
The Application of Bacillus subtilis for Adhesion Inhibition of Pseudomonas and Preservation of Fresh Fish




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Strains
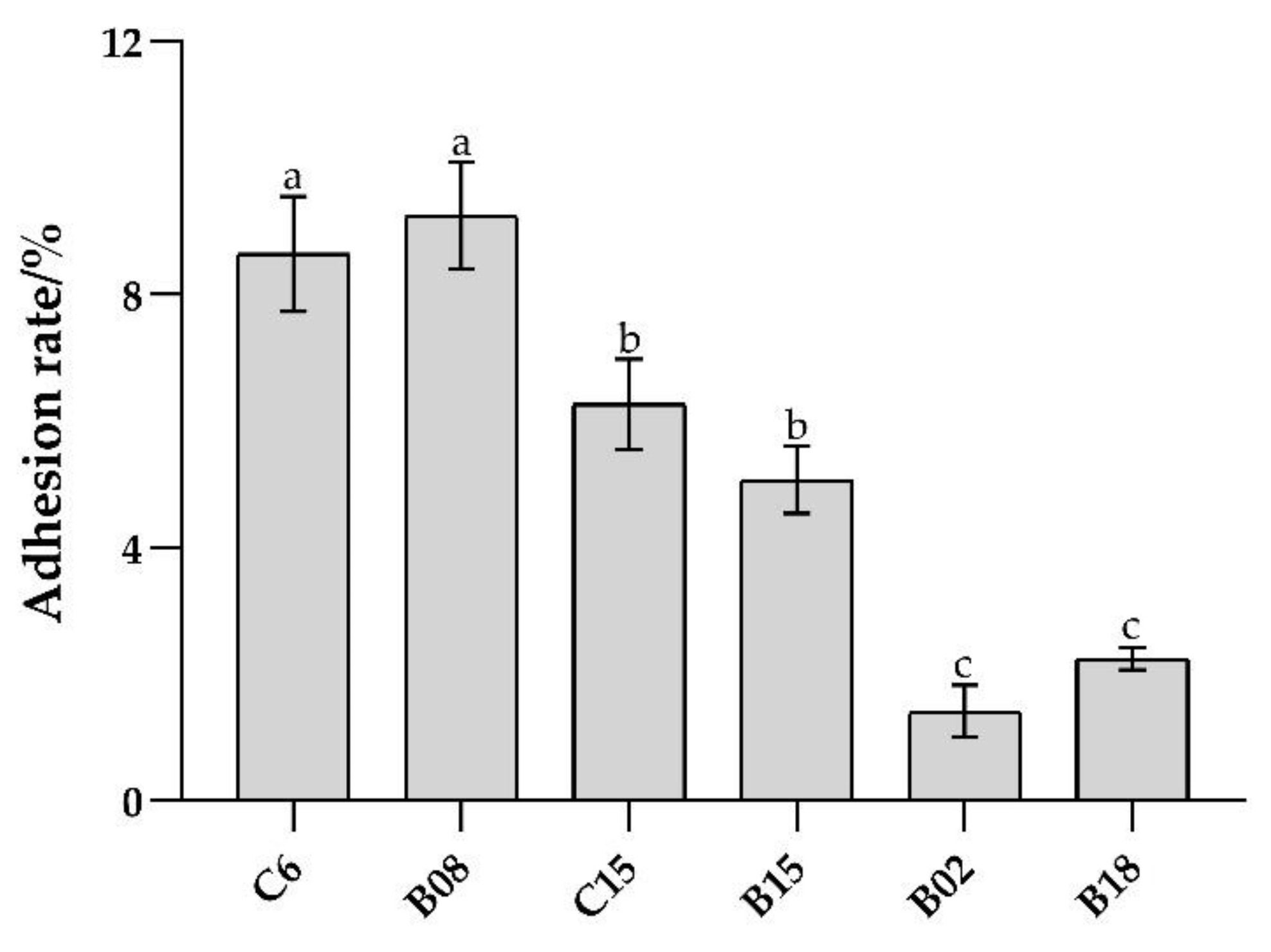
2.2. Determination of the Adhesion Ability of B. subtilis In Vitro
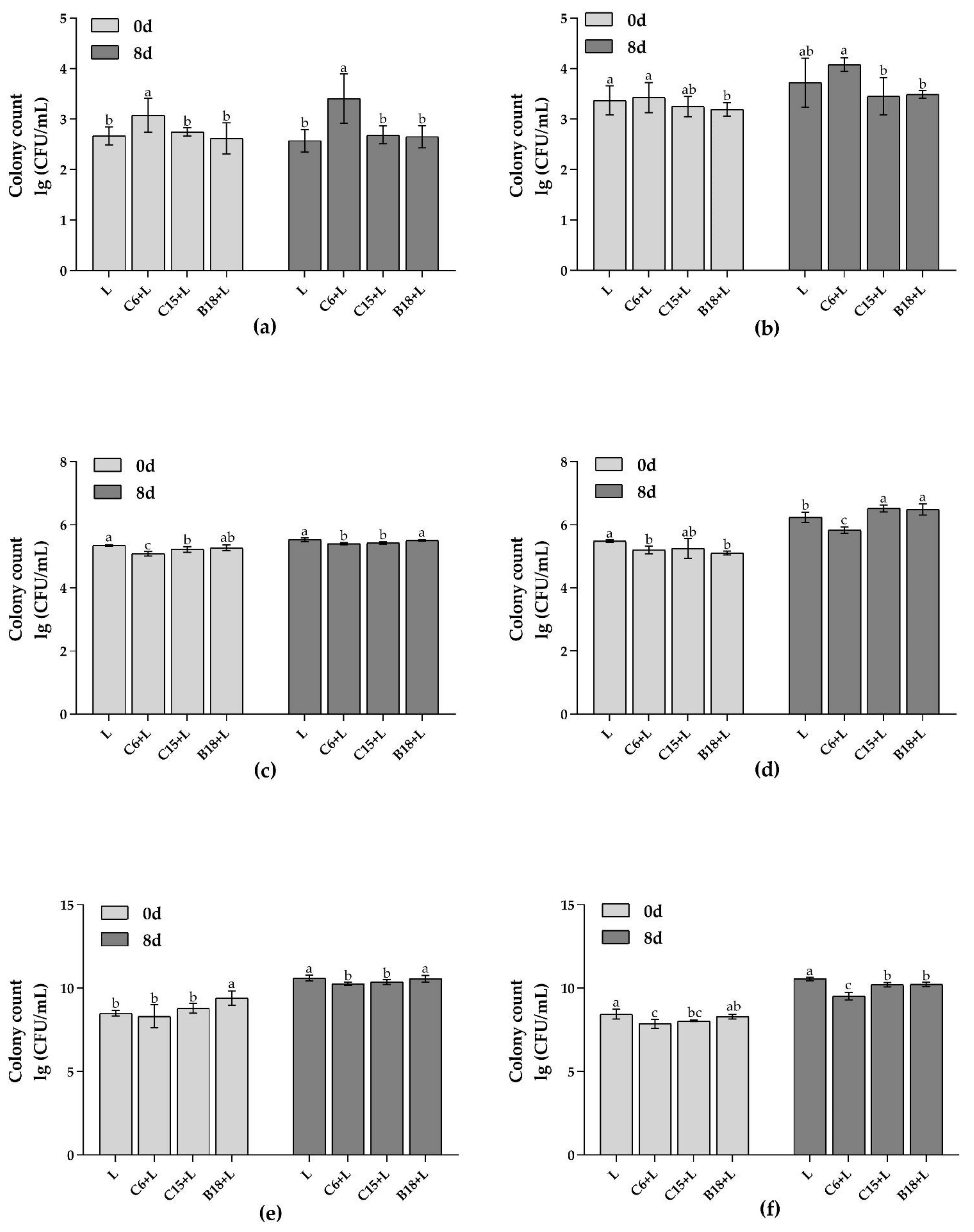
2.3. Determination of the Adhesion Inhibition Ability of B. subtilis to Pseudomonas spp.
2.4. In Vivo Adhesion Test
2.4.1. Feeding Experiment of Tilapia
2.4.2. Sample Collection and Processing
2.5. Determination of Total Volatile Basic Nitrogen (TVB-N)
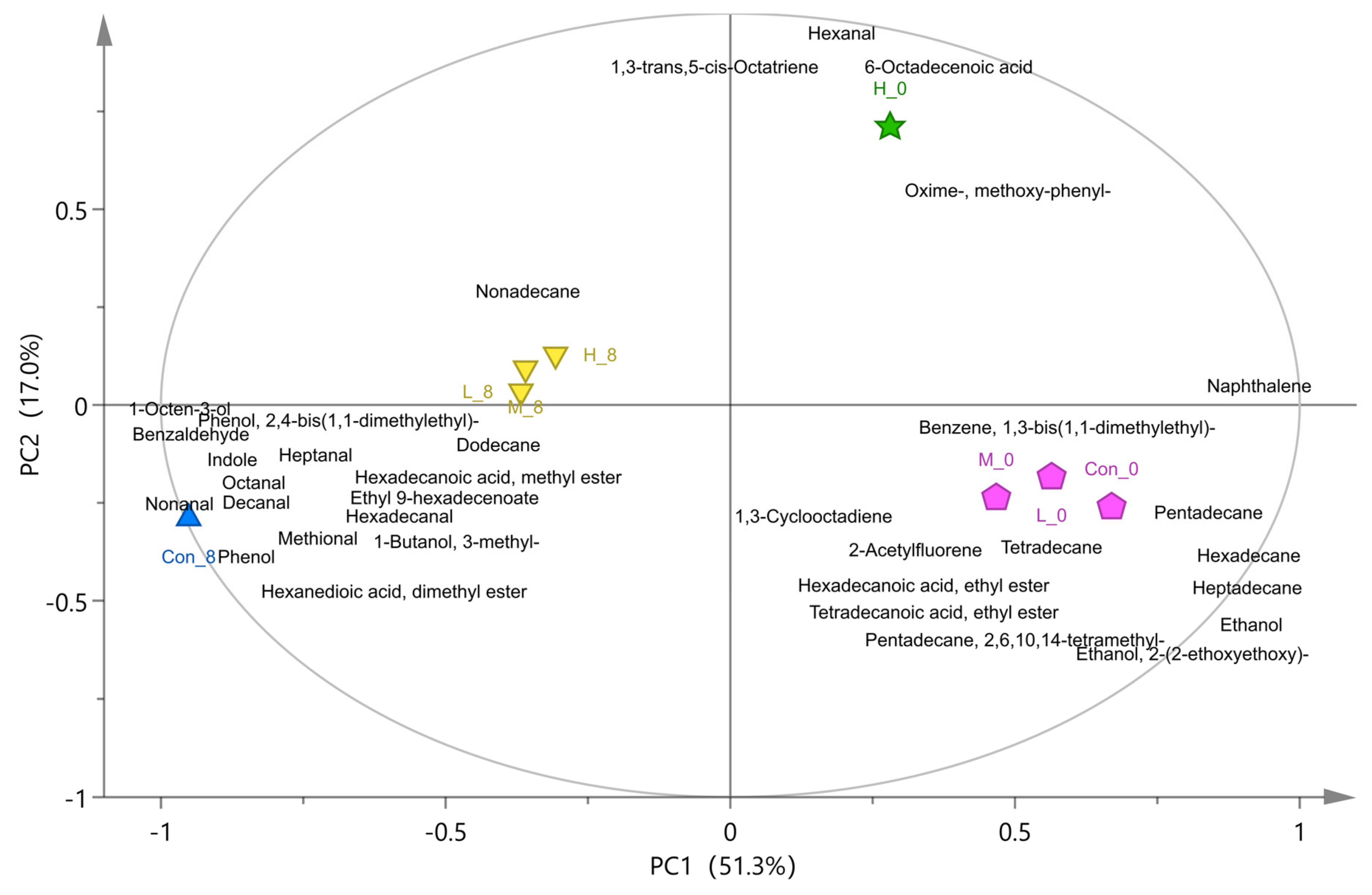
2.6. Determination of Volatile Flavor Compounds by GC-MS
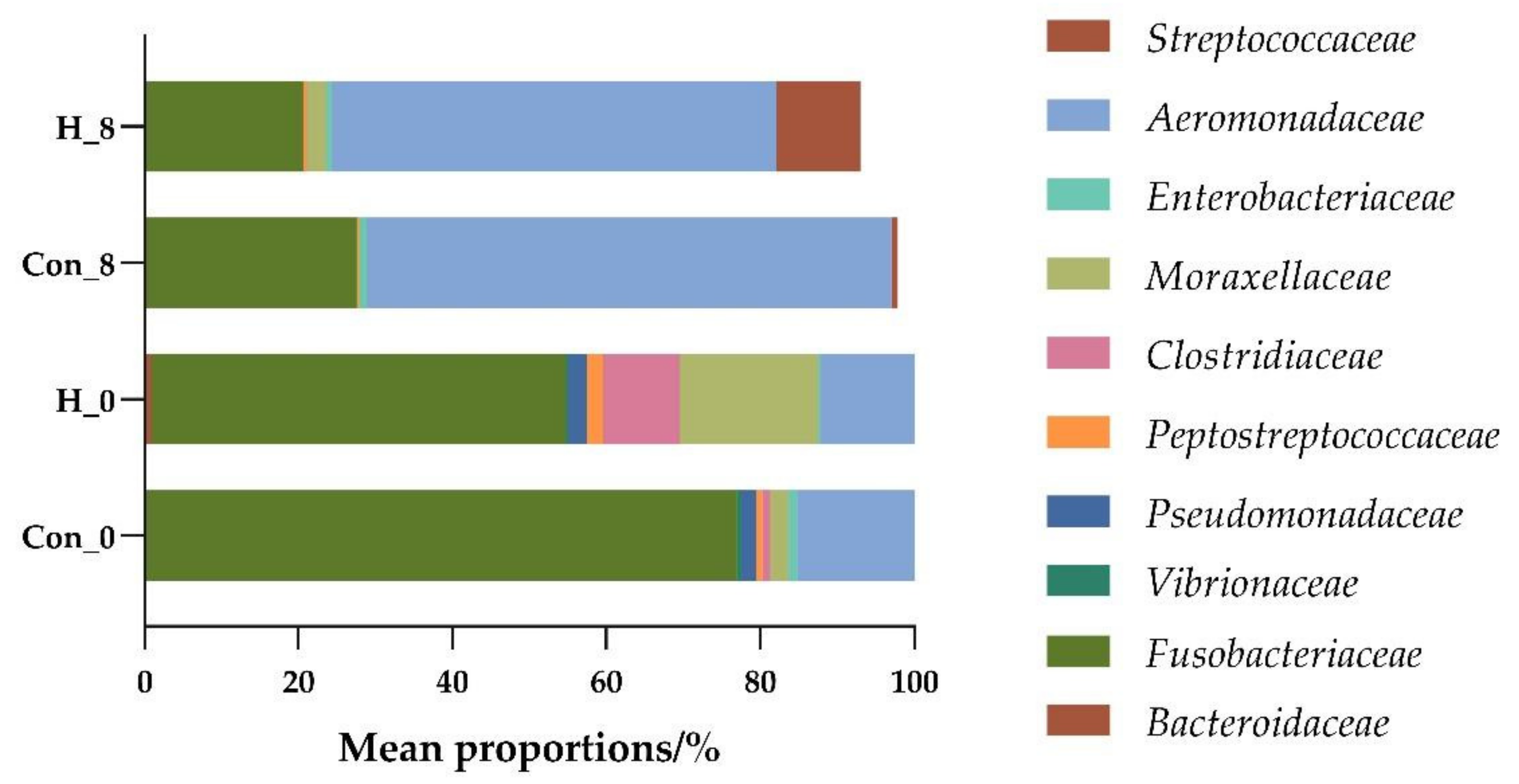
2.7. Analysis of Changes of Intestinal Microflora in Tilapia by High-Throughput Sequencing
2.8. Detection of Specific Microflora in Intestine and Flesh of Tilapia by Real-Time Quantitative PCR (qPCR)
2.9. Statistical Analysis
3. Results
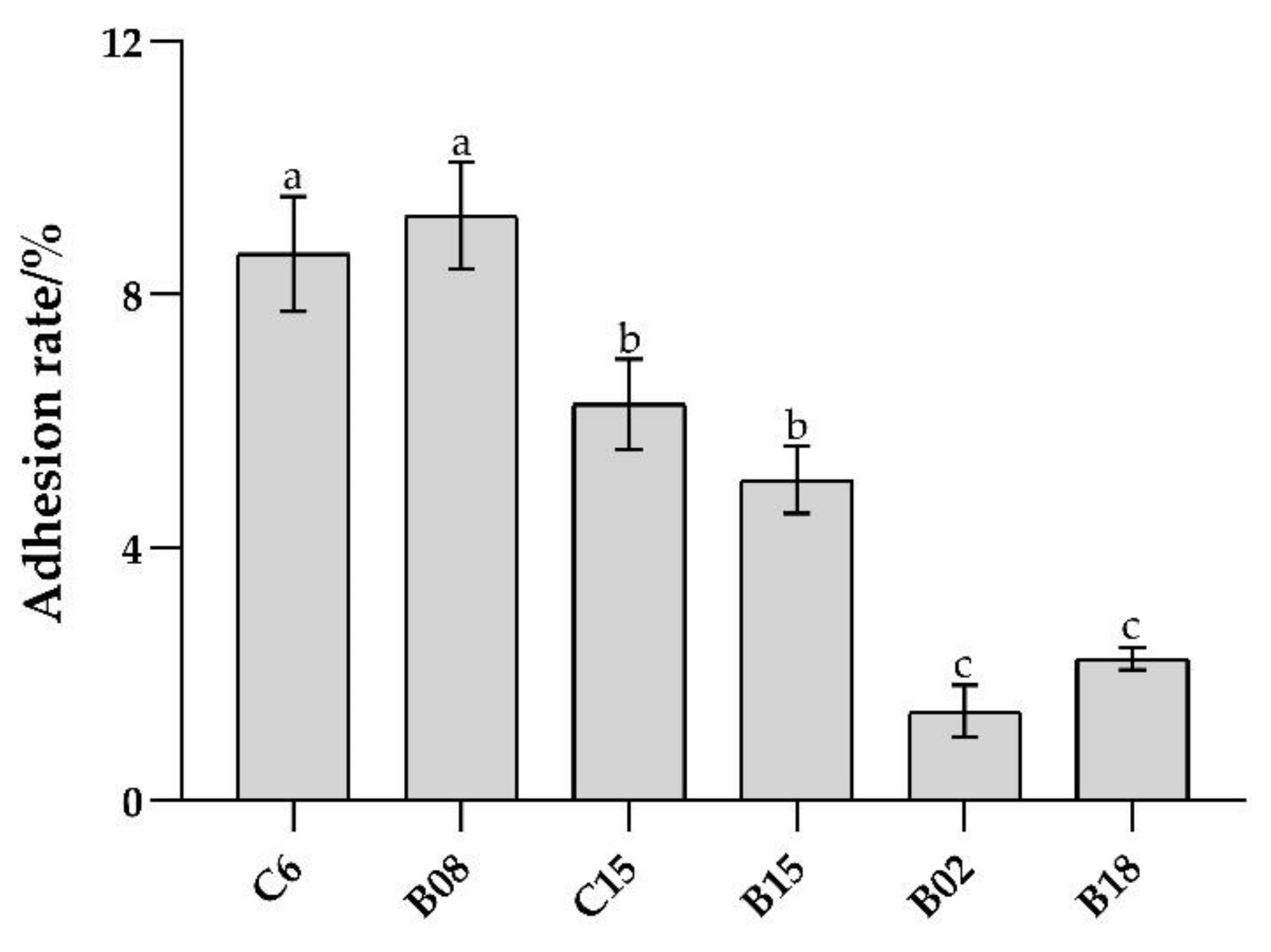
3.1. Adhesion Ability of B. subtilis In Vitro and Adhesion Inhibition to Pseudomonas spp.
3.2. In Vivo Adhesion of B. subtilis and the Preservation Effect on Tilapia
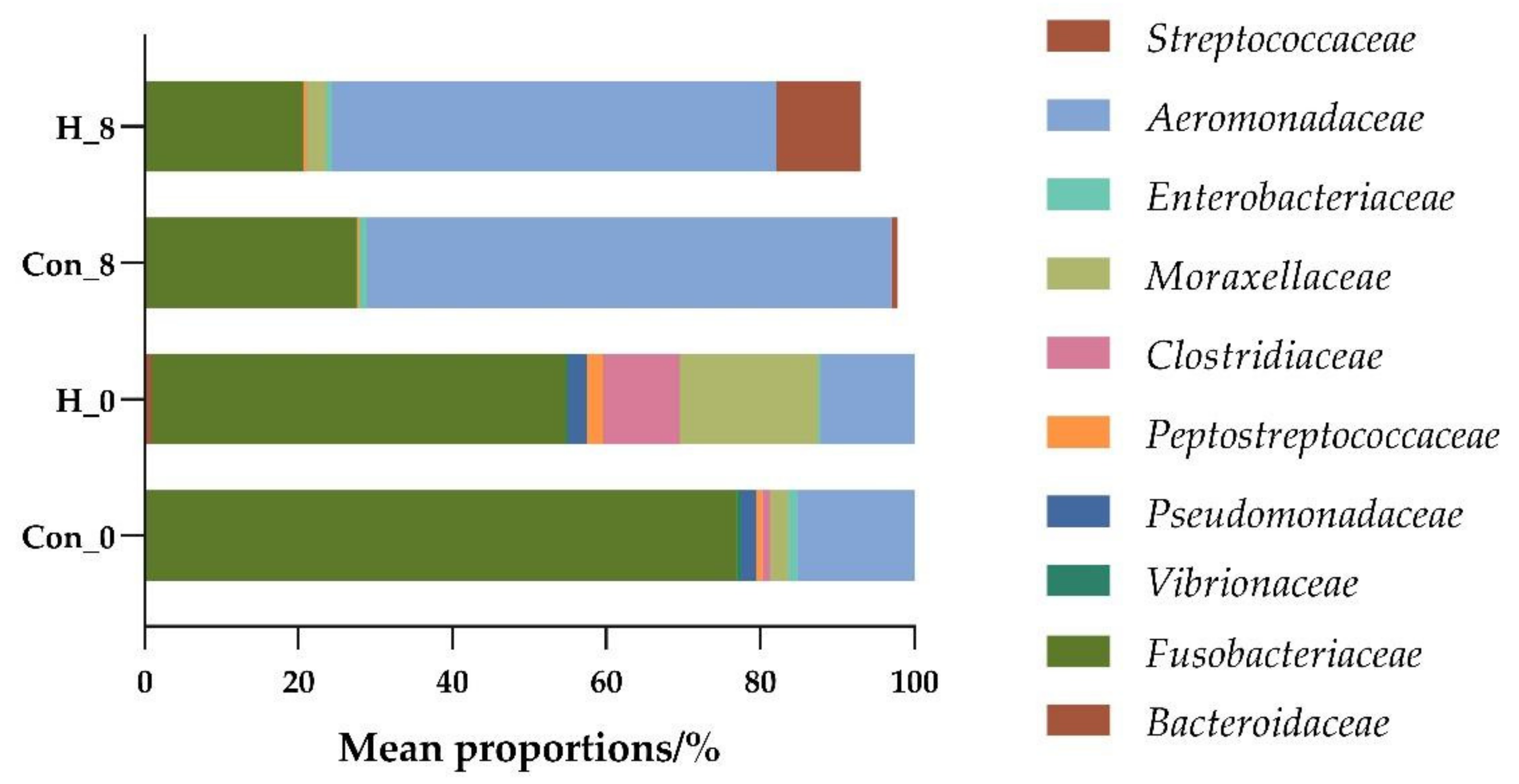
3.2.1. Effects of B. subtilis on Intestinal Flora Diversity of Tilapia
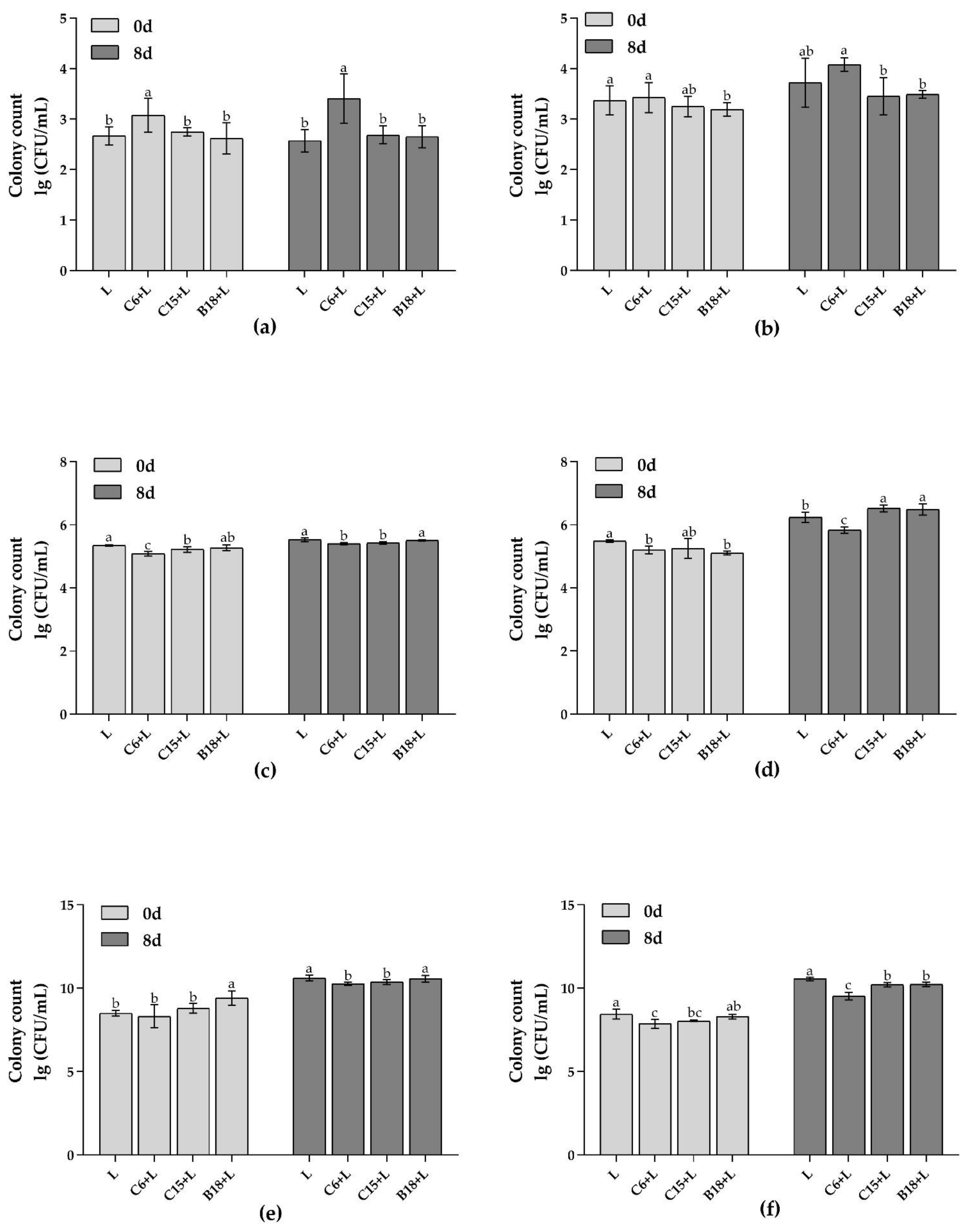
3.2.2. Analysis of Target Flora in Fish Intestines and Flesh by qPCR
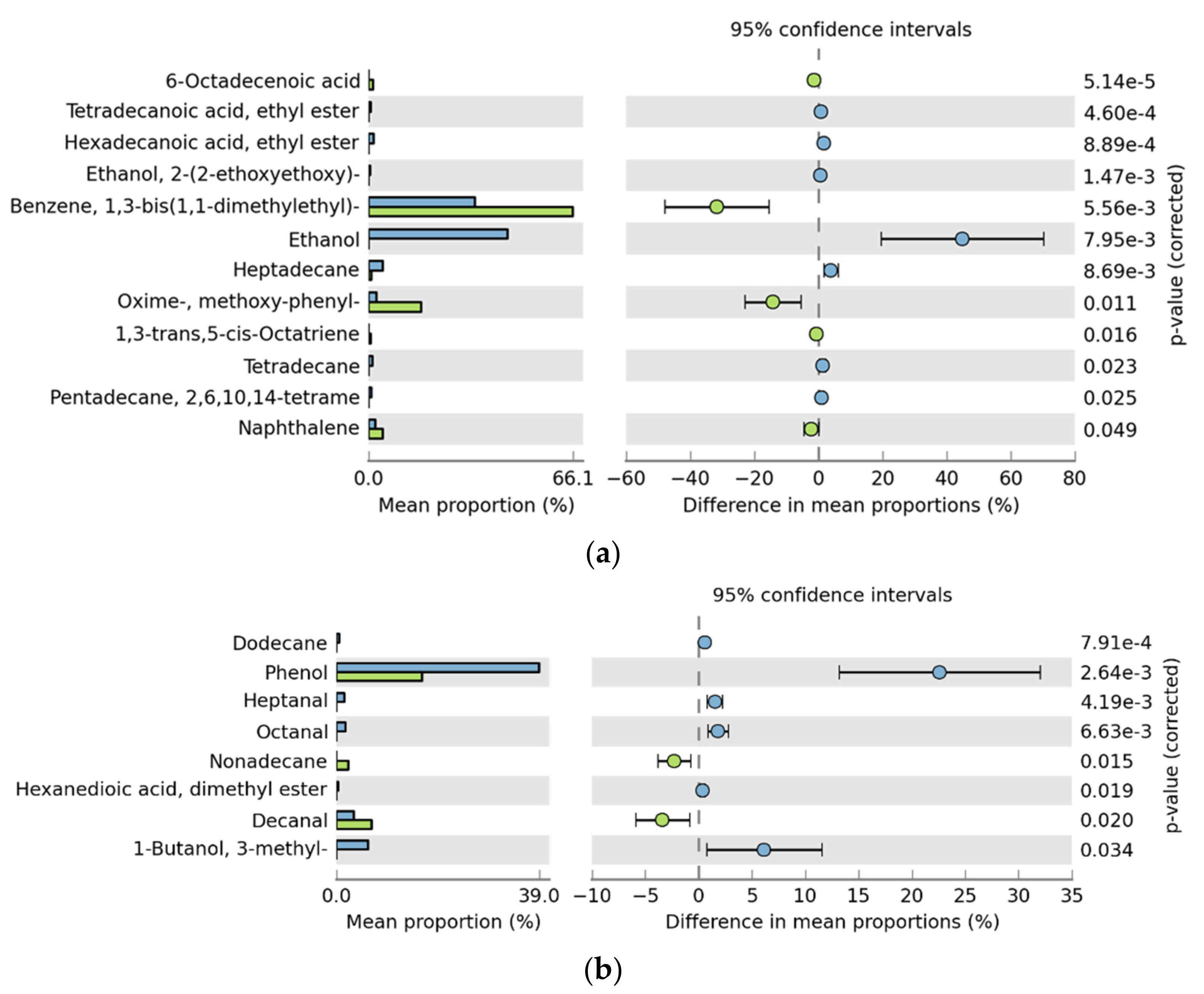
3.3. Analysis of Volatile Flavor Compounds in Fish Flesh
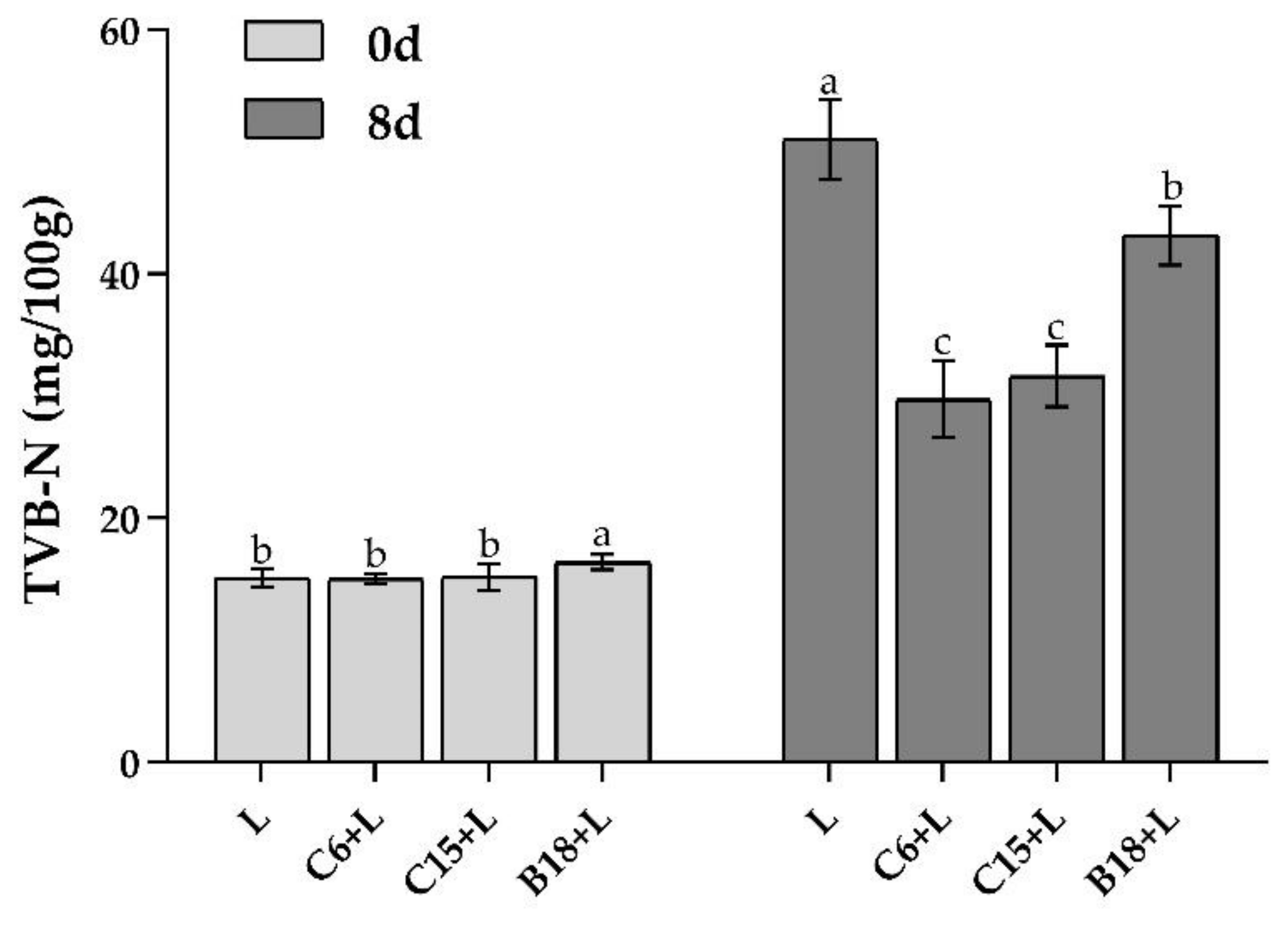
3.4. Analysis of TVB-N Content in Fish Flesh
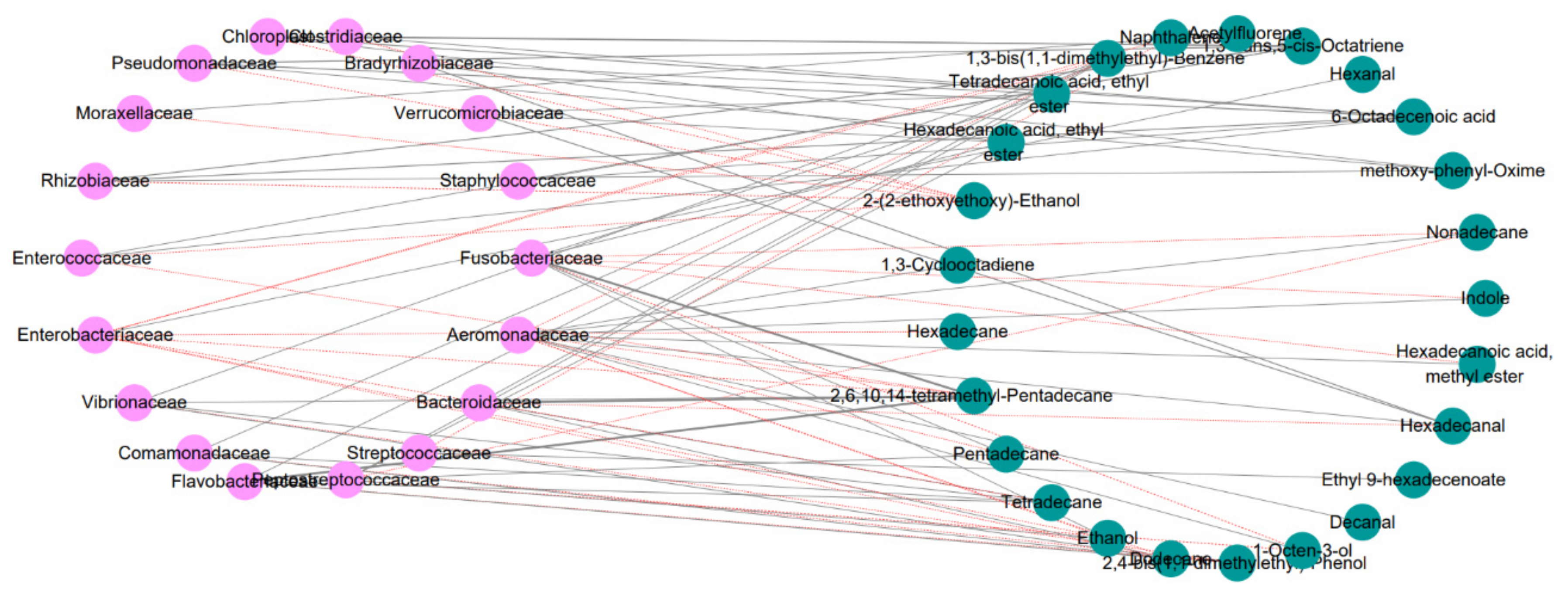
3.5. Correlation Analysis between Intestinal Flora and Volatile Flavor Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newaj-Fyzul, A.; Austin, B. Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J. Fish Dis. 2015, 38, 937–955. [Google Scholar] [CrossRef] [PubMed]
- Lorentzen, G.; Ageeva, T.N.; Heide, M.; Esaiassen, M. Temperature fluctuations in processing and distribution: Effect on the shelf life of fresh cod fillets (Gadus morhua L.). Food Control 2020, 112, 107102. [Google Scholar] [CrossRef]
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of probiotic Bacillus aerius B81e isolated from healthy hybrid catfish on growth, disease resistance and innate immunity of Pla-mong Pangasius bocourti. Fish Shellfish Immunol. 2018, 73, 1–10. [Google Scholar] [CrossRef]
- Ding, H.; Ruan, D.; Jiang, Y.; Xu, G.; Huang, J. Analysis and comparison of cooked high-value sea fish surimi volatile flavor. Food Ferment. Ind. 2015, 41, 163–169. [Google Scholar]
- Molham, F.; Khairalla, A.S.; Azmy, A.F.; El-Gebaly, E.; El-Gendy, A.O.; AbdelGhani, S. Anti-Proliferative and Anti-Biofilm Potentials of Bacteriocins Produced by Non-PathogenicEnterococcussp. Probiotics Antimicrob. Proteins 2021, 13, 571–585. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Yin, F.; Huang, L.S.; Teng, H.F.; Shen, T.Y.; Qin, H.L. Long-term and continuous administration of Bacillus subtilis during remission effectively maintains the remission of inflammatory bowel disease by protecting intestinal integrity, regulating epithelial proliferation, and reshaping microbial structure and function. Food Funct. 2021, 12, 2201–2210. [Google Scholar] [PubMed]
- Gram, L.; Dalgaard, P. Fish spoilage bacteria–problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Aliakbarpour, H.R.; Chamani, M.; Rahimi, G.; Sadeghi, A.A.; Qujeq, D. The Bacillus subtilis and Lactic Acid Bacteria Probiotics Influences Intestinal Mucin Gene Expression, Histomorphology and Growth Performance in Broilers. Asian-Aust. J. Anim. Sci. 2012, 25, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Ran, C.; Wang, Y.; Zhang, Z.; Ding, Q.; Yang, Y.; Olsen, R.E.; Ringo, E.; Bindelle, J.; Zhou, Z. Use of probiotics in aquaculture of China-a review of the past decade. Fish Shellfish Immunol. 2019, 86, 734–755. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, F.; Jatt, A.N.; Liu, H.; Niu, L.; Zhang, L.; Liu, Y. Characterization of co-culture of Aeromonas and Pseudomonas bacterial biofilm and spoilage potential on refrigerated grass carp (Ctenopharyngodon idellus). Lett. Appl. Microbiol 2020, 71, 337–344. [Google Scholar]
- Dabadé, D.S.; den Besten, H.M.; Azokpota, P.; Nout, M.R.; Hounhouigan, D.J.; Zwietering, M.H. Spoilage evaluation, shelf-life prediction, and potential spoilage organisms of tropical brackish water shrimp (Penaeus notialis) at different storage temperatures. Food Microbiol. 2015, 48, 8–16. [Google Scholar] [CrossRef]
- Kesel, S.; Mader, A.; Seeberger, P.H.; Lieleg, O.; Opitz, M. Carbohydrate Coating Reduces Adhesion of Biofilm-Forming Bacillus subtilis to Gold Surfaces. Appl. Environ. Microbiol. 2014, 80, 5911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, K.; Matsumoto, S. Bacterial adhesion: From mechanism to control. Biochem. Eng. J. 2010, 48, 424–434. [Google Scholar] [CrossRef]
- Slizewska, K.; Chlebicz-Wojcik, A.; Nowak, A. Probiotic Properties of New Lactobacillus Strains Intended to Be Used as Feed Additives for Monogastric Animals. Probiotics Antimicrob. Proteins 2021, 13, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Li, P.; Yu, Q.; Yang, Q. Bacillus subtilis inhibition of enterotoxic Escherichia coli-induced activation of MAPK signaling pathways in Caco-2 cells. Ann. Microbiol. 2013, 63, 577–581. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Giri, S.S.; Ryu, E.C.; Sukumaran, V.; Park, S.C. Antioxidant, antibacterial, and anti-adhesive activities of biosurfactants isolated from Bacillus strains. Microb. Pathog. 2019, 132, 66–72. [Google Scholar] [CrossRef]
- Lau, L.Y.J.; Chye, F.Y. Antagonistic effects of Lactobacillus plantarum 0612 on the adhesion of selected foodborne enteropathogens in various colonic environments. Food Control 2018, 91, 237–247. [Google Scholar]
- Zhang, W.; Lv, X.; Liu, Z.; Ni, L. The spoilage and adhesion inhibitory effects of Bacillus subtilis against Shewanella and Pseudomonas in large yellow croaker (Pseudosciaena crocea). Food Sci. Technol. 2021, 10, 02721. [Google Scholar] [CrossRef]
- Larsen, M.H.; Larsen, J.L.; Olsen, J.E. Chemotaxis of Vibrio anguillarum to fish mucus: Role of the origin of the fish mucus, the fish species and the serogroup of the pathogen. FEMS Microbiol. Ecol. 2001, 38, 77–80. [Google Scholar] [CrossRef]
- Forestier, C.; Champs, C.D.; Vatoux, C.; Joly, B. Probiotic activities of Lactobacillus casei rhamnosus: In vitro adherence to intestinal cells and antimicrobial properties. Res. Microbiol. 2001, 152, 167–173. [Google Scholar] [CrossRef]
- GB 5009.228-2016. National Food Safety Standard-Determination of Volatile Salt-Based Nitrogen in Foods; Standards Press of China: Beijing, China, 2016; Volume 1, pp. 3–5. [Google Scholar]
- Xue, W.; Chun, L.; Jing, W.; Qiao, Z.; Zhi, D. Identification and analysis of the flavor components in the fermentation of Siniperca Scherzeri. Chin. J. Food Sci. 2015, 15, 222–229. [Google Scholar]
- Guojun, G.; Qingpi, Y.; Wenzheng, Z.; Rensun, Z.; Changsheng, C.; Qiang, C. Influence of antagonistic bacteria on adhesion of pathogenic vibrio alginoyticus to skin MUCUS of pseudoscaena crocea. Ocean. Lakes 2009, 40, 738–744. [Google Scholar]
- Weng, Z.; Dan, B.; Cheng, R.; Xiang, S. Adhesion and biofilm characteristics of spoilage bacteria from large yellow croaker pseudosciaena crocea. Food Sci. 2019, 40, 14. [Google Scholar]
- Wang, X.Y.; Xie, J. Growth Kinetics and Spoilage Potential of Co-culturing Acinetobacter johnsonii and Pseudomonas fluorescens from Bigeye Tuna (Thunnus obesus) During Refrigerated Storage. Curr. Microbiol. 2020, 77, 1637–1646. [Google Scholar] [CrossRef]
- Guo, H.X.; Sun, Z.Y.; Hao, Y.; Zhang, L.Y.; Ren, Y.T.; Zhang, Y.; Chen, Z.J. Correlation between bacterial communities and organic acids in the fermentation stage of traditional Chinese sour porridge. Int. J. Food Prop. 2020, 23, 1430–1440. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Michailidou, S.; Anagnostopoulos, D.A.; Koromilas, S.; Kios, K.; Pasentsis, K.; Psomopoulos, F.; Argiriou, A.; Haroutounian, S.A.; Boziaris, I.S. Bacterial communities and potential spoilage markers of whole blue crab (Callinectes sapidus) stored under commercial simulated conditions. Food Microbiol. 2019, 82, 325–333. [Google Scholar] [CrossRef]
- Wu, S.Z.; Hou, W.J.; Suo, X.K.; Guo, X.P.; Li, H. Bacillus subtilis extracellular polymeric substances conditioning layers inhibit Escherichia coli adhesion to silicon surfaces: A potential candidate for interfacial antifouling additives. Biointerphases 2021, 16, 011003. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Cai, D.D.; Li, W.Q.; Blank, R.; Liu, Y. Application of gas chromatography-ion mobility spectrometry (GC-IMS) and ultrafast gas chromatography electronic-nose (uf-GC E-nose) to distinguish four Chinese freshwater fishes at both raw and cooked status. J. Food Biochem. 2021, 1, 13840. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Li, L.; Yang, X.; Chen, S.; Wu, Y.; Zhao, Y.; Wei, Y. Analysis of volatile flavor components and fatty acids in fish sauces during fermentation by GC-MS. J. Fish. China 2018, 42, 984–995. [Google Scholar]
- Liu, Y.; Huang, Y.Z.; Wang, Z.M.; Cai, S.H.; Zhu, B.W.; Dong, X.P. Recent advances in fishy odour in aquatic fish products, from formation to control. Int. J. Food Sci. Technol. 2021, 10, 15269. [Google Scholar] [CrossRef]
- Chen, J.H.; Tao, L.N.; Zhang, T.; Zhang, J.J.; Wu, T.T.; Luan, D.L.; Ni, L.; Wang, X.C.; Zhong, J. Effect of four types of thermal processing methods on the aroma profiles of acidity regulator-treated tilapia muscles using E-nose, HS-SPME-GC-MS, and HS-GC-IMS. LWT-Food Sci. Technol. 2021, 147, 111585. [Google Scholar] [CrossRef]
- Wu, Y.; You, G.; Li, L.; Yang, X.; Deng, J.; Chen, S. Comparison of flavor components between low-salt lactic acid fermented fish and traditional salted fish. J. Fish. China 2014, 38, 601–612. [Google Scholar]
- Kang, C.; Shi, W.; Fang, L.; Wang, X. Effects of Different Freezing Methods on the Volatile Components of Grass Carp Meat. Food Sci. 2018, 39, 229–235. [Google Scholar]
- Curioni, P.M.G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Li, Y.; Huang, J.; Yuan, C.; Ding, T.; Chen, S.; Hu, Y. Developing a new spoilage potential algorithm and identifying spoilage volatiles in small yellow croaker (Larimichthys polyactis) under vacuum packaging condition. Lebensm. Wiss. Technol 2019, 106, 209–217. [Google Scholar] [CrossRef]
- Singh, T.P.; Gurpreet, K.; Suman, K.; Malik, R.K. Antagonistic Activity of Lactobacillus reuteri Strains on the Adhesion Characteristics of Selected Pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Gómez, N.C.; Ramiro, J.M.; Quecan, B.X.; de Melo Franco, B.D. Use of Potential Probiotic Lactic Acid Bacteria (LAB) Biofilms for the Control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157:H7 Biofilms Formation. Front. Microbiol. 2016, 7, 863. [Google Scholar] [CrossRef] [Green Version]
- Dandan, Z.; Mengting, W.; Yuxing, G. Purification of Lactobacillus acidophilus surface-layer protein and its immunomodulatory effects on RAW264.7 cells. J. Sci. Food Agric. 2017, 97, 12. [Google Scholar]
- Shi, F.; Zi, Y.J.; Lu, Z.J.; Li, F.L.; Yang, M.X.; Zhan, F.B.; Li, Y.N.; Li, J.; Zhao, L.J.; Lin, L.; et al. Bacillus subtilis H2 modulates immune response, fat metabolism and bacterial flora in the gut of grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2020, 106, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.V.; Antony, M.L.; Praveen, N.K.; Sayooj, P.; Swaminathan, T.R.; Vijayan, K.K. Evaluation of in vitro and in vivo potential of Bacillus subtilis MBTDCMFRI Ba37 as a candidate probiont in fish health management. Microb. Pathog. 2021, 152, 104610. [Google Scholar] [CrossRef]
- Zhu, J.; Zhao, A.; Feng, L.; Gao, H. Quorum sensing signals affect spoilage of refrigerated large yellow croaker (Pseudosciaena crocea) by Shewanella baltica. Int. J. Food Microbiol. 2016, 217, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D.; Casaburi, A.; Nasi, A.; Ferrocino, I.; di Monaco, R.; Ferranti, P.; Mauriello, G.; Villani, F. Different molecular types of Pseudomonas fragi have the same overall behaviour as meat spoilers. Int. J. Food Microbiol. 2010, 142, 120–131. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, L.; Liu, Y.; Zhou, S.; Yang, Y.; Xu, N.; Yang, Q.; Ai, X. Resveratrol influences the pathogenesis of Aeromonas hydrophila by inhibiting production of aerolysin and biofilm. Food Control 2021, 126, 108083. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Wu, Y.Y.; Wang, Y.Q.; Li, L.H.; Li, C.S.; Zhao, Y.Q.; Yang, S.L. Contribution of autochthonous microbiota succession to flavor formation during Chinese fermented mandarin fish (Siniperca chuatsi). Food Chem. 2021, 348, 129107. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Wang, L.; Zhang, K.; Qiu, Z.; Zhang, K.; Yue, C.; Zhang, Y.; Li, J.; Zhao, X. The safety and potential probiotic properties analysis of Streptococcus alactolyticus strain FGM isolated from the chicken cecum. Ann. Microbiol. 2021, 71, 1–14. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | GenBank Accession Number |
|---|---|
| Bacillus subtilis B02 | OL423511 |
| Bacillus subtilis B08 | OL423512 |
| Bacillus subtilis B15 | OL423514 |
| Bacillus subtilis B18 | OL423516 |
| Bacillus subtilis C6 | OL423515 |
| Bacillus subtilis C15 | OL423517 |
| Pseudomonas spp. L. | OL423525 |
| Strain | Primer | Sequence (5′→3′) |
|---|---|---|
| Total bacteria | 27-F | AGAGTTTGATCCTGGCTCAG |
| 1429-R | GGTTACCTTGTTACGACTT | |
| Pseudomonas | Pse-F | CTGCATCATGGCCGGTGACAACATTT |
| Pse-R | GTCGCATGGCTGTCGGTCTTCAGATC | |
| Bacillus subtilis | Bac-F | AAAGTCTGACGGAGCAACGC |
| Bac-R | ACCGCCCTATTCGAACGGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Tong, Q.; You, J.; Lv, X.; Liu, Z.; Ni, L. The Application of Bacillus subtilis for Adhesion Inhibition of Pseudomonas and Preservation of Fresh Fish. Foods 2021, 10, 3093. https://doi.org/10.3390/foods10123093
Zhang W, Tong Q, You J, Lv X, Liu Z, Ni L. The Application of Bacillus subtilis for Adhesion Inhibition of Pseudomonas and Preservation of Fresh Fish. Foods. 2021; 10(12):3093. https://doi.org/10.3390/foods10123093
Chicago/Turabian StyleZhang, Wen, Qiuxia Tong, Jiahong You, Xucong Lv, Zhibin Liu, and Li Ni. 2021. "The Application of Bacillus subtilis for Adhesion Inhibition of Pseudomonas and Preservation of Fresh Fish" Foods 10, no. 12: 3093. https://doi.org/10.3390/foods10123093
APA StyleZhang, W., Tong, Q., You, J., Lv, X., Liu, Z., & Ni, L. (2021). The Application of Bacillus subtilis for Adhesion Inhibition of Pseudomonas and Preservation of Fresh Fish. Foods, 10(12), 3093. https://doi.org/10.3390/foods10123093