Abstract
This study evaluated the anti-obesity effects of lactic acid bacteria. Thirty-one lactic acid bacteria were examined in vitro for their ability to inhibit α-glucosidase activity, lipase activity, and 3T3-L1 cell differentiation. Four selected lactic acid bacteria were administered to obese C57BL/6J mice models for 8 weeks. The degree of improvement in obesity was determined by weight gain and serum biochemical analysis. The expression levels of genes (Fas and Cpt-2) related to obesity in the liver were analyzed by quantitative reverse transcription (qRT)-PCR. In addition, antioxidant protein levels (SOD-2, CAT, and GPx-1) in the liver were evaluated. The lactic acid bacteria-treated groups (PPGK1, LFNK3, LPNK2, and LFNK4) showed lower weight increase rate than the control group. The total cholesterol (T-chol), triglyceride (TG), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) levels in the blood serum of the LFNK4 group were the lowest among other groups, compared to the control group. The expression levels of lipid metabolism-related genes (Fas and Cpt-2) in the liver of the LFNK4 group were lower in Fas and higher in Cpt-2 than in the control group. The antioxidant protein expression levels (SOD-2, CAT, and GPx-1) in the liver tissue were also higher in the LFNK4 group. These results indicate that L. fermentum SMFM2017-NK4 has anti-obesity effects.
1. Introduction
Obesity is a medical condition in which excess fat accumulates in the body, which can adversely affect health [1]. It is closely related to an imbalance between energy intake and expenditure [2]. Obesity has recently been classified as a disease rather than a symptom [3]. Worldwide, more than 300 million people are considered to be overweight, and the current obesity rate in Korea has increased from 31.7% in 2007 to 34.1% in 2017 for adults over 19 years of age [4].
Most of the medicines used to treat obesity have side effects such as increased blood pressure, abdominal pain, anxiety, constipation, insomnia, and headaches [5,6]. Functional foods can substitute for medicines, and several studies are being conducted [7]. Probiotics have been developed as a functional food. They are live microorganisms that exert various beneficial effects on the host [8,9], and the genus Lactobacillus is a representative heterogeneous group of lactic acid bacteria (LAB), which are frequently used as probiotics [10]. Studies have reported that lactic acid bacteria regulate expression of genes involved in energy homeostasis to suppress obesity [11].
The lactic acid bacteria Weissella cibaria WIKIM28 and Weissella onfusa WIKIM51 isolated from kimchi at the Kimchi Research Institute showed anti-obesity effect by inhibiting the differentiation of adipocytes and lipid accumulation, and reducing the expression of adipocyte differentiation-related factors [12,13]. In addition, as fat is deposited in the liver due to obesity, alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels, which are predictors for liver inflammation, increase in blood serum [14]. Kwon et al. (2004) [15] reported that ALT and AST levels decreased after administration of lactic acid bacteria to obese mice, thereby alleviating fatty liver.
The purpose of this study was to develop probiotics with lactic acid bacteria that have anti-obesity effects using obese mouse models.
2. Materials and Methods
2.1. Preparation of Lactic Acid Bacteria
Thirty-one lactic acid bacteria strains were used (Table 1), which were isolated from kimchi in South Korea [16]. One hundred microliters of 20% glycerol stock of each strain was cultured in 10 mL Lactobacilli MRS (de Man, Rogosa and Sharpe) broth (Becton, Dickinson and Company, Franklin lakes, NJ, USA) and incubated anaerobically at 37 °C for 24 h, using an anaerobic pack (AnaerogenTM, Oxoid, Hampshire, UK). Then, 100 μL of the culture medium was transferred into fresh 10 mL Lactobacilli MRS broth (Becton, Dickinson and Company) and incubated at 37 °C for 24 h. The cultured medium was precipitated by centrifuge at 1912× g and 4 °C for 15 min, and the cell pellets were washed twice with phosphate-buffered saline (PBS; pH 7.4; KH2PO4 0.2 g, Na2HPO4 1.5 g, NaCl 8.0 g, KCl 0.2 g/distilled water 1 L). The final cell pellets were suspended in 10 mL of PBS to obtain 9–10 log CFU/mL.

Table 1.
Strains of 31 of lactic acid bacteria used for in vitro test.
2.2. Measurement of α-Glucosidase Inhibition Activity
The inhibitory effect of lactic acid bacteria on α-glucosidase activity was analyzed according to the method described by Lordan et al. (2013) [17]. Ten microliters of 1 U/mL α-glucosidase (Sigma-Aldrich, St. Louis, MO, USA) solution and 50 μL of 100 mM potassium phosphate buffer (pH 6.8) were mixed and 20 μL aliquots of the lactic acid bacteria strains were then added. The mixture was incubated at 37 °C for 15 min and 20 μL of 5 mM p-nitrophenyl α-D-glucopyranoside (Acros Organics, Waltham, MA, USA) was added and mixed well. After reacting at 37 °C for 20 min, 50 μL of 0.1 M Na2CO3 (Duksan Pure Chemicals Co., Ltd., Ansan, Gyeonggido, Korea) was added to stop the reaction. In a blank of the sample, lactic acid bacteria were added without α-glucosidase. Lactic acid bacteria were not added in a control sample (with α-glucosidase) and a blank of the control sample (without α-glucosidase). The reactants were placed into a 96-well plate (SPL Life Sciences, Pocheon, Gyeonggido, Korea) and absorbance was measured three times for each sample at a wavelength of 405 nm using an EpochTM microplate spectrophotometer (BioTek Instruments, Inc., Winooski, VT, USA).
Inhibition activity (%) = {1 − (AbsS − AbsSB)/(AbsC − AbsCB)} × 100
- AbsS: absorbance of a sample
- AbsSB: absorbance of a blank of the sample
- AbsC: absorbance of a control sample
- AbsCB: absorbance of a blank of the control sample
2.3. Measurement of Pancreatic Lipase Inhibition Activity
The inhibitory effect of lactic acid bacteria on pancreatic lipase activity was analyzed according to the method described by Ana and Thomas (2019) [18]. Five hundred microliters of Tris-HCl buffer (Tech&Innovation, Chuncheon, Gangwondo, Korea), adjusted to pH 8.5, and 12.5 μL of 5 mM 4-nitrophenyl octanoate (NPC; Alfa Aesar, Haverhill, MA, USA) were added and mixed with 125 μL of the inoculum. Then, 12.5 μL of pancreatic lipase solution (5 mg/mL) (Sigma-Aldrich) was added and mixed for 2 min without forming bubbles. The mixed solution was incubated at 37 °C for 30 min, and 150 μL was placed into a 96-well plate (SPL Life Sciences). In a blank of the sample, lactic acid bacteria were added with dimethyl sulfoxide (DMSO, Sigma-Aldrich) instead of NPC. Lactic acid bacteria were not added in a control sample (with NPC) and a blank of the control sample (with DMSO instead of NPC). The absorbance was measured three times for each sample at a wavelength of 412 nm using an EpochTM Microplate Spectrophotometer (BioTek Instruments, Inc.), and the inhibition activity was calculated as follows:
Inhibition activity (%) = 100 − {(AbsS − AbsSB)/(AbsRC − AbsB) × 100}
- AbsS: absorbance of a sample
- AbsSB: absorbance of a blank of the sample
- AbsRC: absorbance of a control sample
- AbsB: absorbance of a blank of the control sample
2.4. Measurement of Inhibition for 3T3-L1 Adipocyte Differentiation
3T3-L1 preadipocyte cells were purchased from the Korean Cell Line Bank (KCLB; Seoul, Korea). The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone, Logan, UT, USA) supplemented with 10% newborn calf serum (NBCS; Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and 1% penicillin–streptomycin solution (PS; Oxoid, Basingstoke, Hampshire, UK) in a 72T flask (Corning Inc., New York, NY, USA) and incubated at 37 °C and 5% CO2 for 2 days. Cultured cells were then sub-cultured in fresh medium for another 2 days and washed with Dulbecco’s phosphate-buffered saline (DPBS; Welgene, Gyeongsan, Gyeongsangbukdo, Korea). The cells were then detached with 0.05% trypsin-0.02% EDTA (Gibco). Then, the cells were centrifuged at 1000 rpm and 25°C for 5 min and mixed with the medium to be used in the experiment. 3T3-L1 cells were dispensed into 12-well plates (SPL Life Sciences) at a concentration of 2 × 104 cells/mL, and cultured at 37 °C with 5% CO2. 3T3-L1 cells were cultured in MDI medium (DMEM; Hyclone), 10% fetal bovine serum (FBS; Gibco), 1% penicillin–streptomycin solution (PS; Oxoid), and 5 μg/mL insulin for 2 days to differentiate into adipocytes. The medium was then replaced with fresh growth media (DMEM, 10% FBS, and 1% PS) containing the lactic acid bacteria strains and refed every 2 days for four times to complete differentiation. The differentiated cell cultures were washed three times with DPBS (Welgene) and treated with 10% formalin for 1 h to fix the cells. The differentiated cells with spherical shape were stained with oil red O (Sigma-Aldrich) solution for 30 min in the dark and washed twice with distilled water. The cells were dissolved in isopropyl alcohol (Duksan Pure Chemicals Co., Ltd.) and absorbance was measured three times at a wavelength of 520 nm using an EpochTM Microplate Spectrophotometer (BioTek Instruments, Inc.). The inhibitory activity was calculated using the following equation:
The control optical density (O.D.) is the absorbance of the test solution with PBS instead of the bacterial solution, and sample O.D. shows the absorbance of the test solution with bacterial solution.
2.5. Development of Obese Mouse Model and Administration of Lactic Acid Bacteria
Of 31 lactic acid bacteria, 10 strains were ranked for high α-glucosidase inhibition activity and pancreatic lipase inhibition activity, and 4 strains commonly ranked for both enzyme inhibition activities were selected. The strains were identified as Pediococcus pentosaceus SMFM2017-GK1, Lactobacillus fermentum SMFM2017-NK3, Lactobacillus plantarum SMFM2017-NK2, and Lactobacillus fermentum SMFM2017-NK4. For animal experiments, 48 male SPF C57BL/6J mice 4 weeks old were purchased from Raonbio Inc. (Yongin, Gyeonggido, Korea). The mice were adapted for a week at 18–26 °C and 40–60% humidity. Then, the mice were randomly divided into 6 groups of 8 mice each (normal (normal mice treated with PBS), control (obesity-induced mice treated with PBS), PPGK1 (obesity-induced mice treated with P. pentosaceus SMFM2017-GK1), LFNK3 (obesity-induced mice treated with L. fermentum SMFM2017-NK3), LPNK2 (obesity-induced mice treated with L. plantarum SMFM2017-NK2), and LFNK4 (obesity-induced mice treated with L. fermentum SMFM2017-NK4) groups). Obesity in mice was induced by supplying a 60% high-fat diet (Research Diets, Inc., New Brunswick, NJ, USA) for 6 weeks to the control, PPGK1, LFNK3, LPNK2, and LFNK4 groups. The normal group was fed a normal diet (Research Diets, Inc.). The body weight of the mice was measured every 3 days. Two hundred microliters of lactic acid bacteria were administered orally to the obese mice in PPGK1, LFNK3, LPNK2, and LFNK4 groups, and 200 μL PBS was administered to the control and normal groups. The weight gain was calculated by comparing the difference between the weight after induction of obesity at 8 weeks and the weight after oral administration at 14 weeks. After 8 weeks of treatment, all the mice were sacrificed using CO2 gas. The animal experiments were performed after approval by the Institutional Animal Care and Use Committee of Sookmyung Women’s University (approval number: SMWU-IACUC-1912-027).
2.6. Analysis of Serum Cholesterol and Lipid
Blood was collected from the posterior aorta of the mice after sacrifice and placed in a serum separation tube (Microtainer; Becton, Dickinson and Company) for 30 min. Serum was obtained by centrifugation of the blood at 4 °C and 5000 rpm for 10 min and stored at −70 °C. Blood serum was analyzed using a Mindray BS-220 biochemistry analyzer (Mindray Medical International Ltd., Shenzhen, China).
2.7. Pathological Analysis of Adipose and Liver Tissue
The epididymal fat and the liver were fixed with 10% formalin (Hisko, Gunpo, Gyeonggido, Korea), and a paraffin block was prepared to cut 3–4 μM sections. The adipose tissues of the epididymal fat and the liver tissues were observed using a microscope (Leica, Wetzlar, Germany) after hematoxylin and eosin (H&E) staining, and the size and area of adipocytes and steatosis of hepatocytes were analyzed using the ImageJ program (National Institutes of Health, Bethesda, MD, USA).
2.8. Transcriptome Analysis of the Liver Tissue
To investigate the mRNA expression in the liver tissues, liver mRNA was extracted using an RNeasy mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Complementary DNA (cDNA) was synthesized using the extracted mRNA, according to the manufacturer’s instructions, using a QuantiTect Reverse Transcription kit (Qiagen). cDNA was then used to evaluate the gene expression by quantitative reverse transcription PCR (qRT-PCR) and a Rotor-Gene SYBR® Green PCR kit (Qiagen), and qRT-PCR was performed using a Rotor-Gene Q (Qiagen). In brief, 25 μL reaction mixture was used comprising 1 μL template cDNA, 12.5 μL 2 × rotor-gene SYBR® green PCR master mix, 6.5 μL RNase-free water, 2.5 μL forward primer (10 pmol), and 2.5 μL reverse primer (10 pmol). The amplification conditions were as follows: 5 min preheating at 95 °C, followed by 35 cycles of 5 s denaturation at 95 °C, and 10 s annealing at 60 °C. Gene expression levels of β-actin, Fas, and Cpt-2 were analyzed using specific primers for each gene (Table 2), and the relative gene expression was calculated using the 2−ΔΔCt method [19].
Table 2.
Primer sequences of obesity related to genes used for qRT-PCR.
2.9. Immunoblotting
To analyze protein expression in the liver, protein was extracted from 30 to 50 mg of liver using a PRO-PREPTM protein extraction solution (iNtRON Biotechnology Inc., Seongnam, Gyeonggido, Korea) according to the manufacturer’s instructions. The concentration of the extracted protein was quantified using a DCTM Protein Assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Protein expression was quantified by Western blotting as follows: 20 μg of protein was separated by 12% sodium dodecyl sulfate (SDS)-PAGE at 200 V for 30 min. The proteins were then transferred to a polyvinylidene difluoride (PVDF) membrane (GE Healthcare Life Sciences, Marlborough, MA, USA) at 60 V for 2.5 h. Membranes were incubated with 5% skim milk (Sigma-Aldrich) at room temperature (20 °C) for 1 h. The primary antibodies (mouse-anti-SOD-2 (sc-130345, 1:100), mouse-anti-CAT (sc-271803, 1:1000), and GPx-1 (sc-133160, 1:500)) (Santa Cruz Biotechnology Inc., Santa Cruz, TX, USA) were used for immunoblotting, and mouse-anti-β-actin (sc-81178, 1:1000, Santa Cruz Biotechnology Inc.) was used as a reference protein. Anti-mouse IgG HRP (sc-2005, 1:5000, Santa Cruz Biotechnologies, Inc., Dallas, TX, USA) was used as the secondary antibody. The immunoreactive bands were visualized with an LAS-3000 Imager (Fujifilm, Tokyo, Japan), and the intensity of the bands was quantified using GelQuant software v.2.7. (DNR Imaging Systems Ltd., Jerusalem, Israel).
2.10. Statistical Analysis
SAS version 9.4 (SAS Institute Inc., Cary, NC, USA) was used for the statistical analysis. Significant differences were determined between the two groups (normal and lactic acid bacteria-treated groups) using a general linear model (GLM) with a pairwise t-test at α = 0.05. The least-squares (LS) means were compared to analyze the differences.
3. Results/Discussion
3.1. Inhibition of α-Glucosidase Activity by Lactic Acid Bacteria
α-Glucosidase is an enzyme that catalyzes the last step in the digestion of carbohydrates, converting them to glucose [23,24]. α-Glucosidase inhibitors in the small intestine inhibit the process of disaccharide-to-monosaccharide degradation [25]. The activity of α-glucosidase inhibitors delays digestion and absorption of sugars in the intestine [26]. To select lactic acid bacteria that have anti-obesity effects, we examined whether they inhibit α-glucosidase activity. Table 3 shows the results of the inhibition of α-glucosidase activity by the 31 lactic acid bacteria strains. Thirty-one lactic acid bacteria strains showed 6.58–41.45% inhibitory activity against α-glucosidase. The top 10 strains with the highest α-glucosidase inhibitory activity were LAB 66 (41.45%), 29 (40.64%), 50 (39.13%), 70 (39.09%), 75 (37.22%), 45 (37.04%), 44 (34.64%), 73 (33.11%), 20 (30.68%), and 98-1-3 (30.16%). Among the 31 lactic acid bacteria, strain 66 showed the highest (41.45%) α-glucosidase inhibitory activity. EI-shiekh et al. (2019) [27] showed that the activities of various plant extracts including Solenostemma argel were 37.29%–89.08%. Kim and Lim (2020) [28] reported that α-glucosidase inhibitory activity of the milk fermented with L. plantarum KI134 strain was 2.74%, which was the highest among other 255 strains. Compared to these activities, the α-glucosidase inhibitory activity in our study presented above can be considered similar or high. Thus, it considered that the lactic acid bacteria strains may have an effect on obesity. Hence, further analysis was performed.
Table 3.
Inhibition ability of lactic acid bacteria for α-glucosidase activity.
3.2. Inhibition of Pancreatic Lipase Activity by Lactic Acid Bacteria
Pancreatic lipase is an enzyme that hydrolyzes fat triacylglycerol into 2-monoacylglycerols and fatty acids [29]. Orlistat, a representative inhibitor of pancreatic lipase, has excellent efficacy and inhibits approximately 30% hydrolysis of the ingested fat; however, orlistat has side effects such as gastrointestinal disorders, irritability, bile secretion disorders, and inhibition of absorption of fat-soluble vitamins [30]. Recently, studies have been conducted to develop inhibitors derived from natural products without side effects [31]. Based on these findings, we examined the potential of 31 lactic acid bacteria to inhibit obesity by investigating their ability to inhibit pancreatic lipase activity. The results are presented in Table 4. The 31 lactic acid bacteria strains showed 10.58%–18.92% inhibitory activity against pancreatic lipase, and the top 10 lactic acid bacteria strains with the highest pancreatic lipase inhibitory activity were LAB 20 (18.92%), 11 (18.80%), 26 (18.10%), 30 (15.22%), 29 (14.43%), 73 (13.74%), 72 (10.96%), 50 (10.95%), 38-Y (10.39%), and 31 (10.15%). In the studies by Kang et al. (2018) [32], the lipase inhibitory activity of Lactobacillus fermentum MG4231 was 12.24%, similar to the results of this study, and it has been shown to have a good inhibitory effect on adipogenic differentiation. The results of the inhibition of α-glucosidase activity and pancreatic lipase activity by the lactic acid bacteria showed that LAB 20, 29, 50, and 73 strains had excellent inhibitory activities.
Table 4.
Inhibition ability of lactic acid bacteria for pancreatic lipase activity.
3.3. Inhibition of 3T3-L1 Cell Differentiation by Lactic Acid Bacteria
Triglycerides are accumulated during differentiation of 3T3-L1 pre-adipocyte in cells by various transcription factors such as peroxisome proliferator-activated receptor γ (PPARγ), CCAAT-enhancer binding protein α (C/EBPα), and hormones [33]. Several studies analyzed the effect of obesity prevention through the inhibition of 3T3-L1 cell differentiation. Kim et al. (2010) [34] evaluated the effect of soy milk fermented with Bacillus subtilis KC-3 on the inhibition of 3T3-L1 cell differentiation and found that the fermented soy milk reduced 19% of fat accumulation, and Samjeonghwan fermented with lactic acid bacteria reduced the number of adipocytes [35]. In a study by Kim and Lim (2017) [36], L. plantarum K6 was selected as a lactic acid bacterium that has an anti-obesity effect; it inhibited differentiation of 3T3-L1 by 27.4%. These are similar to the highest inhibitory value of 21.44% (strain LAB 73) obtained in our study (Table 5). The strains LAB 29, 20, 50, and 73 were identified and named as L. fermentum SMFM2017-NK3, P. pentosaceus SMFM2017-GK1, L. plantarum SMFM2017-NK2, and L. fermentum SMFM2017-NK4, respectively (Table 5).
Table 5.
Inhibition ability of lactic acid bacteria (LAB) strains (L. fermentum SMFM2017-NK4, L. fermentum SMFM2017-NK3, P. pentosaceus SMFM2017-GK1, and L. plantarum SMFM2017-NK2) for 3T3-L1 pre-adipocyte cell differentiation.
3.4. Weight Gain of the Obese Mouse Model
Obesity was induced in mice by a 60% high-fat diet for 6 weeks, and the weight of the control group (38.32 g) was significantly higher than that of the normal group (25.81 g) (p < 0.05). After administration of lactic acid bacteria in each group (P. pentosaceus SMFM2017-GK1 (PPGK1), L. fermentum SMFM2017-NK3 (LFNK3), L. plantarum SMFM2017-NK2 (LPNK2), and L. fermentum SMFM2017-NK4 (LFNK4)), the weight gain, which is the difference between the maximum body weight after induction of obesity and the minimum body weight after administration of lactic acid bacteria, in all four groups (PPGK1 (9.51 g), LFNK3 (9.01 g), LPNK2 (9.35 g), and LFNK4 (9.28 g)) were similar (Figure 1). These weight gains were lower by 2.0 g (PPGK1), 2.5 g (LFNK3), 2.1 g (LPNK2), and 2.2 g (LFNK4), respectively, compared to the control group (p < 0.05). In a study by Park et al. (2018) [37], the obese C57BL/6J mice which were treated with lactic acid bacteria (combination of Lactobacillus casei WK3, Enterococcus faecium WK5, Bifidobacterium longum WK9, and L. plantarum K-1) for 6 weeks had lower weight gain of 0.4 g, compared to the control group. In addition, fat accumulation in epididymal adipose tissue and symptoms of fatty liver were found to be alleviated [37]. Thus, lower weight gains of 2.0–2.5 g would be effective against obesity, and lactic acid bacteria (P. pentosaceus SMFM2017-GK1 (PPGK1), L. fermentum SMFM2017-NK3 (LFNK3), L. plantarum SMFM2017-NK2 (LPNK2), and L. fermentum SMFM2017-NK4 (LFNK4)) might reduce fat accumulation.
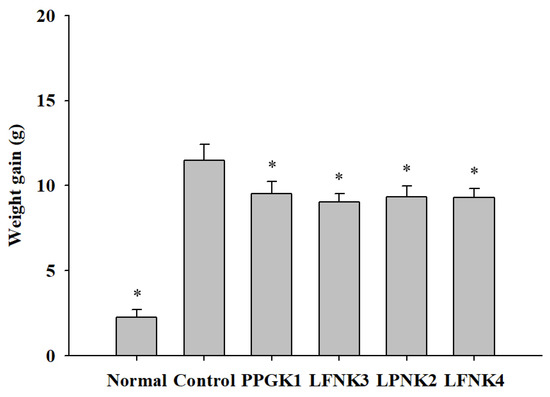
Figure 1.
Weight gain (g) in mice after treatment of lactic acid bacteria for 8 weeks. * Statistically significant, compared to the control group by pairwise t-test (p < 0.05).
Data are presented as mean and standard error. Normal: normal diet + phosphate-buffered saline (PBS); Control: 60% high fat diet (HFD) + PBS; PPGK1: HFD + P. pentosaceus SMFM2017-GK1; LFNK3: HFD + L. fermentum SMFM2017-NK3; LPNK2: HFD + L. plantarum SMFM2017-NK2; LFNK4: HFD + L. fermentum SMFM2017-NK4.
3.5. Indicators of Lipid and Inflammation-Related Markers in Blood Serum
After treatment of the obese mice with lactic acid bacteria for 8 weeks, the concentrations of serum total cholesterol (T-chol) were 116.6, 264.8, 227.0, 258.8 and 204.0 mg/dL in the normal, control, PPGK1, LPNK2, and LFNK4 groups, respectively (Figure 2a). The concentrations of triglycerides (TG) were 65.6 and 115.0 mg/dL in normal and control groups, respectively. After treatment with lactic acid bacteria, the concentrations of TG in the PPGK1 (87.2 mg/dL), LFNK3 (81.0 mg/dL), LPNK2 (79.7 mg/dL) and LFNK4 (72.2 mg/dL) groups were lower (p < 0.05) than that in the control group (115.0 mg/dL), and the level in the LFNK4 group was the lowest (p < 0.05) (Figure 2b). HDL-cholesterol (107.0 mg/dL) and LDL-cholesterol (6.8 mg/dL) levels in the normal group were lower (p < 0.05) than those of the control group (HDL-cholesterol: 190.3 mg/dL, LDL-cholesterol: 29.1 mg/dL). After treatment of lactic acid bacteria, HDL-cholesterol levels were 181.9, 203.7, 191.7 and 167.2 mg/dL in PPGK1, LFNK3, LPNK2 and LFNK4 (Figure 2c). LDL-cholesterol levels were 24.4, 28.6, 25.4 and 21.8 mg/dL in PPGK1, LFNK3, LPNK2 and LFNK4 groups, and the level in the LFNK4 group was the lowest among the groups administered lactic acid bacteria (Figure 2d). For ALT and AST, the levels were 41.2 U/L and 73.1 U/L in the normal group, which were lower (p < 0.05) than those in the control group (ALT: 171.0 U/L, AST: 161.5 U/L). The ALT levels were 147.9, 148.4, 144.9 and 102.3 U/L, and AST levels were 149.5, 154.2 U/L, 153.0 and 118.3 U/L in PPGK1, LFNK3, LPNK2 and LFNK4 groups, respectively, after lactic acid bacteria treatment (Figure 2d,e). These levels in the LFNK4 group were the lowest among the groups administered lactic acid bacteria. Based on these results, the L. fermentum SMFM2017-NK4 strain, which was administered to the LFNK4 group, is the most effective in reducing blood serum lipids, especially TG and alleviating inflammation-related markers among the four lactic acid bacteria.
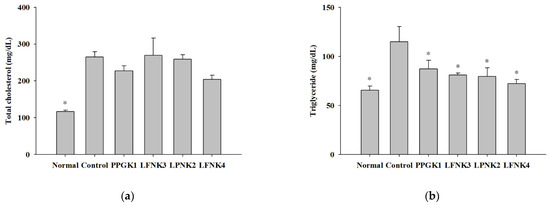
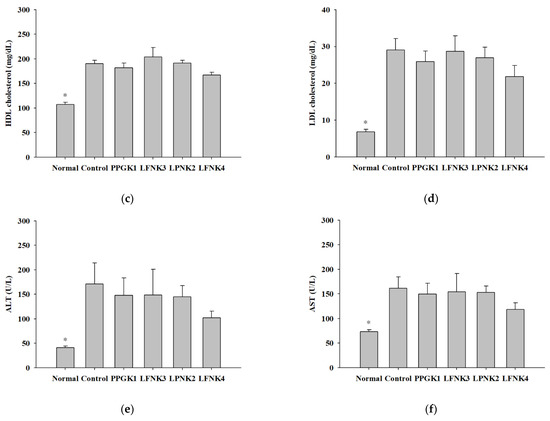
Figure 2.
Serum level of total cholesterol (T-chol) (a), triglyceride (TG) (b), HDL-cholesterol (c), LDL-cholesterol (d), alanine aminotransferase (ALT) (e), and aspartate aminotransferase (AST) (f) in serum after the treatment of lactic acid bacteria for 8 weeks. Data are presented as mean and standard error. Normal: normal diet + phosphate-buffered saline (PBS); Control: 60% high-fat diet (HFD) + PBS; PPGK1: HFD + P. pentosaceus SMFM2017-GK1; LFNK3: HFD + L. fermentum SMFM2017-NK3; LPNK2: HFD + L. plantarum SMFM2017-NK2; LFNK4: HFD + L. fermentum SMFM2017-NK4. * Statistically significant, compared to the control group by pairwise t-test (p < 0.05).
3.6. Pathological Features of Adipose and Liver Tissue
In general, the size of epididymal adipose tissue fat cells increases significantly when obesity is induced by the intake of a high-fat diet [38]. Ryu et al. (2011) [39] reported that the high-fat diet group increased the size of adipocytes compared to the normal group, and the group fed with white beech mushroom (Hypsizigus marmoreus) powder decreased the size of adipocytes, thereby inhibiting fat accumulation in adipose tissue. In our study, the adipocytes of the mice in the control group, which were administered a high-fat diet, were larger and irregularly shaped, compared to those of the normal group in the same ×40 magnification (Figure 3). In the groups administered lactic acid bacteria, the size and shape of adipocytes were similar to those in the normal group (Figure 3). The average area of adipocytes increased by 189.7% in the control group compared to the normal group (p < 0.05), and it decreased in the group administered with lactic acid bacteria, compared to the control group. After treatment with lactic acid bacteria, the average area of adipocytes in the LFNK4 group (135.7%) was significantly decreased (p < 0.05) compared to the control group, followed by LFNK3 (137.1%), PPGK1 (162.3%) and LPNK2 (217.4%) groups (Figure 4). The average area of adipocytes in the LFNK4 group decreased the most by 154.0% compared to that of the control group. In a study by Song et al. (2016) [40], the average area of adipocytes increased significantly by 83.2% in the control group compared to the normal group and treatment with lactic acid bacteria resulted in a 29.0% decrease in the average area of adipocytes, compared to the control group [40]. Thus, it was confirmed that the L. fermentum SMFM2017-NK4 (LFNK4) was effective in decreasing fat synthesis and the size of fat cells.

Figure 3.
Representative microscopic images (×40) on epididymal white adipose tissues from mice groups. Scale bar, 50 μm. (a) Normal: normal diet + phosphate-buffered saline (PBS), (b) Control: 60% high-fat diet (HFD) + PBS, (c) PPGK1: HFD + P. pentosaceus SMFM2017-GK1, (d) LFNK3: HFD + L. fermentum SMFM2017-NK3, (e) LPNK2: HFD + L. plantarum SMFM2017-NK2, (f) LFNK4: HFD + L. fermentum SMFM2017-NK4.
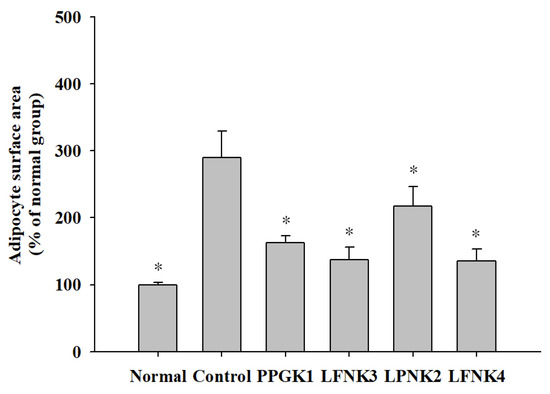
Figure 4.
Epididymal adipocyte surface area. Data are presented as mean and standard error. Normal: normal diet + phosphate-buffered saline (PBS); Control: 60% high-fat diet (HFD) + PBS; PPGK1: HFD + P. pentosaceus SMFM2017-GK1; LFNK3: HFD + L. fermentum SMFM2017-NK3; LPNK2: HFD + L. plantarum SMFM2017-NK2; LFNK4: HFD + L. fermentum SMFM2017-NK4. * Statistically significant, compared to the control group by pairwise t-test (p < 0.05).
In the liver tissue of obese mice, severe steatosis was observed according to the International Harmonization of Nomenclature and Diagnostic Criteria (INHAND) [41]. In a study by Yu et al. (2019) [42], the livers of mice fed a normal diet were normal, but fat particles were observed on the surface of the liver of mice fed a 60% high-fat diet for 14 weeks. In a study by Park et al. (2015) [43], fat deposition in the liver tissue was reduced in the group administered with lactic acid bacteria-fermented Dioscoreae Rhizoma powder, compared to the control group. In a study by Lee et al. (2017) [44], a large amount of fat was accumulated in the liver in the control group that was fed a high-fat diet, and liver fat in the group administered ethanol extract of Aster yomena was reduced. In our study, severe steatosis was observed in the control group, which had fat accumulation in the liver. After administration of the lactic acid bacteria, fewer lesions were observed in the LFNK3 and LFNK4 groups; however, many steatosis lesions were observed in the PPGK1 and LPNK2 groups (Figure 5) (Table 6). The degree of steatosis in the liver was severe in the control, PPGK1, and LPNK2 groups, and it was moderate in the LFNK3 and LFNK4 groups (Table 6). Our results showed that administration of lactic acid bacteria, especially L. fermentum SMFM2017-NK3 (LFNK3) and L. fermentum SMFM2017-NK4 (LFNK4), reduced fat accumulation in the liver.
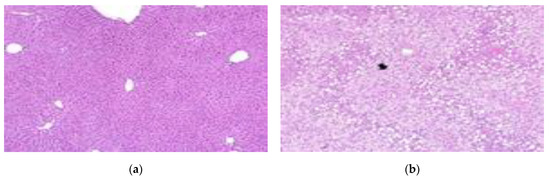
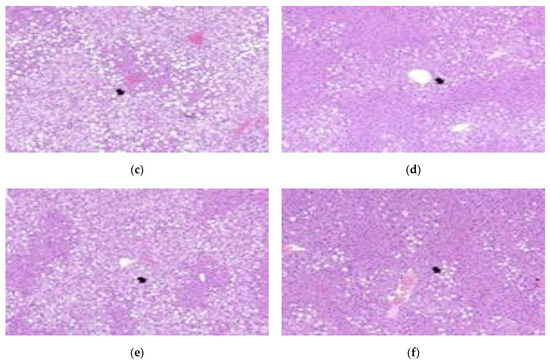
Figure 5.
Representative microscopic images on liver tissues from mice groups (×100). (a) Normal: normal diet + phosphate-buffered saline (PBS), (b) Control: 60% high-fat diet (HFD) + PBS, (c) PPGK1: HFD + P. pentosaceus SMFM2017-GK1, (d) LFNK3: HFD + L. fermentum SMFM2017-NK3, (e) LPNK2: HFD + L. plantarum SMFM2017-NK2, (f) LFNK4: HFD + L. fermentum SMFM2017-NK4.
Table 6.
Histopathological scores in liver tissue from mice groups.
3.7. Transcriptome in the Liver Tissue
Fas is an enzyme involved in the synthesis of fatty acids from acetyl-CoA and malonyl-CoA [45]. Fatty acid synthesis decreases if the expression levels of Fas are suppressed, which reduces accumulation [46]. In Ikarashi et al. (2011) [47], the level of Fas gene expression in the liver decreased in the group administered with acacia polyphenol compared to the high-fat diet group. In this study, the expression levels of Fas were significantly lower (p < 0.05) in the LFNK4 group than in the other lactic acid bacteria-treated groups, compared to the control group (Figure 6a). In detail, the Fas expression level in the control group increased by 1.9 times, compared to that of the normal group; however, the level in the LFNK4 group increased by 0.6 times, compared to the normal group (Figure 6a). In the case of a study by Kim et al. (2019) [48], the expression level of Fas gene in the liver increased by 1.7 times in the high-fat diet group, compared to that of the normal group. The level decreased by 1.3 times in the group administered with the mixture of Atractylodes macrocephala and Amomum villosum extracts, compared to the normal group. Thus, it is considered that L. fermentum SMFM2017-NK4 (LFNK4) was more effective on lowering Fas gene expression than the mixture of Kim et al. (2019) [48]. Cpt-2 is required for the synthesis of carnitine palmitoyl transferase 2, which is an essential enzyme for fatty acid oxidation [49]. Fatty acids must move from cells to mitochondria to be subjected to degradation by β-oxidation, and Cpt-2 is an enzyme that catalyzes this process [50]. Therefore, the oxidation rate of fatty acids increases when the expression of Cpt-2 increases and body fat decreases [51]. In our study, the expression levels of Cpt-2 appeared the highest in the LFNK4 group among the lactic acid bacteria-treated groups, compared to the control group, although there was no statistically significant difference (Figure 6b). Therefore, L. fermentum SMFM2017-NK4 (LFNK4) was effective on regulating gene expressions in the liver, especially Fas gene.
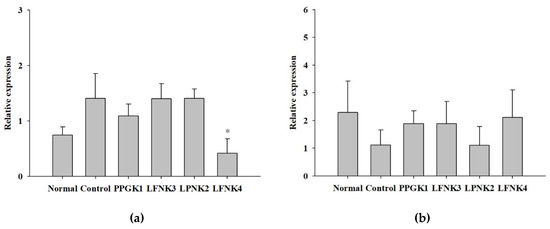
Figure 6.
Relative expression levels of Fas (a) and Cpt-2 (b) in liver tissues. Data are presented as mean and standard error. Normal: normal diet + phosphate-buffered saline (PBS); control: 60% high-fat diet (HFD) + PBS; PPGK1: HFD + P. pentosaceus SMFM2017-GK1; LFNK3: HFD + L. fermentum SMFM2017-NK3; LPNK2: HFD + L. plantarum SMFM2017-NK2; LFNK4: HFD + L. fermentum SMFM2017-NK4. * Statistically significant, compared to the control group by pairwise t-test (p < 0.05).
3.8. Protein Expression Related to Antioxidant Enzymes in the Liver Tissue
Based on the results of the in vivo experiments presented above, the LFNK4 bacteria were the most effective in improving obesity among the other lactic acid bacteria. Thus, the protein expression levels in the liver tissue of the mice of the various groups were further analyzed. There are many studies on antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx). Obesity affects the activity of these enzymes resulting in abnormalities in the antioxidant defense mechanism and an increase in free radicals in the liver. For this reason, studies on the antioxidant system in the liver of obese animal models are being conducted. The expression levels of antioxidant-related proteins (SOD-2, CAT, and GPx-1) in the liver tissues were analyzed (Figure 7). The expression levels of SOD-2 and CAT appeared higher in the LFNK4 group than in the control group, although there was no statistically significant difference (Figure 7a,b), and the expression levels of GPx-1 were significantly increased (p < 0.05) in the LFNK4 group compared to the control group (Figure 7c). According to a study by Kim et al. (2019) [52], the expression levels of the antioxidant proteins SOD, CAT, and GPx were increased after treatment with Banggihwanggitang-hap-yeonggyechulgam-tang, a medicinal herb used to treat obesity, in an obese mouse model, compared to the group that did not receive treatment. Our study also showed that the expression levels of the antioxidant proteins, especially GPx-1, increased in the LFNK4 group compared to that in the control group. Therefore, L. fermentum SMFM2017-NK4 (LFNK4) was found to be effective at reducing obesity-induced oxidative stress in the liver.
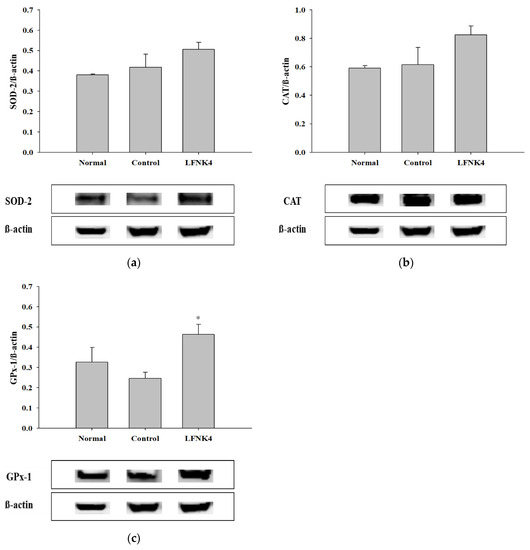
Figure 7.
The immunoreactive intensity of SOD-2 (a), CAT (b), and GPx-1 (c) in liver tissues. Data are presented as mean and standard error. Normal: normal diet + phosphate-buffered saline (PBS); Control: 60% high-fat diet (HFD) + PBS; LFNK4: HFD + L. fermentum SMFM2017-NK4. * Statistically significant, compared to the control group by pairwise t-test (p < 0.05).
As described in Figure 8, L. fermentum SMFM2017-NK4 downregulated the expression of the Fas gene, which resulted in decreased fat synthesis in the liver. Furthermore, it upregulated the protein expression of GPx-1, increasing the antioxidant capacity that was reduced by obesity. As a result, it affected lowering the concentration of TG in the blood serum, adipocyte surface area, and steatosis in the liver; thus, the weight gain decreased.
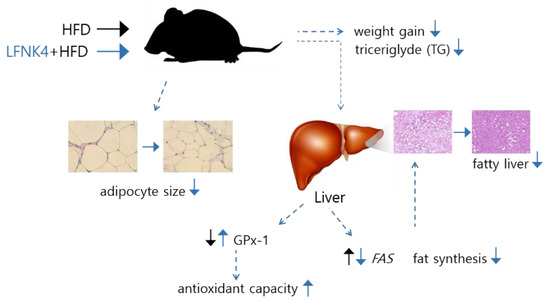
Figure 8.
Effect of L. fermentum SMFM2017-NK4 administration on the improvement of fatty liver and lipid accumulation. A high-fat diet (HFD) causes fatty liver by increasing the expression of Fas gene in the liver. In addition, HFD decreases antioxidant capacity by decreasing the expression of GPx-1 protein in the liver. After administration of L. fermentum SMFM2017-NK4, fat synthesis was inhibited by decreasing Fas expression in the liver. In addition, antioxidant capacity was improved by increasing GPx-1 expression in the liver. L. fermentum SMFM2017-NK4 eventually inhibited the development of fatty liver and lipid accumulation.
4. Conclusions
In conclusion, L. fermentum SMFM2017-NK4, which had inhibition ability for α-glucosidase and pancreatic lipase activity, and 3T3-L1 pre-adipocyte cell differentiation, decreased TG in the blood serum, the size of adipocytes, and the steatosis of the liver. It was affected by downregulating expression of the Fas and upregulating GPx-1. Thus, weight gain was eventually decreased. Therefore, L. fermentum SMFM2017-NK4 can alleviate obesity.
Author Contributions
Conceptualization, Y.Y. (Yohan Yoon) and Y.C.; methodology, D.K. and Y.C.; validation, S.K., J.H. and H.O.; formal analysis, D.K.; investigation, Y.L., Y.K. and Y.S.; resources, J.K., E.P. and Y.Y. (Yoonjeong Yoo); data curation, D.K.; writing—original draft preparation, D.K.; writing—review and editing, Y.C. and Y.Y (Yohan Yoon).; supervision, S.L. and H.L.; project administration, Y.Y. (Yohan Yoon). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Animal Care and Use Committee of Sookmyung Women’s University (approval number: SMWU-IACUC-1912-027, date of approval: 2020.7.31).
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, S.K.; Lim, S.D. Separation and purification of lipase inhibitory peptide from fermented milk by Lactobacillus plantarum Q180. Food Sci. Anim. Resour. 2020, 40, 87–95. [Google Scholar] [CrossRef]
- Lee, J.S.; Hyun, I.K.; Yoon, J.W.; Seo, H.J.; Kang, S.S. Bioconversion products of whey by lactic acid bacteria exert anti-adipogenic effect. Food Sci. Anim. Resour. 2021, 41, 145–152. [Google Scholar] [CrossRef] [PubMed]
- McGee, D.L. Body mass index and mortality: A meta analysis based on person-level data from twenty-six observational studies. Ann. Epidemiol. 2005, 15, 87–97. [Google Scholar] [CrossRef] [PubMed]
- KCDC (Korea Centers for Disease Control and Prevention). Trends in Prevalence of Obesity among Korean Adults 2007–2016. 2018. Available online: http://www.cdc.go.kr/board.es?mid=a20602010000&bid=0034&act=view&list_no=140978 (accessed on 29 June 2020).
- Lee, K.S.; Kim, M.G.; Lee, N.Y. Antimicrobial effect of the extracts of Cactus Cheonnyuncho (Opuntiahumifusa) against food borne pathogens. J. Korean Soc. Food. Nutr. 2004, 33, 1268–1272. [Google Scholar]
- Lee, Y.A.; Kim, M.H. The effect of sea tangle extract on serum lipid level in ovariectomized rats. J. Life Sci. 2008, 18, 249–254. [Google Scholar] [CrossRef]
- Lee, S.J.; Park, H.J.; Song, Y.O.; Jang, S.H.; Goo, J.; Ko, Y.G.; Cho, J.H. Antioxidant activity and anti-obesity effect of Coprinus comatus in Zucker rat (fa/fa). Korean J. Vet. Serv. 2014, 37, 51–58. [Google Scholar] [CrossRef][Green Version]
- De Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Wulandari, E.; Yurmiati, H.; Subroto, T.; Suradi, K. Quality and probiotic lactic acid bacteria diversity of rabbit meat Bekasam-fermented meat. Food Sci. Anim. Resour. 2020, 40, 362–376. [Google Scholar] [CrossRef]
- Kariyawasam, K.M.G.M.M.; Yang, S.J.; Lee, N.K.; Paik, H.D. Probiotic properties of Lactobacillus brevis KU200019 and synergistic activity with fructooligosaccharides in antagonistic activity against foodborne pathogens. Food Sci. Anim. Resour. 2020, 40, 297–310. [Google Scholar] [CrossRef]
- Miyazawa, K.; He, F.; Yoda, K.; Hiramatsu, M. Potent effects of, and mechanisms for, modification of cross-talk between macrophages and adipocytes by lactobacilli. Microbiol. Immunol. 2012, 56, 847–854. [Google Scholar] [CrossRef]
- Choi, H.J.; Oh, Y.J.; Jang, J.Y.; Lee, J.E.; Kwon, M.S.; Lim, S.K. Weissella cibaria WIKIM28 Having Anti-Obesity Activity and Composition for Comprising the Same. KR Patent 1834383, 26 February 2018. [Google Scholar]
- Choi, H.J.; Oh, Y.J.; Jang, J.Y.; Lee, J.E. Weissella confusa WIKIM51 Having Anti-Obesity Activity and Composition for Comprising the Same. KR Patent 1834379, 26 February 2018. [Google Scholar]
- Seo, J.Y.; Kim, J.R.; Yoo, B.W. Interpretation of liver function test. Korean J. Fam. Pract. 2012, 2, 207–213. [Google Scholar]
- Kwon, J.Y.; Cheigh, H.S.; Song, Y.O. Weight reduction and lipid lowering effects of kimchi lactic acid powder in rats fed high fas diets. Korean J. Food. Sci. Technol. 2004, 36, 1014–1019. [Google Scholar]
- Jang, H.J. Potential Use of Lactic Acid Bacteria Isolated from Kimchi as Probiotics. Master’s Thesis, Sookmyung Women’s University, Seoul, Korea, 2018. [Google Scholar]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Ana, M.R.; Thomas, P.B. Lipase inhibitory activity of skim milk fermented with different strains of lactic acid bacteria. J. Funct. Foods 2019, 60, 103413. [Google Scholar]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Kim, K.A.; Gu, W.; Lee, I.A.; Joh, E.H.; Kim, D.H. High fat diet-induced gut microbiota exacerbates infalmmation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar]
- Ritze, Y.; Bárdos, G.; Claus, A.; Ehrmann, V.; Berqheim, I.; Schwiertz, A.; Bischoff, S.C. Lactobacillus rhamnosus GG protects against non-alcholic fatty liver disease in mice. PLoS ONE 2014, 9, e80169. [Google Scholar] [CrossRef]
- Yoo, S.R.; Kim, Y.J.; Park, D.Y.; Jung, U.J.; Jeon, S.M.; Ahn, Y.T.; Huh, C.S.; McGregor, R.; Choi, M.S. Probiotics L. plantarum and L. curvatus in combination alter hepatic lipid metabolism and suppress diet-induced obesity. Obesity 2013, 21, 2421–2603. [Google Scholar] [CrossRef] [PubMed]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef]
- Bertozzi, C.R.; Kiessling, L.L. Chemical glycobiology. Science 2001, 23, 2357–2364. [Google Scholar] [CrossRef]
- Ko, K.H.; Kim, J.Y.; Kim, Y.G.; Oh, K.Y.; Goo, Y.M.; Son, Y.H.; Park, B.R.; Kim, G.S.; Lim, J.H.; Nam, S.H. Isolation and identification of α-glucosidase inhibitory compounds from artemisia annua L. leaves and stems. J. Agric. Life Sci. 2016, 50, 161–173. [Google Scholar] [CrossRef]
- Yoshio, K. Obesity and Related Diseases; Koonja Publishing Company: Seoul, Korea, 2005; pp. 691–692. [Google Scholar]
- EI-shiekh, R.A.; AI-Mahdy, D.A.; Hifnawy, M.S.; Abdel-Sattar, E.A. In-vitro screening of selected traditional medicinal plants for their anti-obesity and anti-oxidant activities. J. Bot. 2019, 123, 43–50. [Google Scholar]
- Kim, S.K.; Lim, S.D. Physiological characteristics and anti-obesity effect of milk fermented by Lactobacillus plantarum KI134. J. Dairy Sci. Biotechnol. 2020, 38, 207–221. [Google Scholar] [CrossRef]
- Charles, M. Fat digestion and absorption. In Encyclopeida of Gastroenterology; Leonard, R.J., Ed.; Academic Press: Cambridge, MA, USA, 2004; pp. 23–30. [Google Scholar]
- Bray, G.A. Medications for obesity: Mechanisms and applications. Clin. Chest Med. 2009, 30, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Lee, S.Y.; Lee, A.R. The effect of Selaginella tamariscina on inhibition of pancreatic lipase and lipid accumulation. J. Korean Food. Nutr. 2018, 32, 27–32. [Google Scholar]
- Kang, C.H.; Jeong, Y.A.; Han, S.H.; Kim, J.S.; Kim, Y.G.; Park, H.M.; Choi, S.I.; Paek, N.S. In vitro probiotic evaluation of potential antiobesity lactic acid bacteria isolated from human vagina and shellfish. KSBB J. 2018, 33, 161–167. [Google Scholar] [CrossRef]
- Kim, E.J.; Kang, M.J.; Seo, Y.B.; Nam, S.W.; Kim, G.D. Acer okamotoanum Nakai leaf extract inhibits adipogenesis via suppressing expression of PPAR γ and C/EBP α in 3T3-L1 cells. J. Microbiol. Biotechnol. 2018, 28, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Jeong, J.E.; Moon, S.H.; Park, K.Y. Anti-obesity effect of Bacillus subtilis KC-3 fermented soymilk in 3T3-L1 adipocytes. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1126–1131. [Google Scholar]
- Song, M.Y.; Bose, S.; Kim, H.J. Effect of probiotics-fermented Samjunghwan on differentiation in 3T3-L1 preadipocytes. J. Korean Soc. Food Sci. Nutr. 2013, 42, 1–7. [Google Scholar] [CrossRef]
- Kim, S.K.; Lim, S.D. Physiological characteristics and anti-obesity effect of Lactobacillus plantarum K6 isolated from Kimchi. J. Milk Sci. Biotechnol. 2017, 35, 221–231. [Google Scholar] [CrossRef]
- Park, H.E.; Do, K.H.; Jeong, J.W.; Jung, Y.H.; Lee, W.K. Cholesterol and visceral fat owering effects of combined lactic acid bacteria (Lactobacillus casei WK3, Enterococus faecium WK5, Bifidobacterium longum WK9 and Lactobacillus plantarum K-1) in high fat diet-fed C57BL/6J Mice. J. Korean Soc. Food Sci. Nutr. 2018, 12, 1210–1216. [Google Scholar] [CrossRef]
- Go, G.Y.; Park, J.H.; Kwak, J.Y.; Ahn, T.W. Anti-obesity effect of yeoldahanso-tang on obesity mice. J. Sassng Const. Med. 2017, 29, 154–173. [Google Scholar]
- Ryu, H.J.; Um, M.Y.; Ahn, J.Y.; Jung, C.H.; Huh, D.; Kim, T.W.; Ha, T.Y. Anti-obesity effect of Hypsizigus marmoreus in high fat-fed mice. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1708–1714. [Google Scholar] [CrossRef]
- Song, M.Y.; Jung, H.W.; Park, Y.K. Anti-obesity effect of water extract of Coix lacrymajobi var. mayuen in high fat fed C5BL/6 mice. J. Korean Med. Obes. Res. 2016, 16, 27–35. [Google Scholar] [CrossRef]
- Bob, T.; Robert, R.M.; Takanori, H.; Abraham, N.; Colin, R.; Thomas, N.; David, E.M.; Wolfgang, K.; Karin, K.; Ulrich, D.; et al. Proliferative and nonproliferative lesions of the rat and mouse hepatobiliary system. J. Toxicol. Pathol. 2010, 38, 5S–81S. [Google Scholar]
- Yu, Z.; Mao, C.; Fu, X.; Ma, M. High density lipoprotein from egg yolk (EYHDL) improves dyslipidemia by mediating fatty acids metabolism in high fat diet-induced obese mice. Food Sci. Anim. Resour. 2019, 39, 179–196. [Google Scholar] [CrossRef]
- Park, Y.M.; Oh, H.G.; Kang, Y.G.; Kim, Y.P.; Sin, H.S.; Jang, S.H.; Kim, H.J.; Lee, S.W.; Lee, H.Y. Anti-obesity effects of lactic acid bacteria-fermented Dioscoreae Rhizoma powder on high fat diet-fed animal model. J. Physiol. Pathol. Korean Med. 2015, 29, 267–272. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.S.; Seo, S.W. Anti-obesity effect of Aster yomena ethanol extract in high fat diet-induced obese mice. J. Physiol. Pathol. Korean Med. 2017, 31, 348–355. [Google Scholar] [CrossRef]
- Hur, H.J.; Hwang, J.T. Effect of silybin on body weight and glucose tolerance in high-fat-diet induced obese mice. KSBB J. 2011, 26, 78–82. [Google Scholar] [CrossRef]
- Park, S.J.; Choi, C.Y.; Jeong, J.C.; Lee, R.K.; Hwang, Y.P. Artemisia annua extract ameliorates high-fat diet-induced fatty liver by activating AMPK. J. Biomed. Transl. Res. 2020, 21, 59–71. [Google Scholar] [CrossRef]
- Ikarashi, N.; Toda, T.; Okaniwa, T.; Ito, K.; Ochiai, W.; Sugiyama, K. Anti-obesity and anti-diabetic effects of Acacia polyphenol in obese diabetic KKAy mice fed high-fat diet. Evid. Based Complement. Alternat. Med. 2011, 2011, 952031. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Choi, B.K.; Jung, H.J. Anti-obesity effects of mixture of Atractylodes macrocephala and Amomum villosum extracts. J. Physiol. Pathol. Korean Med. 2019, 33, 282–287. [Google Scholar] [CrossRef]
- Cho, J.K.; Kang, H.S.; Yoon, J.H. Exercise strategies for the prevention and treatment of non-alcoholic fatty liver disease. Korean J. Obes. 2015, 24, 190–196. [Google Scholar] [CrossRef]
- Lee, J.B.; Lee, J.W.; Hong, Y.M. Serum fatty acid and carnitine levels in obese children with fatty livers. J. Korean Pediatr. Soc. 2002, 45, 1083–1089. [Google Scholar]
- Lee, J.W.; Hong, Y.M. Changes of serum fatty acid and carnitine levels after administration of L-carnitine in rats. J. Korean Pediatr. Soc. 2002, 45, 1075–1082. [Google Scholar]
- Kim, T.R.; Kim, Y.J.; Woo, C.H. Anti-obesity effects of Banggihwnggi-tang-hap-yeonggyechulgam-tang in high fat diet induced obese mice. J. Korean Med. Rehabil. 2019, 29, 29–45. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).