
Impact of Processing Technology on Macro- and Micronutrient Profile of Protein-Enriched Products from Fish Backbones


Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Backbone Samples
2.2. pH-Shift Process and Mechanical Separation
2.3. Protein Recovery Measurement
2.4. Compositional Analyses
2.5. Amino Acid Analysis
2.6. Fatty Acid Composition Measurement
2.7. Measurement of Vitamins D, E and C
2.8. Measurement of Minerals
2.9. Statistical Analysis
3. Results and Discussion
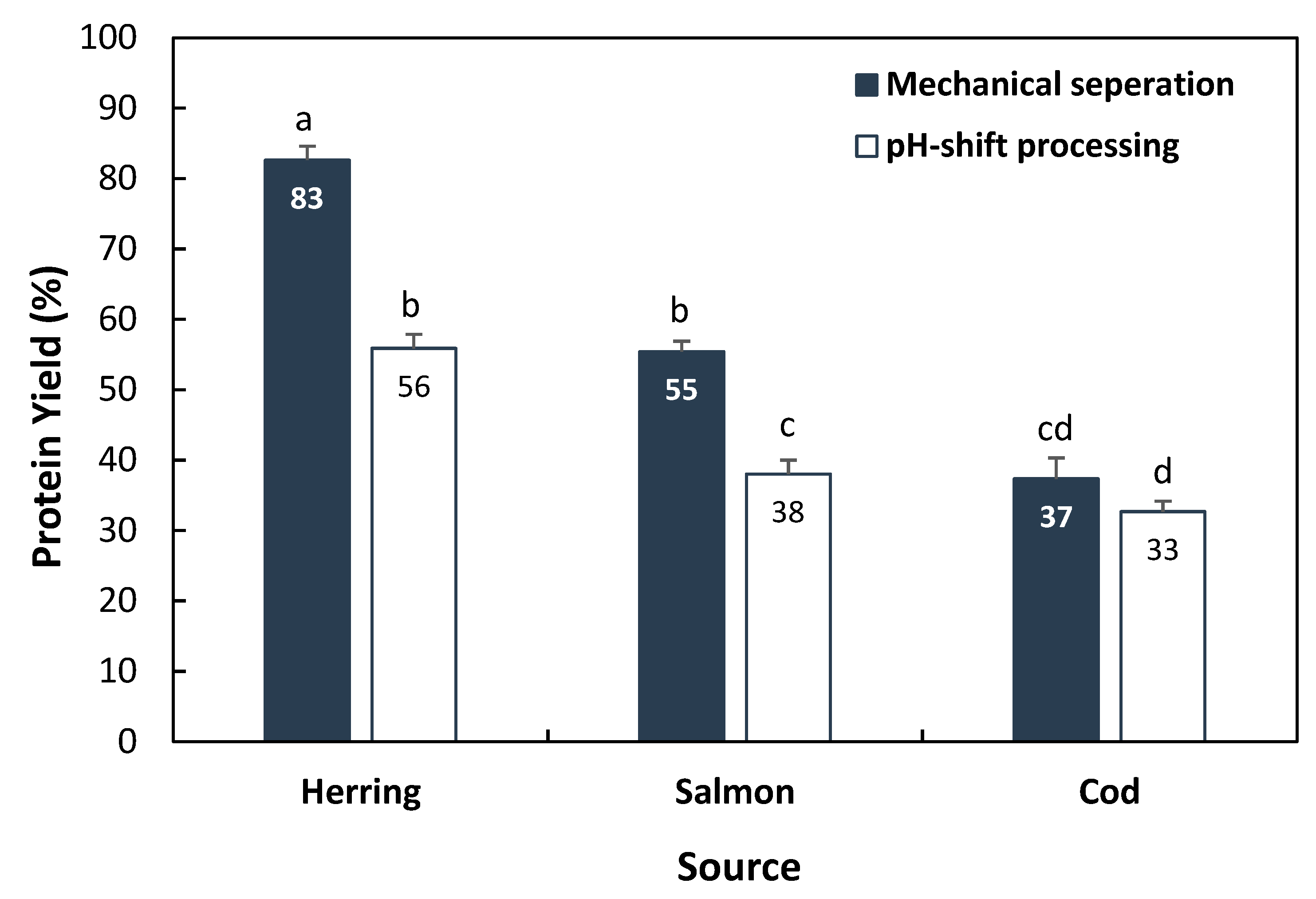
3.1. Protein Recovery from Fish Backbones
3.2. Macronutrients of the Recovered Protein-Enriched Products
3.3. Amino Acid Composition
3.4. Fatty Acid Composition
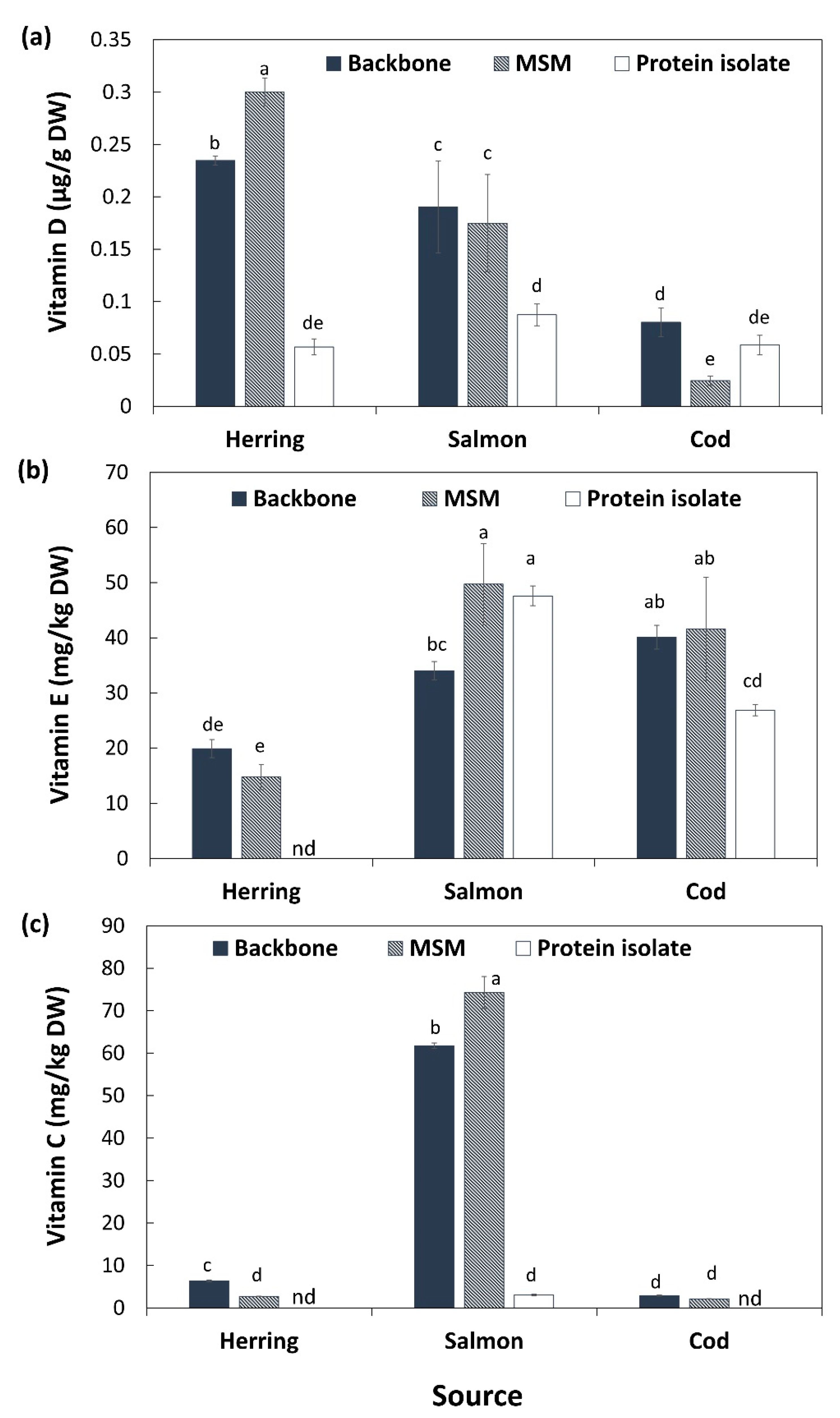
3.5. Vitamins
3.6. Content of Minerals
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization. World Fisheries and Aquaculture Sofia Report; FAO: Rome, Italy, 2018; ISBN 9789251072257. [Google Scholar]
- Ferraro, V.; Cruz, I.B.; Jorge, R.F.; Malcata, F.X.; Pintado, M.E.; Castro, P.M.L. Valorisation of natural extracts from marine source focused on marine by-products: A review. Food Res. Int. 2010, 43, 2221–2233. [Google Scholar] [CrossRef]
- Yuste, J.; Mor-Mur, M.; Capellas, M.; Guamis, B.; Pla, R. Mechanically recovered poultry meat sausages manufactured with high hydrostatic pressure. Poult. Sci. 1999, 78, 914–921. [Google Scholar] [CrossRef]
- Palmeira, K.R.; Mársico, E.T.; Monteiro, M.L.G.; Lemos, M.; Conte Junior, C.A. Ready-to-eat products elaborated with mechanically separated fish meat from waste processing: Challenges and chemical quality. CYTA J. Food 2016, 14, 227–238. [Google Scholar] [CrossRef]
- Secci, G.; Borgogno, M.; Mancini, S.; Paci, G.; Parisi, G. Mechanical separation process for the value enhancement of Atlantic horse mackerel (Trachurus trachurus), a discard fish. Innov. Food Sci. Emerg. Technol. 2017, 39, 13–18. [Google Scholar] [CrossRef]
- Newman, P.B. The separation of meat from bone-A review of the mechanics and the problems. Meat Sci. 1981, 5, 171–200. [Google Scholar] [CrossRef]
- Hrynets, Y.; Omana, D.A.; Xu, Y.; Betti, M. Comparative study on the effect of acid- and alkaline-aided extractions on mechanically separated turkey meat (MSTM): Chemical characteristics of recovered proteins. Process Biochem. 2011, 46, 335–343. [Google Scholar] [CrossRef]
- Secci, G.; Borgogno, M.; Lupi, P.; Rossi, S.; Paci, G.; Mancini, S.; Bonelli, A.; Parisi, G. Effect of mechanical separation process on lipid oxidation in European aquacultured sea bass, gilthead sea bream, and rainbow trout products. Food Control 2016, 67, 75–81. [Google Scholar] [CrossRef]
- Abdollahi, M.; Undeland, I. A novel cold biorefinery approach for isolation of high quality fish oil in parallel with gel-forming proteins. Food Chem. 2020, 127294. [Google Scholar] [CrossRef]
- Chen, Y.C.; Tou, J.C.; Jaczynski, J. Amino acid, fatty acid, and mineral profiles of materials recovered from rainbow trout (Oncorhynchus mykiss) processing by-products using isoelectric solubilization/precipitation. J. Food Sci. 2007, 72. [Google Scholar] [CrossRef]
- Abdollahi, M.; Undeland, I. Structural, functional, and sensorial properties of protein isolate produced from salmon, cod, and herring by-products. Food Bioprocess Technol. 2018, 11, 1733–1749. [Google Scholar] [CrossRef]
- Surasani, V.K.R. Acid and alkaline solubilization (pH shift) process: A better approach for the utilization of fish processing waste and by-products. Environ. Sci. Pollut. Res. 2018, 25, 18345–18363. [Google Scholar] [CrossRef] [PubMed]
- Abbasiliasi, S.; Tan, J.; Masomian, M. pH Shift Solubilization and Precipitation Protein Extraction from the Wastes of the Threadfin Bream, Nemipterus japonicus. J. Biochem. Microbiol. Biotechnol. 2019, 7, 10–16. [Google Scholar]
- Abdollahi, M.; Undeland, I. Physicochemical and gel-forming properties of protein isolated from salmon, cod and herring by-products using the pH-shift method. Lwt 2019, 101, 678–684. [Google Scholar] [CrossRef]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting nitrogen into protein—Beyond 6.25 and Jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Lee, C.M.; Trevino, B.; Chaiyawat, M. A simple and rapid solvent extraction method for determining total lipids in fish tissue. J. AOAC Int. 1995, 79, 487–492. [Google Scholar] [CrossRef]
- Undeland, I.; Hultin, H.O.; Richards, M.P. Added triacylglycerols do not hasten hemoglobin-mediated lipid oxidation in washed minced cod muscle. J. Agric. Food Chem. 2002, 50, 6847–6853. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef]
- Özcan, S.; Şenyuva, H.Z. Improved and simplified liquid chromatography/atmospheric pressure chemical ionization mass spectrometry method for the analysis of underivatized free amino acids in various foods. J. Chromatogr. A 2006, 1135, 179–185. [Google Scholar] [CrossRef]
- Tullberg, C.; Larsson, K.; Carlsson, N.G.; Comi, I.; Scheers, N.; Vegarud, G.; Undeland, I. Formation of reactive aldehydes (MDA, HHE, HNE) during the digestion of cod liver oil: Comparison of human and porcine in vitro digestion models. Food Funct. 2016, 7, 1401–1412. [Google Scholar] [CrossRef]
- Cavonius, L.R.; Carlsson, N.G.; Undeland, I. Quantification of total fatty acids in microalgae: Comparison of extraction and transesterification methods. Anal. Bioanal. Chem. 2014, 406, 7313–7322. [Google Scholar] [CrossRef]
- Standal, I.B.; Mozuraityte, R.; Rustad, T.; Alinasabhematabadi, L.; Carlsson, N.G.; Undeland, I. Quality of Filleted Atlantic Mackerel (Scomber Scombrus) During Chilled and Frozen Storage: Changes in Lipids, Vitamin D, Proteins, and Small Metabolites, including Biogenic Amines. J. Aquat. Food Prod. Technol. 2018, 27, 338–357. [Google Scholar] [CrossRef]
- Jäpelt, R.B.; Silvestro, D.; Smedsgaard, J.; Jensen, P.E.; Jakobsen, J. LC-MS/MS with atmospheric pressure chemical ionisation to study the effect of UV treatment on the formation of vitamin D3 and sterols in plants. Food Chem. 2011, 129, 217–225. [Google Scholar] [CrossRef]
- Larsson, K.; Almgren, A.; Undeland, I. Hemoglobin-mediated lipid oxidation and compositional characteristics of washed fish mince model systems made from cod (Gadus morhua), herring (Clupea harengus), and salmon (Salmo salar) muscle. J. Agric. Food Chem. 2007, 55, 9027–9035. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Perez Trueba, G.; Poulsen, H.E.; Christen, S. Vitamin C deficiency in weanling guinea pigs: Differential expression of oxidative stress and DNA repair in liver and brain. Br. J. Nutr. 2007, 98, 1116–1119. [Google Scholar] [CrossRef]
- Hornsey, H.C. The colour of cooked cured pork. I.—Estimation of the Nitric oxide-Haem Pigments. J. Sci. Food Agric. 1956, 7, 534–540. [Google Scholar] [CrossRef]
- Borgogno, M.; Husein, Y.; Secci, G.; Masi, S.; Parisi, G. Technological and nutritional advantages of mechanical separation process applied to three European aquacultured species. LWT Food Sci. Technol. 2017, 84, 298–305. [Google Scholar] [CrossRef]
- Chomnawang, C.; Yongsawatdigul, J. Protein Recovery of Tilapia Frame By-Products by pH-Shift Method. J. Aquat. Food Prod. Technol. 2013, 22, 112–120. [Google Scholar] [CrossRef]
- Shi, L.; Beamer, S.K.; Yin, T.; Matak, K.E.; Yang, H.; Jaczynski, J. Mass balance for isoelectric solubilization/precipitation of carp, chicken, menhaden, and krill. LWT Food Sci. Technol. 2017, 81, 26–34. [Google Scholar] [CrossRef]
- Marmon, S.K.; Undeland, I. Protein isolation from gutted herring (Clupea harengus) using pH-shift processes. J. Agric. Food Chem. 2010, 58, 10480–10486. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Tou, J.C.; Jaczynski, J. Amino acid and mineral composition of protein and other components and their recovery yields from whole Antarctic krill (Euphausia superba) using isoelectric solubilization/precipitation. J. Food Sci. 2009, 74, H31–H39. [Google Scholar] [CrossRef] [PubMed]
- Taskaya, L.; Chen, Y.-C.; Beamer, S.; Tou, J.C.; Jaczynski, J. Compositional characteristics of materials recovered from whole gutted silver carp (Hypophthalmichthys molitrix) using isoelectric solubilization/precipitation. J. Agric. Food Chem. 2009, 57, 4259–4266. [Google Scholar] [CrossRef] [PubMed]
- WHO; FAO; UNU. Protein and amino acid requirements in human nutrition. World Health Organ. Tech. Rep. Ser. 2007, 1–265. [Google Scholar]
- Zeng, D.; Mai, K.; Ai, Q.; Milley, J.E.; Lall, S.P. Lipid and fatty acid compositions of cod (Gadus morhua), haddock (Melanogrammus aeglefinus) and halibut (Hippoglossus hippoglossus). J. Ocean Univ. China 2010, 9, 381–388. [Google Scholar] [CrossRef]
- Castro-Gómez, M.P.; Holgado, F.; Rodríguez-Alcalá, L.M.; Montero, O.; Fontecha, J. Comprehensive Study of the Lipid Classes of Krill Oil by Fractionation and Identification of Triacylglycerols, Diacylglycerols, and Phospholipid Molecular Species by Using UPLC/QToF-MS. Food Anal. Methods 2015, 8, 2568–2580. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- EFSA Assesses Safety of Long-Chain Omega-3 Fatty Acids. Available online: https://www.efsa.europa.eu/en/press/news/120727 (accessed on 1 March 2021).
- EFSA Commission Regulation (EU) No 432/2012 of 16 May 2012. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?ur (accessed on 1 March 2021).
- Schmid, A.; Walther, B. Natural Vitamin D Content in Animal Products. Adv. Nutr. 2013, 4, 453–462. [Google Scholar] [CrossRef]
- Mattila, P.; Piironen, V.; Uusi-Rauva, E.; Koivistoinen, P. Cholecalciferol and 25-Hydroxycholecalciferol contents in fish and fish products. J. Food Compos. Anal. 1995, 8, 232–243. [Google Scholar] [CrossRef]
- Afonso, C.; Bandarra, N.M.; Nunes, L.; Cardoso, C. Tocopherols in Seafood and Aquaculture Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 128–140. [Google Scholar] [CrossRef]
- Hamre, K.; Berge, R.K.; Lie, Ø. Oxidative stability of Atlantic salmon (Salmo salar, L.) fillet enriched in α-, γ-, and δ-tocopherol through dietary supplementation. Food Chem. 1998, 62, 173–178. [Google Scholar] [CrossRef]
- Petillo, D.; Hultin, H.O.; Krzynowek, J.; Autio, W.R. Kinetics of Antioxidant Loss in Mackerel Light and Dark Muscle. J. Agric. Food Chem. 1998, 46, 4128–4137. [Google Scholar] [CrossRef]
- Abdollahi, M.; Olofsson, E.; Zhang, J.; Alminger, M.; Undeland, I. Minimizing lipid oxidation during pH-shift processing of fish by-products by cross-processing with lingonberry press cake, shrimp shells or brown seaweed. Food Chem. 2020, 327, 127078. [Google Scholar] [CrossRef] [PubMed]
- Andersen, F.; Lygren, B.; Maage, A.; Waagbø, R. Interaction between two dietary levels of iron and two forms of ascorbic acid and the effect on growth, antioxidant status and some non-specific immune parameters in Atlantic salmon (Salmo salar) smolts. Aquaculture 1998, 161, 437–451. [Google Scholar] [CrossRef]
- Niki, E. Synergistic Inhibition of Oxidation by Vitamin E and Vitamin C; CRC Press: Boca Raton, FL, USA, 1988; pp. 111–120. [Google Scholar]
- Ssepuuya, G.; Nakimbugwe, D.; De Winne, A.; Smets, R.; Claes, J.; Van Der Borght, M. Effect of heat processing on the nutrient composition, colour, and volatile odour compounds of the long-horned grasshopper Ruspolia differens serville. Food Res. Int. 2020, 129. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, G.-W.; Yoon, I.S.; Park, S.H.; Park, S.Y.; Kim, J.-S.; Heu, M.S. Preparation and characterization of protein isolate from Yellowfin tuna Thunnus albacares roe by isoelectric solubilization/precipitation process. Fish. Aquat. Sci. 2016, 19, 1–10. [Google Scholar] [CrossRef]
- Abdollahi, M.; Marmon, S.; Chaijan, M.; Undeland, I. Tuning the pH-shift Protein-Isolation Method for Maximum Hemoglobin-removal from Blood Rich Fish Muscle. Food Chem. 2016, 212, 213–224. [Google Scholar] [CrossRef]
- Sannaveerappa, T.; Cai, H.; Richards, M.P.; Undeland, I. Factors affecting the binding of trout HbI and HbIV to washed cod mince model system and their influence on lipid oxidation. Food Chem. 2014, 143, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461–1467. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Moisture (%) | Protein * (% dw) | Fat * (% dw) | Ash * (% dw) | ||
|---|---|---|---|---|---|
| Herring | Backbone | 72.91 ± 0.20 e | 50.82 ± 0.31 e | 34.22 ± 3.07 b | 10.24 ± 0.59 c |
| MSM | 76.58 ± 0.51 d | 58.50 ± 0.74 d | 31.62 ± 3.56 b | 5.12 ± 0.30 d | |
| Protein isolate | 80.54 ± 0.08 ab | 85.66 ± 0.98 a | 7.43 ± 0.70 e | 2.41 ± 0.03 f | |
| Salmon | Backbone | 57.95 ± 0.11 g | 34.60 ± 0.93 g | 52.03 ± 2.96 a | 28.38 ± 0.45 b |
| MSM | 68.02 ± 2.43 f | 46.78 ± 1.5 f | 43.11 ± 1.32 b | 3.70 ± 0.53 e | |
| Protein isolate | 79.69 ± 0.00 b | 72.33 ± 0.52 c | 20.58 ± 0.24 d | 2.18 ± 0.04 f | |
| Cod | Backbone | 77.77 ± 0.30 cd | 60.25 ± 1.60 d | 6.22 ± 0.38 e | 29.69 ± 0.22 a |
| MSM | 82.92 ± 0.55 a | 81.07 ± 2.68 b | 8.85 ± 1.88 e | 5.49 ± 0.30 d | |
| Protein isolate | 81.01 ± 0.20 a | 82.85 ± 0.30 b | 9.29 ± 0.58 f | 2.51 ± 0.28 f |
| Amino Acid (mg/g Protein) | Herring | Salmon | Cod | FAO/WHO Adult (Infant) (mg/g Protein) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | ||
| Valine * | 45.3 ± 1.6 b | 45.4 ± 1.6 b | 50.4 ± 1.8 b | 50.1.1 ± 0.9 b | 50.3 ± 5.6 b | 56.2 ± 1.3 a | 39.9 ± 2.7 c | 45.5 ± 2.2 b | 49.2 ± 1.4 b | 39(55) |
| Threonine * | 45.1 ± 1.2 de | 45.7 ± 1.0 cde | 48.4 ± 1.8 bcd | 52.3 ± 2.2 ab | 51.0 ± 6.5 abc | 55.3 ± 1.0 a | 42.3 ± 3.0 e | 46.2 ± 2.0 cde | 48.5 ± 1.9 bcd | 23(31) |
| Isoleucine * | 41.1 ± 0.8 d | 43.5 ± 1.3 cd | 48.0 ± 1.8 b | 48.7 ± 1.7 b | 49.9 ± 5.5 b | 55.8 ± 0.9 a | 39.4 ± 2.6 d | 46.3 ± 1.8 bc | 49.6 ± 1.8 b | 30(32) |
| Leucine * | 41.1 ± 2.5 e | 50.0 ± 2.7 d | 61.2 ± 1.6 b | 52.1 ± 0.0 cd | 55.1 ± 4.7 c | 67.7 ± 1.4 a | 44.2 ± 2.2 e | 55.4 ± 1.2 c | 64.3 ± 1.2 ab | 59(66) |
| Lysine * | 77.5 ± 0.9 bcd | 73.2 ± 1.9 cd | 77.3 ± 1.8 bcd | 86.0 ± 4.0 ab | 83.7 ± 11.1 ab | 88.4 ± 1.3 a | 68.9 ± 4.6 d | 76.6 ± 3.5 bcd | 79.8 ± 2.5 abc | 45(57) |
| Methionine * | 46.9 ± 1.3 abc | 44.3 ± 0.7 bc | 44.6 ± 1.8 bc | 39.2 ± 7.7 a | 52.3 ± 2.3 ab | 51.2 ± 1.5 a | 42.1 ± 2.9 c | 45.2 ± 2.2 bc | 46.8 ± 2.2 abc | 17(42) |
| Histidine * | 76.5 ± 0.5 ab | 68.0 ± 0.5 bcd | 62.0 ± 3.6 d | 82.4 ± 4.7 a | 76.9 ± 12.3 ab | 72.5 ± 2.03 abc | 63.2 ± 4.6 cd | 64.8 ± 4.2 cd | 62.8 ± 3.1 cd | 15(20) |
| Phenylalanine * | 48.7 ± 2.8 bc | 47.8 ± 0.7 bc | 48.8 ± 2.8 bc | 55.8 ± 2.4 a | 53.4 ± 6.9 ab | 56.4 ± 1.1 a | 44.4 ± 3.3 c | 48.2 ± 2.4 bc | 49.9 ± 1.6 bc | 19(72) |
| Glycine | 51.0 ± 0.2 bc | 44.3 ± 0.4 cde | 42.3 ± 1.8 e | 60.7 ± 1.8 a | 50.4 ± 7.1 bc | 49.5 ± 0.9 bcd | 53.2 ± 5.8 b | 44.4 ± 2.1 cde | 43.3 ± 1.8 de | |
| Alanine | 54.0 ± 0.6 ab | 48.5 ± 1.5 b | 49.9 ± 2.6 b | 58.6 ± 2.0 a | 53.8 ± 6.5 ab | 57.0 ± 1.7 a | 48.9 ± 4.5 b | 49.2 ± 2.1 b | 50.5 ± 1.2 b | |
| Serine | 58.8 ± 0.9 abcd | 54.4 ± 0.3 bcd | 52.0 ± 2.5 d | 65.2 ± 3.4 a | 60.9 ± 9.0 ab | 60.4 ± 1.5 abc | 52.8 ± 4.4 cd | 53.8 ± 2.8 bcd | 54.0 ± 2.2 bcd | |
| Proline | 48.8 ± 1.0 bc | 45.9 ± 1.0 bc | 45.8 ± 2.3 bc | 58.7 ± 1.7 a | 51.1 ± 7.0 bc | 52.0 ± 0.8 c | 49.3 ± 1.7 bc | 44.7 ± 2.2 c | 44.5 ± 2.1 bc | |
| Aspartic acid | 122.5 ± 3.0 d | 107.0 ± 1.1 bc | 98.7 ± 5.3 c | 129.2 ± 7.4 a | 120.7 ± 19.4 ab | 114.6 ± 3.0 abc | 99.5 ± 7.3 c | 102.3 ± 6.5 c | 100.4 ± 4.9 c | |
| Glutamic acid | 118.1 ± 2.8 ab | 104.3 ± 1.0 bcd | 99.3 ± 4.7 cd | 125.3 ± 7.0 a | 117.1 ± 18.4 ab | 113.8 ± 2.5 abc | 96.9 ± 7.0 d | 101.0 ± 5.5 cd | 102.0 ± 3.7 cd | |
| Arginine | 121.1 ± 3.1 ab | 105.9 ± 0.7 bcd | 97.5 ± 5.2 d | 128.5 ± 5.5 a | 120.0 ± 19.1 ab | 113.8 ± 3.0 abc | 99.4 ± 7.4 cd | 101.9 ± 6.4 cd | 99.6 ± 4.4 cd | |
| Tyrosine | 92.3 ± 2.3 ab | 81.3 ± 0.6 bcd | 74.7 ± 4.3 d | 98.0 ± 15.7 a | 91.8 ± 14.7 ab | 87.8 ± 2.5 abc | 75.5 ± 5.4 d | 78.4 ± 4.9 cd | 76.7 ± 4.0 cd | |
| Total EAA | 422.5 ± 8.16 cde | 418 ± 9.5 de | 440.8 ± 18.5 bcd | 479.0 ± 18.4 ab | 470.4 ± 59.6 abc | 503.6 ± 9.2 a | 384.5 ± 26.1 e | 428.3 ± 19.4 cde | 450.9 ± 15.8 bcd | |
| EAA/AA | 0.41 c | 0.41 c | 0.44 a | 0.40 d | 0.41 c | 0.43 a | 0.40 d | 0.42 b | 0.44 a | |
| Fatty Acids | Herring | Salmon | Cod | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | |
| ∑SFA | 79.25 ± 8.74 c (31.0 ± 0.37) | 68.33 ± 7.84 c (29.26 ± 0.66) | 11.56 ± 1.89 g (28.26 ± 2.31) | 219.7 ± 13.0 a (57.2 ± 0.25) | 181.3 ± 4.89 b (55.9 ± 0.09) | 80.79 ± 0.66 c (57.8 ± 0.22) | 16.70 ± 2.83 f (48.4 ± 4.21) | 25.04 ± 1.71 e (45.8 ± 1.69) | 46.30 ± 2.02 d (25.5 ± 0.05) |
| ∑MUFA | 106 ± 18.75 a (41.4 ± 4.26) | 86.51 ± 9.07 a (37.0 ± 0.62) | 7.30 ± 0.53 e (17.91 ± 0.13) | 64.84 ± 4.78 b (16.8 ± 0.17) | 53.32 ± 1.38 b (16.4 ± 0.01) | 19.21 ± 0.12 c (13.7 ± 0.15) | 5.87 ± 0.42 f (17.0 ± 2.52) | 9.08 ± 0.00 d (16.6 ± 0.59) | 59.94 ± 3.06 b (33.0 ± 0.27) |
| ∑PUFA | 60.70 ± 4.69 b (23.8 ± 3.61) | 67.19 ± 3.87 b (28.8 ± 0.87) | 17.47 ± 1.72 d (42.84 ± 0.21) | 84.39 ± 6.64 a (21.9 ± 0.24) | 67.67 ± 1.48 b (20.8 ± 0.13) | 30.01 ± 0.23 c (21.47 ± 0.00) | 8.60 ± 0.30 e (24.8 ± 1.00) | 15.20 ± 2.40 d (27.7 ± 2.96) | 33.78 ± 1.45 c (21.1 ± 0.12) |
| C20:5 n3 (EPA) | 18.06 ± 2.05 a (7.0 ± 0.28) | 15.69 ± 2.11 a (6.7 ± 0.31) | 3.78 ± 0.37 b (9.26 ± 0.18) | 0.28 ± 0.00 d (0.07 ± 0.00) | 0.22 ± 0.00 e (0.07 ± 0.00) | 0.06 ± 0.00 f (0.04 ± 0.00) | 2.27 ± 0.05 c (6.5 ± 0.34) | 3.03 ± 0.62 bc (7.2 ± 0.84) | 3.98 ± 0.30 b (1.31 ± 0.02) |
| C22:5 n3 (DPA) | 0.49 ± 0.37 cd (0.31 ± 0.00) | 0.71 ± 0.09 c (0.30 ± 0.01) | 0.09 ± 0.01 f (0.23 ± 0.02) | 2.19 ± 0.20 a (0.05 ± 0.01) | 1.80 ± 0.00 b (0.05 ± 0.01) | 0.98 ± 0.00 c (0.7 ± 0.00) | 0.16 ± 0.00 e (0.46 ± 0.03) | 0.21 ± 0.02 d (0.48 ± 0.05) | 0.26 ± 0.04 d (0.97 ± 0.06) |
| C22:6 n3 (DHA) | 24.95 ± 1.78 a (9.7 ± 0.02) | 23.01 ± 2.99 a (9.85 ± 0.41) | 11.66 ± 0.85 c (28.6 ± 0.18) | 11.05 ± 0.90 cd (2.8 ± 0.05) | 9.92 ± 0.07 d (3.0 ± 0.10) | 8.89 ± 0.09 e (6.3 ± 0.04) | 5.21 ± 0.18 f (15.0 ± 0.60) | 8.77 ± 1.13 de (16.5 ± 2.36) | 9.07 ± 1.62 cde (0.76 ± 0.05) |
| ∑LC n-3 PUFA | 43.81 ± 3.85 a (17.1 ± 0.24) | 39.42 ± 5.21 a (16.8 ± 0.75) | 15.88 ± 1.27 c (38.1 ± 0.02) | 13.53 ± 1.11 bc (3.5 ± 0.06) | 11.96 ± 0.08 c (3.6 ± 0.11) | 9.94 ± 0.10 d (7.1 ± 0.04) | 7.65 ± 0.23 e (22.1 ± 0.98) | 13.35 ± 2.27 bc (24.3 ± 3.26) | 14.53 ± 0.15 b (3.0 ± 0.14) |
| ∑n-3 PUFA | 47.26 ± 3.34 a (18.0 ± 0.96) | 42.82 ± 5.60 a (18.32 ± 0.24) | 15.98 ± 0.85 c (39.2 ± 0.3) | 17.45 ± 1.42 bc (4.5 ± 0.08) | 15.03 ± 0.06 c (4.6 ± 0.13) | 11.10 ± 0.09 d (7.9 ± 0.03) | 7.74 ± 0.24 e (22.3 ± 0.98) | 13.40 ± 2.24 c (24.51 ± 3.27) | 17.73 ± 0.68 b (9.7 ± 0.13) |
| ∑n-6 PUFA | 19.56 ± 3.30 bc (7.4 ± 1.86) | 22.40 ± 1.00 b (9.62 ± 0.43) | 1.49 ± 0.33 d (3.64 ± 0.52) | 66.63 ± 5.19 a (17.3 ± 0.16) | 52.39 ± 1.34 a (16.17 ± 0.00) | 18.82 ± 0.11 c (13.46 ± 0.03) | 0.74 ± 0.05 e (2.14 ± 0.00) | 1.48 ± 0.02 d (2.72 ± 0.11) | 16.05 ± 0.77 c (8.8 ± 0.05) |
| n-3/n6 ratio | 2.42 ± 0.71 b | 1.90 ± 0.16 b | 10.88 ± 1.64 a | 0.26 ± 0.00 f | 0.28 ± 0.00 f | 0.59 ± 0.00 e | 10.42 ± 0.41 a | 9.03 ± 1.57 a | 1.10 ± 0.00 c |
| Minerals (mg/kg DW) | Herring | Salmon | Cod | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | Backbone | MSM | Protein Isolate | |
| Sodium | 6435 ± 247 ab | 6201 ± 354 ab | 6749 ± 409 a | 2548 ± 209 c | 2648 ± 32 c | 6418 ± 307 ab | 5794 ± 421 b | 6694 ± 183 a | 6501 ± 133 a |
| Calcium | 7818 ± 373 b | 652 ± 32 cd | 165 ± 13 d | 8456 ± 783 b | 586 ± 41 cd | 204 ± 23 d | 9274 ± 528 a | 1251 ± 79 c | 173 ± 16 d |
| Potassium | 12,491 ± 269 b | 11,321 ± 803 bc | 1240 ± 108 f | 4896 ± 508 e | 7299 ± 940 d | 986 ± 18 f | 10,732 ± 700 c | 14,893 ± 231 a | 1277 ± 68 f |
| Selenium | 284 ± 11 a | 274 ± 14 ab | 234 ± 20 b | 253 ± 12 ab | 247 ± 15 ab | 178 ± 22 c | 281 ± 15 ab | 248 ± 27 ab | 194 ± 28 bc |
| Zinc | 31.9 ± 2.5 cd | 22.9 ± 1.8 de | 33.1 ± 5.2 cd | 32.9 ± 3.6 cd | 17.7 ± 2.3 e | 56.8 ± 7.2 a | 43.9 ± 3.1 b | 33.9 ± 5.7 bc | 39.5 ± 4.8 bc |
| Copper | 5.65 ± 0.45 b | 5.05 ± 0.84 bc | 9.85 ± 1.58 a | 5.36 ± 0.25 bc | 2.60 ± 0.82 c | 11.11 ± 1.27 a | 4.64 ± 0.12 bc | 4.51 ± 1.38 bc | 10.81 ± 2.27 a |
| Iron | 45.8 ± 2.1 bc | 52.7 ± 5.7 b | 71.7 ± 7.3 a | 18.2 ± 0.65 d | 21.0 ± 1.64 d | 21.2 ± 2.4 d | 39.9 ± 7.1 c | 43.6 ± 3.5 bc | 42.9 ± 4.9 bc |
| Heme-iron | 31.90 ± 2.02 ab | 33.76 ± 0.23 a | 27.11 ± 0.28 b | 7.03 ± 0.39 e | 9.97 ± 0.17 d | 8.26 ± 0.001 de | 16.60 ± 1.48 c | 31.80 ± 1.60 ab | 29.62 ± 1.15 b |
| Magnesium | 1496 ± 34 b | 1286 ± 82 c | 55 ± 4 f | 927 ± 101 d | 631 ± 21 e | 75 ± 0.9 f | 2246 ± 83 a | 1183 ± 37 c | 70 ± 2 f |
| Manganese | 12.2 ± 1.7 c | 1.9 ± 0.3 d | 5.3 ± 0.7 c | 14.6 ± 1.1 ab | 4.2 ± 0.8 cd | 4.3 ± 0.9 cd | 15.5 ± 0.2 a | 5.9 ± 1.0 c | 5.8 ± 1.9 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdollahi, M.; Wu, H.; Undeland, I. Impact of Processing Technology on Macro- and Micronutrient Profile of Protein-Enriched Products from Fish Backbones. Foods 2021, 10, 950. https://doi.org/10.3390/foods10050950
Abdollahi M, Wu H, Undeland I. Impact of Processing Technology on Macro- and Micronutrient Profile of Protein-Enriched Products from Fish Backbones. Foods. 2021; 10(5):950. https://doi.org/10.3390/foods10050950
Chicago/Turabian StyleAbdollahi, Mehdi, Haizhou Wu, and Ingrid Undeland. 2021. "Impact of Processing Technology on Macro- and Micronutrient Profile of Protein-Enriched Products from Fish Backbones" Foods 10, no. 5: 950. https://doi.org/10.3390/foods10050950
APA StyleAbdollahi, M., Wu, H., & Undeland, I. (2021). Impact of Processing Technology on Macro- and Micronutrient Profile of Protein-Enriched Products from Fish Backbones. Foods, 10(5), 950. https://doi.org/10.3390/foods10050950