
Yeast Particle Encapsulation of Scaffolded Terpene Compounds for Controlled Terpene Release

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
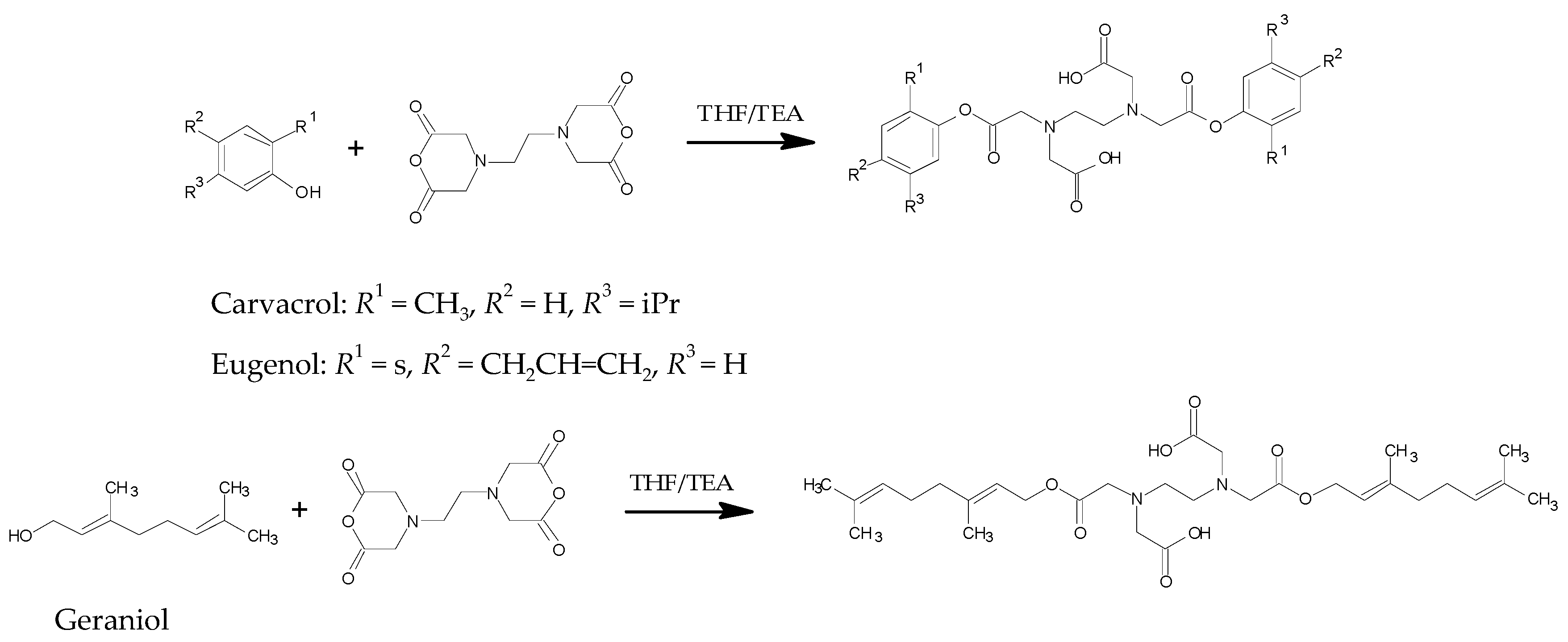
2.2.1. Synthesis of Pro-Terpene Compounds
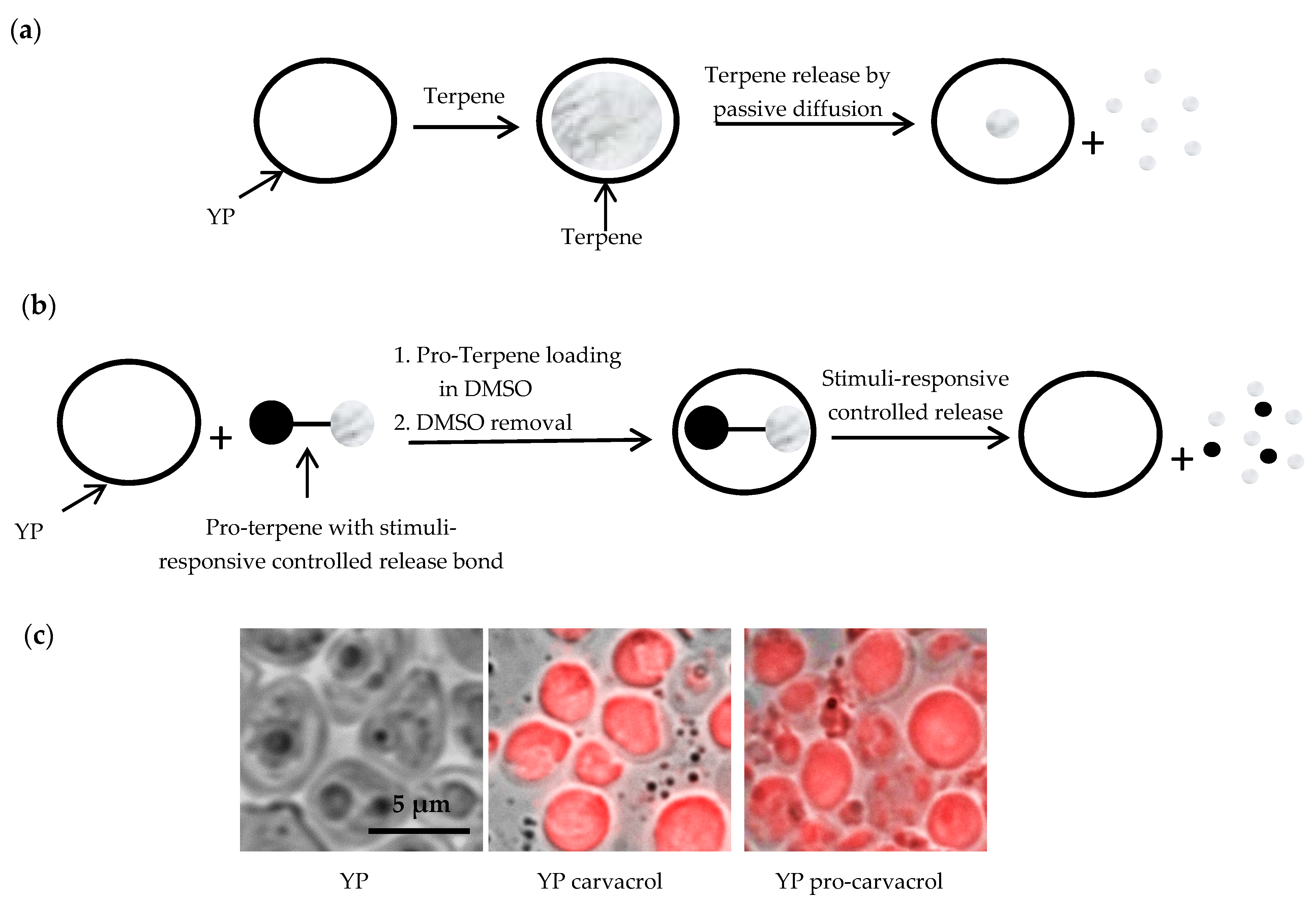
2.2.2. YP Loading of Terpenes (YP Terpene)
2.2.3. YP Loading of Pro-Terpenes (YP Pro-Terpene)
2.2.4. Characterization of Terpene and Pro-Terpene Loading Efficiency
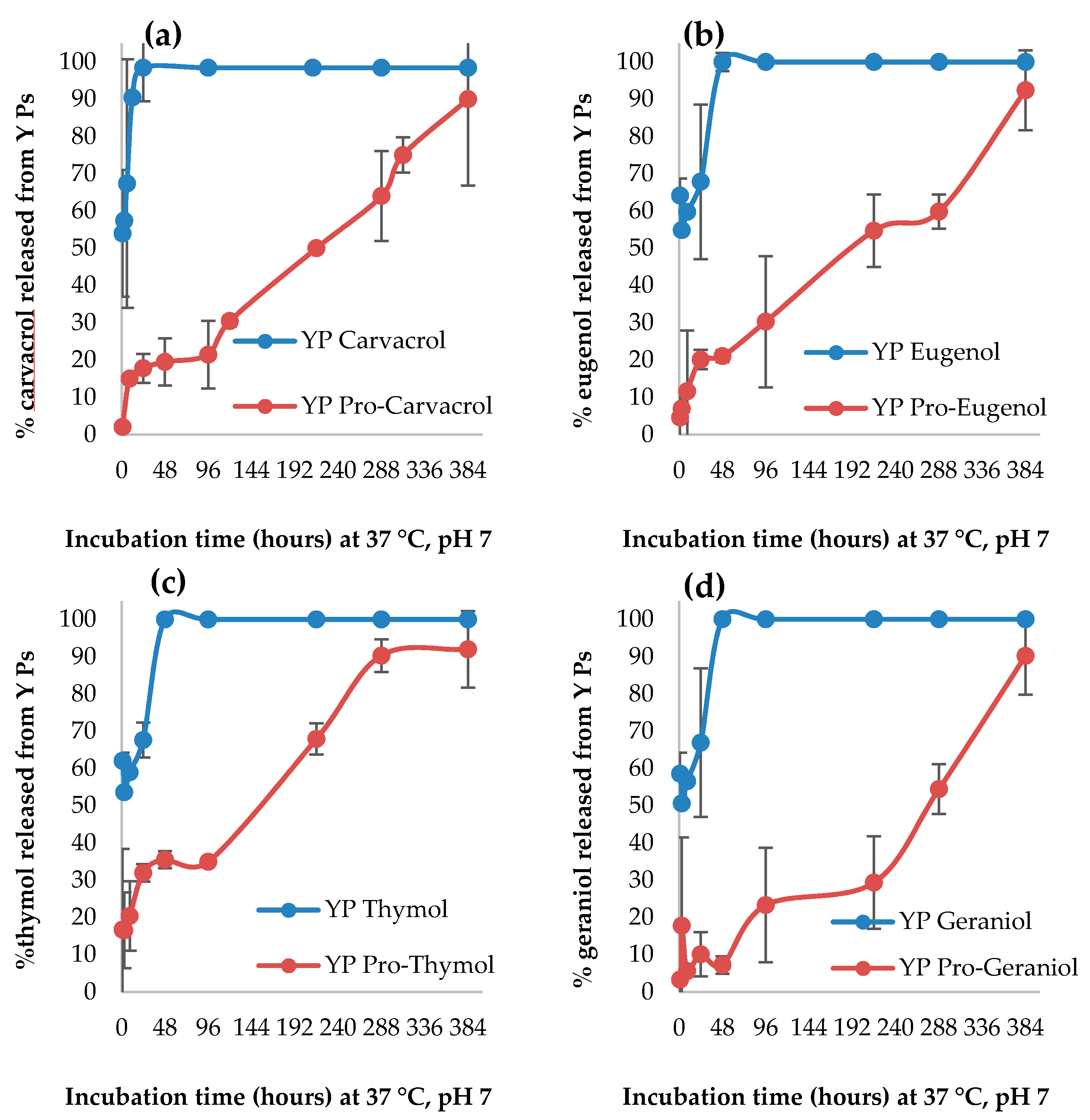
2.2.5. Terpene Release from YPs
2.2.6. Antimicrobial Activity Assays in Model Bacterial and Fungal Organisms
2.2.7. Antimicrobial Assay with Fractionated Samples
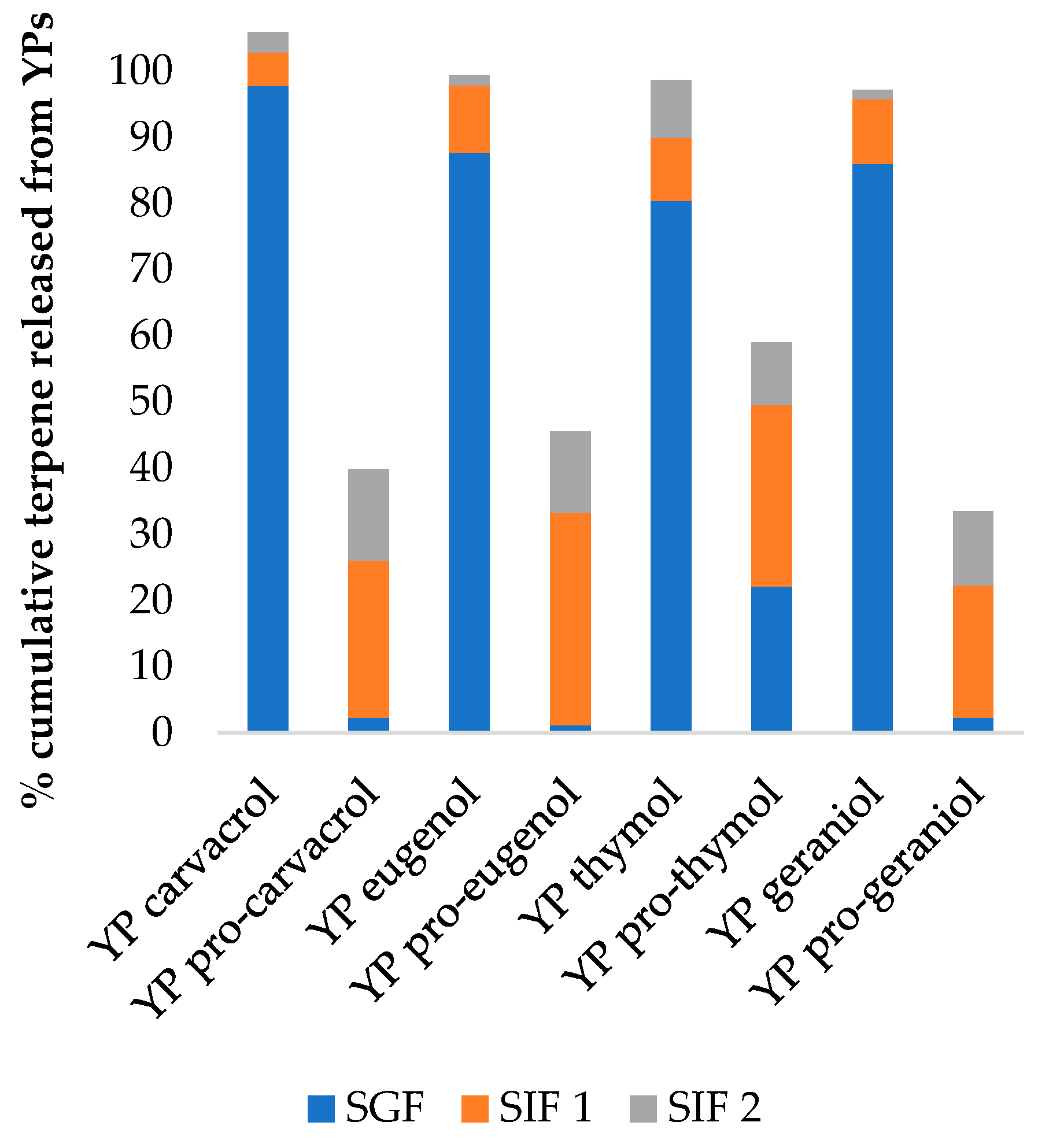
2.2.8. Simulated Digestion Assay
2.2.9. Nematode Extract Assay
2.2.10. Larval Development (Egg-to-Larvae (E2L)) Assay
2.2.11. Adult Worm In Vitro Screening
3. Results
3.1. Preparation and Characterization of Yeast Particle Encapsulated Terpenes and Pro-Terpenes
3.1.1. Synthesis of Pro-Terpenes
3.1.2. Yeast Particle Encapsulation of Terpenes and Pro-Terpenes
3.1.3. Terpene Release from YP Terpene and YP Pro-Terpene Samples
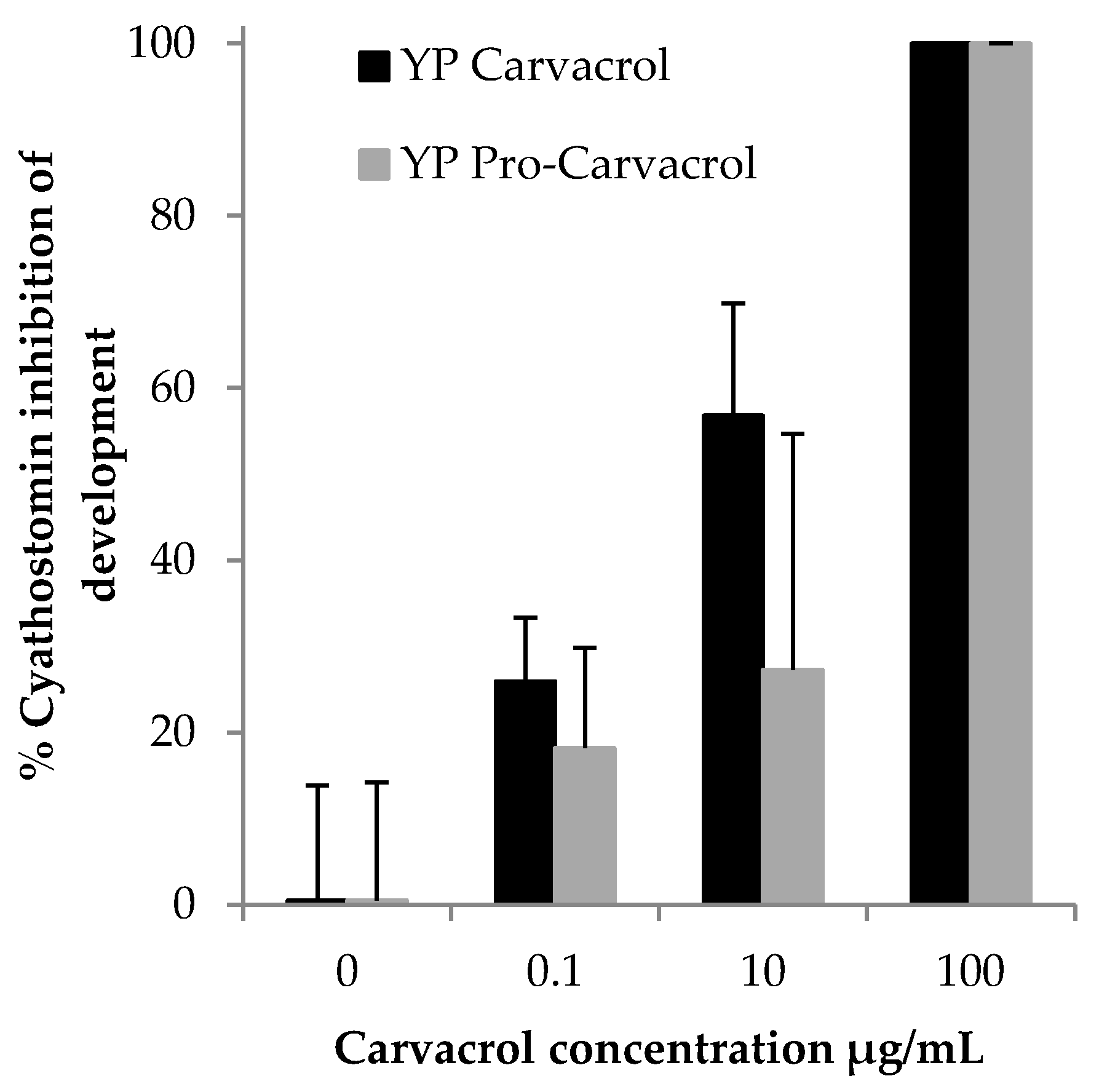
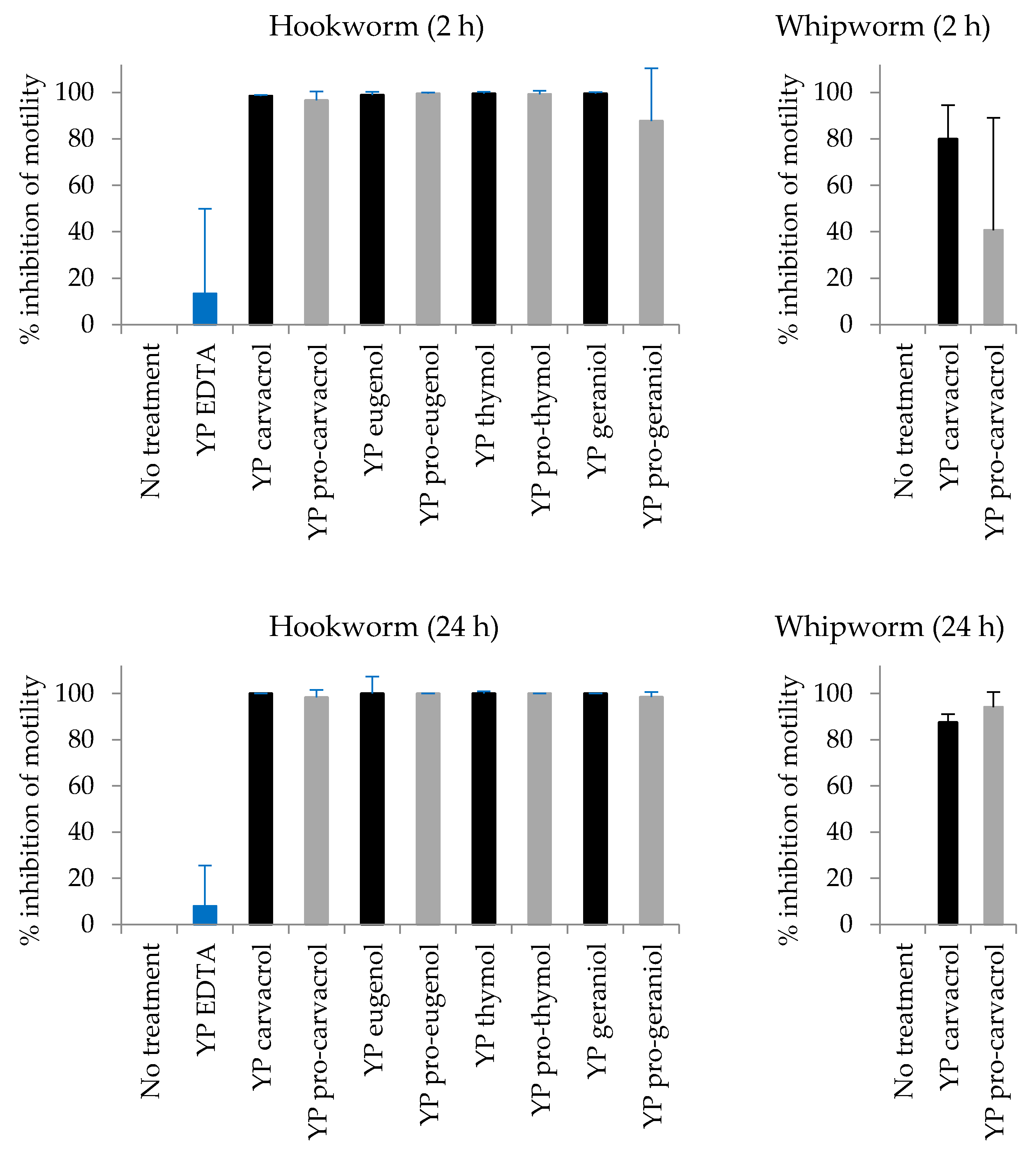
3.2. Biological Activity of YP Pro-Terpenes
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Falleh, H.; Ben Jemaa, M.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef]
- Ben Salha, G.; Abderrabba, M.; Labidi, J. A status review of terpenes and their separation methods. Rev. Chem. Eng. 2019, 37, 433–447. [Google Scholar] [CrossRef]
- Bakry, A.M.; Abbas, S.; Ali, B.; Majeed, H.; Abouelwafa, M.Y.; Mousa, A.; Liang, L. Microencapsulation of Oils: A Comprehensive Review of Benefits, Techniques, and Applications. Compr. Rev. Food Sci. Food Saf. 2016, 15, 143–182. [Google Scholar] [CrossRef]
- Calo, J.R.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems—A review. Food Control 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Saifullah, M.; Shishir, M.R.I.; Ferdowsi, R.; Tanver Rahman, M.R.; Van Vuong, Q. Micro and nano encapsulation, retention and controlled release of flavor and aroma compounds: A critical review. Trends Food Sci. Technol. 2019, 86, 230–251. [Google Scholar] [CrossRef]
- Bhalerao, Y.P.; Wagh, S.J. A Review on thymol encapsulation and its controlled release through biodegradable polymer shells. Int. J. Pharm. Sci. Res. 2018, 9, 4522–4532. [Google Scholar]
- De Matos, S.P.; Teixeira, H.F.; De Lima, A.A.N.; Veiga-Junior, V.F.; Koester, L.S. Essential oils and isolated terpenes in nanosystems designed for topical administration: A review. Biomolecules 2019, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.R.; Ostroff, G.R. Characterization of multilayered nanoparticles encapsulated in yeast cell wall particles for DNA. Bioconjug. Chem. 2008, 19, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Aouadi, M.; Tesz, G.J.; Nicoloro, S.M.; Wang, M.; Chouinard, M.; Soto, E.; Ostroff, G.R.; Czech, M.P. Orally delivered siRNA targeting macrophage Map4k4 suppresses systemic inflammation. Nature 2009, 458, 1180–1184. [Google Scholar] [CrossRef]
- Soto, E.; Ostroff, G. Glucan particles as carriers of nanoparticles for macrophage-targeted delivery. In Nanomaterials for Biomedicine; American Chemical Society: Washington, DC, USA, 2012; pp. 57–79. [Google Scholar] [CrossRef]
- Mirza, Z.; Soto, E.R.; Dikengil, F.; Levitz, S.M.; Ostroff, G.R. Beta-glucan particles as vaccine adjuvant carriers. In Vaccines for Invasive Fungal Infections; Humana Press: Totowa, NJ, USA, 2017; pp. 143–157. [Google Scholar]
- Soto, E.R.; O’Connell, O.; Dikengil, F.; Peters, P.J.; Clapham, P.R.; Ostroff, G.R. Targeted Delivery of Glucan Particle Encapsulated Gallium Nanoparticles Inhibits HIV Growth in Human Macrophages. J. Drug Deliv. 2016, 2016, 8520629. [Google Scholar] [CrossRef]
- Hamza, Z.; El-Hashash, M.; Aly, S.; Hathout, A.; Soto, E.; Sabry, B.; Ostroff, G. Preparation and characterization of yeast cell wall beta-glucan encapsulated humic acid nanoparticles as an enhanced aflatoxin B1 binder. Carbohydr. Polym. 2019, 203, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Specht, C.A.; Levitz, S.M. Robust Stimulation of Humoral and Cellular Immune Responses following Vaccination with Antigen-Loaded β-Glucan Particles. MBio. 2010, 1, e00164-10. [Google Scholar] [CrossRef]
- Franklin, L.; Ostroff, G.; Harman, G. Terpene-Containing Compositions and Methods of Making and Using Them. U.S. Patent No. 10,667,512, 2 June 2020. [Google Scholar]
- Franklin, L.; Ostroff, G. Compositions and Methods Comprising Terpenes or Terpene Mixtures Selected from Thymol, Eugenol, Geraniol, Citral and L-Carvone. U.S. Patent No. 10,258,033, 16 April 2019. [Google Scholar]
- Franklin, L.; Ostroff, G. Compositions Containing a Hollow Glucan Particle or a Cell Wall Particle Encapsulating a Terpene Component, Methods of Making and Using Them. U.S. Patent No. 10,638,750, 5 May 2020. [Google Scholar]
- Franklin, L.; Ostroff, G. Nematicidal Compositions and Methods of Using Them. U.S. Patent No. 10,004,229, 26 June 2018. [Google Scholar]
- Franklin, L.; Ostroff, G. Nematicidal Compositions and Methods of Using Them. U.S. Patent No. 9,655,360, 23 May 2017. [Google Scholar]
- Franklin, L.; Ostroff, G. Compositions and Methods Comprising Terpenes or Terpene Mixtures Selected from Thymol, Eugenol, Geraniol, Citral, and L-Carvone. U.S. Patent No. 9,439,416, 13 September 2016. [Google Scholar]
- Mirza, Z.; Soto, E.R.; Hu, Y.; Nguyen, T.T.; Koch, D.; Aroian, R.V.; Ostroff, G.R. Anthelmintic activity of yeast particle-encapsulated terpenes. Molecules 2020, 25, 2958. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.; Rus, F.; Ostroff, G.R. Yeast Cell Wall Particle Encapsulation of Pro-Terpene Payloads. Nanotech 2019 TechConnect Briefs. 2019. Available online: https://briefs.techconnect.org/papers/yeast-cell-wall-particle-encapsulation-of-pro-terpene-payloads/ (accessed on 10 January 2021).
- Carbone-Howell, A.L.; Stebbins, N.D.; Uhrich, K.E. Poly(anhydride-esters) comprised exclusively of naturally occurring antimicrobials and EDTA: Antioxidant and antibacterial activities. Biomacromolecules 2014, 15, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Sultanbawa, Y.; Cusack, A.; Currie, M.; Davis, C. An innovative microplate assay to facilitate the detection of antimicrobial activity in plant extracts. J. Rapid Methods Autom. Microbiol. 2009, 17, 519–534. [Google Scholar] [CrossRef]
- Harley, C.A.; Tipper, D.J. The role of charged residues in determining transmembrane protein insertion orientation in yeast. J. Biol. Chem. 1996, 271, 24625–24633. [Google Scholar] [CrossRef]
- U.S. Pharmacopeia. Simulated Gastric Fluid, TS. National Formulary No. 24/19; National Publishing: Philadelphia, PA, USA, 2000. [Google Scholar]
- U.S. Pharmacopeia. Simulated Intestinal Fluid; United States Pharmacopeial Convention: Rockville, MD, USA, 1995. [Google Scholar]
- Urban, J.F., Jr.; Hu, Y.; Miller, M.M.; Scheib, U.; Yiu, Y.Y.; Aroian, R.V. Bacillus thuringiensis-derived Cry5B has potent anthelmintic activity against Ascaris suum. PLoS Negl. Trop. Dis. 2013, 7, e2263. [Google Scholar] [CrossRef]
- Hu, Y.; Miller, M.; Zhang, B.; Nguyen, T.T.; Nielsen, M.K.; Aroian, R.V. In vivo and in vitro studies of Cry5B and nicotinic acetylcholine receptor agonist anthelmintics reveal a powerful and unique combination therapy against intestinal nematode parasites. PLoS Negl. Trop. Dis. 2018, 12, e0006506. [Google Scholar] [CrossRef]
- Lyons, E.T.; Drudge, J.H.; Tolliver, S.C. Prevalence of some internal parasites found (1971-1989) in horses born on a farm in Central Kentucky. J. Equine Vet. Sci. 1990, 10, 99–107. [Google Scholar] [CrossRef]
- Hu, Y.; Zhan, B.; Keegan, B.; Yiu, Y.Y.; Miller, M.M.; Jones, K.; Aroian, R.V. Mechanistic and Single-Dose In Vivo Therapeutic Studies of Cry5B Anthelmintic Action against Hookworms. PLoS Negl. Trop. Dis. 2012, 6, e1900. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xiao, S.H.; Aroian, R.V. The new anthelmintic tribendimidine is an L-type (Levamisole and Pyrantel) nicotinic acetylcholine receptor agonist. PLoS Negl. Trop. Dis. 2009, 3, e499. [Google Scholar] [CrossRef] [PubMed]
- Marcellino, C.; Gut, J.; Lim, K.C.; Singh, R.; McKerrow, J.; Sakanari, J. WormAssay: A novel computer application for whole-plate motion-based screening of macroscopic parasites. PLoS Negl. Trop. Dis. 2012, 6, e1494. [Google Scholar] [CrossRef] [PubMed]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 17 December 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Melting Point (°C) | Solubility in Water |
|---|---|---|
| Carvacrol 1 | 3.5 | 1.25 mg/mL |
| Pro-carvacrol 2 | 156–158 | Insoluble |
| Eugenol 1 | −9.2 | 1.44 mg/mL |
| Pro-eugenol 2 | 167–169 | <20 µg/mL |
| Thymol 1 | 51.5 | 0.90 mg/mL |
| Pro-thymol 2 | 135–138 | Insoluble |
| Geraniol 1 | −15 | 0.1 mg/mL |
| Pro-geraniol 2 | Decomposes above 145 | Insoluble |
| Sample | MIC 75% (Average of n = 4) | |
|---|---|---|
| Negative controls | Empty YP | Not active |
| YP + EDTA | 4244 ± 1400 µg EDTA/mL | |
| Carvacrol samples | Carvacrol | 625 ± 0 µg carvacrol/mL |
| YP carvacrol | 677 ± 45 µg carvacrol/mL | |
| YP pro-carvacrol | 625 ± 0 µg carvacrol/mL | |
| Eugenol samples | Eugenol | 938 ± 361 µg eugenol/mL |
| YP eugenol | 312 ± 0 µg eugenol/mL | |
| YP pro-eugenol | 625 ± 442 µg eugenol/mL | |
| Geraniol samples | Geraniol | 1094 ± 313 µg geraniol/mL |
| YP geraniol | 781 ± 312 µg geraniol/mL | |
| YP pro-geraniol | 1094 ± 312 µg geraniol/mL | |
| LB | Fraction | MIC 75% (µg Carvacrol/mL, Average of n = 3) |
|---|---|---|
| Fresh | Supernatant | Not active |
| Pellet | 521 ± 180 | |
| Spent | Supernatant | 500 ± 216 |
| Pellet | 729 ± 252 |
| Sample | MIC 75% (Average of n = 3) | |
|---|---|---|
| Negative controls | Empty YP | Not active |
| YP + EDTA | Not active | |
| Eugenol samples | Eugenol | 703 ± 773 g eugenol/mL |
| YP eugenol | 390 ± 193 µg eugenol/mL | |
| YP pro-eugenol | 781 ± 362 µg eugenol/mL | |
| Thymol samples | Thymol | 312 ± 110 µg thymol/mL |
| YP thymol | 1250 ± 442 µg thymol/mL | |
| YP pro-thymol | 781 ± 312 µg thymol/mL | |
| Geraniol samples | Geraniol | 703 ± 773 µg geraniol/mL |
| YP geraniol | 312 ± 221 µg geraniol/mL | |
| YP pro-geraniol | 260 ± 90 µg geraniol/mL | |
| Buffer ± Ascaris Extract | % Carvacrol Released from YPs after 24 h Incubation | MIC 75% (µg carvacrol/mL, Average of n = 3) | |
|---|---|---|---|
| Supernatant | Pellet | ||
| PBS (pH 7) | 4.5 ± 1.8 | Not active | 597 |
| Ascaris extract in PBS (pH 7) | 27.9 ± 6.0 | 291 ± 101 | 451 ± 0 |
| Acetate buffer (pH 5) | 6.5 ± 0.9 | 325 | 584 |
| Ascaris extract in acetate buffer (pH 5) | 37.3 ± 9.5 | 155 ± 67 | 261 ± 113 |
| Carbonate buffer (pH 10) | 95.6 ± 1.2 | 598 | Not active |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, E.R.; Rus, F.; Li, H.; Garceau, C.; Chicca, J.; Elfawal, M.; Gazzola, D.; Nielsen, M.K.; Urban, J.F., Jr.; Aroian, R.V.; et al. Yeast Particle Encapsulation of Scaffolded Terpene Compounds for Controlled Terpene Release. Foods 2021, 10, 1207. https://doi.org/10.3390/foods10061207
Soto ER, Rus F, Li H, Garceau C, Chicca J, Elfawal M, Gazzola D, Nielsen MK, Urban JF Jr., Aroian RV, et al. Yeast Particle Encapsulation of Scaffolded Terpene Compounds for Controlled Terpene Release. Foods. 2021; 10(6):1207. https://doi.org/10.3390/foods10061207
Chicago/Turabian StyleSoto, Ernesto R., Florentina Rus, Hanchen Li, Carli Garceau, Jeffrey Chicca, Mostafa Elfawal, David Gazzola, Martin K. Nielsen, Joseph F. Urban, Jr., Raffi V. Aroian, and et al. 2021. "Yeast Particle Encapsulation of Scaffolded Terpene Compounds for Controlled Terpene Release" Foods 10, no. 6: 1207. https://doi.org/10.3390/foods10061207
APA StyleSoto, E. R., Rus, F., Li, H., Garceau, C., Chicca, J., Elfawal, M., Gazzola, D., Nielsen, M. K., Urban, J. F., Jr., Aroian, R. V., & Ostroff, G. R. (2021). Yeast Particle Encapsulation of Scaffolded Terpene Compounds for Controlled Terpene Release. Foods, 10(6), 1207. https://doi.org/10.3390/foods10061207