
New Insights on the Impact of Cattle Handling on Post-Mortem Myofibrillar Muscle Proteome and Meat Tenderization

 , , ,
, , ,  and
and
Abstract

1. Introduction
2. Materials and Methods
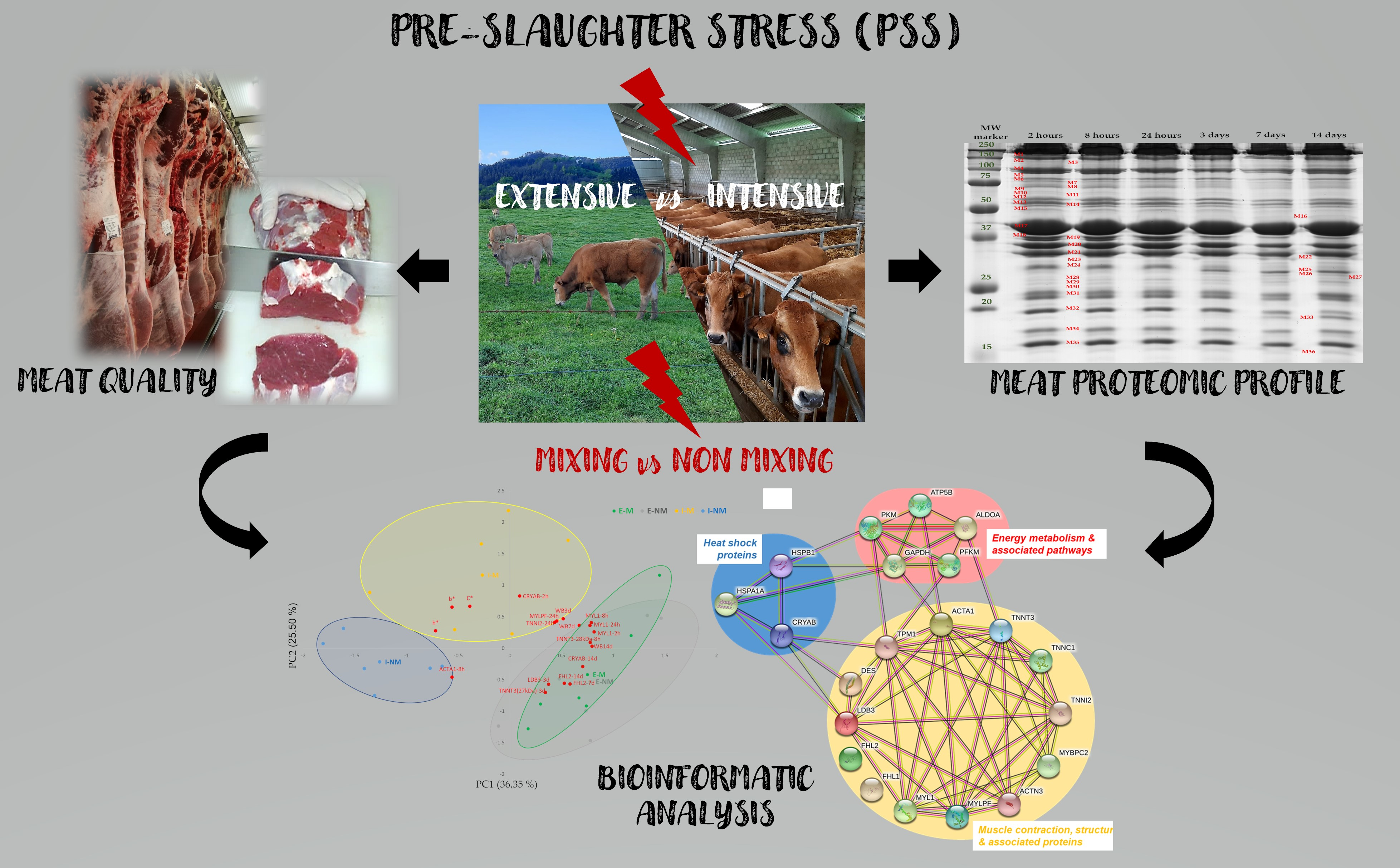
2.1. Animals and Experimental Design
- 1)
- Intensive (“I”) (n = 12), with animals managed indoors, in pens of 6 × 6 m (6 animals per pen) and finished for 100 days before slaughter with 8 kg/day of concentrate (84% barley meal, 10% soya meal, 3% fat, 3% minerals, vitamins and oligoelements) and 2 kg/day of barley straw
- 2)
- Extensive (“E”) (n = 12), with animals managed outdoors in two 1.5 ha plots (6 animals per plot) and finished for 100 days before slaughter grazing on ryegrass and clover pasture + 3.5 kg/day of supplementation with concentrate.
2.2. Muscle Sampling and Meat Quality Measurements
2.3. Myofibrillar Protein Extraction
2.4. Myofibrillar Subproteome Analysis (1D SDS-PAGE) and Protein Identification
2.5. Statistical and Bioinformatics Analyses
3. Results
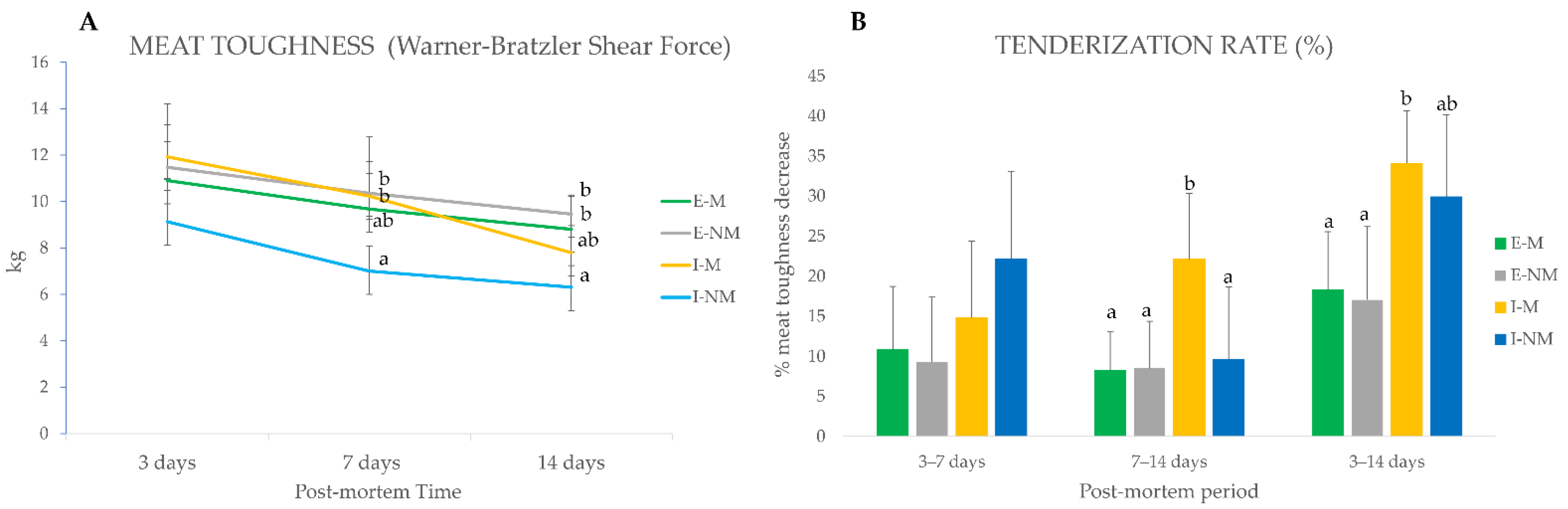
3.1. Meat Quality Attributes
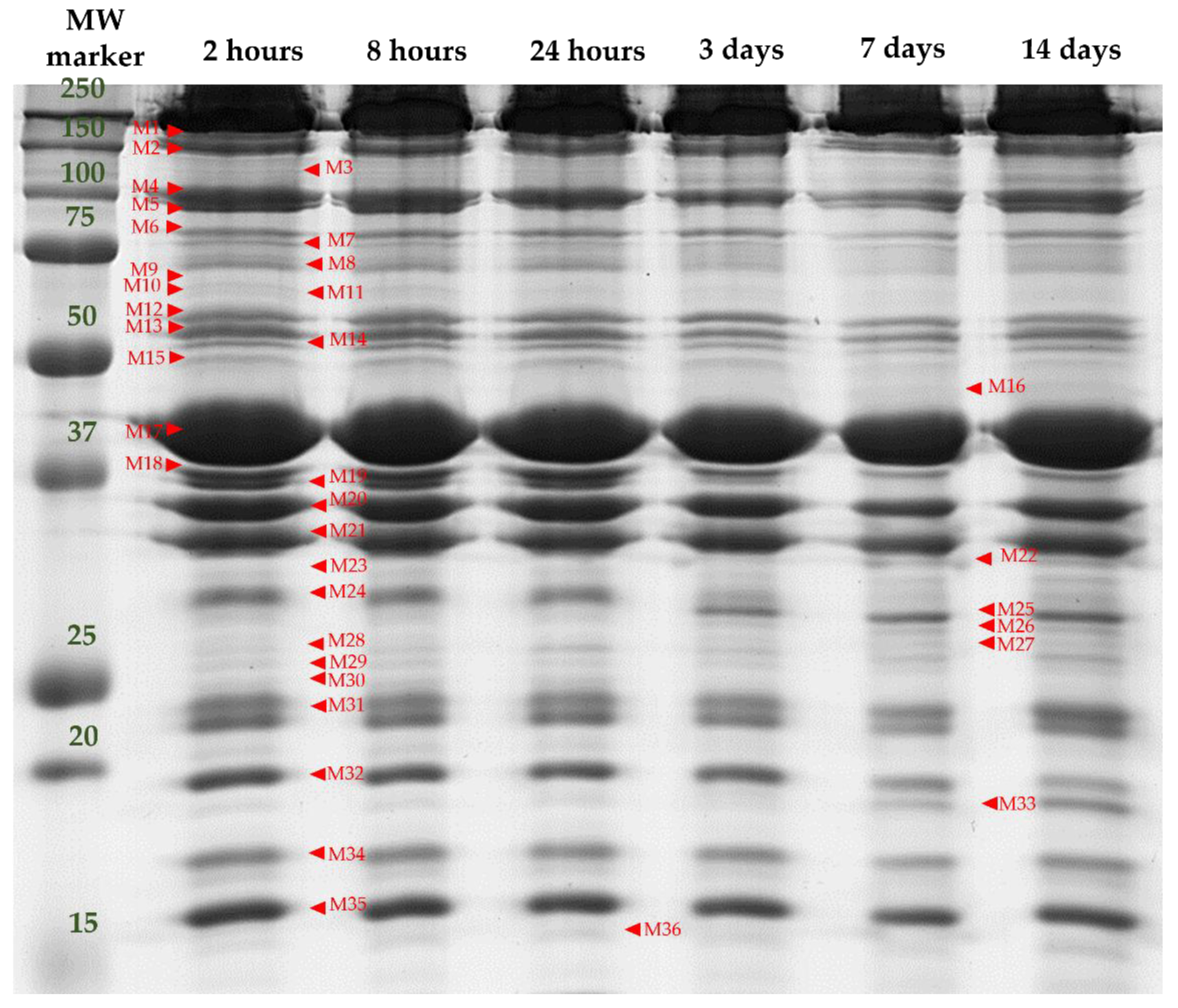
3.2. Separation and Identification of Myofibrillar Subproteome
- ➢
- Muscle contraction, structure and associated proteins: M1 (Myosin-binding protein C, fast-type isoform X2 “MYBPC2”), M4 (Alpha-actinin-3 “ACTN3”), M13 (Desmin, partial “DES”), M17 (Actin, alpha skeletal muscle “ACTA1”), M21 (Tropomyosin alpha-1 chain “TPM1”), M23 (LIM domain-binding protein 3 isoform X5 “LDB3”), M24 (Four and a half LIM domains protein 1 isoform 1 “FHL1”), M25 and M27 (Troponin T, fast skeletal muscle isoform X31 “TNNT3”), M26 (Four and a half LIM domains protein 1 isoform 2 “FHL2”), M31 (Myosin light chain 1/3 skeletal muscle isoform “MYL1”), M32 (Troponin I, fast skeletal muscle “TNNI2”), M34 (Troponin C, skeletal muscle “TNNC1” and M35 (Myosin regulatory light chain 2, skeletal muscle isoform “MYLPF”);
- ➢
- Energy metabolism and associated pathways: M6 (ATP-dependent 6-phosphofructokinase, muscle type “PFKM”), M12 (Pyruvate kinase PKM isoform X1 “PKM”), M15 (ATP synthase subunit beta, mitochondrial precursor “ATP5F1B”), M18 y M19 (Fructose-biphosphate aldolase A “ALDOA”) and M20 (Glyceraldehide-3-phosphate dehydrogenase, “GAPDH”);
- ➢
- Heat shock proteins: M9 (Heat Shock 70 kDa protein 1A “HSPA1A”), M30 (Heat Shock protein family B member 1 variant 1 “HSPB1”) and M33 (Alpha-crystallin B chain “CRYAB”).
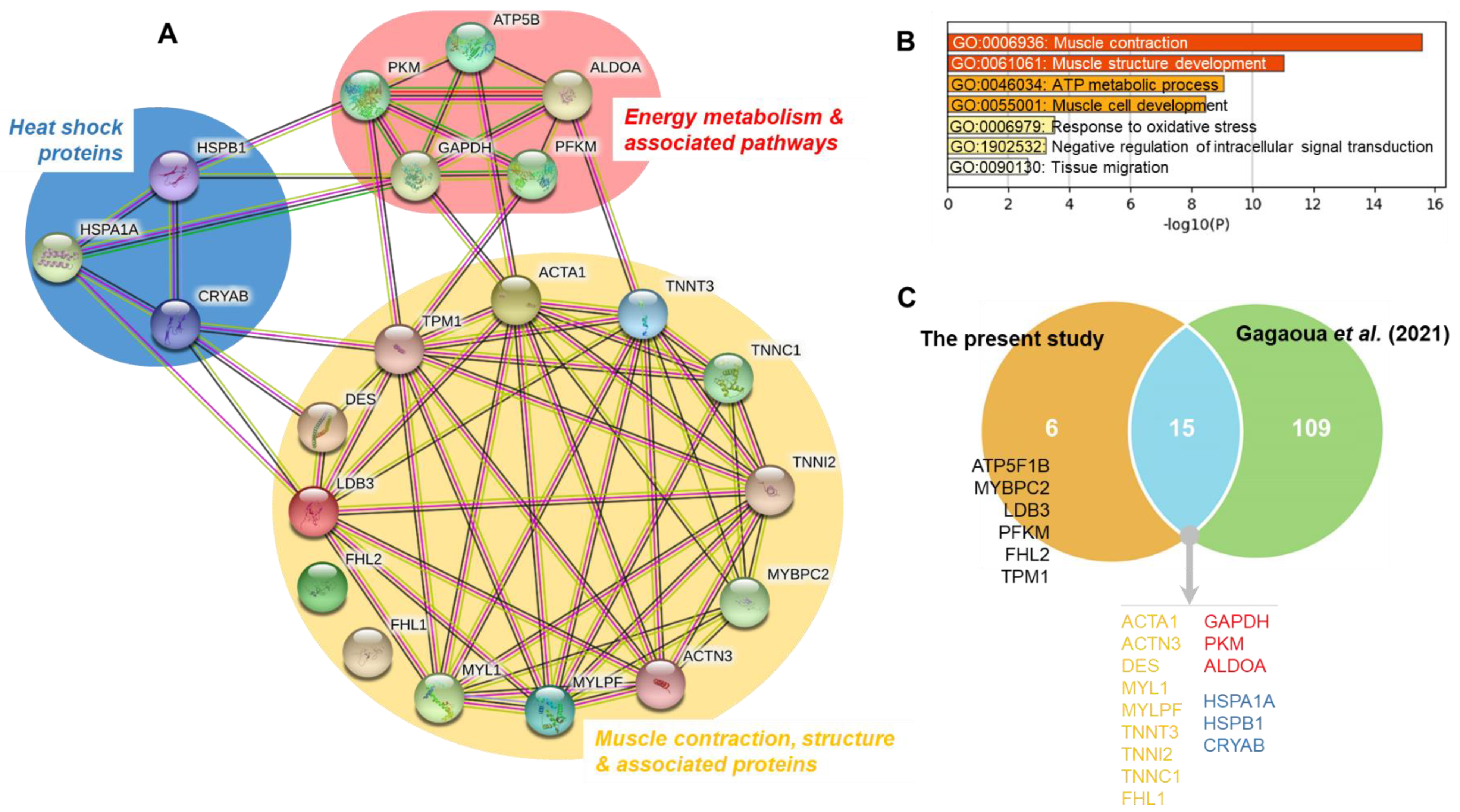
3.2.1. Handling Effects on the Muscle Contraction, Structure and Associated Proteins
3.2.2. Handling Effects on the Energy Metabolism and Associated Pathways Proteins
3.2.3. Handling Effects on the Heat Shock Proteins
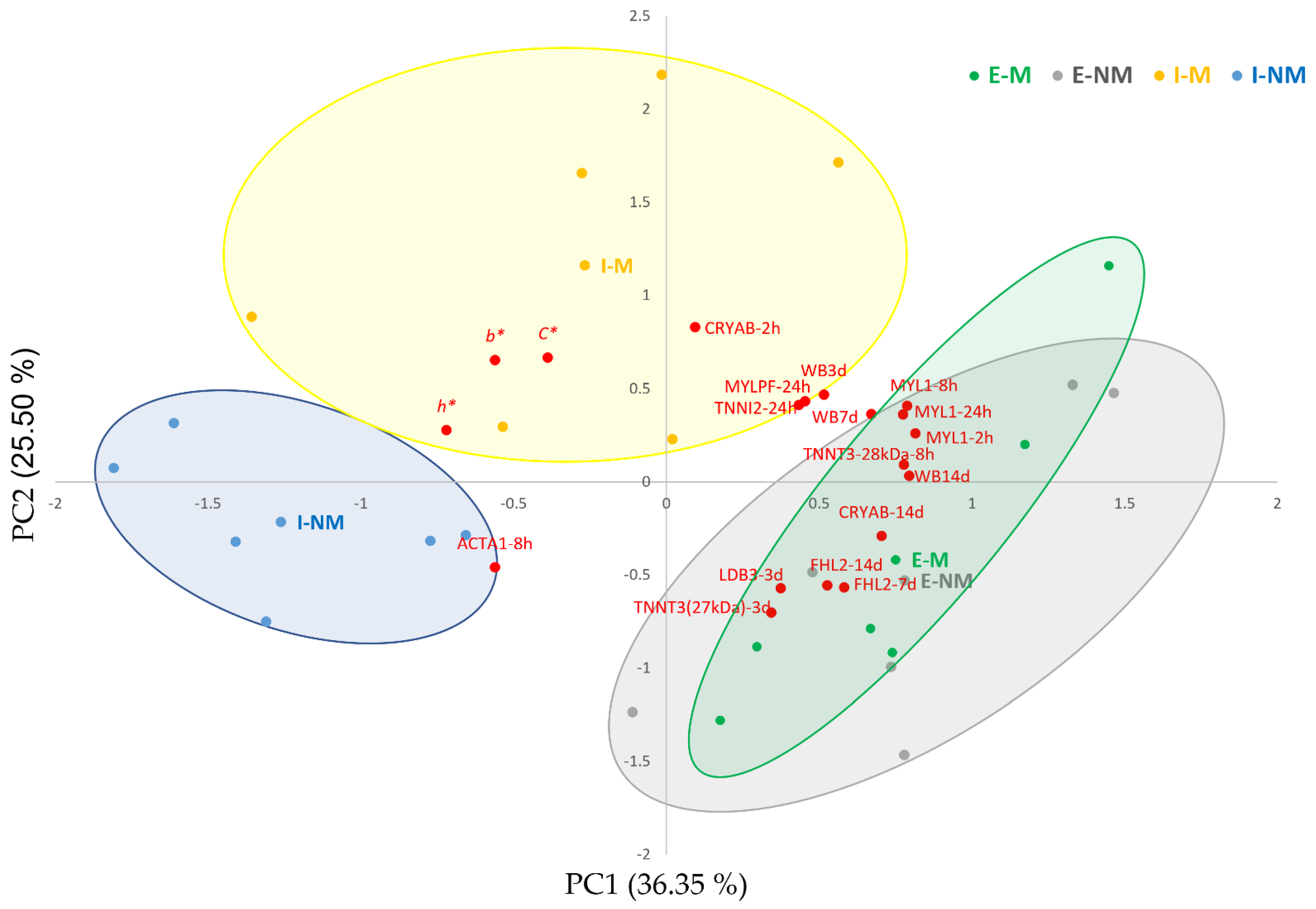
3.3. Relationship between Meat Quality Traits and the Significantly Changing Myofibrillar Proteins
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Picard, B.; Gagaoua, M.; Micol, D.; Cassar-Malek, I.; Hocquette, J.F.; Terlouw, C.E. Inverse relationships between biomarkers and beef tenderness according to contractile and metabolic properties of the muscle. J. Agric. Food Chem. 2014, 62, 9808–9818. [Google Scholar] [CrossRef] [PubMed]
- Gagaoua, M.; Monteils, V.; Picard, B. Data from the farmgate-to-meat continuum including omics-based biomarkers to better understand the variability of beef tenderness: An integromics approach. J. Agric. Food Chem. 2018, 66, 13552–13563. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Rodríguez-Vázquez, R.; Purriños, L.; Oliván, M.; García-Torres, S.; Sentandreu, M.Á.; Lorenzo, J.M.; Zapata, C.; Franco, D. Sensory and physicochemical analysis of meat from bovine breeds in different livestock production systems, pre-slaughter handling conditions, and ageing time. Foods 2020, 9, 176. [Google Scholar] [CrossRef]
- Gagaoua, M.; Picard, B.; Soulat, J.; Monteils, V. Clustering of sensory eating qualities of beef: Consistencies and differences within carcass, muscle, animal characteristics and rearing factors. Livest. Sci. 2018, 214, 245–258. [Google Scholar] [CrossRef]
- Priolo, A.; Micol, D.; Agabriel, J. Effects of grass feeding systems on ruminant meat colour and flavour. A review. Anim. Res. 2001, 50, 185–200. [Google Scholar] [CrossRef]
- Andersen, H.J.; Oksbjerg, N.; Young, J.F.; Therkildsen, M. Feeding and meat quality—A future approach. Meat Sci. 2005, 70, 543–554. [Google Scholar] [CrossRef]
- Bouissou, M.F.; Boissy, A.; le Neindre, P.; Veissier, I. The social behaviour of cattle. Soc. Behav. Farm Anim. 2001, 113–145. [Google Scholar] [CrossRef]
- Marco-Ramell, A.; Arroyo, L.; Saco, Y.; García-Heredia, A.; Camps, J.; Fina, M.; Piedrafita, J.; Bassols, A. Proteomic analysis reveals oxidative stress response as the main adaptative physiological mechanism in cows under different production systems. J. Proteom. 2012, 75, 4399–4411. [Google Scholar] [CrossRef]
- Díaz, F.; Díaz-Luis, A.; Sierra, V.; Diñeiro, Y.; González, P.; García-Torres, S.; Oliván, M. What functional proteomic and biochemical analysis tell us about animal stress in beef? J. Proteom. 2020, 218, 103722. [Google Scholar] [CrossRef] [PubMed]
- Terlouw, E.M.C.; Picard, B.; Deiss, V.; Berri, C.; Hocquette, J.F.; Lebret, B.; Lefèvre, F.; Hamill, R.; Gagaoua, M. Understanding the determination of meat quality using biochemical characteristics of the muscle: Stress at slaughter and other missing keys. Foods 2021, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Bøe, K.E.; Færevik, G. Grouping and social preferences in calves, heifers and cows. Appl. Anim. Behav. Sci. 2003, 80, 175–190. [Google Scholar] [CrossRef]
- Disanto, C.; Celano, G.; Varvara, M.; Fusiello, N.; Fransvea, A.; Bozzo, G.; Celano, G.V. Stress factors during cattle slaughter. Ital. J. Food Saf. 2014, 3, 143–144. [Google Scholar] [CrossRef][Green Version]
- Rubio-González, A.; Potes, Y.; Illán-Rodríguez, D.; Vega-Naredo, I.; Sierra, V.; Caballero, B.; Fàbrega, E.; Velarde, A.; Dalmau, A.; Oliván, M.; et al. Effect of animal mixing as a stressor on biomarkers of autophagy and oxidative stress during pig muscle maturation. Animal 2015, 9, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Potes, Y.; Oliván, M.; Rubio-González, A.; de Luxán-Delgado, B.; Díaz, F.; Sierra, V.; Arroyo, L.; Peña, R.; Bassols, A.; González, J.; et al. Pig cognitive bias affects the conversion of muscle into meat by antioxidant and autophagy mechanisms. Animal 2017, 11, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Mato, A.; Salgado, F.J.; López-Pedrouso, M.; Carrera, M.; Bravo, S.; Parrado, M.; Gallardo, J.M.; Zapata, C. Tackling proteome changes in the Longissimus thoracis bovine muscle in response to pre-slaughter stress. J. Proteomics 2015, 122, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Luis, A.; Díaz, F.; Diñeiro, Y.; González-Blanco, L.; Arias, E.; Coto-Montes, A.; Oliván, M.; Sierra, V. Nuevos indicadores de carnes (DFD): Estrés oxidativo, autofagia y apoptosis. ITEA 2020, 117, 3–18. [Google Scholar] [CrossRef]
- Fuente-García, C.; Aldai, N.; Sentandreu, E.; Oliván, M.; Franco, D.; García-Torres, S.; Barron, L.J.R.; Sentandreu, M.Á. Caspase activity in post-mortem muscle and its relation to cattle handling practices. J. Sci. Food Agric. 2021, 101, 6258–6264. [Google Scholar] [CrossRef]
- García-Torres, S.; Cabeza de Vaca, M.; Tejerina, D.; Romero-Fernández, M.P.; Ortiz, A.; Franco, D.; Sentandreu, M.Á.; Oliván, M. Assessment of stress by serum biomarkers in calves and their relationship to ultimate pH as an indicator of meat quality. Animals 2021, 11, 2291. [Google Scholar] [CrossRef] [PubMed]
- Ouali, A.; Gagaoua, M.; Boudida, Y.; Becila, S.; Boudjellal, A.; Herrera-Mendez, C.; Sentandreu, M.Á. Biomarkers of meat tenderness: Present knowledge and perspectives in regard to our current understanding of the mechanisms involved. Meat Sci. 2013, 95, 854–870. [Google Scholar] [CrossRef]
- Picard, B.; Gagaoua, M. Chapter 11-proteomic investigations of beef tenderness. In Proteomics in Food Science: From Farm to Fork; Colgrave, M.L., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 177–197. [Google Scholar] [CrossRef]
- Gagaoua, M.; Terlouw, E.M.C.; Mullen, A.M.; Franco, D.; Warner, R.D.; Lorenzo, J.M.; Purslow, P.P.; Gerrard, D.; Hopkins, D.L.; Troy, D.; et al. Molecular signatures of beef tenderness: Underlying mechanisms based on integromics of protein biomarkers from multi-platform proteomics studies. Meat Sci. 2021, 172, 108311. [Google Scholar] [CrossRef] [PubMed]
- Gagaoua, M.; Warner, R.B.; Purslow, P.; Ramanathan, R.; Mullen, A.M.; López-Pedrouso, M.; Franco, D.; Lorenzo, J.M.; Tomasevic, I.; Picard, B.; et al. Dark-cutting beef: A brief review and an integromics meta-analysis at the proteome level to decipher the underlying pathways. Meat Sci. 2021, 181, 108611. [Google Scholar] [CrossRef]
- Warner, R.; Wheeler, T.L.; Ha, M.; Li, X.; Bekhit, A.E.D.; Morton, J.; Vaskoska, R.; Dunshea, F.; Liu, R.; Purslow, P.; et al. Meat tenderness: Advances in biology, biochemistry, molecular mechanisms and new technologies. Meat Sci. 2021, 108657. [Google Scholar] [CrossRef]
- Zapata, I.; Zerby, H.N.; Wick, M. Functional proteomic analysis predicts beef tenderness and the tenderness differential. J. Agric. Food Chem. 2009, 57, 4956–4963. [Google Scholar] [CrossRef] [PubMed]
- Sierra, V.; Fernández-Suárez, V.; Castro, P.; Osoro, K.; Vega-Naredo, I.; García-Macía, M.; Rodríguez-Colunga, P.; Coto-Montes, A.; Oliván, M. Identification of biomarkers of meat tenderisation and its use for early classification of Asturian beef into fast and late tenderising meat. J. Sci. Food Agric. 2012, 92, 2727–2740. [Google Scholar] [CrossRef]
- Fuente-Garcia, C.; Sentandreu, E.; Aldai, N.; Oliván, M.; Sentandreu, M.Á. Characterization of the myofibrillar proteome as a way to better understand differences in bovine meats having different ultimate pH values. J. Proteomics 2020, 20, 2000012. [Google Scholar] [CrossRef]
- Gagaoua, M. The path from protein profiling to biomarkers: The potential of proteomics and data integration in beef quality research. IOP Conf. Ser. Earth Environ. Sci. 2021, 854, 012029. [Google Scholar] [CrossRef]
- Piedrafita, J.; Quintanilla, R.; Sañudo, C.; Olleta, J.L.; Campo, M.M.; Panea, B.; Renand, G.; Turin, F.; Jabet, S.; Osoro, K.; et al. Carcass quality of ten beef cattle breeds of the south-west of europe. Livest. Prod. Sci. 2003, 82, 1–13. [Google Scholar] [CrossRef]
- Sierra, V.; Guerrero, L.; Fernández-Suárez, V.; Martínez, A.; Castro, P.; Osoro, K.; Rodríguez-Colunga, M.J.; Coto-Montes, A.; Oliván, M. Eating quality of beef from biotypes included in the PGI “Ternera Asturiana” showing distinct physicochemical characteristics and tenderization pattern. Meat Sci. 2010, 86, 343–351. [Google Scholar] [CrossRef]
- Mapama, I.G.P. Ternera Asturiana. Available online: https://www.mapa.gob.es/es/alimentacion/temas/calidad-diferenciada/dop-igp/carnes/IGP_Ternera_Asturiana.aspx (accessed on 7 August 2021).
- AMSA (American Meat Science Association). Meat Color Measurement Guidelines, Champaign, IL, USA, 2017. Available online: http://www.meatscience.org (accessed on 10 October 2021).
- Bjarnadóttir, S.G.; Hollung, K.; Faergestad, E.M.; Veiseth-Kent, E. Proteome changes in bovine Longissimus thoracis muscle during the first 48 h postmortem: Shifts in energy status and myofibrillar stability. J. Agric. Food Chem. 2010, 58, 7408–7414. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gagaoua, M.; Troy, D.; Mullen, A.M. The extent and rate of the appearance of the major 110 and 30 kDa proteolytic fragments during post-mortem aging of beef depend on the glycolysing rate of the muscle and aging time: An LC–MS/MS approach to decipher their proteome and associated pathways. J. Agric. Food Chem. 2021, 69, 602–614. [Google Scholar] [CrossRef]
- Vestergaard, M.; Oksbjerg, N.; Henckel, P. Influence of feeding intensity, grazing and finishing feeding on muscle fibre characteristics and meat colour of semitendinosus, longissimus dorsi and supraspinatus muscles of young bulls. Meat Sci. 2000, 54, 177–185. [Google Scholar] [CrossRef]
- Realini, C.E.; Duckett, S.K.; Brito, G.W.; dalla Rizza, M.; de Mattos, D. Effect of pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Sci. 2004, 66, 567–577. [Google Scholar] [CrossRef]
- Gagaoua, M.; Monteils, V.; Couvreur, S.; Picard, B. Identification of biomarkers associated with the rearing practices, carcass characteristics, and beef quality: An integrative approach. J. Agric. Food Chem. 2017, 65, 8264–8278. [Google Scholar] [CrossRef]
- Vestergaard, M.; Therkildsen, M.; Henckel, P.; Jensen, L.R. Influence of feeding intensity, grazing and finishing feeding on meat and eating quality of young bulls and the relationship between muscle fibre characteristics, fibre fragmentation and meat tenderness. Meat Sci. 2000, 54, 187–195. [Google Scholar] [CrossRef]
- Nuernberg, K.; Dannenberger, D.; Nuernberg, G.; Ender, K.; Voigt, J.; Scollan, N.D.; Wood, J.D.; Nute, G.R.; Richardson, R.I. Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of Longissimus muscle in different cattle breeds. Livest. Prod. Sci. 2005, 94, 137–147. [Google Scholar] [CrossRef]
- Laville, E.; Sayd, T.; Morzel, M.; Blinet, S.; Chambon, C.; Lepetit, J.; Renand, G.; Hocquette, J.F. Proteome changes during meat aging in tough and tender beef suggest the importance of apoptosis and protein solubility for beef aging and tenderization. J. Agric. Food Chem. 2009, 57, 10755–10764. [Google Scholar] [CrossRef]
- Paulsen, G.; Vissing, K.; Kalhovde, J.M.; Ugelstad, I.; Bayer, M.L.; Kadi, F.; Schjerling, P.; Hallén, J.; Raastad, T. Maximal eccentric exercise induces a rapid accumulation of small heat shock proteins on myofibrils and a delayed HSP70 response in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, 844–853. [Google Scholar] [CrossRef] [PubMed]
- González-Blanco, L.; Diñeiro, Y.; Díaz-Luis, A.; Coto-Montes, A.; Oliván, M.; Sierra, V. Impact of extraction method on the detection of quality biomarkers in normal vs. DFD meat. Foods 2021, 10, 1097. [Google Scholar] [CrossRef]
- Ho, C.Y.; Stromer, M.H.; Robson, R.M. Identification of the 30 kDa polypeptide in post mortem skeletal muscle as a degradation product of troponin-T. Biochimie 1994, 76, 369–375. [Google Scholar] [CrossRef]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness—An enzymatic view. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef]
- Picard, B.; Gagaoua, M. Meta-proteomics for the discovery of protein biomarkers of beef tenderness: An overview of integrated studies. Food Res. Int. 2020, 127, 108739. [Google Scholar] [CrossRef] [PubMed]
- Flashman, E.; Redwood, C.; Moolman-Smook, J.; Watkins, H. Cardiac myosin binding protein C: Its role in physiology and disease. Circ. Res. 2004, 94, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, N.; Bonnet, M.; Jurie, C.; Picard, B. Functional analysis of beef tenderness. J. Proteomics 2011, 75, 352–365. [Google Scholar] [CrossRef]
- Gagaoua, M.; Hughes, J.; Terlouw, E.M.C.; Warner, R.D.; Purslow, P.P.; Lorenzo, J.M.; Picard, B. Proteomic biomarkers of beef colour. Trends Food Sci. Technol. 2020, 101, 234–252. [Google Scholar] [CrossRef]
- Wu, G.; Clerens, S.; Farouk, M.M. LC MS/MS identification of large structural proteins from bull muscle and their degradation products during post mortem storage. Food Chem. 2014, 150, 137–144. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Hollung, K.; Therkildsen, M.; Hildrum, K.I.; Bendixen, E. Proteome analysis of early post-mortem changes in two bovine muscle types: M. longissimus dorsi and M. semitendinosis. J. Proteom. 2006, 6, 936–944. [Google Scholar] [CrossRef]
- Sawdy, J.C.; Kaiser, S.A.; St-Pierre, N.R.; Wick, M.P. Myofibrillar 1-D fingerprints and myosin heavy chain MS analyses of beef loin at 36 h postmortem correlate with tenderness at 7 days. Meat Sci. 2004, 67, 421–426. [Google Scholar] [CrossRef]
- Morzel, M.; Terlouw, C.; Chambon, C.; Micol, D.; Picard, B. Muscle proteome and meat eating qualities of Longissimus thoracis of “blonde d’Aquitaine” young bulls: A central role of HSP27 isoforms. Meat Sci. 2008, 78, 297–304. [Google Scholar] [CrossRef]
- Zhou, Q.; Chu, P.H.; Huang, C.; Cheng, C.F.; Martone, M.E.; Knoll, G.; Shelton, G.D.; Evans, S.; Chen, J. Ablation of Cypher, a PDZ-LIM domain Z-line protein, causes a severe form of congenital myopathy. J. Cell Biol. 2001, 155, 605–612. [Google Scholar] [CrossRef]
- Shathasivam, T.; Kislinger, T.; Gramolini, A.O. Genes, proteins and complexes: The multifaceted nature of FHL family proteins in diverse tissues. J. Cell Mol. Med. 2021, 14, 2702–2720. [Google Scholar] [CrossRef] [PubMed]
- Morzel, M.; Chambon, C.; Hamelin, M.; Sante-Lhoutellier, V.; Sayd, T.; Monin, G. Proteome changes during pork meat ageing following use of two different pre-slaughter handling procedures. Meat Sci. 2004, 67, 689–696. [Google Scholar] [CrossRef]
- Gagaoua, M.; Bonnet, M.; Ellies-Oury, M.P.; de Koning, L.; Picard, B. Reverse phase protein arrays for the identification/validation of biomarkers of beef texture and their use for early classification of carcasses. Food Chem. 2018, 250, 245–252. [Google Scholar] [CrossRef]
- Hughes, J.; Clarke, F.; Li, Y.; Purslow, P.; Warner, R. Differences in light scattering between pale and dark beef Longissimus thoracis muscles are primarily caused by differences in the myofilament lattice, myofibril and muscle fibre transverse spacings. Meat Sci. 2019, 149, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Oliván, M.; Fernández-Suárez, V.; Díaz-Martínez, F.; Sierra, V.; Coto-Montes, A.; Luxán-Delgado, B.; Peña, R.; Bassols, A.; Fàbrega, E.; Dalmau, A.; et al. Identification of biomarkers of stress in meat of pigs managed under different mixing treatments. Br. Biotechnol. J. 2016, 11, 1–13. [Google Scholar] [CrossRef]
- Yu, J.; Tang, S.; Bao, E.; Zhang, M.; Hao, Q.; Yue, Z. The effect of transportation on the expression of heat shock proteins and meat quality of M. longissimus dorsi in pigs. Meat Sci. 2009, 83, 474–478. [Google Scholar] [CrossRef]
- Ma, D.; Kim, Y.H.B. Proteolytic changes of myofibrillar and small heat shock proteins in different bovine muscles during aging: Their relevance to tenderness and Water-holding capacity. Meat Sci. 2020, 163, 108090. [Google Scholar] [CrossRef]
- Lomiwes, D.; Hurst, S.M.; Dobbie, P.; Frost, D.A.; Hurst, R.D.; Young, O.A.; Farouk, M.M. The protection of bovine skeletal myofibrils from proteolytic damage post-mortem by small heat shock proteins. Meat Sci. 2014, 97, 548–557. [Google Scholar] [CrossRef]
- Balan, P.; Kim, Y.H.; Blijenburg, R. Small heat shock protein degradation could be an indicator of the extent of myofibrillar protein degradation. Meat Sci. 2014, 97, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.; Penick, M.L.; Waddell, J.N.; Bidwell, C.A.; Kim, Y.H.B. A new insight into meat toughness of callipyge lamb loins—The relevance of anti-apoptotic systems to decreased proteolysis. Meat Sci. 2018, 140, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Hildrum, K.I.; Westad, F.; Kummen, E.; Aass, L.; Hollung, K. Changes in enzymes associated with energy metabolism during the early post-mortem period in Longissimus thoracis bovine muscle analyzed by proteomics. J. Proteome Res. 2006, 5, 1763–1769. [Google Scholar] [CrossRef]
- Jia, X.H.; Ekman, M.; Grove, H.; Faergestad, E.M.; Aass, L.; Hildrum, K.I.; Hollung, K. Proteome changes in bovine Longissimus thoracis muscle during the early post-mortem storage period. J. Proteome Res. 2007, 6, 2720–2731. [Google Scholar] [CrossRef] [PubMed]
- Canto, A.C.; Suman, S.P.; Nair, M.N.; Li, S.; Rentfrow, G.; Beach, C.M.; Silva, T.J.; Wheeler, T.L.; Shackelford, S.D.; Grayson, A.; et al. Differential abundance of sarcoplasmic proteome explains animal effect on beef Longissimus lumborum color stability. Meat Sci. 2015, 102, 90–98. [Google Scholar] [CrossRef]
- Wu, W.; Gao, X.G.; Dai, Y.; Fu, Y.; Li, X.M.; Dai, R.T. Post-mortem changes in sarcoplasmic proteome and its relationship to meat color traits in M. semitendinosus of Chinese Luxi yellow cattle. Food Res. Int. 2015, 72, 98–105. [Google Scholar] [CrossRef]
- Wu, W.; Yu, Q.Q.; Fu, Y.; Tian, X.J.; Jia, F.; Li, X.M.; Dai, R.T. Towards muscle specific meat color stability of Chinese Luxi yellow cattle: A proteomic insight into post-mortem storage. J. Proteomics 2016, 147, 108–118. [Google Scholar] [CrossRef]
- Hughes, J.; Clarke, F.; Purslow, P.; Warner, R. High pH in beef longissimus thoracis reduces muscle fibre transverse shrinkage and light scattering which contributes to the dark colour. Food Res. Int. 2017, 101, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Wu, W.; Tian, X.; Jia, F.; Xu, L.; Dai, R.; Li, X. Comparative proteomics to reveal muscle-specific beef color stability of Holstein cattle during post-mortem storage. Food Chem. 2017, 229, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Zhang, X.Z.; Wang, T.T.; Hopkins, D.L.; Mao, Y.W.; Liang, R.R.; Yang, G.F.; Luo, X.; Zhu, L.X. Implications of step-chilling on meat color investigated using proteome analysis of the sarcoplasmic protein fraction of beef Longissimus lumborum muscle. J. Integr. Agric. 2018, 17, 2118–2125. [Google Scholar] [CrossRef]
- Mahmood, S.; Turchinsky, N.; Paradis, F.; Dixon, W.T.; Bruce, H.L. Proteomics of dark cutting Longissimus thoracis muscle from heifer and steer carcasses. Meat Sci. 2018, 137, 47–57. [Google Scholar] [CrossRef]
- Gagaoua, M.; Terlouw, E.M.C.; Picard, B. The study of protein biomarkers to understand the biochemical processes underlying beef color development in young bulls. Meat Sci. 2017, 134, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P.; Warner, R.D.; Clarke, F.M.; Hughes, J.M. Variations in meat colour due to factors other than myoglobin chemistry; a synthesis of recent findings (invited review). Meat Sci. 2020, 159, 107941. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Oiseth, S.K.; Purslow, P.P.; Warner, R.D. A structural approach to understanding the interactions between colour, water-holding capacity and tenderness. Meat Sci. 2014, 98, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P.; Gagaoua, M.; Warner, R.D. Insights on meat quality from combining traditional studies and proteomics. Meat Science. 2021, 174, 108423. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm Management (F) | Transport and Lairage (TL) | Significance | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Quality Traits | I | E | SEM | NM | M | SEM | F | TL | F × TL |
| pH | 5.6 | 5.46 | 0.05 | 5.49 | 5.58 | 0.05 | NS | NS | NS |
| L* | 41.86 | 38.20 | 0.99 | 38.73 | 41.33 | 0.99 | * | NS | NS |
| a* | 12.03 | 10.82 | 0.523 | 10.4 | 12.45 | 0.523 | NS | * | * |
| b* | 15.49 | 10.28 | 0.522 | 12.19 | 13.58 | 0.522 | *** | NS | NS |
| C* | 19.64 | 14.97 | 0.681 | 16.13 | 18.49 | 0.681 | *** | * | * |
| h* | 52.49 | 43.61 | 1.04 | 49.07 | 47.03 | 1.04 | *** | NS | NS |
| Farm Management (F) | Transport and Lairage (TL) | Post-Mortem Time (t) | Significance | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Band [MWe] | I | E | SEM | NM | M | SEM | 2 h | 8 h | 24 h | 3 d | 7 d | 14 d | SEM | F | TL | t | F × TL | F × t | TL × t | F × TL× t |
| M1 (155.03) | 4.66 | 3.67 | 0.25 | 4.76 | 3.87 | 0.15 | 4.08 | 4.39 | 4.22 | 4.4 | 4.36 | 3.57 | 0.24 | * | ** | NS | NS | NS | NS | NS |
| M4 (99.16) | 5.94 | 4.97 | 0.29 | 5.48 | 5.43 | 0.17 | 6.04 b | 5.99 b | 5.71 ab | 5.03ab | 5.03ab | 4.93 a | 0.28 | NS | NS | ** | NS | NS | NS | NS |
| M6 (83.56) | 2.18 | 2.0 | 0.09 | 2.102 | 2.08 | 0.06 | 2.05 ab | 2.24 b | 2.28 b | 2.13 b | 2.07 ab | 1.78 a | 0.09 | NS | NS | ** | NS | NS | NS | NS |
| M9 (71.02) | 1.23 | 1.36 | 0.06 | 1.27 | 1.32 | 0.04 | 1.37 b | 1.43 b | 1.33 b | 1.37 b | 1.21 a | 1.05 a | 0.06 | NS | NS | *** | NS | NS | NS | NS |
| M12 (59.93) | 2.63 | 2.22 | 0.15 | 2.63 | 2.22 | 0.09 | 2.35 | 2.53 | 2.61 | 2.60 | 2.35 | 2.12 | 0.15 | NS | ** | NS | *** | NS | NS | NS |
| M13 (56.24) | 2.42 | 2.49 | 0.09 | 2.47 | 2.44 | 0.05 | 2.47 ab | 2.70 b | 2.59 b | 2.37 ab | 2.37 ab | 2.25 a | 0.09 | NS | NS | ** | NS | NS | NS | NS |
| M15 (50.74) | 1.5 | 1.57 | 0.06 | 1.49 | 1.58 | 0.04 | 1.47 ab | 1.62 b | 1.62 b | 1.64 b | 1.52 ab | 1.34 a | 0.06 | NS | NS | ** | * | NS | NS | NS |
| M17 (40.53) | 16.14 | 19.3 | 0.79 | 17.64 | 17.8 | 0.47 | 19.19 ab | 16.23 a | 17.08 a | 16.93 a | 16.91 a | 19.98 b | 0.76 | * | NS | ** | NS | NS | NS | NS |
| M18 (37.64) | 2.96 | 2.98 | 0.12 | 2.96 | 2.98 | 0.07 | 3.27 bc | 3.49 c | 3.38 c | 2.82 ab | 2.52 a | 2.34 a | 0.12 | NS | NS | *** | NS | NS | NS | NS |
| M19 (36.88) | 2.34 | 1.94 | 0.07 | 2.24 | 2.05 | 0.04 | 2.61 c | 2.73 c | 2.67 c | 1.99 b | 1.49 a | 1.38 a | 0.07 | ** | ** | *** | NS | ** | NS | NS |
| M20 (35.03) | 7.47 | 7.04 | 0.22 | 7.28 | 7.23 | 0.13 | 7.09 abc | 7.58 bc | 8.02 c | 7.68 b | 6.84 ab | 6.33 a | 0.21 | NS | NS | *** | NS | NS | NS | NS |
| M21 (32.95) | 6.1 | 5.9 | 0.17 | 6.01 | 5.99 | 0.1 | 5.26 a | 5.7 ab | 5.74 ab | 6.56 c | 6.62 c | 6.1 c | 0.16 | NS | NS | *** | NS | NS | NS | NS |
| M23 (30.89) | 1.03 | 1.25 | 0.05 | 1.15 | 1.13 | 0.03 | 1.05 a | 1.04 a | 1.04 a | 1.06 a | 1.21 ab | 1.44 b | 0.05 | * | NS | *** | NS | NS | NS | NS |
| M24 (29.75) | 2.4 | 2.6 | 0.1 | 2.49 | 2.51 | 0.06 | 3.04 d | 2.89 d | 2.74 cd | 2.32 bc | 2.17 ab | 1.84 a | 0.10 | NS | NS | *** | *** | NS | NS | NS |
| M25 (28.67) | 1.58 | 1.71 | 0.08 | 1.61 | 1.68 | 0.05 | 0.89 a | 0.92 a | 0.97 a | 1.68 b | 2.51 c | 2.89 d | 0.08 | NS | NS | *** | NS | * | NS | NS |
| M26 (27.82) | 0.65 | 0.8 | 0.03 | 0.71 | 0.73 | 0.02 | 0.54 a | 0.53 a | 0.51 a | 0.73 b | 0.95 c | 1.08 c | 0.03 | * | NS | *** | * | * | NS | NS |
| M27 (27.25) | 0.56 | 0.69 | 0.02 | 0.6 | 0.65 | 0.01 | 0.47 a | 0.45 a | 0.45 a | 0.62 b | 0.83 c | 0.93 d | 0.02 | ** | * | *** | NS | NS | NS | NS |
| M30 (25.10) | 1.22 | 1.33 | 0.06 | 1.35 | 1.2 | 0.04 | 1.33 bc | 1.37 bc | 1.42 c | 1.24 abc | 1.16 ab | 1.11 a | 0.06 | NS | ** | ** | *** | NS | NS | NS |
| M31 (23.20) | 5.53 | 6.04 | 0.13 | 5.61 | 5.96 | 0.08 | 5.81 | 5.77 | 5.59 | 5.69 | 5.86 | 5.97 | 0.12 | * | ** | NS | NS | NS | NS | NS |
| M32 (19.89) | 3.83 | 3.7 | 0.13 | 3.66 | 3.86 | 0.07 | 4.11 bc | 4.09 bc | 3.98 bc | 4.23 c | 3.64 b | 2.52 a | 0.12 | NS | ** | *** | NS | *** | NS | NS |
| M33 (19.07) | 1.17 | 1.22 | 0.08 | 1.2 | 1.18 | 0.05 | 0.79 a | 0.77 a | 0.79 a | 1.05 a | 1.54 b | 2.21 c | 0.08 | NS | NS | *** | NS | *** | NS | NS |
| M34 (17.54) | 3.84 | 3.92 | 0.11 | 3.96 | 3.8 | 0.07 | 3.82 | 3.81 | 3.78 | 3.86 | 3.94 | 4.06 | 0.13 | NS | NS | NS | * | NS | NS | NS |
| M35 (16.05) | 5.76 | 5.37 | 0.14 | 5.43 | 5.7 | 0.08 | 5.45 | 5.49 | 5.34 | 5.62 | 5.73 | 5.76 | 0.05 | NS | * | NS | NS | NS | NS | NS |
| Band: Gene Name | Identification | Accession Number | MOWSE Scores | Sequence Coverage (%) | Matched Queries | MWt |
|---|---|---|---|---|---|---|
| M1: MYBPC2 | Myosin-binding protein C, fast-type isoform X2 | E1BNV1 | 295 | 24 | 25 | 128.5 |
| M4: ACTN3 | Alpha-actinin-3 | Q0III9 | 506 | 38 | 35 | 103.7 |
| M6: PFKM | ATP-dependent 6-phosphofructokinase, muscle type | Q0IIG5 | 352 | 40 | 42 | 86.1 |
| M9: HSPA1A | Heat Shock 70 kDa protein 1A | Q27975 | 288 | 38 | 24 | 70.5 |
| M12: PKM | Pyruvate kinase PKM, isoform X1 | A5D984 | 822 | 66 | 46 | 58.5 |
| M13: DES | Desmin, partial | O62654 | 246 | 54 | 21 | 52.6 |
| M15: ATP5F1B | ATP synthase subunit beta, mitochondrial precursor | P00829 | 445 | 48 | 25 | 56.2 |
| M17: ACTA1 | Actin, alpha skeletal muscle | P68138 | 522 | 52 | 24 | 42.4 |
| M18: ALDOA | Fructose-biphosphate aldolase A | A6QLL8 | 430 | 62 | 24 | 39.9 |
| M19: ALDOA | Fructose-biphosphate aldolase A | A6QLL8 | 286 | 60 | 21 | 39.9 |
| M20: GAPDH | Glyceraldehide-3-phosphate dehydrogenase | P10096 | 394 | 45 | 20 | 36.1 |
| M21: TPM1 | Tropomyosin alpha-1 chain | Q5KR49 | 199 | 41 | 15 | 32.7 |
| M23: LDB3 | LIM domain-binding protein 3 isoform X5 | G3N3C9 | 180 | 55 | 19 | 30.9 |
| M24: FHL1 | Four and a half LIM domains protein 1 isoform 1 | G3MZ95 | 671 | 87 | 34 | 35.5 |
| M25: TNNT3 | Troponin T, fast skeletal muscle isoform X31 | Q8MKI3 | 206 | 45 | 18 | 28.9 |
| M26: FHL2 | Four and a half LIM domains protein 1 isoform 2 | Q2KI95 | 95 | 37 | 11 | 33.8 |
| M27: TNNT3 | Troponin T, fast skeletal muscle isoform X31 | Q8MKI3 | 178 | 43 | 15 | 28.9 |
| M30: HSPB1 | Heat Shock protein family B member 1 variant 1 | Q3T149 | 368 | 73 | 14 | 22.4 |
| M31: MYL1 | Myosin light chain 1/3 skeletal muscle isoform | A0JNJ5 | 425 | 77 | 18 | 21.1 |
| M32: TNNI2 | Troponin I, fast skeletal muscle | F6QIC1 | 194 | 70 | 23 | 21.6 |
| M33: CRYAB | Alpha-crystallin B chain | P02510 | 163 | 69 | 13 | 20.1 |
| M34: TNNC1 | Troponin C, skeletal muscle | P63315 | 333 | 56 | 15 | 18.3 |
| M35: MYLPF | Myosin regulatory light chain 2, skeletal muscle isoform | Q0P571 | 517 | 66 | 21 | 19.11 |
| PFKM | HSPA1A | PKM | DES | ALDOA M18 | ALDOA M19 | GAPDH | TPM1 | LDB3 | FHL1 | TNNT3 M25 | TNNT3 M27 | HSPB1 | TNNI2 | CRYAB | MYLPF | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L*(60) | 2 h | 0.691 * | −0.654 * | −0.651 * | 0.624 * | 0.702 * | |||||||||||
| 8 h | 0.731 ** | 0.613 * | 0.608 * | −0.581 * | 0.604 * | ||||||||||||
| 24 h | 0.615 * | 0.697 * | |||||||||||||||
| 3 d | −0.687 * | 0.768 ** | −0.630* | 0.610 * | |||||||||||||
| 7 d | −0.652 * | −0.588 * | 0.583 * | −0.777 ** | .578 * | ||||||||||||
| 14 d | 0.599 * | .633* | 0.615 * | ||||||||||||||
| a*(60) | 2 h | −0.695 * | −0.683 * | −0.593 * | |||||||||||||
| 8 h | −0.588 * | −0.850 ** | |||||||||||||||
| 24 h | −0.694 * | −0.675 * | |||||||||||||||
| 3 d | −0.726 * | 0.685 * | −0.581 * | −0.707 * | |||||||||||||
| 7 d | 0.821 ** | 0.701 * | −0.592 * | −0.686* | −0.611 * | ||||||||||||
| 14 d | −0.585 * | −0.632 * | |||||||||||||||
| b*(60) | 2 h | 0.645 * | −0.633 * | 0.608 * | |||||||||||||
| 8 h | 0.612 * | 0.606 * | 0.627 * | ||||||||||||||
| 24 h | 0.588 * | 0.683 * | |||||||||||||||
| 3 d | −0.664 * | −0.624 * | 0.634 * | −0.597 * | |||||||||||||
| 7 d | 0.581* | −0.650 * | −0.738 ** | ||||||||||||||
| 14 d | |||||||||||||||||
| C*(60) | 2 h | −0.605 * | −0.584 * | −0.592 * | |||||||||||||
| 8 h | −0.613 * | −0.644 * | |||||||||||||||
| 24 h | −0.714 * | −0.599 * | |||||||||||||||
| 3 d | −0.637 * | 0.651 * | |||||||||||||||
| 7 d | 0.666 * | −0.634* | |||||||||||||||
| 14 d | |||||||||||||||||
| h* (60) | 2 h | 0.592 * | 0.588 * | −0.617 * | −0.599 * | 0.741 ** | |||||||||||
| 8 h | 0.700 * | 0.800 ** | |||||||||||||||
| 24 h | 0.670 * | 0.610 * | 0.578 * | ||||||||||||||
| 3 d | 0.676 * | 0.755 ** | 0.721 ** | ||||||||||||||
| 7 d | −0.695 * | −0.751 ** | 0.626 * | −0.641 * | 0.654 * | ||||||||||||
| 14 d | 0.689 * | 0.618 * |
| ACTN3 | HSPA1A | PKM | DES | ATP5F1B | ACTA1 | ALDOA M18 | ALDOA M19 | GAPDH | FHL1 | TNNT3 M25 | FHL2 | TNNT3 M27 | MYL1 | TNNI2 | CRYAB | TNNC1 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WB3 d | 2 h | −0.482 * | −0.427 * | −0.437 * | 0.539 ** | 0.415 * | 0.497 * | 0.423 * | ||||||||||
| 8 h | −0.493 * | 0.439 * | ||||||||||||||||
| 24 h | 0.415 * | 0.472 * | −0.545 ** | 0.428 * | ||||||||||||||
| 3 d | 0.495 * | |||||||||||||||||
| 7 d | −0.450 * | |||||||||||||||||
| 14 d | −0.521 ** | −0.417 * | ||||||||||||||||
| WB7 d | 2 h | 0.556 ** | 0.503 * | |||||||||||||||
| 8 h | −0.411 * | 0.529 ** | 0.516 ** | |||||||||||||||
| 24 h | −0.502 * | 0.453 * | ||||||||||||||||
| 3 d | 0.472 * | |||||||||||||||||
| 7 d | −0.451 * | |||||||||||||||||
| 14 d | 0.447 * | −0.599 ** | 0.413 * | 0.514 * | ||||||||||||||
| WB14 d | 2 h | −0.441 * | 0.514 * | 0.483 * | 0.417 * | |||||||||||||
| 8 h | −0.524 ** | 0.625 ** | 0.503 * | 0.405 * | 0.441 * | |||||||||||||
| 24 h | −0.421 * | 0.445 * | 0.456 * | |||||||||||||||
| 3 d | 0.542 ** | |||||||||||||||||
| 7 d | 0.521 * | |||||||||||||||||
| 14 d | −0.567 ** | 0.429 * | 0.614 ** | |||||||||||||||
| % TR. 3–14 d | 2 h | 0.592 ** | ||||||||||||||||
| 8 h | ||||||||||||||||||
| 24 h | −0.505 * | |||||||||||||||||
| 3 d | 0.442 * | −0.491 * | −0.564 ** | |||||||||||||||
| 7 d | −0.546 ** | −0.538 ** | ||||||||||||||||
| 14 d | 0.453 * | −0.557 ** | −0.469 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sierra, V.; González-Blanco, L.; Diñeiro, Y.; Díaz, F.; García-Espina, M.J.; Coto-Montes, A.; Gagaoua, M.; Oliván, M. New Insights on the Impact of Cattle Handling on Post-Mortem Myofibrillar Muscle Proteome and Meat Tenderization. Foods 2021, 10, 3115. https://doi.org/10.3390/foods10123115
Sierra V, González-Blanco L, Diñeiro Y, Díaz F, García-Espina MJ, Coto-Montes A, Gagaoua M, Oliván M. New Insights on the Impact of Cattle Handling on Post-Mortem Myofibrillar Muscle Proteome and Meat Tenderization. Foods. 2021; 10(12):3115. https://doi.org/10.3390/foods10123115
Chicago/Turabian StyleSierra, Verónica, Laura González-Blanco, Yolanda Diñeiro, Fernando Díaz, María Josefa García-Espina, Ana Coto-Montes, Mohammed Gagaoua, and Mamen Oliván. 2021. "New Insights on the Impact of Cattle Handling on Post-Mortem Myofibrillar Muscle Proteome and Meat Tenderization" Foods 10, no. 12: 3115. https://doi.org/10.3390/foods10123115
APA StyleSierra, V., González-Blanco, L., Diñeiro, Y., Díaz, F., García-Espina, M. J., Coto-Montes, A., Gagaoua, M., & Oliván, M. (2021). New Insights on the Impact of Cattle Handling on Post-Mortem Myofibrillar Muscle Proteome and Meat Tenderization. Foods, 10(12), 3115. https://doi.org/10.3390/foods10123115