horseradish root, wasabi powder, and wall rocket leaves; Figure S2: Signal recorded at wavelengths between 400 and 550 nm for the residual activity of the 3CLPro without inhibition (Control), after co-incubation with chamomile extracts at concentrations of 200 µg mL−1 or 500 µg mL−1, after co-incubation with the inhibitor GC376 at 50 µM (GC376), and the signal of the raw chamomile extract at a concentration of 500 µg mL−1 without incubation with 3CLPro (Raw).
Author Contributions
Conceptualization, J.P. and A.F.; methodology, C.G.-R., M.P. and A.F.; validation, C.G.-R., M.P. and A.F.; formal analysis, C.G.-R., A.R.-B. and J.P.; investigation, C.G.-R. and M.P.; resources, A.R.-B. and J.P.; data curation, C.G.-R.; writing—original draft preparation, C.G.-R., A.R.-B. and A.F.; writing—review and editing, A.R.-B. and J.P.; supervision, J.P. and A.F.; project administration, A.R.-B. and J.P.; funding acquisition, A.R.-B. and J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article or Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Salian, V.S.; Wright, J.A.; Vedell, P.T.; Nair, S.; Li, C.; Kandimalla, M.; Tang, X.; Porquera, E.M.C.; Kalari, K.R.; Kandimalla, K.K. COVID-19 transmission, current treatment, and future therapeutic strategies. Mol. Pharm. 2021, 18, 754–771. [Google Scholar] [CrossRef]
- World Health Organization. Coronavirus Disease (COVID-19) Weekly Epidemiological Update and Weekly Operational Update. Coronavirus Disease (COVID-19). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 8 April 2021).
- Astuti, I.; Ysrafil. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An overview of viral structure and host response. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Hoque, M.N.; Rahman, M.S.; Ul Alam, A.S.M.R.; Akther, M.; Puspo, J.A.; Akter, S.; Sultana, M.; Crandall, K.A.; Hossain, M.A. Genome-wide analysis of SARS-CoV-2 virus strains circulating worldwide implicates heterogeneity. Sci. Rep. 2020, 10, 14004. [Google Scholar] [CrossRef]
- Laamarti, M.; Alouane, T.; Kartti, S.; Chemao-Elfihri, M.W.; Hakmi, M.; Essabbar, A.; Laamarti, M.; Hlali, H.; Bendani, H.; Boumajdi, N.; et al. Large scale genomic analysis of 3067 SARS-CoV-2 genomes reveals a clonal geodistribution and a rich genetic variations of hotspots mutations. PLoS ONE 2020, 15, e0240345. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, J.; Gao, K.; Hozumi, Y.; Yin, C.; Wei, G.-W. Analysis of SARS-CoV-2 mutations in the United States suggests presence of four substrains and novel variants. Commun. Biol. 2021, 4, 228. [Google Scholar] [CrossRef]
- Forni, G.; Mantovani, A.; On Behalf of the COVID-19 Commission of Accademia Nazionale dei Lincei, Rome. COVID-19 vaccines: Where we stand and challenges ahead. Cell Death Differ. 2021, 28, 626–639. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. SARS-CoV-2-Increased Circulation of Variants of Concern and Vaccine Rollout in the EU/EEA-14th Update; ECDC: Stockholm, Sweden, 2021; pp. 1–29.
- Centers for Disease Control and Prevention. Interim Clinical Considerations for Use of COVID-19 Vaccines Currently Authorized in the United States. Available online: https://www.cdc.gov/vaccines/covid-19/info-by-product/clinical-considerations.html (accessed on 8 March 2021).
- World Health Organization. The Moderna COVID-19 (mRNA-1273) Vaccine: What You Need to Know. Available online: https://www.who.int/news-room/feature-stories/detail/the-moderna-covid-19-mrna-1273-vaccine-what-you-need-to-know (accessed on 8 March 2021).
- World Health Organization. COVID-19 Clinical Management. Living Guidance 25 January 2021; No. WHO/2019-nCoV/clinical/2021.1; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved a-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Liu, D.; Yang, Y.; Guo, J.; Feng, Y.; Zhang, X.; Cheng, S.; Feng, J. Phylogenetic supertree reveals detailed evolution of SARS-CoV-2. Sci. Rep. 2020, 10, 22366. [Google Scholar] [CrossRef]
- Blanchard, J.E.; Elowe, N.H.; Huitema, C.; Fortin, P.D.; Cechetto, J.D.; Eltis, L.D.; Brown, E.D. High-throughput screening identifies inhibitors of the SARS Coronavirus Main Proteinase. Chem. Biol. 2004, 11, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.N.; Lin, C.P.C.; Huang, K.K.; Chen, W.C.; Hsieh, H.P.; Liang, P.H.; Hsu, J.T.A. Inhibition of SARS-CoV 3C-like protease activity by theaflavin-3,3′- digallate (TF3). Evid. Based Complement. Altern. Med. 2005, 2, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-W.; Tsai, F.-J.; Tsai, C.-H.; Lai, C.-C.; Wan, L.; Ho, T.-Y.; Hsieh, C.; Chao, P.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Xian, Y.; Zhang, J.; Bian, Z.; Zhou, H.; Zhang, Z.; Lin, Z.; Xu, H. Bioactive natural compounds against human coronaviruses: A review and perspective. Acta Pharm. Sin. B 2020, 10, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Antonio, A.; Wiedemann, L.S.M.; Veiga-Junior, V.F. Natural products’ role against COVID-19. RSC Adv. 2020, 10, 23379–23393. [Google Scholar] [CrossRef]
- Vougogiannopoulou, K.; Corona, A.; Tramontano, E.; Alexis, M.N.; Skaltsounis, A.-L. Natural and nature-derived products targeting human coronaviruses. Molecules 2021, 26, 448. [Google Scholar] [CrossRef] [PubMed]
- Romeo, I.; Mesiti, F.; Lupia, A.; Alcaro, S. Current updates on naturally occurring compounds recognizing SARS-CoV-2 druggable targets. Molecules 2021, 26, 632. [Google Scholar] [CrossRef] [PubMed]
- El-Missiry, M.A.; Fekri, A.; Kesar, L.A.; Othman, A.I. Polyphenols are potential nutritional adjuvants for targeting COVID-19. Phyther. Res. 2020, 35, 2879–2889. [Google Scholar] [CrossRef]
- Zhou, J.; Huang, J. Current Findings Regarding Natural Components with Potential Anti-2019-nCoV Activity. Front. Cell Dev. Biol. 2020, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Ferreira, I.C. In vivo antioxidant activity of phenolic compounds: Facts and gaps. Trends Food Sci. Technol. 2016, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Godos, J.; Caraci, F.; Castellano, S.; Currenti, W.; Galvano, F.; Ferri, R.; Grosso, G. Association between dietary flavonoids intake and cognitive function in an Italian cohort. Biomolecules 2020, 10, 1300. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zhang, Y.; Talalay, P. Broccoli sprouts: An exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc. Natl. Acad. Sci. USA 1997, 94, 10367–10372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Liu, Y.; Ma, L.; Ji, R.; Qu, Y.; Xin, Y.; Lv, G. Chemopreventive activity of sulforaphane. Drug Des. Dev. Ther. 2018, 12, 2905–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Chillón, M.T.; Carazo-Díaz, C.; Prieto-Merino, D.; Zafrilla, P.; Moreno, D.A.; Villaño, D. Effects of long-term consumption of broccoli sprouts on inflammatory markers in overweight subjects. Clin. Nutr. 2019, 38, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Schepici, G.; Bramanti, P.; Mazzon, E. Efficacy of sulforaphane in neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 8637. [Google Scholar] [CrossRef]
- Lin, C.; Kim, J.; Du, W.; Wei, C. Bactericidal Activity of Isothiocyanate against Pathogens on Fresh Produce. J. Food Prot. 2000, 63, 25–30. [Google Scholar] [CrossRef]
- Kaiser, S.J.; Mutters, N.T.; Blessing, B.; Günther, F. Natural isothiocyanates express antimicrobial activity against developing and mature biofilms of Pseudomonas aeruginosa. Fitoterapia 2017, 119, 57–63. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, V.; Chao, T.-C.; Hsiao, C.-D.; Lin, A.; Chang, M.-F.; Chow, L.-P. Screening of drugs by FRET analysis identifies inhibitors of SARS-CoV 3CL protease. Biochem. Biophys. Res. Commun. 2005, 333, 194–199. [Google Scholar] [CrossRef]
- Kuo, C.-J.; Chi, Y.-H.; Hsu, J.T.-A.; Liang, P.-H. Characterization of SARS main protease and inhibitor assay using a fluorogenic substrate. Biochem. Biophys. Res. Commun. 2004, 318, 862–867. [Google Scholar] [CrossRef]
- Park, J.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Guijarro-Real, C.; Adalid-Martínez, A.M.; Aguirre, K.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Growing conditions affect the phytochemical composition of edible wall rocket (Diplotaxis erucoides). Agronomy 2019, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Guijarro-Real, C.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Consumers acceptance and volatile profile of wall rocket (Diplotaxis erucoides). Food Res. Int. 2020, 132, 109008. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I. Review of the flavonoids quercetin, hesperetin, and naringenin. Dietary sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 2004, 24, 851–874. [Google Scholar] [CrossRef]
- Manthey, J.A.; Grohmann, K. Concentrations of hesperidin and other orange peel flavonoids in citrus processing byproducts. J. Agric. Food Chem. 1996, 44, 811–814. [Google Scholar] [CrossRef]
- Li, Y.; Kandhare, A.D.; Mukherjee, A.A.; Bodhankar, S.L. Acute and sub-chronic oral toxicity studies of hesperidin isolated from orange peel extract in Sprague Dawley rats. Regul. Toxicol. Pharmacol. 2019, 105, 77–85. [Google Scholar] [CrossRef]
- Garcia-Castello, E.M.; Rodriguez-Lopez, A.D.; Mayor, L.; Ballesteros, R.; Conidi, C.; Cassano, A. Optimization of conventional and ultrasound assisted extraction of flavonoids from grapefruit (Citrus paradisi L.) solid wastes. LWT Food Sci. Technol. 2015, 64, 1114–1122. [Google Scholar] [CrossRef]
- Zhang, M.; Duan, C.; Zang, Y.; Huang, Z.; Liu, G. The flavonoid composition of flavedo and juice from the pummelo cultivar (Citrus grandis (L.) Osbeck) and the grapefruit cultivar (Citrus paradisi) from China. Food Chem. 2011, 129, 1530–1536. [Google Scholar] [CrossRef]
- Xi, W.; Lu, J.; Qun, J.; Jiao, B. Characterization of phenolic profile and antioxidant capacity of different fruit part from lemon (Citrus limon Burm.) cultivars. J. Food Sci. Technol. 2017, 54, 1108–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Mejía, E.; Rosales-Conrado, N.; León-González, M.E.; Madrid, Y. Citrus peels waste as a source of value-added compounds: Extraction and quantification of bioactive polyphenols. Food Chem. 2019, 295, 289–299. [Google Scholar] [CrossRef]
- De la Rosa, J.D.P.; Ruiz-Palomino, P.; Arriola-Guevara, E.; García-Fajardo, J.; Sandoval, G.; Guatemala-Morales, G.M. A green process for the extraction and purification of hesperidin from mexican lime peel (Citrus aurantifolia Swingle) that is extendible to the citrus genus. Processes 2018, 6, 266. [Google Scholar] [CrossRef] [Green Version]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid composition of fruit tissues of citrus species. Biosci. Biotechnol. Biochem. 2006, 70, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guijarro-Real, C.; Prohens, J.; Rodriguez-Burruezo, A.; Adalid-Martínez, A.M.; López-Gresa, M.P.; Fita, A. Wild edible fool’s watercress, a potential crop with high nutraceutical properties. PeerJ 2019, 7, e6296. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Lean, M.E.J.; McDonald, M.S.; Black, C. Quantitative Analysis of the Flavonoid Content of Commercial Tomatoes, Onions, Lettuce, and Celery. J. Agric. Food Chem. 1997, 45, 590–595. [Google Scholar] [CrossRef]
- Justesen, U.; Knuthsen, P.; Leth, T. Quantitative analysis of flavonols, flavones, and flavanones in fruits, vegetables and beverages by high performance liquid chromatography with photo-diode array and mass spectrometric detection. J. Chromatogr. A 1998, 799, 101–110. [Google Scholar] [CrossRef]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, A.C. Characterization of phenolic composition in lamiaceae spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Santos-Buelga, C.; Henriques, M.; Silva, S.; Ferreira, I.C. Decoction, infusion and hydroalcoholic extract of Origanum vulgare L.: Different performances regarding bioactivity and phenolic compounds. Food Chem. 2014, 158, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Justesen, U.; Knuthsen, P. Composition of flavonoids in fresh herbs and calculation of flavonoid intake by use of herbs in traditional Danish dishes. Food Chem. 2001, 73, 245–250. [Google Scholar] [CrossRef]
- Srivastava, J.K.; Gupta, S. Extraction, Characterization, Stability and Biological Activity of Flavonoids Isolated from Chamomile Flowers. Mol. Cell. Pharmacol. 2009, 1, 138. [Google Scholar] [CrossRef]
- Svehlıkova, V.; Bennett, R.N.; Mellon, F.A.; Needs, P.W.; Piacente, S.; Kroon, P.A.; Bao, Y. Isolation, identification and stability of acylated derivatives of apigenin 7-O-glucoside from chamomile (Chamomilla recutita [L.] Rauschert). Phytochemistry 2004, 65, 2323–2332. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Cheng, L.Z.; Zhang, T.; Yaron, S.; Jiang, H.X.; Sui, Z.Q.; Corke, H. Phenolic profiles, antioxidant, and antiproliferative activities of turmeric (Curcuma longa). Ind. Crops Prod. 2020, 152, 112561. [Google Scholar] [CrossRef]
- Sepahpour, S.; Selamat, J.; Manap, M.Y.A.; Khatib, A.; Razis, A.F.A. Comparative analysis of chemical composition, antioxidant activity and quantitative characterization of some phenolic compounds in selected herbs and spices in different solvent extraction systems. Molecules 2018, 23, 402. [Google Scholar] [CrossRef] [Green Version]
- Al Shahawany, A.W.; Al Hattab, Z.N.; Al Tahhan, S.F. Qualitative and Quantitative Analysis of Sinigrin in Different Parts In Vitro and In Vivo of Brassica nigra Plants. Biomed. Biotechnol. 2016, 4, 19–24. [Google Scholar] [CrossRef]
- Rangkadilok, N.; Nicolas, M.E.; Bennett, R.N.; Premier, R.R.; Eagling, D.R.; Taylor, P. Developmental changes of sinigrin and glucoraphanin in three Brassica species (Brassica nigra, Brassica juncea and Brassica oleracea var. italica). Sci. Hortic. 2002, 96, 11–26. [Google Scholar] [CrossRef]
- Agneta, R.; Lelario, F.; De Maria, S.; Möllers, C.; Bufo, S.A.; Rivelli, A.R. Glucosinolate profile and distribution among plant tissues and phenological stages of field-grown horseradish. Phytochemistry 2014, 106, 178–187. [Google Scholar] [CrossRef]
- Popović, M.; Maravić, A.; Čulić, V.Č.; Đulović, A.; Burčul, F.; Blažević, I. Biological effects of glucosinolate degradation products from horseradish: A horse that wins the race. Biomolecules 2020, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Di Gioia, F.; Avato, P.; Serio, F.; Argentieri, M.P. Glucosinolate profile of Eruca sativa, Diplotaxis tenuifolia and Diplotaxis erucoides grown in soil and soilless systems. J. Food Compos. Anal. 2018, 69, 197–204. [Google Scholar] [CrossRef]
- Borges-Argáez, R.; Chan-Balan, R.; Cetina-Montejo, L.; Ayora-Talavera, G.; Sansores-Peraza, P.; Gómez-Carballo, J.; Cáceres-Farfán, M. In vitro evaluation of anthraquinones from Aloe vera (Aloe barbadensis Miller) roots and several derivatives against strains of influenza virus. Ind. Crops Prod. 2019, 132, 468–475. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakasha, G.K.; Jifon, J.; Patil, B.S. Extraction efficiency and validation of an HPLC method for flavonoid analysis in peppers. Food Chem. 2012, 130, 751–758. [Google Scholar] [CrossRef]
- Selmi, S.; Rtibi, K.; Grami, D.; Sebai, H.; Marzouki, L. Protective effects of orange (Citrus sinensis L.) peel aqueous extract and hesperidin on oxidative stress and peptic ulcer induced by alcohol in rat. Lipids Health Dis. 2017, 16, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doheny-Adams, T.; Redeker, K.; Kittipol, V.; Bancroft, I.; Hartley, S.E. Development of an efficient glucosinolate extraction method. Plant Methods 2017, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, R.; Yu, Q.; Potter, J.; Chiba, M. Direct and simultaneous analysis of sinigrin and allyl isothiocyanate in mustard samples by high-performance liquid chromatography. J. Agric. Food Chem. 2002, 50, 4749–4753. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Coronavirus Disease 2019 (COVID-19): Situation Report-94; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Hilgenfeld, R. From SARS to MERS: Crystallographic studies on coronaviral proteases enable antiviral drug design. FEBS J. 2014, 281, 4085–4096. [Google Scholar] [CrossRef] [Green Version]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M. Unravelling lead antiviral phytochemicals for the inhibition of SARS-CoV-2 M pro enzyme through in silico approach. Life Sci. 2020, 255, 117831. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Yiu, C.P.B.; Wong, K.Y. Prediction of the SARS-CoV-2 (2019-nCoV) 3C-like protease (3CLpro) structure: Virtual screening reveals velpatasvir, ledipasvir, and other drug repurposing candidates. F1000Research 2020, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Naguyen, T.T.H.; Park, S.J.; Chang, J.S.; Park, K.H.; et al. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef]
- Cheng, L.; Zheng, W.; Li, M.; Huang, J.; Bao, S.; Xu, Q.; Ma, Z. Citrus fruits are rich in flavonoids for immunoregulation and potential targeting ACE2. Preprints 2020. [Google Scholar] [CrossRef]
- Utomo, R.Y.; Ikawati, M.; Meiyanto, E. Revealing the Potency of Citrus and Galangal Constituents to Halt SARS-CoV-2 Infection. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Adem, S.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Ali, M. Identification of potent COVID-19 main protease (Mpro) inhibitors from natural polyphenols: An in silico strategy unveils a hope against CORONA. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Pendyala, B.; Patrasa, A. In silico screening of food bioactive compounds to predict potential inhibitors of COVID-19 Main protease (Mpro) and RNA-dependent RNA polymerase (RdRp). ChemRxiv 2020. [Google Scholar] [CrossRef]
- Khaerunnisa, S.; Kurniawan, H.; Awaluddin, R.; Suhartati, S.; Soetjipto, S. Potential inhibitor of COVID-19 Main Protease (Mpro) from several medicinal plant compounds by molecular docking study. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Tayyem, R.F.; Heath, D.D.; Al-Delaimy, W.K.; Rock, C.L. Curcumin content of turmeric and curry powders. Nutr. Cancer 2006, 55, 126–131. [Google Scholar] [CrossRef]
- Chao, I.-C.; Wang, C.-M.; Li, S.-P.; Lin, L.-G.; Ye, W.-C.; Zhang, Q.-W. Simultaneous quantification of three curcuminoids and three volatile components of Curcuma longa using pressurized liquid extraction and high-performance liquid chromatography. Molecules 2018, 23, 1568. [Google Scholar] [CrossRef] [Green Version]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef]
- Abian, O.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Vega, S.; Reyburn, H.T.; Rizzuti, B.; Velazquez-Campoy, A. Structural stability of SARS-CoV-2 3CLpro and identification of quercetin as an inhibitor by experimental screening. Int. J. Biol. Macromol. 2020, 164, 1693–1703. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Abu Ghoush, M.; Al-Nabulsi, A.A.; Holley, R.A. Control of Salmonella enterica and Listeria monocytogenes in hummus using allyl isothiocyanate. Int. J. Food Microbiol. 2018, 278, 73–80. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Ferreira, C.; Saavedra, M.J.; Simões, L.C.; Simões, M. Antibacterial activity and mode of action of selected glucosinolate hydrolysis products against bacterial pathogens. J. Food Sci. Technol. 2015, 52, 4737–4748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, D.; Srivastava, S.K.; Lew, K.L.; Zeng, Y.; Hershberger, P.; Johnson, C.S.; Trump, D.L.; Singh, S.V. Allyl isothiocyanate, a constituent of cruciferous vegetables, inhibits proliferation of human prostate cancer cells by causing G2/M arrest and inducing apoptosis. Carcinogenesis 2003, 24, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.K.; Xiao, D.; Lew, K.L.; Hershberger, P.; Kokkinakis, D.M.; Johnson, C.S.; Trump, D.L.; Singh, S.V. Allyl isothiocyanate, a constituent of cruciferous vegetables, inhibits growth of PC-3 human prostate cancer xenograft in vivo. Carcinogenesis 2003, 24, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Sávio, A.L.V.; da Silva, G.N.; Salvadori, D.M.F. Inhibition of bladder cancer cell proliferation by allyl isothiocyanate (mustard essential oil). Mutat. Res. 2015, 771, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citi, V.; Martelli, A.; Brancaleone, V.; Brogi, S.; Gojon, G.; Montanaro, R.; Morales, G.; Testai, L.; Calderone, V. Anti-inflammatory and antiviral roles of hydrogen sulfide: Rationale for considering H2S donors in COVID-19 therapy. Br. J. Pharmacol. 2020, 177, 4931–4941. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).