
Oral Mucosa as a Potential Site for Diagnosis and Treatment of Allergic and Autoimmune Diseases

 ,
, 

Abstract
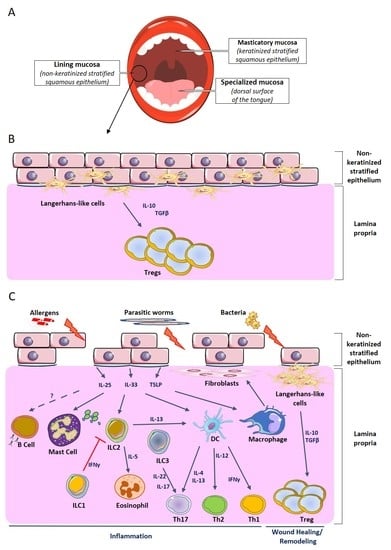
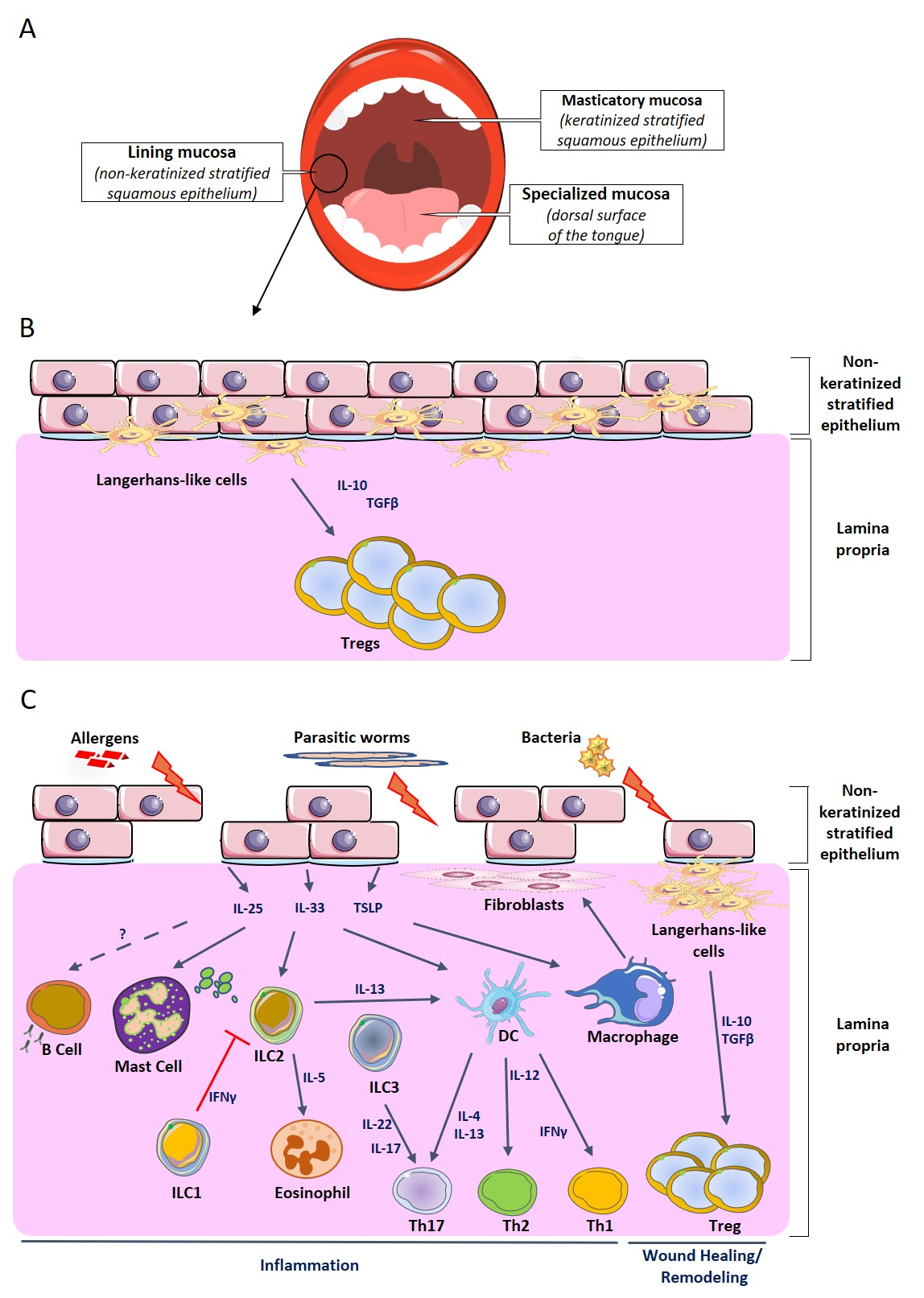{kind=link}
1. Introduction
2. Oral Mucosa
2.1. Structure of the Oral Mucosa
2.2. Oral Mucosa-Associated Immune System
3. Oral Mucosa in Food Allergy
4. Oral Mucosa in Other Allergies and Inflammatory Pathologies
4.1. Oral Mucosa in Respiratory Allergies with No Associated Food Allergy
4.2. Oral Contact Mucosal Allergies
4.3. Oral Mucosa in Celiac Disease (CD)
4.4. Oral Manifestations of Gastrointestinal Disease
4.5. Oral Manifestations in Autoimmunity
4.6. Oral Mucosa and Systemic Disease
5. Potential of Oral Mucosa Characterization in Diagnosis and Treatment
5.1. Research Approaches
5.2. Diagnosis
5.3. Treatment
5.3.1. Micronutrients
5.3.2. Oral Immunotherapy or Desensitization
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Moutsopoulos, N.M.; Konkel, J.E. Tissue-Specific Immunity at the Oral Mucosal Barrier. Trends Immunol. 2018, 39, 276–287. [Google Scholar] [CrossRef]
- Leoni, G.; Neumann, P.-A.; Sumagin, R.; Denning, T.L.; Nusrat, A. Wound repair: Role of immune–epithelial interactions. Mucosal Immunol. 2015, 8, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Novak, N.; Haberstok, J.; Bieber, T.; Allam, J.-P. The immune privilege of the oral mucosa. Trends Mol. Med. 2008, 14, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Moutsopoulos, N.M.; Moutsopoulos, H.M. The oral mucosa: A barrier site participating in tissue-specific and systemic immunity. Oral Dis. 2018, 24, 22–25. [Google Scholar] [CrossRef]
- Rosace, D.; Gomez-Casado, C.; Fernandez, P.; Perez-Gordo, M.; del Carmen Dominguez, M.; Vega, A.; Belver, M.T.; Ramos, T.; Vega, F.; Marco, G.; et al. Profilin-mediated food-induced allergic reactions are associated with oral epithelial remodeling. J. Allergy Clin. Immunol. 2019, 143, 681–690.e1. [Google Scholar] [CrossRef]
- Sanchez-Solares, J.; Delgado-Dolset, M.I.; Mera-Berriatua, L.; Hormias-Martin, G.; Cumplido, J.A.; Saiz, V.; Carrillo, T.; Moreno-Aguilar, C.; Escribese, M.M.; Gomez-Casado, C.; et al. Respiratory allergies with no associated food allergy disrupt oral mucosa integrity. Allergy 2019, 74, 2261–2265. [Google Scholar] [CrossRef]
- Sanchez-Solares, J.; Sanchez, L.; Pablo-Torres, C.; Diaz-Fernandez, C.; Sørensen, P.; Barber, D.; Gomez-Casado, C. Celiac Disease Causes Epithelial Disruption and Regulatory T Cell Recruitment in the Oral Mucosa. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Squier, C.; Brogden, K.A. Human Oral Mucosa: Development, Structure and Function; John Wiley & Sons, Ltd.: West Sussex, UK, 2013; ISBN 9781118710470. [Google Scholar]
- Shetty, S.; Gokul, S. Keratinization and its Disorders. Oman Med. J. 2012, 27, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Groeger, S.; Meyle, J. Oral Mucosal Epithelial Cells. Front. Immunol. 2019, 10, 208. [Google Scholar] [CrossRef]
- Samiei, M.; Ahmadian, E.; Eftekhari, A.; Eghbal, M.A.; Rezaie, F.; Vinken, M. Cell junctions and oral health. EXCLI J. 2019, 18, 317–330. [Google Scholar] [CrossRef]
- Wan, H.; Gadmor, H.; Brown, L. Anchoring junctions in the oral mucosa: Adherens junctions and desmosomes. In Oral Mucosa in Health and Disease; Springer: Cham, Switzerland, 2018; pp. 31–51. [Google Scholar]
- Liu, W.; Cui, Y.; Wei, J.; Sun, J.; Zheng, L.; Xie, J. Gap junction-mediated cell-to-cell communication in oral development and oral diseases: A concise review of research progress. Int. J. Oral Sci. 2020, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Ewert, P.; Aguilera, S.; Alliende, C.; Kwon, Y.-J.; Albornoz, A.; Molina, C.; Urzúa, U.; Quest, A.F.G.; Olea, N.; Pérez, P.; et al. Disruption of tight junction structure in salivary glands from Sjögren’s syndrome patients is linked to proinflammatory cytokine exposure. Arthritis Rheum. 2010, 62, 1280–1289. [Google Scholar] [CrossRef]
- Martin, T.A.; Mansel, R.E.; Jiang, W.G. Loss of occludin leads to the progression of human breast cancer. Int. J. Mol. Med. 2010, 26, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, P.P.; Bharadwaj, R.R.; Machado, J.; Macmillan, C.; Pintilie, M.; Sukhai, M.A.; Perez-Ordonez, B.; Gullane, P.; Irish, J.; Kamel-Reid, S. Claudin 1 overexpression increases invasion and is associated with aggressive histological features in oral squamous cell carcinoma. Cancer 2008, 113, 3169–3180. [Google Scholar] [CrossRef]
- Villaret, D.B.; Wang, T.; Dillon, D.; Xu, J.; Sivam, D.; Cheever, M.A.; Reed, S.G. Identification of genes overexpressed in head and neck squamous cell carcinoma using a combination of complementary DNA subtraction and microarray analysis. Laryngoscope 2000, 110, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Groeger, S.E.; Meyle, J. Epithelial barrier and oral bacterial infection. Periodontology 2000 2015, 69, 46–67. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, O.A.; Den Beste, K.; Hoddeson, E.K.; Parkos, C.A.; Nusrat, A.; Wise, S.K. House dust mite allergen Der p 1 effects on sinonasal epithelial tight junctions. Int. Forum Allergy Rhinol. 2013, 3, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Golebski, K.; Röschmann, K.I.L.; Toppila-Salmi, S.; Hammad, H.; Lambrecht, B.N.; Renkonen, R.; Fokkens, W.J.; van Drunen, C.M. The multi-faceted role of allergen exposure to the local airway mucosa. Allergy 2013, 68, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Steelant, B.; Farré, R.; Wawrzyniak, P.; Belmans, J.; Dekimpe, E.; Vanheel, H.; Van Gerven, L.; Kortekaas Krohn, I.; Bullens, D.M.A.; Ceuppens, J.L.; et al. Impaired barrier function in patients with house dust mite-induced allergic rhinitis is accompained by decreased occludin and zonula occludens-1 expression. J. Allergy Clin. Immunol. 2016, 137, 1043–1053.e5. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. Allergens and the airway epithelium response: Gateway to allergic sensitization. J. Allergy Clin. Immunol. 2014, 134, 499–507. [Google Scholar] [CrossRef]
- Edelblum, K.L.; Turner, J.R. The tight junction in inflammatory disease: Communication breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Van Deventer, S.J.H. Anti-TNF antibody treatment of Crohn’s disease. Ann. Rheum. Dis. 1999, 58 (Suppl. 1), i114–i120. [Google Scholar] [CrossRef]
- Abe, A.; Takano, K.; Kojima, T.; Nomura, K.; Kakuki, T.; Kaneko, Y.; Yamamoto, M.; Takahashi, H.; Himi, T. Interferon-gamma increased epithelial barrier function via upregulating claudin-7 expression in human submandibular gland duct epithelium. J. Mol. Histol. 2016, 47, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.; Xiang, R.-L.; Cong, X.; Zhang, Y.; Li, J.; Yi, X.; Park, K.; Han, J.-Y.; Wu, L.-L.; Yu, G.-Y. Claudin-3 is required for modulation of paracellular permeability by TNF-α through ERK1/2/slug signaling axis in submandibular gland. Cell. Signal. 2015, 27, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Baker, O.J.; Camden, J.M.; Redman, R.S.; Jones, J.E.; Seye, C.I.; Erb, L.; Weisman, G.A. Proinflammatory cytokines tumor necrosis factor-alpha and interferon-gamma alter tight junction structure and function in the rat parotid gland Par-C10 cell line. Am. J. Physiol. Cell Physiol. 2008, 295, C1191–C1201. [Google Scholar] [CrossRef] [PubMed]
- Roan, F.; Obata-Ninomiya, K.; Ziegler, S.F. Epithelial cell–derived cytokines: More than just signaling the alarm. J. Clin. Investig. 2019, 129, 1441–1451. [Google Scholar] [CrossRef]
- Moharamzadeh, K.; Brook, I.M.; Van Noort, R.; Scutt, A.M.; Thornhill, M.H. Tissue-engineered Oral Mucosa: A Review of the Scientific Literature. J. Dent. Res. 2007, 86, 115–124. [Google Scholar] [CrossRef]
- Berin, M.C.; Sampson, H.A. Mucosal Immunology of Food Allergy. Curr. Biol. 2013, 23, R389–R400. [Google Scholar] [CrossRef]
- Hovav, A.-H. Dendritic cells of the oral mucosa. Mucosal Immunol. 2014, 7, 27–37. [Google Scholar] [CrossRef]
- Henri, S.; Poulin, L.F.; Tamoutounour, S.; Ardouin, L.; Guilliams, M.; de Bovis, B.; Devilard, E.; Viret, C.; Azukizawa, H.; Kissenpfennig, A.; et al. CD207+ CD103+ dermal dendritic cells cross-present keratinocyte-derived antigens irrespective of the presence of Langerhans cells. J. Exp. Med. 2010, 207, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Mascarell, L.; Lombardi, V.; Louise, A.; Saint-Lu, N.; Chabre, H.; Moussu, H.; Betbeder, D.; Balazuc, A.-M.; Van Overtvelt, L.; Moingeon, P. Oral dendritic cells mediate antigen-specific tolerance by stimulating TH1 and regulatory CD4+ T cells. J. Allergy Clin. Immunol. 2008, 122, 603–609.e5. [Google Scholar] [CrossRef] [PubMed]
- Allam, J.-P.; Stojanovski, G.; Friedrichs, N.; Peng, W.; Bieber, T.; Wenzel, J.; Novak, N. Distribution of Langerhans cells and mast cells within the human oral mucosa: New application sites of allergens in sublingual immunotherapy? Allergy 2008, 63, 720–727. [Google Scholar] [CrossRef]
- Santoro, A.; Majorana, A.; Roversi, L.; Gentili, F.; Marrelli, S.; Vermi, W.; Bardellini, E.; Sapelli, P.; Facchetti, F. Recruitment of dendritic cells in oral lichen planus. J. Pathol. 2005, 205, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Jotwani, R.; Cutler, C.W. Multiple Dendritic Cell (DC) Subpopulations in Human Gingiva and Association of Mature DCs with CD4+ T-cells in situ. J. Dent. Res. 2003, 82, 736–741. [Google Scholar] [CrossRef]
- Dutzan, N.; Konkel, J.E.; Greenwell-Wild, T.; Moutsopoulos, N.M. Characterization of the human immune cell network at the gingival barrier. Mucosal Immunol. 2016, 9, 1163–1172. [Google Scholar] [CrossRef]
- Panda, S.K.; Colonna, M. Innate Lymphoid Cells in Mucosal Immunity. Front. Immunol. 2019, 10, 861. [Google Scholar] [CrossRef]
- Dutzan, N.; Abusleme, L.; Bridgeman, H.; Greenwell-Wild, T.; Zangerle-Murray, T.; Fife, M.E.; Bouladoux, N.; Linley, H.; Brenchley, L.; Wemyss, K.; et al. On-going Mechanical Damage from Mastication Drives Homeostatic Th17 Cell Responses at the Oral Barrier. Immunity 2017, 46, 133–147. [Google Scholar] [CrossRef]
- Park, J.Y.; Chung, H.; Choi, Y.; Park, J.H. Phenotype and Tissue Residency of Lymphocytes in the Murine Oral Mucosa. Front. Immunol. 2017, 8, 250. [Google Scholar] [CrossRef]
- Glowacki, A.J.; Yoshizawa, S.; Jhunjhunwala, S.; Vieira, A.E.; Garlet, G.P.; Sfeir, C.; Little, S.R. Prevention of inflammation-mediated bone loss in murine and canine periodontal disease via recruitment of regulatory lymphocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 18525–18530. [Google Scholar] [CrossRef]
- Araujo-Pires, A.C.; Vieira, A.E.; Francisconi, C.F.; Biguetti, C.C.; Glowacki, A.; Yoshizawa, S.; Campanelli, A.P.; Trombone, A.P.F.; Sfeir, C.S.; Little, S.R.; et al. IL-4/CCL22/CCR4 Axis Controls Regulatory T-Cell Migration That Suppresses Inflammatory Bone Loss in Murine Experimental Periodontitis. J. Bone Miner. Res. 2015, 30, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nagashima, H.; Bando, K.; Lu, L.; Ozaki, A.; Morita, Y.; Fukumoto, S.; Ishii, N.; Sugawara, S. Oral CD103−CD11b+ classical dendritic cells present sublingual antigen and induce Foxp3+ regulatory T cells in draining lymph nodes. Mucosal Immunol. 2017, 10, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Koppelman, S.; Smits, M.; Tomassen, M.; de Jong, G.; Baumert, J.; Taylor, S.; Witkamp, R.; Veldman, R.; Pieters, R.; Wichers, H. Release of Major Peanut Allergens from Their Matrix under Various pH and Simulated Saliva Conditions—Ara h2 and Ara h6 Are Readily Bio-Accessible. Nutrients 2018, 10, 1281. [Google Scholar] [CrossRef] [PubMed]
- Sugita, K.; Steer, C.A.; Martinez-Gonzalez, I.; Altunbulakli, C.; Morita, H.; Castro-Giner, F.; Kubo, T.; Wawrzyniak, P.; Rückert, B.; Sudo, K.; et al. Type 2 innate lymphoid cells disrupt bronchial epithelial barrier integrity by targeting tight junctions through IL-13 in asthmatic patients. J. Allergy Clin. Immunol. 2018, 141, 300–310.e11. [Google Scholar] [CrossRef]
- Boyce, J.A.; Assa’ad, A.; Burks, A.W.; Jones, S.M.; Sampson, H.A.; Wood, R.A.; Plaut, M.; Cooper, S.F.; Fenton, M.J.; Arshad, S.H.; et al. Guidelines for the diagnosis and management of food allergy in the United States: Report of the NIAID-sponsored expert panel. J. Allergy Clin. Immunol. 2010, 126, S1–S58. [Google Scholar] [CrossRef] [PubMed]
- Celebi Sözener, Z.; Cevhertas, L.; Nadeau, K.; Akdis, M.; Akdis, C.A. Environmental factors in epithelial barrier dysfunction. J. Allergy Clin. Immunol. 2020, 145, 1517–1528. [Google Scholar] [CrossRef]
- Moran, T.P. The External Exposome and Food Allergy. Curr. Allergy Asthma Rep. 2020, 20, 37. [Google Scholar] [CrossRef]
- Agache, I.; Miller, R.; Gern, J.E.; Hellings, P.W.; Jutel, M.; Muraro, A.; Phipatanakul, W.; Quirce, S.; Peden, D. Emerging concepts and challenges in implementing the exposome paradigm in allergic diseases and asthma: A Practall document. Allergy 2019, 74, 449–463. [Google Scholar] [CrossRef]
- Gómez-Casado, C.; Díaz-Perales, A. Allergen-Associated Immunomodulators: Modifying Allergy Outcome. Arch. Immunol. Ther. Exp. 2016, 64, 339–347. [Google Scholar] [CrossRef]
- Wesemann, D.R.; Nagler, C.R. The Microbiome, Timing, and Barrier Function in the Context of Allergic Disease. Immunity 2016, 44, 728–738. [Google Scholar] [CrossRef]
- Akdis, C.A. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat. Rev. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, K.M.; Konstantinou, G.N.; Pilapil, M.; Arrieta, M.-C.; Noone, S.; Sampson, H.A.; Meddings, J.; Nowak-Węgrzyn, A. Intestinal permeability in children with food allergy on specific elimination diets. Pediatr. Allergy Immunol. 2013, 24, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Barau, E.; Molkhou, P.; Raynaud, F.; Barbet, J.P.; Dehennin, L. Food-Induced Alterations of Intestinal Permeability in Children with Cow’s Milk-Sensitive Enteropathy and Atopic Dermatitis. J. Pediatr. Gastroenterol. Nutr. 1989, 8, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.; Andre, F.; Colin, L.; Cavagna, S. Measurement of intestinal permeability to mannitol and lactulose as a means of diagnosing food allergy and evaluating therapeutic effectiveness of disodium cromoglycate. Ann. Allergy 1987, 59, 127–130. [Google Scholar] [PubMed]
- Brough, H.A.; Nadeau, K.C.; Sindher, S.B.; Alkotob, S.S.; Chan, S.; Bahnson, H.T.; Leung, D.Y.M.; Lack, G. Epicutaneous sensitization in the development of food allergy: What is the evidence and how can this be prevented? Allergy 2020, 75, 2185–2205. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.T.; Sasieni, P.; du Toit, G.; Syed, H.; Lack, G. Household peanut consumption as a risk factor for the development of peanut allergy. J. Allergy Clin. Immunol. 2009, 123, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Brough, H.A.; Santos, A.F.; Makinson, K.; Penagos, M.; Stephens, A.C.; Douiri, A.; Fox, A.T.; Du Toit, G.; Turcanu, V.; Lack, G. Peanut protein in household dust is related to household peanut consumption and is biologically active. J. Allergy Clin. Immunol. 2013, 132, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ramírez, G.; Pazos-Castro, D.; Gómez Torrijos, E.; Yuste Montalvo, A.; Romero-Sahagun, A.; González-Klein, Z.; Jimeno-Nogales, L.; Escribese, M.M.; Extremera Ortega, A.; Nuñez-Borque, E.; et al. Group 1 allergens, transported by mold spores, induce asthma exacerbation in a mouse model. Allergy 2020, 75, 2388–2391. [Google Scholar] [CrossRef]
- Tsakok, T.; Woolf, R.; Smith, C.H.; Weidinger, S.; Flohr, C. Atopic dermatitis: The skin barrier and beyond. Br. J. Dermatol. 2019, 180, 464–474. [Google Scholar] [CrossRef]
- Chalmers, J.R.; Haines, R.H.; Bradshaw, L.E.; Montgomery, A.A.; Thomas, K.S.; Brown, S.J.; Ridd, M.J.; Lawton, S.; Simpson, E.L.; Cork, M.J.; et al. Daily emollient during infancy for prevention of eczema: The BEEP randomised controlled trial. Lancet 2020, 395, 962–972. [Google Scholar] [CrossRef]
- Skjerven, H.O.; Rehbinder, E.M.; Vettukattil, R.; LeBlanc, M.; Granum, B.; Haugen, G.; Hedlin, G.; Landrø, L.; Marsland, B.J.; Rudi, K.; et al. Skin emollient and early complementary feeding to prevent infant atopic dermatitis (PreventADALL): A factorial, multicentre, cluster-randomised trial. Lancet 2020, 395, 951–961. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Burton, O.T.; Oettgen, H.C. Beyond immediate hypersensitivity: Evolving roles for IgE antibodies in immune homeostasis and allergic diseases. Immunol. Rev. 2011, 242, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J.; Sutton, B.J. IgE in allergy and asthma today. Nat. Rev. Immunol. 2008, 8, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Selb, R.; Eckl-Dorna, J.; Neunkirchner, A.; Schmetterer, K.; Marth, K.; Gamper, J.; Jahn-Schmid, B.; Pickl, W.F.; Valenta, R.; Niederberger, V. CD23 surface density on B cells is associated with IgE levels and determines IgE-facilitated allergen uptake, as well as activation of allergen-specific T cells. J. Allergy Clin. Immunol. 2017, 139, 290–299.e4. [Google Scholar] [CrossRef]
- Oettgen, H.C.; Burton, O.T. IgE receptor signaling in food allergy pathogenesis. Curr. Opin. Immunol. 2015, 36, 109–114. [Google Scholar] [CrossRef]
- Palaniyandi, S.; Tomei, E.; Li, Z.; Conrad, D.H.; Zhu, X. CD23-Dependent Transcytosis of IgE and Immune Complex across the Polarized Human Respiratory Epithelial Cells. J. Immunol. 2011, 186, 3484–3496. [Google Scholar] [CrossRef]
- Yu, L.C.H.; Yang, P.-C.; Berin, M.C.; Di Leo, V.; Conrad, D.H.; Mckay, D.M.; Satoskar, A.R.; Perdue, M.H. Enhanced transepithelial antigen transport in intestine of allergic mice is mediated by IgE/CD23 and regulated by interleukin-4. Gastroenterology 2001, 121, 370–381. [Google Scholar] [CrossRef]
- Barber, D.; Díaz-Perales, A.; Villalba, M.; Chivato, T. Challenges for Allergy Diagnosis in Regions with Complex Pollen Exposures. Curr. Allergy Asthma Rep. 2015, 15, 496. [Google Scholar] [CrossRef]
- Barber, D.; de la Torre, F.; Lombardero, M.; Antépara, I.; Colas, C.; Dávila, I.; Tabar, A.I.; Vidal, C.; Villalba, M.; Salcedo, G.; et al. Component-resolved diagnosis of pollen allergy based on skin testing with profilin, polcalcin and lipid transfer protein pan-allergens. Clin. Exp. Allergy 2009, 39, 1764–1773. [Google Scholar] [CrossRef]
- Ruiz-García, M.; García del Potro, M.; Fernández-Nieto, M.; Barber, D.; Jimeno-Nogales, L.; Sastre, J. Profilin: A relevant aeroallergen? J. Allergy Clin. Immunol. 2011, 128, 416–418. [Google Scholar] [CrossRef]
- Asero, R.; Tripodi, S.; Dondi, A.; Di Rienzo Businco, A.; Sfika, I.; Bianchi, A.; Candelotti, P.; Caffarelli, C.; Povesi Dascola, C.; Ricci, G.; et al. Prevalence and Clinical Relevance of IgE Sensitization to Profilin in Childhood: A Multicenter Study. Int. Arch. Allergy Immunol. 2015, 168, 25–31. [Google Scholar] [CrossRef]
- Mastrorilli, C.; Tripodi, S.; Caffarelli, C.; Perna, S.; Di Rienzo-Businco, A.; Sfika, I.; Asero, R.; Dondi, A.; Bianchi, A.; Povesi Dascola, C.; et al. Endotypes of pollen-food syndrome in children with seasonal allergic rhinoconjunctivitis: A molecular classification. Allergy 2016, 71, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Van Ree, R. Profilins: Mimickers of Allergy or Relevant Allergens. Int. Arch. Allergy Immunol. 2011, 155, 191–204. [Google Scholar] [CrossRef]
- Tsuchiya, T.; Nishimura, Y.; Nishiuma, T.; Kotani, Y.; Funada, Y.; Yoshimura, S.; Yokoyama, M. Airway remodeling of murine chronic antigen exposure model. J. Asthma 2003, 40, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Van Bruaene, N.; Derycke, L.; Perez-Novo, C.A.; Gevaert, P.; Holtappels, G.; De Ruyck, N.; Cuvelier, C.; Van Cauwenberge, P.; Bachert, C. TGF-beta signaling and collagen deposition in chronic rhinosinusitis. J. Allergy Clin. Immunol. 2009, 124, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Takabayashi, T.; Kato, A.; Peters, A.T.; Hulse, K.E.; Suh, L.A.; Carter, R.; Norton, J.; Grammer, L.C.; Cho, S.H.; Tan, B.K.; et al. Excessive fibrin deposition in nasal polyps caused by fibrinolytic impairment through reduction of tissue plasminogen activator expression. Am. J. Respir. Crit. Care Med. 2013, 187, 49–57. [Google Scholar] [CrossRef]
- Elia, C.S.A.; Renata, M.E.; Benito, A.S.M.; Vitorino, M.S.; Maria, G.R. Oral epithelial changes in HIV-positive individuals. Pathol. Res. Pract. 2013, 209, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Dodani, K.; Anumala, N.; Avula, H.; Reddy, K.; Varre, S.; Kalakonda, B.B.; Arora, N.; Suri, C.; Avula, J.K. Periodontal findings in patients with oral submucous fibrosis and comet assay of affected gingival epithelial cells. J. Periodontol. 2012, 83, 1038–1047. [Google Scholar] [CrossRef]
- Porsbjerg, C.; Baines, K.; Gibson, P.; Bergqvist, A.; Erjefält, J.S.; Sverrild, A.; Backer, V. IL-33 is related to innate immune activation and sensitization to HDM in mild steroid-free asthma. Clin. Exp. Allergy 2016, 46, 564–574. [Google Scholar] [CrossRef]
- Rak, G.D.; Osborne, L.C.; Siracusa, M.C.; Kim, B.S.; Wang, K.; Bayat, A.; Artis, D.; Volk, S.W. IL-33-Dependent Group 2 Innate Lymphoid Cells Promote Cutaneous Wound Healing. J. Investig. Dermatol. 2016, 136, 487–496. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, I.; Steer, C.A.; Takei, F. Lung ILC2s link innate and adaptive responses in allergic inflammation. Trends Immunol. 2015, 36, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, M.I.; Jimeno, L.; De La Torre, F.; Boissy, P.; Rivas, B.; Lázaro, M.J.; Barber, D. Profilin as a severe food allergen in allergic patients overexposed to grass pollen. Allergy 2014, 69, 1610–1616. [Google Scholar] [CrossRef]
- Peeters, K.A.B.M.; Koppelman, S.J.; van Hoffen, E.; van der Tas, C.W.H.; den Hartog Jager, C.F.; Penninks, A.H.; Hefle, S.L.; Bruijnzeel-Koomen, C.A.F.M.; Knol, E.F.; Knulst, A.C. Does skin prick test reactivity to purified allergens correlate with clinical severity of peanut allergy? Clin. Exp. Allergy 2007, 37, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, H.S.; Murray, K.S.; Schlichting, D.G.; Chen, X.; Hansen, K.C.; Duncan, M.W.; Dreskin, S.C. Effector activity of peanut allergens: A critical role for Ara h 2, Ara h 6, and their variants. Clin. Exp. Allergy 2009, 39, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Kulis, M.; Chen, X.; Lew, J.; Wang, Q.; Patel, O.P.; Zhuang, Y.; Murray, K.S.; Duncan, M.W.; Porterfield, H.S.; Burks, A.W.; et al. The 2S albumin allergens of Arachis hypogaea, Ara h 2 and Ara h 6, are the major elicitors of anaphylaxis and can effectively desensitize peanut-allergic mice. Clin. Exp. Allergy 2012, 42, 326–336. [Google Scholar] [CrossRef]
- Wüthrich, B. Lethal or life-threatening allergic reactions to food. J. Investig. Allergol. Clin. Immunol. 2000, 10, 59–65. [Google Scholar]
- Bock, S.A.; Muñoz-Furlong, A.; Sampson, H.A. Fatalities due to anaphylactic reactions to foods. J. Allergy Clin. Immunol. 2001, 107, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Mendelson, L.; Rosen, J.P. Fatal and Near-Fatal Anaphylactic Reactions to Food in Children and Adolescents. N. Engl. J. Med. 1992, 327, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Popescu, F.-D. Cross-reactivity between aeroallergens and food allergens. World J. Methodol. 2015, 5, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, R.R.; Davis, M.D.P.; Bruce, A.J.; Farmer, S.A.; Rogers, R.S. Contact allergy in oral disease. J. Am. Acad. Dermatol. 2007, 57, 315–321. [Google Scholar] [CrossRef] [PubMed]
- LeSueur, B.W.; Yiannias, J.A. Contact stomatitis. Dermatol. Clin. 2003, 21, 105–114. [Google Scholar] [CrossRef]
- Collet, E.; Jeudy, G.; Dalac, S. Cheilitis, perioral dermatitis and contact allergy. Eur. J. Dermatol. 2013, 23, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Assimakopoulos, D.; Patrikakos, G.; Fotika, C.; Elisaf, M. Benign migratory glossitis or geographic tongue: An enigmatic oral lesion. Am. J. Med. 2002, 113, 751–755. [Google Scholar] [CrossRef]
- Goregen, M.; Melikoglu, M.; Miloglu, O.; Erdem, T. Predisposition of allergy in patients with benign migratory glossitis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 110, 470–474. [Google Scholar] [CrossRef]
- Bilhan, H.; Bural, C.; Geckili, O. Titanium hypersensitivity. A hidden threat for dental implant patients? N. Y. State Dent. J. 2013, 79, 38–43. [Google Scholar]
- Wiesenthal, A.; Hunter, L.; Wang, S.; Wickliffe, J.; Wilkerson, M. Nanoparticles: Small and mighty. Int. J. Dermatol. 2011, 50, 247–254. [Google Scholar] [CrossRef]
- Teubl, B.J.; Leitinger, G.; Schneider, M.; Lehr, C.-M.; Fröhlich, E.; Zimmer, A.; Roblegg, E. The buccal mucosa as a route for TiO2 nanoparticle uptake. Nanotoxicology 2015, 9, 253–261. [Google Scholar] [CrossRef]
- Roblegg, E.; Fröhlich, E.; Meindl, C.; Teubl, B.; Zaversky, M.; Zimmer, A. Evaluation of a physiological in vitro system to study the transport of nanoparticles through the buccal mucosa. Nanotoxicology 2012, 6, 399–413. [Google Scholar] [CrossRef]
- Teubl, B.J.; Meindl, C.; Eitzlmayr, A.; Zimmer, A.; Fröhlich, E.; Roblegg, E. In-vitro permeability of neutral polystyrene particles via buccal mucosa. Small 2013, 9, 457–466. [Google Scholar] [CrossRef]
- Makihira, S.; Mine, Y.; Nikawa, H.; Shuto, T.; Iwata, S.; Hosokawa, R.; Kamoi, K.; Okazaki, S.; Yamaguchi, Y. Titanium ion induces necrosis and sensitivity to lipopolysaccharide in gingival epithelial-like cells. Toxicology 2010, 24, 1905–1910. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Sun, D.H.; Trindade, M.C.; Maloney, W.J.; Goodman, S.B.; Schurman, D.J.; Smith, R.L. Signaling pathways for tumor necrosis factor-alpha and interleukin-6 expression in human macrophages exposed to titanium-alloy particulate debris in vitro. J. Bone Joint Surg. Am. 1999, 81, 603–615. [Google Scholar] [CrossRef]
- Müller, K.; Valentine-Thon, E. Hypersensitivity to titanium: Clinical and laboratory evidence. Neuro Endocrinol. Lett. 2006, 27 (Suppl. 1), 31–35. [Google Scholar]
- Jimson, S.; Rajesh, E.; Krupaa, R.J.; Kasthuri, M. Burning mouth syndrome. J. Pharm. Bioallied Sci. 2015, 7, S194–S196. [Google Scholar] [CrossRef] [PubMed]
- Coculescu, E.C.; Tovaru, S.; Coculescu, B.I. Epidemiological and etiological aspects of burning mouth syndrome. J. Med. Life 2014, 7, 305–309. [Google Scholar] [PubMed]
- Steele, J.C.; Bruce, A.J.; Davis, M.D.P.; Torgerson, R.R.; Drage, L.A.; Rogers, R.S. Clinically relevant patch test results in patients with burning mouth syndrome. Dermat. Contact Atopic Occup. Drug 2012, 23, 61–70. [Google Scholar] [CrossRef]
- Manavalan, J.S.; Hernandez, L.; Shah, J.G.; Konikkara, J.; Naiyer, A.J.; Lee, A.R.; Ciaccio, E.; Minaya, M.T.; Green, P.H.R.; Bhagat, G. Serum cytokine elevations in celiac disease: Association with disease presentation. Hum. Immunol. 2010, 71, 50–57. [Google Scholar] [CrossRef]
- Leffler, D.A.; Green, P.H.R.; Fasano, A. Extraintestinal manifestations of coeliac disease. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 561–571. [Google Scholar] [CrossRef]
- Nardecchia, S.; Auricchio, R.; Discepolo, V.; Troncone, R. Extra-Intestinal Manifestations of Coeliac Disease in Children: Clinical Features and Mechanisms. Front. Pediatr. 2019, 7. [Google Scholar] [CrossRef]
- Hernandez, L.; Green, P.H. Extraintestinal manifestations of celiac disease. Curr. Gastroenterol. Rep. 2006, 8, 383–389. [Google Scholar] [CrossRef]
- López-Casado, M.; Lorite, P.; Palomeque, T.; Torres, M. Potential role of the IL-33/ST2 axis in celiac disease. Cell. Mol. Immunol. 2017, 14, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Heydari, F.; Rostami-Nejad, M.; Moheb-Alian, A.; Mollahoseini, M.H.; Rostami, K.; Pourhoseingholi, M.A.; Aghamohammadi, E.; Zali, M.R. Serum Cytokines Profile in Treated Celiac Disease Compared with Non-celiac Gluten Sensitivity and Control: A Marker for Differentiation. J. Gastrointest. Liver Dis. 2018, 27, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Björck, S.; Lindehammer, S.R.; Fex, M.; Agardh, D. Serum cytokine pattern in young children with screening detected coeliac disease. Clin. Exp. Immunol. 2015, 179, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Tye-Din, J.A.; Qiao, S.-W.; Russell, A.K.; Mayassi, T.; Ciszewski, C.; Sarna, V.K.; Wang, S.; Goldstein, K.E.; Dzuris, J.L.; et al. Cytokine release and gastrointestinal symptoms after gluten challenge in celiac disease. Sci. Adv. 2019, 5, eaaw7756. [Google Scholar] [CrossRef] [PubMed]
- Romero-Adrián, T.B. Celiac disease: Participation of Cytokines and Other Factors in the Immune Response. J. Gastrointest. Disord. Liver Funct. 2016, 1, 1–6. [Google Scholar] [CrossRef][Green Version]
- Zaiss, D.M.; Minutti, C.M.; Knipper, J.A. Immune- and non-immune-mediated roles of regulatory T-cells during wound healing. Immunology 2019, 157, 190–197. [Google Scholar] [CrossRef]
- Varona, R.; Ramos, T.; Escribese, M.M.; Jimeno, L.; Galán, A.; Würtzen, P.A.; Vega, F.; Marín, A.; Martín, S.; Carrera, A.C.; et al. Persistent regulatory T-cell response 2 years after 3 years of grass tablet SLIT: Links to reduced eosinophil counts, sIgE levels, and clinical benefit. Allergy 2019, 74, 349–360. [Google Scholar] [CrossRef]
- Mejia, L.M. Oral Manifestations of Gastrointestinal Disorders. Atlas Oral Maxillofac. Surg. Clin. N. Am. 2017, 25, 93–104. [Google Scholar] [CrossRef]
- Lankarani, K.B.; Sivandzadeh, G.R.; Hassanpour, S. Oral manifestation in inflammatory bowel disease: A review. World J. Gastroenterol. 2013, 19, 8571–8579. [Google Scholar] [CrossRef]
- Logan, R.M. Links between oral and gastrointestinal health. Curr. Opin. Support. Palliat. Care 2010, 4, 31–35. [Google Scholar] [CrossRef]
- Jajam, M.; Bozzolo, P.; Niklander, S. Oral manifestations of gastrointestinal disorders. J. Clin. Exp. Dent. 2017, 9, e1242–e1248. [Google Scholar] [CrossRef] [PubMed]
- Rowland, M.; Fleming, P.; Bourke, B. Looking in the mouth for Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Muhvić-Urek, M.; Tomac-Stojmenović, M.; Mijandrušić-Sinčić, B. Oral pathology in inflammatory bowel disease. World J. Gastroenterol. 2016, 22, 5655–5667. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, A.; Gullickson, R.G.; Singh, R.; Ro, S.; Omaye, S.T. The Link between Oral and Gut Microbiota in Inflammatory Bowel Disease and a Synopsis of Potential Salivary Biomarkers. Appl. Sci. 2020, 10, 6421. [Google Scholar] [CrossRef]
- Lucas López, R.; Grande Burgos, M.J.; Gálvez, A.; Pérez Pulido, R. The human gastrointestinal tract and oral microbiota in inflammatory bowel disease: A state of the science review. APMIS 2017, 125, 3–10. [Google Scholar] [CrossRef]
- Said, H.S.; Suda, W.; Nakagome, S.; Chinen, H.; Oshima, K.; Kim, S.; Kimura, R.; Iraha, A.; Ishida, H.; Fujita, J.; et al. Dysbiosis of salivary microbiota in inflammatory bowel disease and its association with oral immunological biomarkers. DNA Res. 2014, 21, 15–25. [Google Scholar] [CrossRef]
- Rautava, J.; Pinnell, L.J.; Vong, L.; Akseer, N.; Assa, A.; Sherman, P.M. Oral microbiome composition changes in mouse models of colitis. J. Gastroenterol. Hepatol. 2015, 30, 521–527. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.M.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, M.B.; Porter, S.R.; Smoller, B.R.; Sitaru, C. Oral mucosal manifestations of autoimmune skin diseases. Autoimmun. Rev. 2015, 14, 930–951. [Google Scholar] [CrossRef]
- Magliocca, K.R.; Fitzpatrick, S.G. Autoimmune Disease Manifestations in the Oral Cavity. Surg. Pathol. Clin. 2017, 10, 57–88. [Google Scholar] [CrossRef]
- Lourenço, S.; Hussein, T.; Bologna, S.; Sipahi, A.; Nico, M. Oral manifestations of inflammatory bowel disease: A review based on the observation of six cases. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Schifter, M.; Yeoh, S.-C.; Coleman, H.; Georgiou, A. Oral mucosal diseases: The inflammatory dermatoses. Aust. Dent. J. 2010, 55, 23–38. [Google Scholar] [CrossRef]
- Chi, A.C.; Neville, B.W.; Krayer, J.W.; Gonsalves, W.C. Oral manifestations of systemic disease. Am. Fam. Physician 2010, 82, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Saccucci, M.; Di Carlo, G.; Bossù, M.; Giovarruscio, F.; Salucci, A.; Polimeni, A. Autoimmune Diseases and Their Manifestations on Oral Cavity: Diagnosis and Clinical Management. J. Immunol. Res. 2018, 2018, 6061825. [Google Scholar] [CrossRef] [PubMed]
- Mays, J.W.; Sarmadi, M.; Moutsopoulos, N.M. Oral Manifestations of Systemic Autoimmune and Inflammatory Diseases: Diagnosis and Clinical Management. J. Evid. Based Dent. Pract. 2012, 12, 265–282. [Google Scholar] [CrossRef]
- Fuggle, N.R.; Smith, T.O.; Kaul, A.; Sofat, N. Hand to Mouth: A Systematic Review and Meta-Analysis of the Association between Rheumatoid Arthritis and Periodontitis. Front. Immunol. 2016, 7, 80. [Google Scholar] [CrossRef]
- Dalmády, S.; Kemény, L.; Antal, M.; Gyulai, R. Periodontitis: A newly identified comorbidity in psoriasis and psoriatic arthritis. Expert Rev. Clin. Immunol. 2020, 16, 101–108. [Google Scholar] [CrossRef]
- Okada, M.; Kobayashi, T.; Ito, S.; Yokoyama, T.; Abe, A.; Murasawa, A.; Yoshie, H. Periodontal Treatment Decreases Levels of Antibodies to Porphyromonas gingivalis and Citrulline in Patients With Rheumatoid Arthritis and Periodontitis. J. Periodontol. 2013, 84, e74–e84. [Google Scholar] [CrossRef]
- Teeuw, W.J.; Gerdes, V.E.A.; Loos, B.G. Effect of Periodontal Treatment on Glycemic Control of Diabetic Patients: A systematic review and meta-analysis. Diabetes Care 2010, 33, 421–427. [Google Scholar] [CrossRef]
- Keller, J.J.; Lin, H.-C. The effects of chronic periodontitis and its treatment on the subsequent risk of psoriasis. Br. J. Dermatol. 2012, 167, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Holmstrup, P.; Damgaard, C.; Olsen, I.; Klinge, B.; Flyvbjerg, A.; Nielsen, C.H.; Hansen, P.R. Comorbidity of periodontal disease: Two sides of the same coin? An introduction for the clinician. J. Oral Microbiol. 2017, 9, 1332710. [Google Scholar] [CrossRef]
- Pentony, P.; Duquenne, L.; Dutton, K.; Mankia, K.; Gul, H.; Vital, E.; Emery, P. The initiation of autoimmunity at epithelial surfaces: A focus on rheumatoid arthritis and systemic lupus erythematosus. Discov. Med. 2017, 24, 191–200. [Google Scholar]
- Mankia, K.; Emery, P. Is localized autoimmunity the trigger for rheumatoid arthritis? Unravelling new targets for prevention. Discov. Med. 2015, 20, 129–135. [Google Scholar] [PubMed]
- Wang, S.-S.; Tang, Y.-L.; Pang, X.; Zheng, M.; Tang, Y.-J.; Liang, X.-H. The maintenance of an oral epithelial barrier. Life Sci. 2019, 227, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Mikuls, T.R.; Payne, J.B.; Deane, K.D.; Thiele, G.M. Autoimmunity of the lung and oral mucosa in a multisystem inflammatory disease: The spark that lights the fire in rheumatoid arthritis? J. Allergy Clin. Immunol. 2016, 137, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Nikitakis, N.; Papaioannou, W.; Sakkas, L.; Kousvelari, E. The autoimmunity-oral microbiome connection. Oral Dis. 2017, 23, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Marchesan, J.T.; Gerow, E.A.; Schaff, R.; Taut, A.D.; Shin, S.-Y.; Sugai, J.; Brand, D.; Burberry, A.; Jorns, J.; Lundy, S.K.; et al. Porphyromonas gingivalis oral infection exacerbates the development and severity of collagen-induced arthritis. Arthritis Res. Ther. 2013, 15, R186. [Google Scholar] [CrossRef]
- Gaddey, H.L. Oral manifestations of systemic disease. Gen. Dent. 2017, 65, 23–29. [Google Scholar]
- Napeñas, J.J.; Brennan, M.T.; Elad, S. Oral Manifestations of Systemic Diseases. Dermatol. Clin. 2020, 38, 495–505. [Google Scholar] [CrossRef]
- Lawrence, H.P. Salivary markers of systemic disease: Noninvasive diagnosis of disease and monitoring of general health. J. Can. Dent. Assoc. 2002, 68, 170–174. [Google Scholar]
- Castagnola, M.; Picciotti, P.M.; Messana, I.; Fanali, C.; Fiorita, A.; Cabras, T.; Calò, L.; Pisano, E.; Passali, G.C.; Iavarone, F.; et al. Potential applications of human saliva as diagnostic fluid. Acta Otorhinolaryngol. Ital. 2011, 31, 347–357. [Google Scholar] [PubMed]
- Porter, S.R.; Mercadente, V.; Fedele, S. Oral manifestations of systemic disease. BDJ Team 2018, 5, 18012. [Google Scholar] [CrossRef][Green Version]
- Santosh, A.R.; Muddana, K. Viral infections of oral cavity. J. Fam. Med. Prim. Care 2020, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Rautemaa, R.; Lauhio, A.; Cullinan, M.P.; Seymour, G.J. Oral infections and systemic disease—An emerging problem in medicine. Clin. Microbiol. Infect. 2007, 13, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, S.N.; Hagner, M.; Nogueira, A.V.B.; Franke, A.; Jäger, A.; Deschner, J. Inflammatory bowel disease and oral health: Systematic review and a meta-analysis. J. Clin. Periodontol. 2017, 44, 382–393. [Google Scholar] [CrossRef]
- Bierbaumer, L.; Schwarze, U.Y.; Gruber, R.; Neuhaus, W. Cell culture models of oral mucosal barriers: A review with a focus on applications, culture conditions and barrier properties. Tissue Barriers 2018, 6, 1479568. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Berkovitz, B.K. Oral cavity. In Gray’s Anatomy, 41st ed.; Elsevier: Amsterdam, The Netherlands, Chapter 31; 2015. [Google Scholar]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.A. Barrier properties of mucus. Adv. Drug Deliv. Rev. 2009, 61, 75–85. [Google Scholar] [CrossRef]
- Yadev, N.P.; Murdoch, C.; Saville, S.P.; Thornhill, M.H. Evaluation of tissue engineered models of the oral mucosa to investigate oral candidiasis. Microb. Pathog. 2011, 50, 278–285. [Google Scholar] [CrossRef]
- Nielsen, H.M.; Rassing, M.R. TR146 cells grown on filters as a model of human buccal epithelium: IV. Permeability of water, mannitol, testosterone and beta-adrenoceptor antagonists. Comparison to human, monkey and porcine buccal mucosa. Int. J. Pharm. 2000, 194, 155–167. [Google Scholar] [CrossRef]
- Moharamzadeh, K.; Colley, H.; Murdoch, C.; Hearnden, V.; Chai, W.L.; Brook, I.M.; Thornhill, M.H.; Macneil, S. Tissue-engineered oral mucosa. J. Dent. Res. 2012, 91, 642–650. [Google Scholar] [CrossRef]
- Kinikoglu, B.; Damour, O.; Hasirci, V. Tissue engineering of oral mucosa: A shared concept with skin. J. Artif. Organs 2015, 18, 8–19. [Google Scholar] [CrossRef]
- Mohd Nor, N.H.; Berahim, Z.; Ahmad, A.; Kannan, T.P. Properties of Cell Sources in Tissue-Engineered Three-dimensional Oral Mucosa Model: A Review. Curr. Stem Cell Res. Ther. 2017, 12, 52–60. [Google Scholar] [CrossRef]
- Guerra, E.N.S.; Acevedo, A.C.; de Toledo, I.P.; Combes, A.; Chardin, H. Do mucosal biomarkers reveal the immunological state associated with food allergy? Allergy 2018, 73, 2392–2394. [Google Scholar] [CrossRef]
- Peeters, K.A.B.M.; Lamers, R.-J.A.N.; Penninks, A.H.; Knol, E.F.; Bruijnzeel-Koomen, C.A.F.M.; van Nesselrooij, J.H.J.; Knulst, A.C. A search for biomarkers as diagnostic tools for food allergy: A pilot study in peanut-allergic patients. Int. Arch. Allergy Immunol. 2011, 155, 23–30. [Google Scholar] [CrossRef]
- Tomicić, S.; Norrman, G.; Fälth-Magnusson, K.; Jenmalm, M.C.; Devenney, I.; Böttcher, M.F. High levels of IgG4 antibodies to foods during infancy are associated with tolerance to corresponding foods later in life. Pediatr. Allergy Immunol. 2009, 20, 35–41. [Google Scholar] [CrossRef]
- Streckfus, C.F.; Bigler, L.R. Saliva as a diagnostic fluid. Oral Dis. 2002, 8, 69–76. [Google Scholar] [CrossRef]
- Streckfus, C.; Bigler, L.; Navazesh, M.; Al-Hashimi, I. Cytokine concentrations in stimulated whole saliva among patients with primary Sjögren’s syndrome, secondary Sjögren’s syndrome, and patients with primary Sjögren’s syndrome receiving varying doses of interferon for symptomatic treatment of the condition. Clin. Oral Investig. 2001, 5, 133–135. [Google Scholar] [CrossRef]
- Lorenzo-Pouso, A.I.; Pérez-Sayáns, M.; Bravo, S.B.; López-Jornet, P.; García-Vence, M.; Alonso-Sampedro, M.; Carballo, J.; García-García, A. Protein-Based Salivary Profiles as Novel Biomarkers for Oral Diseases. Dis. Markers 2018, 2018, 6141845. [Google Scholar] [CrossRef]
- Greabu, M.; Battino, M.; Mohora, M.; Totan, A.; Didilescu, A.; Spinu, T.; Totan, C.; Miricescu, D.; Radulescu, R. Saliva—A diagnostic window to the body, both in health and in disease. J. Med. Life 2009, 2, 124–132. [Google Scholar] [PubMed]
- Gröschl, M. Saliva: A reliable sample matrix in bioanalytics. Bioanalysis 2017, 9, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Kaczor-Urbanowicz, K.E.; Martin Carreras-Presas, C.; Aro, K.; Tu, M.; Garcia-Godoy, F.; Wong, D.T. Saliva diagnostics—Current views and directions. Exp. Biol. Med. 2017, 242, 459–472. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Bardellini, E.; Amadori, F.; Ravelli, A.; Salemme, M.; Lonardi, S.; Villanacci, V.; Majorana, A. Histopathological findings in the oral mucosa of celiac patients. Rev. Esp. Enferm. Dig. 2014, 106, 86–91. [Google Scholar] [CrossRef][Green Version]
- Lähteenoja, H.; Toivanen, A.; Viander, M.; Räihä, I.; Rantala, I.; Syrjänen, S.; Mäki, M.; Lahteenoja, H.; Toivanen, A.; Viander, M.; et al. Increase in T-cell subsets of oral mucosa: A late immune response in patients with treated coeliac disease? Scand. J. Immunol. 2000, 52, 602–608. [Google Scholar] [CrossRef]
- Rashid, M.; Zarkadas, M.; Anca, A.; Limeback, H. Oral manifestations of celiac disease: A clinical guide for dentists. J. Can. Dent. Assoc. 2011, 77, b39. [Google Scholar]
- Sapone, A.; Bai, J.C.; Ciacci, C.; Dolinsek, J.; Green, P.H.R.; Hadjivassiliou, M.; Kaukinen, K.; Rostami, K.; Sanders, D.S.; Schumann, M.; et al. Spectrum of gluten-related disorders: Consensus on new nomenclature and classification. BMC Med. 2012, 10. [Google Scholar] [CrossRef]
- Bao, F.; Green, P.H.; Bhagat, G. An Update on Celiac Disease Histopathology and the Road Ahead. Arch. Pathol. Lab. Med. 2012, 136, 735–745. [Google Scholar] [CrossRef]
- Fasano, A.; Sapone, A.; Zevallos, V.; Schuppan, D. Nonceliac gluten sensitivity. Gastroenterology 2015, 148, 1195–1204. [Google Scholar] [CrossRef]
- Bizzaro, N.; Tozzoli, R.; Villalta, D.; Fabris, M.; Tonutti, E. Cutting-edge issues in celiac disease and in gluten intolerance. Clin. Rev. Allergy Immunol. 2012, 42, 279–287. [Google Scholar] [CrossRef]
- Troncone, R.; Jabri, B. Coeliac disease and gluten sensitivity. J. Intern. Med. 2011, 269, 582–590. [Google Scholar] [CrossRef]
- Picarelli, A.; Di Tola, M.; Vallecoccia, A.; Libanori, V.; Magrelli, M.; Carlesimo, M.; Rossi, A. Oral mucosa patch test: A new tool to recognize and study the adverse effects of dietary nickel exposure. Biol. Trace Elem. Res. 2011, 139, 151–159. [Google Scholar] [CrossRef]
- Di Tola, M.; Amodeo, R.; Marino, M.; Tabacco, F.; Casale, R.; Bove, M.; Rossi, A.; Cardelli, P.; Picarelli, A. Peripheral blood lymphocyte typing as a useful tool to objectify the oral mucosa patch test in the diagnosis of allergic contact mucositis to nickel. Biol. Trace Elem. Res. 2014, 159, 81–86. [Google Scholar] [CrossRef]
- Di Tola, M.; Marino, M.; Amodeo, R.; Tabacco, F.; Casale, R.; Portaro, L.; Borghini, R.; Cristaudo, A.; Manna, F.; Rossi, A.; et al. Immunological characterization of the allergic contact mucositis related to the ingestion of nickel-rich foods. Immunobiology 2014, 219, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Picarelli, A.; Borghini, R.; Di Tola, M.; Marino, M.; Urciuoli, C.; Isonne, C.; Puzzono, M.; Porowska, B.; Rumi, G.; Lonardi, S.; et al. Intestinal, Systemic, and Oral Gluten-related Alterations in Patients With Nonceliac Gluten Sensitivity. J. Clin. Gastroenterol. 2016, 50, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Hering, N.A.; Schulzke, J.-D. Therapeutic options to modulate barrier defects in inflammatory bowel disease. Dig. Dis. 2009, 27, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, Y.; Tanabe, S.; Suzuki, T. Cellular zinc is required for intestinal epithelial barrier maintenance via the regulation of claudin-3 and occludin expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G105–G116. [Google Scholar] [CrossRef] [PubMed]
- Valenzano, M.C.; DiGuilio, K.; Mercado, J.; Teter, M.; To, J.; Ferraro, B.; Mixson, B.; Manley, I.; Baker, V.; Moore, B.A.; et al. Remodeling of Tight Junctions and Enhancement of Barrier Integrity of the CACO-2 Intestinal Epithelial Cell Layer by Micronutrients. PLoS ONE 2015, 10, e0133926. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, Z.; Yang, Z.; Zhao, N.; Yuan, W. Self-healing efficiency of cementitious materials containing microcapsules filled with healing adhesive: Mechanical restoration and healing process monitored by water absorption. PLoS ONE 2013, 8, e81616. [Google Scholar] [CrossRef] [PubMed]
- Sturniolo, G.C.; Fries, W.; Mazzon, E.; Di Leo, V.; Barollo, M.; D’inca, R. Effect of zinc supplementation on intestinal permeability in experimental colitis. J. Lab. Clin. Med. 2002, 139, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, D.; Rancan, S.; Orso, G.; Dall’Acqua, S.; Brun, P.; Giron, M.C.; Carrara, M.; Castagliuolo, I.; Ragazzi, E.; Caparrotta, L.; et al. Boswellia serrata Preserves Intestinal Epithelial Barrier from Oxidative and Inflammatory Damage. PLoS ONE 2015, 10, e0125375. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Role of flavonoids in intestinal tight junction regulation. J. Nutr. Biochem. 2011, 22, 401–408. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Morales, P.; Gotteland, M. Polyphenols protect the epithelial barrier function of Caco-2 cells exposed to indomethacin through the modulation of occludin and zonula occludens-1 expression. J. Agric. Food Chem. 2013, 61, 5291–5297. [Google Scholar] [CrossRef]
- Elias, P.M.; Friend, D.S. Vitamin-A-induced mucous metaplasia. An in vitro system for modulating tight and gap junction differentiation. J. Cell Biol. 1976, 68, 173–188. [Google Scholar] [CrossRef]
- Ma, X.; Fan, P.X.; Li, L.S.; Qiao, S.Y.; Zhang, G.L.; Li, D.F. Butyrate promotes the recovering of intestinal wound healing through its positive effect on the tight junctions. J. Anim. Sci. 2012, 90 (Suppl. 4), 266–268. [Google Scholar] [CrossRef]
- Song, P.; Zhang, R.; Wang, X.; He, P.; Tan, L.; Ma, X. Dietary grape-seed procyanidins decreased postweaning diarrhea by modulating intestinal permeability and suppressing oxidative stress in rats. J. Agric. Food Chem. 2011, 59, 6227–6232. [Google Scholar] [CrossRef]
- Rybakovsky, E.; Valenzano, M.C.; Deis, R.; DiGuilio, K.M.; Thomas, S.; Mullin, J.M. Improvement of Human-Oral-Epithelial-Barrier Function and of Tight Junctions by Micronutrients. J. Agric. Food Chem. 2017, 65, 10950–10958. [Google Scholar] [CrossRef]
- Groeger, S.; Jarzina, F.; Windhorst, A.; Meyle, J. Influence of retinoic acid on human gingival epithelial barriers. J. Periodontal Res. 2016, 51, 748–757. [Google Scholar] [CrossRef]
- Bae, M.-J.; Shin, H.S.; See, H.-J.; Jung, S.Y.; Kwon, D.-A.; Shon, D.-H. Baicalein induces CD4+Foxp3+ T cells and enhances intestinal barrier function in a mouse model of food allergy. Sci. Rep. 2016, 6, 32225. [Google Scholar] [CrossRef]
- Xie, R.-D.; Xu, L.-Z.; Yang, L.-T.; Wang, S.; Liu, Q.; Liu, Z.-G.; Yang, P.-C. Galectin-1 inhibits oral-intestinal allergy syndrome. Oncotarget 2017, 8, 13214–13222. [Google Scholar] [CrossRef]
- Antunes, M.M.; Coelho, B.S.L.; Vichi, T.M.; dos Santos, E.A.; Gondim, F.K.B.; Diniz, A.B.; Aguilar, E.C.; Cara, D.C.; Porto, L.C.J.; de Castro, I.C.; et al. Oral supplementation with capsaicin reduces oxidative stress and IL-33 on a food allergy murine model. World Allergy Organ. J. 2019, 12, 100045. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, R.U.; Aijima, R.; Ohyama, Y.; Yoshizumi, J.; Kitsuki, T.; Ohsaki, Y.; Cao, A.-L.; Danjo, A.; Yamashita, Y.; Kiyoshima, T.; et al. Impaired Junctions and Invaded Macrophages in Oral Epithelia With Oral Pain. J. Histochem. Cytochem. 2019, 67, 245–256. [Google Scholar] [CrossRef]
- Shakya, A.K.; Chowdhury, M.Y.E.; Tao, W.; Gill, H.S. Mucosal vaccine delivery: Current state and a pediatric perspective. J. Control. Release 2016, 240, 394–413. [Google Scholar] [CrossRef]
- Hua, S. Advances in Nanoparticulate Drug Delivery Approaches for Sublingual and Buccal Administration. Front. Pharmacol. 2019, 10, 1328. [Google Scholar] [CrossRef]
- New, R.R.C. Formulation technologies for oral vaccines. Clin. Exp. Immunol. 2019, 198, 153–169. [Google Scholar] [CrossRef]
- He, S.; Liu, Z.; Xu, D. Advance in oral delivery systems for therapeutic protein. J. Drug Target. 2019, 27, 283–291. [Google Scholar] [CrossRef]
- Reinholz, J.; Landfester, K.; Mailänder, V. The challenges of oral drug delivery via nanocarriers. Drug Deliv. 2018, 25, 1694–1705. [Google Scholar] [CrossRef]
- Creighton, R.L.; Woodrow, K.A. Microneedle-Mediated Vaccine Delivery to the Oral Mucosa. Adv. Healthc. Mater. 2019, 8, e1801180. [Google Scholar] [CrossRef]
- Incorvaia, C.; Frati, F.; Sensi, L.; Riario-Sforza, G.G.; Marcucci, F. Allergic inflammation and the oral mucosa. Recent Pat. Inflamm. Allergy Drug Discov. 2007, 1, 35–38. [Google Scholar] [CrossRef]
- Incorvaia, C.; Fuiano, N.; Frati, F.; Marcucci, F. Advances of knowledge on allergic inflammation and the oral mucosa. Recent Pat. Inflamm. Allergy Drug Discov. 2014, 8, 173–177. [Google Scholar] [CrossRef]
- Canonica, G.W.; Cox, L.; Pawankar, R.; Baena-Cagnani, C.E.; Blaiss, M.; Bonini, S.; Bousquet, J.; Calderón, M.; Compalati, E.; Durham, S.R.; et al. Sublingual immunotherapy: World Allergy Organization position paper 2013 update. World Allergy Organ. J. 2014, 7, 6. [Google Scholar] [CrossRef]
- Barber, D.; Rico, P.; Blanco, C.; Fernandez-Rivas, M.; Ibañez, M.D.; Escribese, M.M. GRAZAX®: A sublingual immunotherapy vaccine for Hay fever treatment: From concept to commercialization. Hum. Vaccin. Immunother. 2019, 15, 2887–2895. [Google Scholar] [CrossRef]
- Gueguen, C.; Bouley, J.; Moussu, H.; Luce, S.; Duchateau, M.; Chamot-Rooke, J.; Pallardy, M.; Lombardi, V.; Nony, E.; Baron-Bodo, V.; et al. Changes in markers associated with dendritic cells driving the differentiation of either TH2 cells or regulatory T cells correlate with clinical benefit during allergen immunotherapy. J. Allergy Clin. Immunol. 2016, 137, 545–558. [Google Scholar] [CrossRef]
- Suárez-Fueyo, A.; Ramos, T.; Galán, A.; Jimeno, L.; Wurtzen, P.A.; Marin, A.; de Frutos, C.; Blanco, C.; Carrera, A.C.; Barber, D.; et al. Grass tablet sublingual immunotherapy downregulates the TH2 cytokine response followed by regulatory T-cell generation. J. Allergy Clin. Immunol. 2014, 133, 130–138. [Google Scholar] [CrossRef]
- Barker-Tejeda, T.C.; Bazire, R.; Obeso, D.; Mera-Berriatua, L.; Rosace, D.; Vazquez-Cortes, S.; Ramos, T.; Rico, M.D.P.; Chivato, T.; Barbas, C.; et al. Exploring novel systemic biomarker approaches in grass-pollen sublingual immunotherapy using omics. Allergy 2020. [Google Scholar] [CrossRef] [PubMed]
- Enrique, E.; Pineda, F.; Malek, T.; Bartra, J.; Basagaña, M.; Tella, R.; Castelló, J.V.; Alonso, R.; De Mateo, J.A.; Cerdá-Trias, T.; et al. Sublingual immunotherapy for hazelnut food allergy: A randomized, double-blind, placebo-controlled study with a standardized hazelnut extract. J. Allergy Clin. Immunol. 2005, 116, 1073–1079. [Google Scholar] [CrossRef]
- Fernández-Rivas, M.; Garrido Fernández, S.; Nadal, J.A.; Díaz de Durana, M.D.A.; García, B.E.; González-Mancebo, E.; Martín, S.; Barber, D.; Rico, P.; Tabar, A.I. Randomized double-blind, placebo-controlled trial of sublingual immunotherapy with a Pru p 3 quantified peach extract. Allergy 2009, 64, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.; Bogas, G.; Gonzalez, M.; Campo, P.; Salas, M.; Diaz-Perales, A.; Rodriguez, M.J.; Prieto, A.; Barber, D.; Blanca, M.; et al. The clinical and immunological effects of Pru p 3 sublingual immunotherapy on peach and peanut allergy in patients with systemic reactions. Clin. Exp. Allergy 2017, 47, 339–350. [Google Scholar] [CrossRef]
- Palomares, F.; Gomez, F.; Bogas, G.; Campo, P.; Perkins, J.R.; Diaz-Perales, A.; Rodriguez, M.J.; Prieto, A.; Barber, D.; Torres, M.J.; et al. Immunological Changes Induced in Peach Allergy Patients with Systemic Reactions by Pru p 3 Sublingual Immunotherapy. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Fleischer, D.M.; Burks, A.W.; Vickery, B.P.; Scurlock, A.M.; Wood, R.A.; Jones, S.M.; Sicherer, S.H.; Liu, A.H.; Stablein, D.; Henning, A.K.; et al. Sublingual immunotherapy for peanut allergy: A randomized, double-blind, placebo-controlled multicenter trial. J. Allergy Clin. Immunol. 2013, 131, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Burks, A.W.; Wood, R.A.; Jones, S.M.; Sicherer, S.H.; Fleischer, D.M.; Scurlock, A.M.; Vickery, B.P.; Liu, A.H.; Henning, A.K.; Lindblad, R.; et al. Sublingual immunotherapy for peanut allergy: Long-term follow-up of a randomized multicenter trial. J. Allergy Clin. Immunol. 2015, 135, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.; Whyte, A. Immunology of the tonsils. Immunol. Today 1998, 19, 414–421. [Google Scholar] [CrossRef]
- Palomares, O.; Rückert, B.; Jartti, T.; Kücüksezer, U.C.; Puhakka, T.; Gomez, E.; Fahrner, H.B.; Speiser, A.; Jung, A.; Kwok, W.W.; et al. Induction and maintenance of allergen-specific FOXP3+ Treg cells in human tonsils as potential first-line organs of oral tolerance. J. Allergy Clin. Immunol. 2012, 129, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Palomares, O.; Martín-Fontecha, M.; Lauener, R.; Traidl-Hoffmann, C.; Cavkaytar, O.; Akdis, M.; Akdis, C.A. Regulatory T cells and immune regulation of allergic diseases: Roles of IL-10 and TGF-β. Genes Immun. 2014, 15, 511–520. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Jansen, K.; Głobińska, A.; van de Veen, W.; Akdis, M. Regulatory Immune Mechanisms in Tolerance to Food Allergy. Front. Immunol. 2018, 9, 2939. [Google Scholar] [CrossRef]
- Nolte, M.; Barber, D.; Maloney, J.; Li, Z.; Kaur, A.; Galan, A.; Andersen, J.S.; Nolte, H. Timothy specific IgE levels are associated with efficacy and safety of timothy grass sublingual immunotherapy tablet. Ann. Allergy. Asthma Immunol. 2015, 115, 509–515.e2. [Google Scholar] [CrossRef]
- FitzPatrick, M.E.B.; Provine, N.M.; Garner, L.C.; Powell, K.; Amini, A.; Irwin, S.L.; Ferry, H.; Ambrose, T.; Friend, P.; Vrakas, G.; et al. Human intestinal tissue-resident memory T cells comprise transcriptionally and functionally distinct subsets. Cell Rep. 2021, 34, 108661. [Google Scholar] [CrossRef]
- Krishnan, S.; Prise, I.E.; Wemyss, K.; Schenck, L.P.; Bridgeman, H.M.; McClure, F.A.; Zangerle-Murray, T.; O’Boyle, C.; Barbera, T.A.; Mahmood, F.; et al. Amphiregulin-producing γδ T cells are vital for safeguarding oral barrier immune homeostasis. Proc. Natl. Acad. Sci. USA 2018, 115, 10738–10743. [Google Scholar] [CrossRef]
- Zhang, C.; Li, L.; Feng, K.; Fan, D.; Xue, W.; Lu, J. ‘Repair’ Treg Cells in Tissue Injury. Cell. Physiol. Biochem. 2017, 43, 2155–2169. [Google Scholar] [CrossRef]
- Cook, L.; Munier, C.M.L.; Seddiki, N.; van Bockel, D.; Ontiveros, N.; Hardy, M.Y.; Gillies, J.K.; Levings, M.K.; Reid, H.H.; Petersen, J.; et al. Circulating gluten-specific FOXP3 + CD39 + regulatory T cells have impaired suppressive function in patients with celiac disease. J. Allergy Clin. Immunol. 2017, 140, 1592–1603.e8. [Google Scholar] [CrossRef] [PubMed]
- Granzotto, M.; dal Bo, S.; Quaglia, S.; Tommasini, A.; Piscianz, E.; Valencic, E.; Ferrara, F.; Martelossi, S.; Ventura, A.; Not, T. Regulatory T-Cell Function Is Impaired in Celiac Disease. Dig. Dis. Sci. 2009, 54, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Holtan, S.G.; DeFor, T.E.; Panoskaltsis-Mortari, A.; Khera, N.; Levine, J.E.; Flowers, M.E.D.; Lee, S.J.; Inamoto, Y.; Chen, G.L.; Mayer, S.; et al. Amphiregulin modifies the Minnesota Acute Graft-versus-Host Disease Risk Score: Results from BMT CTN 0302/0802. Blood Adv. 2018, 2, 1882–1888. [Google Scholar] [CrossRef]
- Jones, A.C.; Anderson, D.; Galbraith, S.; Fantino, E.; Gutierrez Cardenas, D.; Read, J.F.; Serralha, M.; Holt, B.J.; Strickland, D.H.; Sly, P.D.; et al. Personalized Transcriptomics Reveals Heterogeneous Immunophenotypes in Children with Viral Bronchiolitis. Am. J. Respir. Crit. Care Med. 2019, 199, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Hirahara, K.; Aoki, A.; Morimoto, Y.; Kiuchi, M.; Okano, M.; Nakayama, T. The immunopathology of lung fibrosis: Amphiregulin-producing pathogenic memory T helper-2 cells control the airway fibrotic responses by inducing eosinophils to secrete osteopontin. Semin. Immunopathol. 2019, 41, 339–348. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Casado, C.; Sanchez-Solares, J.; Izquierdo, E.; Díaz-Perales, A.; Barber, D.; Escribese, M.M. Oral Mucosa as a Potential Site for Diagnosis and Treatment of Allergic and Autoimmune Diseases. Foods 2021, 10, 970. https://doi.org/10.3390/foods10050970
Gomez-Casado C, Sanchez-Solares J, Izquierdo E, Díaz-Perales A, Barber D, Escribese MM. Oral Mucosa as a Potential Site for Diagnosis and Treatment of Allergic and Autoimmune Diseases. Foods. 2021; 10(5):970. https://doi.org/10.3390/foods10050970
Chicago/Turabian StyleGomez-Casado, Cristina, Javier Sanchez-Solares, Elena Izquierdo, Araceli Díaz-Perales, Domingo Barber, and María M. Escribese. 2021. "Oral Mucosa as a Potential Site for Diagnosis and Treatment of Allergic and Autoimmune Diseases" Foods 10, no. 5: 970. https://doi.org/10.3390/foods10050970
APA StyleGomez-Casado, C., Sanchez-Solares, J., Izquierdo, E., Díaz-Perales, A., Barber, D., & Escribese, M. M. (2021). Oral Mucosa as a Potential Site for Diagnosis and Treatment of Allergic and Autoimmune Diseases. Foods, 10(5), 970. https://doi.org/10.3390/foods10050970