
Pea Protein Extraction Assisted by Lactic Fermentation: Impact on Protein Profile and Thermal Properties



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
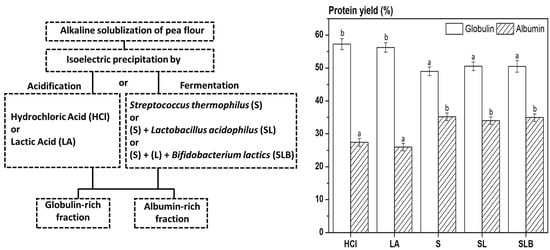
2.2. Pea Protein Extraction and Purification
2.3. Proximate Analysis
2.4. Nitrogen Extraction Yield
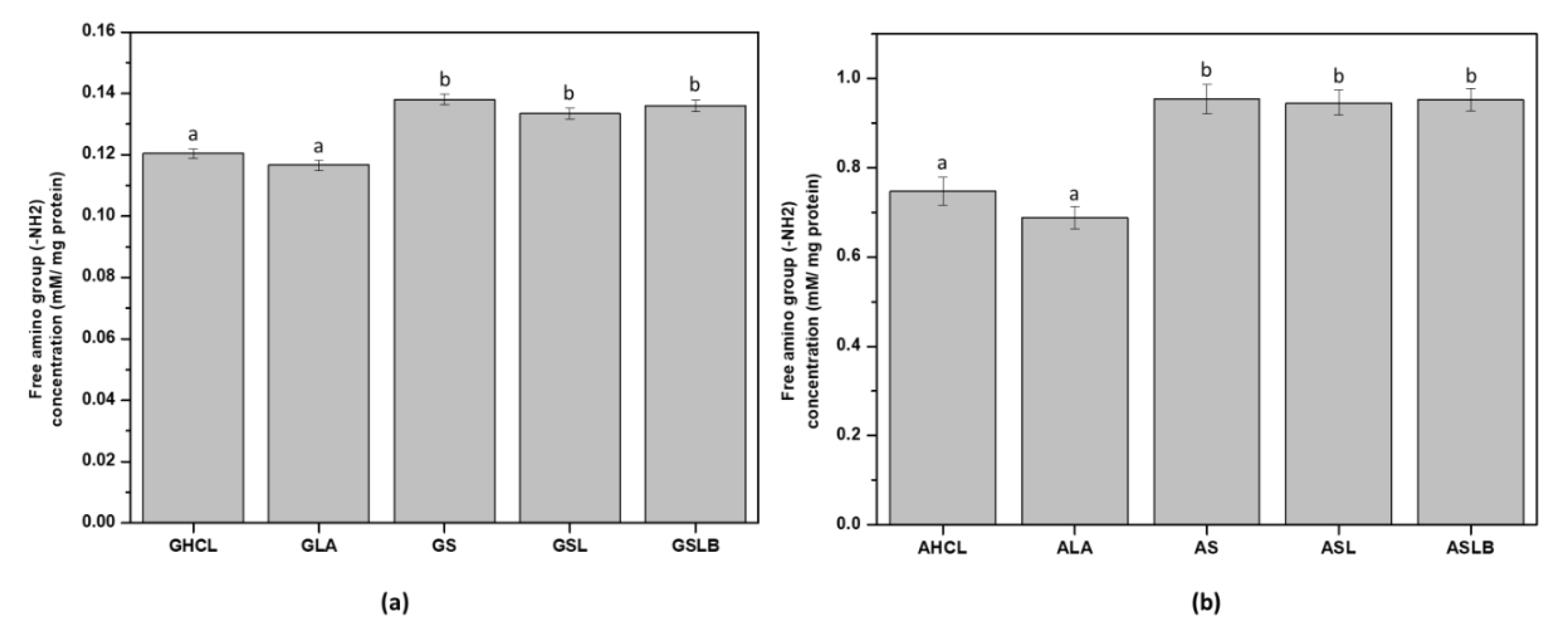
2.5. Protein Composition by SDS PolyAcrylamide Gel Electrophoresis (SDS-PAGE)
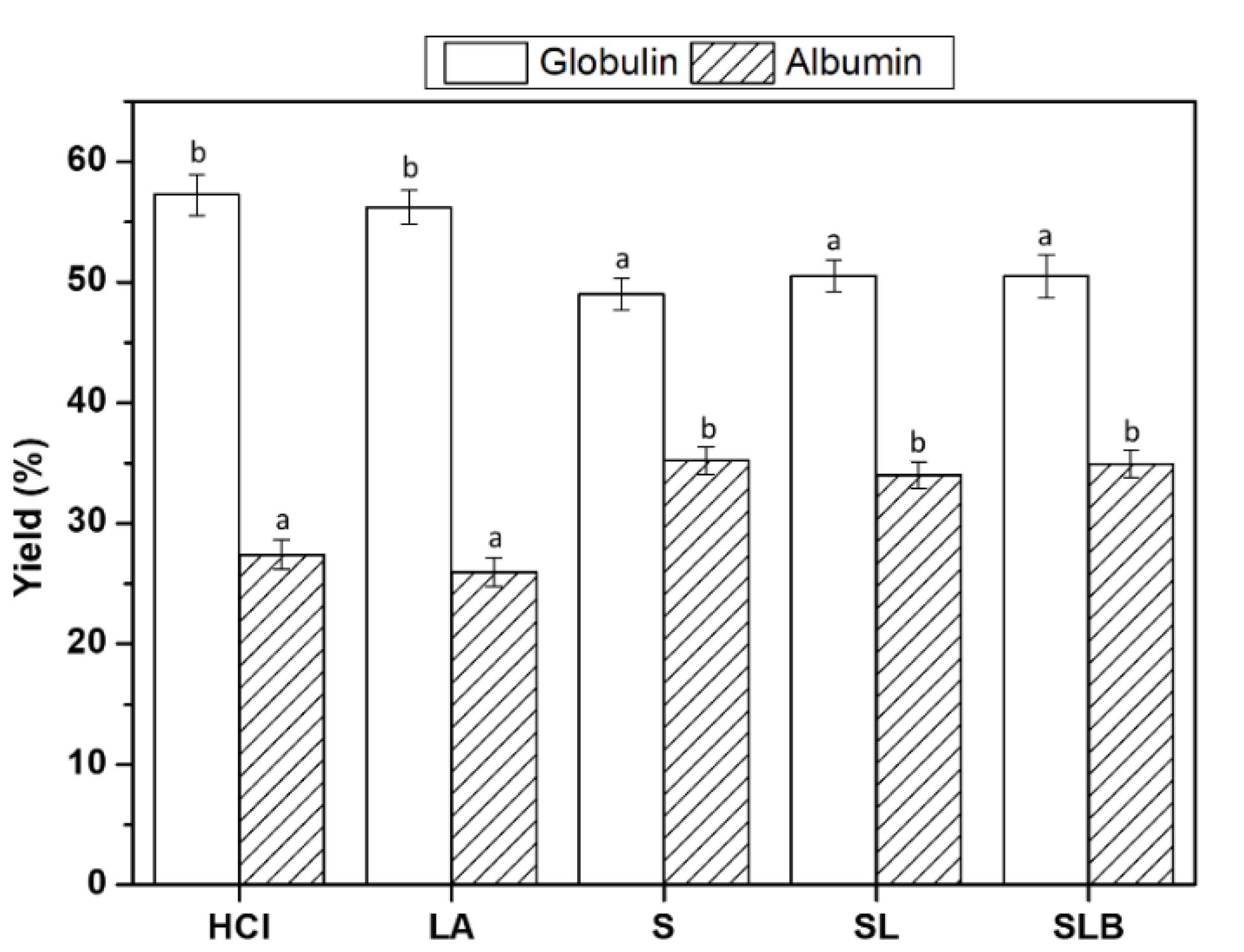
2.6. Free Amino Groups
2.7. Differential Scanning Calorimetry (DSC)
2.8. Statistical Analysis
3. Results
3.1. Acidification Kinetics
3.2. Nitrogen Content in Albumin and Globulin Fractions
3.3. Nitrogen Extraction Yield
3.4. Free Amino Group Content
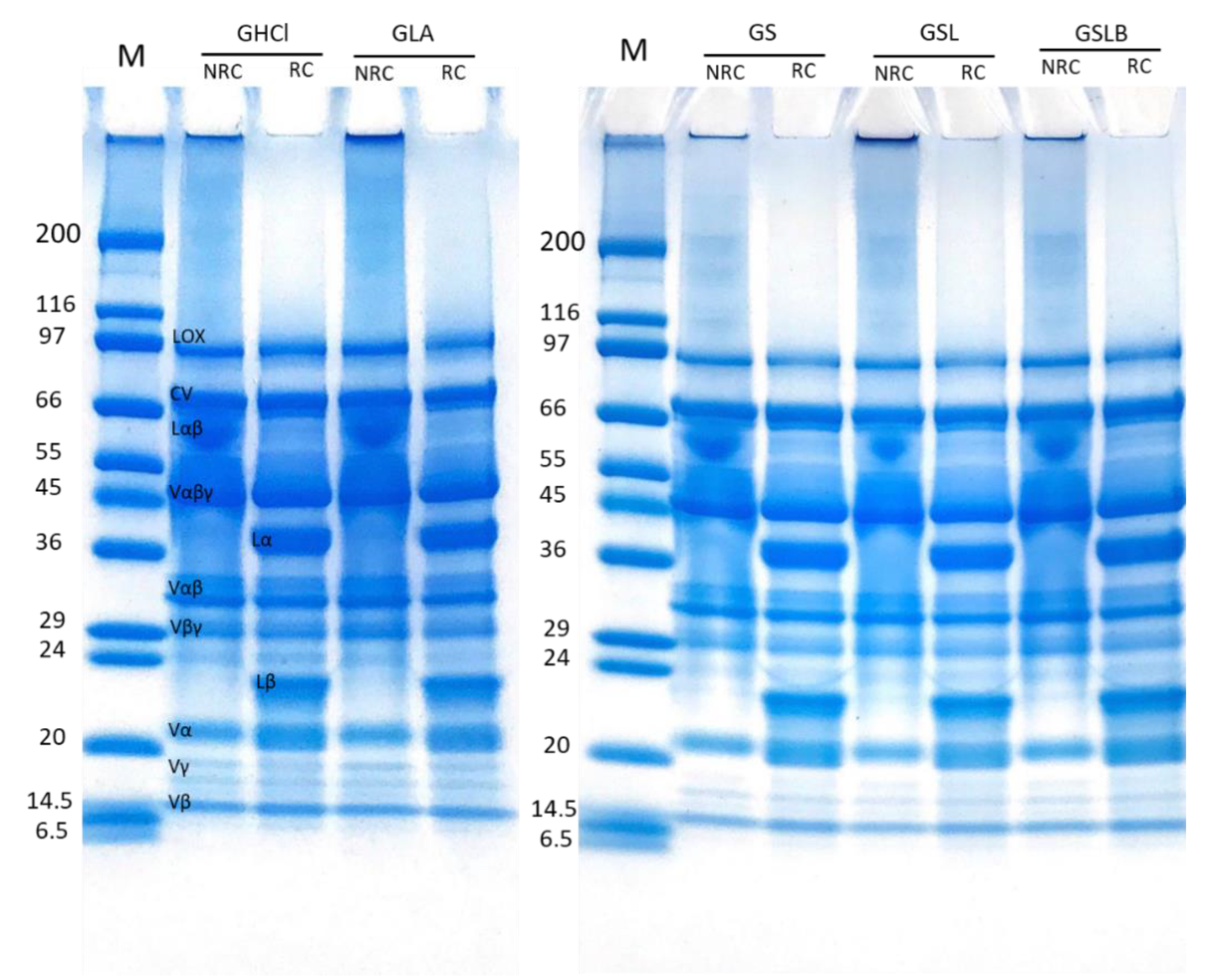
3.5. Polypeptide Profile
3.6. Thermal Properties
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.C.; Helinck, S.; Landaud, S. Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef]
- Pihlanto, A.; Mattila, P.; Makinen, S.; Pajari, A.M. Bioactivities of alternative protein sources and their potential health benefits. Food Funct 2017, 8, 3443–3458. [Google Scholar] [CrossRef]
- Lam, A.C.Y.; Can Karaca, A.; Tyler, R.T.; Nickerson, M.T. Pea protein isolates: Structure, extraction, and functionalityPea protein isolates: Structure, extraction, and functionality. Food Rev. Int. 2018, 34, 126–147. [Google Scholar] [CrossRef]
- Barać, M.B.; Pešić, M.B.; Stanojević, S.P.; Kostić, A.Z.; Čabrilo, S.B. Techno-functional properties of pea (Pisum sativum) protein isolates: A review. Acta Period. Technol. 2015, 46, 1–18. [Google Scholar] [CrossRef]
- Koyoro, H.; Powers, J. Functional properties of pea globulin fractions. Cereal Chem. 1987, 64, 97–101. [Google Scholar]
- Boye, J.I.; Aksay, S.; Roufik, S.; Ribéreau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Int. Food Res. J. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- Crévieu, I.; Berot, S.; Guéguen, J. Large scale procedure for fractionation of albumins and globulins from pea seeds. Mol. Nutr. Food Res. 1996, 40, 237–244. [Google Scholar] [CrossRef]
- Lu, B.Y.; Quillien, L.; Popineau, Y. Foaming and emulsifying properties of pea albumin fractions and partial characterisation of surface-active components. J. Sci. Food Agric. 2000, 80, 1964–1972. [Google Scholar] [CrossRef]
- Gressent, F.; Da Silva, P.; Eyraud, V.; Karaki, L.; Royer, C. Pea Albumin 1 subunit b (PA1b), a promising bioinsecticide of plant origin. Toxins 2011, 3, 1502–1517. [Google Scholar] [CrossRef] [PubMed]
- Gruen, L.C.; Guthrie, R.E.; Blagrove, R.J. Structure of a major pea seed albumin: Implication of a free sulphydryl group. J. Sci. Food Agric. 1987, 41, 167–178. [Google Scholar] [CrossRef]
- Higgins, T.J.; Chandler, P.M.; Randall, P.J.; Spencer, D.; Beach, L.R.; Blagrove, R.J.; Kortt, A.A.; Inglis, A.S. Gene structure, protein structure, and regulation of the synthesis of a sulfur-rich protein in pea seeds. J. Biol. Chem. 1986, 261, 11124–11130. [Google Scholar] [CrossRef]
- O’Kane, F.E.; Happe, R.P.; Vereijken, J.M.; Gruppen, H.; van Boekel, M.A. Characterization of pea vicilin. 2. Consequences of compositional heterogeneity on heat-induced gelation behavior. J. Agric. Food Chem. 2004, 52, 3149–3154. [Google Scholar] [CrossRef]
- Liang, H.-N.; Tang, C.-H. pH-dependent emulsifying properties of pea [Pisum sativum (L.)] proteins. Food Hydrocoll. 2013, 33, 309–319. [Google Scholar] [CrossRef]
- Dagorn-Scaviner, C.; Gueguen, J.; Lefebvre, J. A comparison of interfacial behaviours of pea (Pisum sativum L.) legumin and vicilin at air/water interface. Mol. Nutr. Food Res. 1986, 30, 337–347. [Google Scholar] [CrossRef]
- Tzitzikas, E.N.; Vincken, J.-P.; de Groot, J.; Gruppen, H.; Visser, R.G.F. Genetic variation in pea seed globulin composition. J. Agric. Food Chem. 2006, 54, 425–433. [Google Scholar] [CrossRef]
- Gao, Z.; Shen, P.; Lan, Y.; Cui, L.; Ohm, J.-B.; Chen, B.; Rao, J. Effect of alkaline extraction pH on structure properties, solubility, and beany flavor of yellow pea protein isolate. Food Res. Int. 2020, 131, 109045. [Google Scholar] [CrossRef] [PubMed]
- Stone, A.K.; Karalash, A.; Tyler, R.T.; Warkentin, T.D.; Nickerson, M.T. Functional attributes of pea protein isolates prepared using different extraction methods and cultivars. Food Res. Int. 2015, 76, 31–38. [Google Scholar] [CrossRef]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef]
- Matejčeková, Z.; Dujmić, E.; Liptáková, D.; Valík, Ľ. Modeling of lactic acid fermentation of soy formulation with Lactobacillus plantarum HM1. Food Sci. Technol. Int. 2019, 25, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Rui, X.; Zhang, Q.; Huang, J.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Does lactic fermentation influence soy yogurt protein digestibility: A comparative study between soymilk and soy yogurt at different pH. J. Sci. Food Agric. 2019, 99, 861–867. [Google Scholar] [CrossRef]
- Xing, Q.; Dekker, S.; Kyriakopoulou, K.; Boom, R.M.; Smid, E.J.; Schutyser, M.A.I. Enhanced nutritional value of chickpea protein concentrate by dry separation and solid state fermentation. Innov. Food Sci. Emerg. Technol. 2020, 59, 102269. [Google Scholar] [CrossRef]
- Cabuk, B.; Nosworthy, M.G.; Stone, A.K.; Korber, D.R.; Tanaka, T.; House, J.D.; Nickerson, M.T. Effect of Fermentation on the Protein Digestibility and Levels of Non-Nutritive Compounds of Pea Protein Concentrate. Food Technol. Biotechnol. 2018, 56, 257–264. [Google Scholar] [CrossRef]
- Stanisavljevic, N.; Vukotic, G.; Pastor, F.; Suznjevic, D.; Jovanovic, Z.; Strahinic, I.; Fira, D.; Radovic, S. Antioxidant activity of pea protein hydrolysates produced by batch fermentation with lactic acid bacteria. Arch. Biol. Sci. 2015, 67, 1033–1042. [Google Scholar] [CrossRef]
- Vermeirssen, V.; Van Camp, J.; Decroos, K.; Van Wijmelbeke, L.; Verstraete, W. The impact of fermentation and in vitro digestion on the formation of angiotensin-I-converting enzyme inhibitory activity from pea and whey protein. J. Dairy Sci. 2003, 86, 429–438. [Google Scholar] [CrossRef]
- Schindler, S.; Wittig, M.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Lactic fermentation to improve the aroma of protein extracts of sweet lupin (Lupinus angustifolius). Food Chem. 2011, 128, 330–337. [Google Scholar] [CrossRef]
- Yousseef, M.; Lafarge, C.; Valentin, D.; Lubbers, S.; Husson, F. Fermentation of cow milk and/or pea milk mixtures by different starter cultures: Physico-chemical and sensorial properties. LWT Food Sci. Technol. 2016, 69, 430–437. [Google Scholar] [CrossRef]
- Schlegel, K.; Leidigkeit, A.; Eisner, P.; Schweiggert-Weisz, U. Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates. Foods 2019, 8, 678. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physicochemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Orozco, R.; Frias, J.; Muñoz, R.; Zielinski, H.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C. Fermentation as a bio-process to obtain functional soybean flours. J. Agric. Food Chem. 2007, 55, 8972–8979. [Google Scholar] [CrossRef] [PubMed]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.-B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed]
- Klupsaite, D.; Juodeikiene, G.; Zadeike, D.; Bartkiene, E.; Maknickiene, Z.; Liutkute, G. The influence of lactic acid fermentation on functional properties of narrow-leaved lupine protein as functional additive for higher value wheat bread. LWT 2017, 75, 180–186. [Google Scholar] [CrossRef]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of air classification and fermentation by Lactobacillus plantarum VTT E-133328 on faba bean (Vicia faba L.) flour nutritional properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef]
- Bartkiene, E.; Krungleviciute, V.; Juodeikiene, G.; Vidmantiene, D.; Maknickiene, Z. Solid state fermentation with lactic acid bacteria to improve the nutritional quality of lupin and soya bean. J. Sci. Food Agric. 2015, 95, 1336–1342. [Google Scholar] [CrossRef] [PubMed]
- Rui, X.; Wang, M.; Zhang, Y.; Chen, X.; Li, L.; Liu, Y.; Dong, M. Optimization of soy solid-state fermentation with selected lactic acid bacteria and the effect on the anti-nutritional components. J. Food Process. Preserv. 2017, 41, e13290. [Google Scholar] [CrossRef]
- Lampart-Szczapa, E.; Konieczny, P.; Nogala-Kałucka, M.; Walczak, S.; Kossowska, I.; Malinowska, M. Some functional properties of lupin proteins modified by lactic fermentation and extrusion. Food Chem. 2006, 96, 290–296. [Google Scholar] [CrossRef]
- Masiá, C.; Jensen, P.E.; Buldo, P. Effect of Lactobacillus rhamnosus on physicochemical properties of fermented plant-based raw materials. Foods 2020, 9, 1182. [Google Scholar] [CrossRef]
- Sozer, N.; Melama, L.; Silbir, S.; Rizzello, C.G.; Flander, L.; Poutanen, K. Lactic acid fermentation as a pre-treatment process for faba bean flour and its effect on textural, structural and nutritional properties of protein-enriched gluten-free faba bean breads. Foods 2019, 8, 431. [Google Scholar] [CrossRef]
- Ben-Harb, S.; Saint-Eve, A.; Panouille, M.; Souchon, I.; Bonnarme, P.; Dugat-Bony, E.; Irlinger, F. Design of microbial consortia for the fermentation of pea-protein-enriched emulsions. Int. J. Food Microbiol. 2019, 293, 124–136. [Google Scholar] [CrossRef]
- Czarnecka, M.; Czarnecki, Z.; Nowak, J.; Roszyk, H. Effect of lactic fermentation and extrusion of bean and pea seeds on nutritional and functional properties. Food/Nahrung 1998, 42, 7–11. [Google Scholar] [CrossRef]
- Oliete, B.; Yassine, S.A.; Cases, E.; Saurel, R. Drying method determines the structure and the solubility of microfluidized pea globulin aggregates. Food Res. Int. 2019, 119, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Helrich, K. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 1990; ISBN 978-0935584424. [Google Scholar]
- Awolumate, E.O. Accumulation and quality of storage protein in developing cowpea, mung bean and soya bean seeds. J. Sci. Food Agric. 1983, 34, 1351–1357. [Google Scholar] [CrossRef]
- Sosulski, F.W.; Holt, N.W. Amino acid composition and nitrogen-to-protein factors for grain legumes. Can. J. Plant Sci. 1980, 60, 1327–1331. [Google Scholar] [CrossRef]
- Church, F.C.; Porter, D.H.; Catignani, G.L.; Swaisgood, H.E. An o-phthalaldehyde spectrophotometric assay for proteinases. Anal. Biochem. 1985, 146, 343–348. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J.K. Effect of variety and crude protein content on nutrients and certain antinutrients in field peas (Pisum sativum). J. Sci. Food Agric. 2004, 84, 1021–1029. [Google Scholar] [CrossRef]
- Gänzle, M.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef]
- Boulay, M.; Al Haddad, M.; Rul, F. Streptococcus thermophilus growth in soya milk: Sucrose consumption, nitrogen metabolism, soya protein hydrolysis and role of the cell-wall protease PrtS. Int. J. Food Microbiol. 2020, 335, 108903. [Google Scholar] [CrossRef] [PubMed]
- Blagden, T.D.; Gilliland, S.E. Reduction of levels of volatile components associated with the “beany” flavor in soymilk by lactobacilli and streptococci. J. Food Sci. 2005, 70, M186–M189. [Google Scholar] [CrossRef]
- Murti, T.W.; Bouillanne, C.; Landon, M.; De Smazaud, M.J. Bacterial growth and volatile compounds in yoghurt-type products from soymilk containing Bifidobacterium ssp. J. Food Sci. 1993, 58, 153–157. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Yu, R.-C.; Chou, C.-C. Growth and survival of bifidobacteria and lactic acid bacteria during the fermentation and storage of cultured soymilk drinks. Food Microbiol. 2002, 19, 501–508. [Google Scholar] [CrossRef]
- de Souza Oliveira, R.P.; Perego, P.; de Oliveira, M.N.; Converti, A. Growth, organic acids profile and sugar metabolism of Bifidobacterium lactis in co-culture with Streptococcus thermophilus: The inulin effect. Food Res. Int. 2012, 48, 21–27. [Google Scholar] [CrossRef]
- Helland, M.H.; Wicklund, T.; Narvhus, J.A. Growth and metabolism of selected strains of probiotic bacteria in milk-and water-based cereal puddings. Int. Dairy J. 2004, 14, 957–965. [Google Scholar] [CrossRef]
- Moon, N.J.; Reinbold, G. Commensalism and competition in mixed cultures of Lactobacillus bulgaricus and Streptococcus thermophilus. J. Milk Food Technol. 1976, 39, 337–341. [Google Scholar] [CrossRef]
- Garcia, S.L.A.; da Silva, G.M.; Medeiros, J.M.S.; de Queiroga, A.P.R.; de Queiroz, B.B.; de Farias, D.R.B.; Correia, J.O.; Florentino, E.R.; Buriti, F.C.A. Influence of co-cultures of Streptococcus thermophilus and probiotic lactobacilli on quality and antioxidant capacity parameters of lactose-free fermented dairy beverages containing Syzygium cumini (L.) skeels pulp. RSC Adv. 2020, 10, 10297–10308. [Google Scholar] [CrossRef]
- Božanić, R.; Pletikapić, G.; Lovković, S. Influence of temperature and glucose addition on growth and survival of bacteria from BCT culture in soymilk. Mljekarstvo 2008, 58, 171–179. Available online: https://hrcak.srce.hr/23062 (accessed on 15 October 2020).
- Gueguen, J.; Barbot, J. Quantitative and qualitative variability of pea (Pisum sativum L.) protein composition. J. Sci. Food Agric. 1988, 42, 209–224. [Google Scholar] [CrossRef]
- Aguirre, L.; Garro, M.S.; Savoy de Giori, G. Enzymatic hydrolysis of soybean protein using lactic acid bacteria. Food Chem. 2008, 111, 976–982. [Google Scholar] [CrossRef]
- Rui, X.; Huang, J.; Xing, G.; Zhang, Q.; Li, W.; Dong, M. Changes in soy protein immunoglobulin E reactivity, protein degradation, and conformation through fermentation with Lactobacillus plantarum strains. LWT 2019, 99, 156–165. [Google Scholar] [CrossRef]
- Verni, M.; Wang, C.; Montemurro, M.; De Angelis, M.; Katina, K.; Rizzello, C.G.; Coda, R. Exploring the Microbiota of Faba Bean: Functional Characterization of Lactic Acid Bacteria. Front. Microbiol. 2017, 8, 2461. [Google Scholar] [CrossRef]
- Xu, Y.; Coda, R.; Holopainen-Mantila, U.; Laitila, A.; Katina, K.; Tenkanen, M. Impact of in situ produced exopolysaccharides on rheology and texture of fava bean protein concentrate. Food Res. Int. 2019, 115, 191–199. [Google Scholar] [CrossRef]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef]
- Dziuba, J.; Szerszunowicz, I.; Nałęcz, D.; Dziuba, M. Proteomic analysis of albumin and globulin fractions of pea (Pisum sativum L.) seeds. Acta Sci. Pol. Technol. Aliment. 2014, 13, 181–190. [Google Scholar] [CrossRef]
- Gatehouse, J.A.; Lycett, G.W.; Croy, R.R.D.; Boulter, D. The post-translational proteolysis of the subunits of vicilin from pea (Pisum sativum L.). Biochem 1982, 207, 629–632. [Google Scholar] [CrossRef]
- Shand, P.J.; Ya, H.; Pietrasik, Z.; Wanasundara, P.K.J.P.D. Physicochemical and textural properties of heat-induced pea protein isolate gels. Food Chem. 2007, 102, 1119–1130. [Google Scholar] [CrossRef]
- Barkholt, V.; Jergensen, P.B.; Sørensen, D.; Bahrenscheer, J.; Haikara, A.; Lemola, E.; Laitila, A.; Frøkiær, H. Protein modification by fermentation: Effect of fermentation on the potential allergenicity of pea. Allergy 1998, 53, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Mession, J.-L.; Sok, N.; Assifaoui, A.; Saurel, R. Thermal Denaturation of Pea Globulins (Pisum sativum L.)-Molecular Interactions Leading to Heat-Induced Protein Aggregation. J. Agric. Food Chem. 2013, 61, 1196–1204. [Google Scholar] [CrossRef]
- Ricci, L.; Umiltà, E.; Righetti, M.C.; Messina, T.; Zurlini, C.; Montanari, A.; Bronco, S.; Bertoldo, M. On the thermal behavior of protein isolated from different legumes investigated by DSC and TGA. J. Sci. Food Agric. 2018, 98, 5368–5377. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.D.; Arntfield, S.D. Molecular forces involved in heat-induced pea protein gelation: Effects of various reagents on the rheological properties of salt-extracted pea protein gels. Food Hydrocoll. 2012, 28, 325–332. [Google Scholar] [CrossRef]
- Sun, X.D.; Arntfield, S.D. Gelation properties of salt-extracted pea protein induced by heat treatment. Food Res. Int. 2010, 43, 509–515. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, G.; Ren, J.; Zhao, M.; Yang, B. Effect of denaturation during extraction on the conformational and functional properties of peanut protein isolate. Innov. Food Sci. Emerg. Technol. 2011, 12, 375–380. [Google Scholar] [CrossRef]
- Mession, J.-L.; Assifaoui, A.; Cayot, P.; Saurel, R. Effect of pea proteins extraction and vicilin/legumin fractionation on the phase behavior in admixture with alginate. Food Hydrocoll. 2012, 29, 335–346. [Google Scholar] [CrossRef]
- Djoullah, A. Réticulation Enzymatique des Protéines de Pois Pour la Formation de Microparticules: Application à L’encapsulation de la Riboflavine., in Alimentation et Nutrition. Doctoral Thesis, Université de Bourgogne, Dijon, France, 2015. Available online: https://tel.archives-ouvertes.fr/tel-01375883 (accessed on 21 February 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tonset (°C) | Td1 (°C) | Td2 (°C) | ΔHd Total (J/g Protein) | Peak 1/Peak2 Area Ratio |
|---|---|---|---|---|---|
| GHCl | 61.58 ± 0.76 a 1 | 71.69± 0.3 a | 83.29 ± 0.69 a | 12.57 ± 0.24 a | 3.5 ± 0.4 b |
| GLA | 61.83 ± 0.18 a | 72.43 ± 0.14 ab | 83.05 ± 0.34 a | 13.05 ± 0.26 a | 3.5 ± 0.2 b |
| GS | 61.93 ± 0.36 a | 72.90 ± 0.04 b | 83.31 ± 0.31 a | 15.11 ± 0.40 b | 2.2 ± 0.2 a |
| GSL | 61.79 ± 0.06 a | 72.15 ± 0.65 ab | 81.97 ± 0.44 a | 14.66 ± 0.33 b | 2.6 ± 0.3 a |
| GSLB | 61.84 ± 0.46 a | 71.91 ± 0.45 a | 82.23 ± 0.78 a | 14.76 ± 0.17 b | 2.4 ± 0.3 a |
| Sample | Tonset (°C) | Td1 (°C) | Td2 (°C) | ΔHd Total (J/g Protein) |
|---|---|---|---|---|
| AHCl | 54.16 ± 0.24 a 1 | 62.14 ± 0.29 a | 76.64 ± 0.18 a | 7.87 ± 0.5 a |
| ALA | 55.17 ± 0.89 a | 62.21 ± 0.25 a | 76.69 ± 0.04 a | 8.36 ± 0.55 a |
| AS | 54.11 ± 0.71 a | 63.88 ± 0.13 b | 78.56 ± 0.72 a | 9.42 ± 0.49 b |
| ASL | 55.70 ± 0.35 a | 63.95 ± 0.16 b | 78.15 ± 0.77 a | 9.55 ± 0.54 b |
| ASLB | 55.20 ± 0.47 a | 63.71 ± 0.03 b | 77.81 ± 0.38 a | 9.43 ± 0.45 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emkani, M.; Oliete, B.; Saurel, R. Pea Protein Extraction Assisted by Lactic Fermentation: Impact on Protein Profile and Thermal Properties. Foods 2021, 10, 549. https://doi.org/10.3390/foods10030549
Emkani M, Oliete B, Saurel R. Pea Protein Extraction Assisted by Lactic Fermentation: Impact on Protein Profile and Thermal Properties. Foods. 2021; 10(3):549. https://doi.org/10.3390/foods10030549
Chicago/Turabian StyleEmkani, Mehrsa, Bonastre Oliete, and Rémi Saurel. 2021. "Pea Protein Extraction Assisted by Lactic Fermentation: Impact on Protein Profile and Thermal Properties" Foods 10, no. 3: 549. https://doi.org/10.3390/foods10030549
APA StyleEmkani, M., Oliete, B., & Saurel, R. (2021). Pea Protein Extraction Assisted by Lactic Fermentation: Impact on Protein Profile and Thermal Properties. Foods, 10(3), 549. https://doi.org/10.3390/foods10030549