
Effects of Dietary Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism, Liver Inflammation and Gut Microbiota in Rats

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Equipment
2.2. Conditions and Processes of the Three Cooking Methods
- (1)
- BCM: Five hundred grams of pork belly was cooked in 3500 mL of water at 100 °C for 15 min and then cut into 2 × 3 × 3 cm blocks, which were sautéed at 160 °C for 5 min without water. Next, 120 g of green onions, 120 g of ginger, 30 g of cooking wine, 2500 mL of water, 20 g of dark soy sauce, 20 g of light soy sauce, 30 g of white sugar and 2 g of salt were added to the pork, which was cooked at 200 W for 90 min in an induction cooker. The obtained samples were collected.
- (2)
- SCM: Five hundred grams of pork belly was cut into 0.6 × 0.5 × 0.5 cm granules and mixed with 6 g of green onions, 6 g of ginger, 8 g of salt and 10 g of starch. The pork pieces were boiled in 2500 mL of water at 100 °C for 3 min, and then the cooked samples were transferred to a saucepan with 2500 mL of water. After stewing at 200 W for 120 min, samples were collected.
- (3)
- DCM: Five hundred grams of pork belly was finely pureed and mixed with 6 g of green onions, 6 g of ginger, 3 g of salt and 10 g of starch. From this mixture, pork balls with a diameter of 2.0 cm (30 g/sample) were made by hand and fried in 2500 mL of soybean oil at 150 °C for 8 min. Samples were collected.
2.3. Determination of Fatty Acids
- (1)
- For fat extraction, 5 g of pork sample obtained from meat samples cooked using each of the three methods was placed in a Petri dish and dried at 102 °C for 1 h in an oven. The dried samples were ground. Amounts of 0.5 g of each dry sample and 2 mL of a benzene–petroleum ether solution (1:1) were mixed in a centrifuge tube and reacted for 24 h.
- (2)
- After the extraction, 2 mL of a 0.4 mol/L potassium hydroxide-methanol solution was added to each sample, followed by vortexing for 3 min. Next, 100 μL of an internal standard was added to each sample. After a 30 min incubation, an ultrapure water layer was added, and the upper layer of the solution was collected. Next, a certain amount of anhydrous sodium sulphate was added for use.
- (3)
- A 100 μL aliquot of the collected sample layer was diluted with 1 mL of n-hexane, mixed well and passed through a 0.22 µm membrane filter for injection. The free fatty acids in each sample were analyzed by gas chromatography-mass spectrometry (GC-MS).
2.4. Preparation of Animal Feed
2.5. Animals and Experimental Design
- (1)
- Collection of fecal samples: Fresh feces were collected by the extrusion method and snap frozen in liquid nitrogen. The samples were stored in a freezer at −80 °C until testing.
- (2)
- Collection of serum samples: On days 0 (starting time), 14, 28, 42 and 56 of the experiment, capillary blood samples were collected from the rats via the retroorbital route. The samples were centrifuged for 10 min at 5000 rpm and 4 °C to separate the sera. The serum layer of each sample was removed, placed in a 0.5 mL centrifuge tube and stored at −80 °C until testing.
- (3)
- Rat organ collection: On day 56 of feeding, the rats were killed by spinal dislocation, after which the rats were quickly dissected, and the liver tissues were harvested. After washing in normal saline, the tissues were weighed. Two 1.5 × 1.5 cm sections of tissue were cut from the same part of the large liver lobe, snap frozen in liquid nitrogen and stored at −80 °C until testing.
2.6. Determination of Rat Body Mass Index
2.7. Determination of Biochemical Indicators of Glucose and Lipids in Rats
2.8. Rat Liver Oil Red O Staining Method
2.9. Determination of Inflammatory Factor Expression in Rat Livers
2.10. Analysis of the Rat Gut Microbiota Structure
- (1)
- Collection and storage of fecal samples. Feces were collected before the rats were sacrificed on day 56 of the feeding period. The collected feces were immediately placed into labeled sterile test tubes, sealed and stored at −80 °C.
- (2)
- DNA extraction and quality identification of fecal flora. Total DNA was extracted from the fecal samples according to the instructions provided with the EZNA Soil kit. The DNA concentration and purity were tested using a NanoDrop 2000 spectrophotometer, and the DNA extraction quality was tested via PCR amplification of the V3–V4 variable region of bacterial 16S rRNA with the 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) primers. The following amplification procedure was used: pre-denaturation for 3 min at 95 °C; 27 cycles of denaturation for 30 s at 95 °C, annealing for 30 s at 55 °C and extension for 30 s at 72 °C; and a final extension for 10 min at 72 °C. All amplification reactions had a total volume of 20 µL, containing 4 µL of 5FastPfu buffer, 2 µL of 2.5 mM dNTPs, 0.8 µL of each primer (5 µM), 0.4 µL of FastPfu polymerase and 10 ng of DNA template. The PCR products were subjected to 1% agarose gel electrophoresis.
- (3)
- Analysis of 16S rRNA data from rat fecal flora. Rat feces were collected after day 56 of feeding, and the DNA was extracted from faces, purified using the AxyPrep DNA Gel Extraction Kit, eluted with Tris-HCL and detected by 2% agarose electrophoresis using QuantiFluorTM-ST for quantification. The purified amplification products were used to construct a PE2300 library according to the Illumina Miseq platform standard operating procedures. Sequencing was performed on the Illumina Miseq PE300 platform (Meiji Biomedical Technology Co., Ltd., Shanghai, China).
2.11. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Fatty Acid Composition of Pork Fat Cooked Using Different Methods
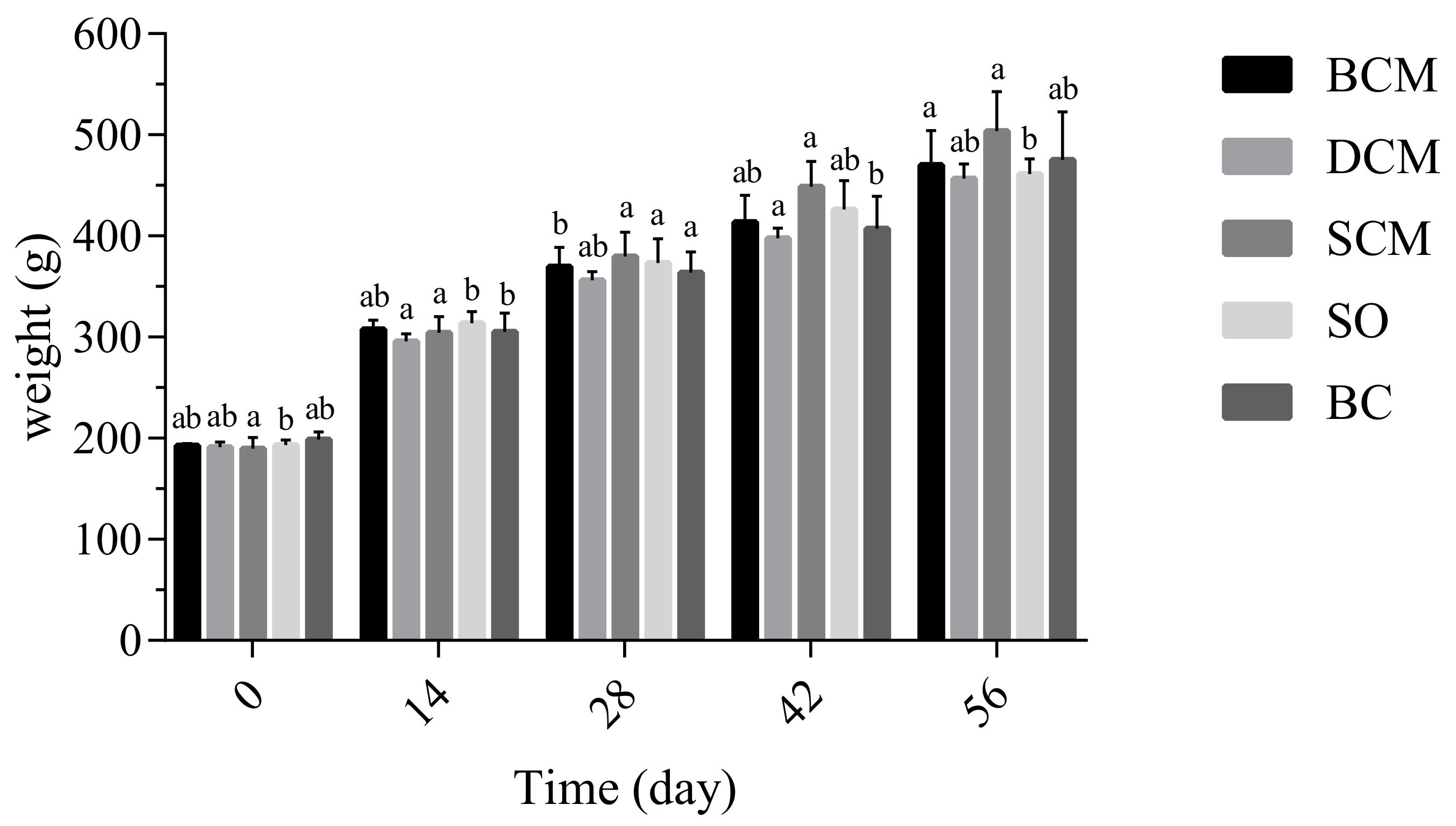
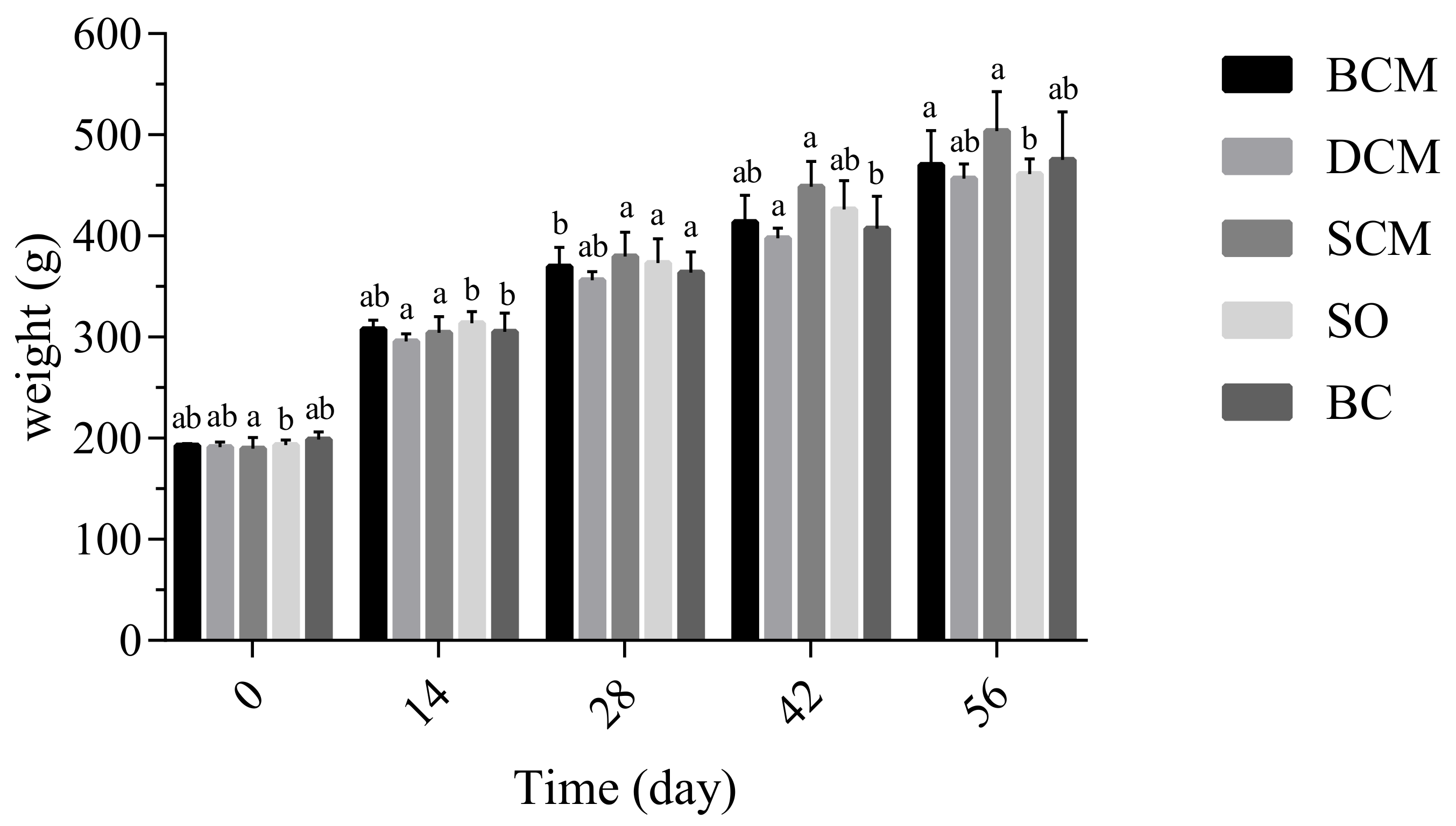
3.2. Effects of Consuming Pork Fat Cooked Using Different Methods on the Growth of Rats
3.3. Effects of Consuming Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism in Rats
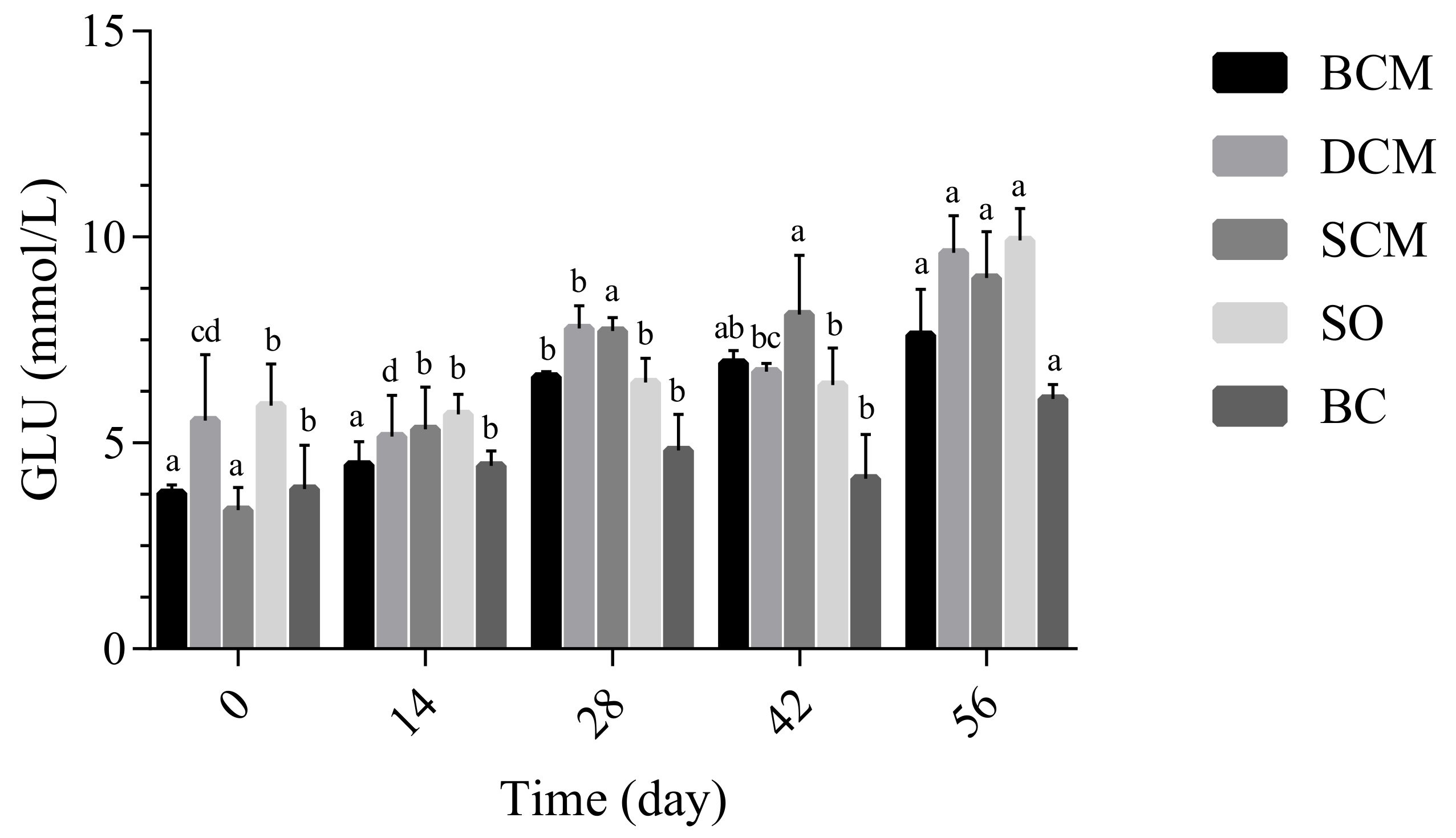
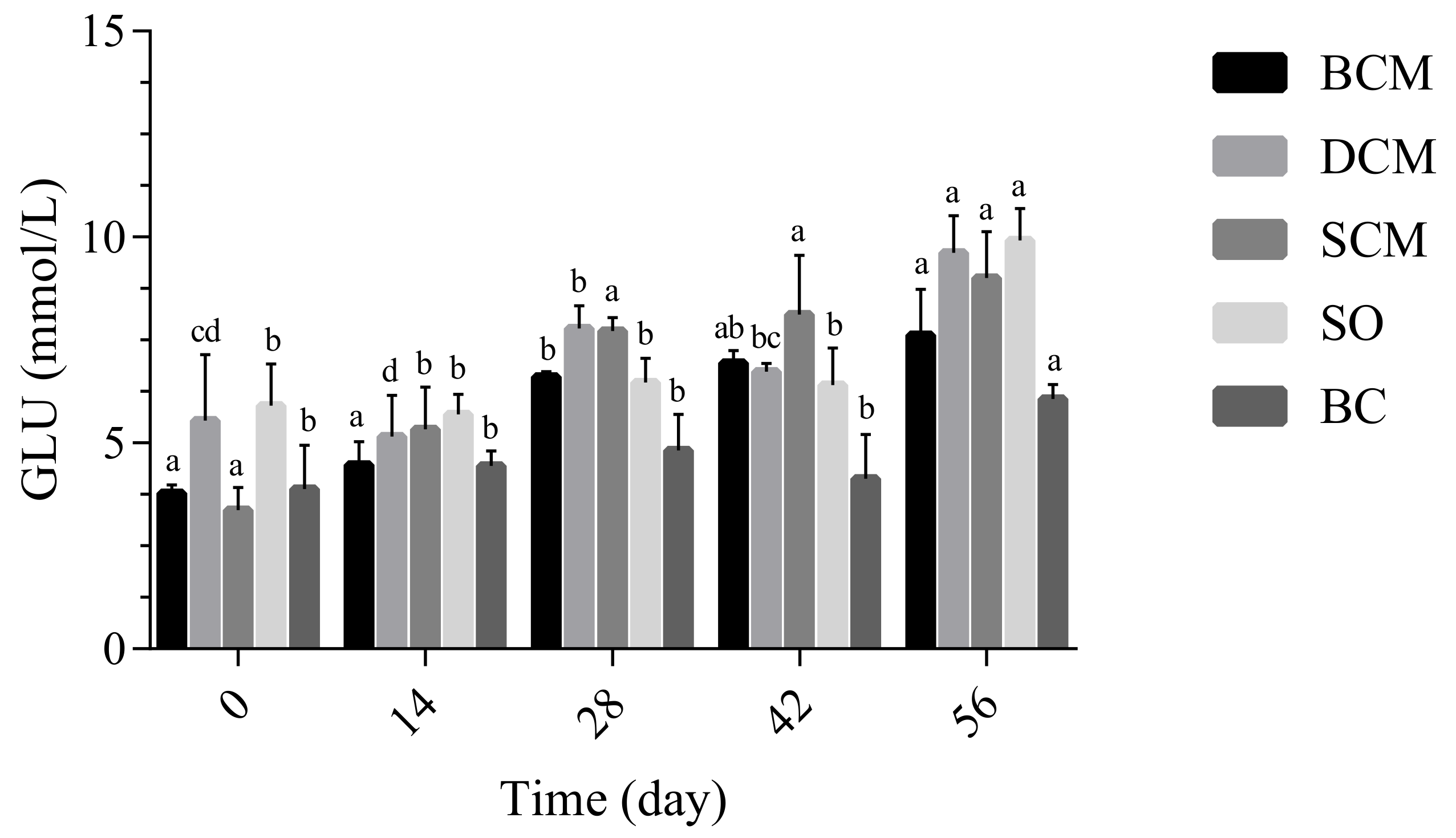
3.3.1. Effects of Pork Fat Cooked Using Different Methods on Blood Glucose
3.3.2. Effects of Pork Fat Cooked Using Different Methods on Serum Lipid Concentrations in Rats
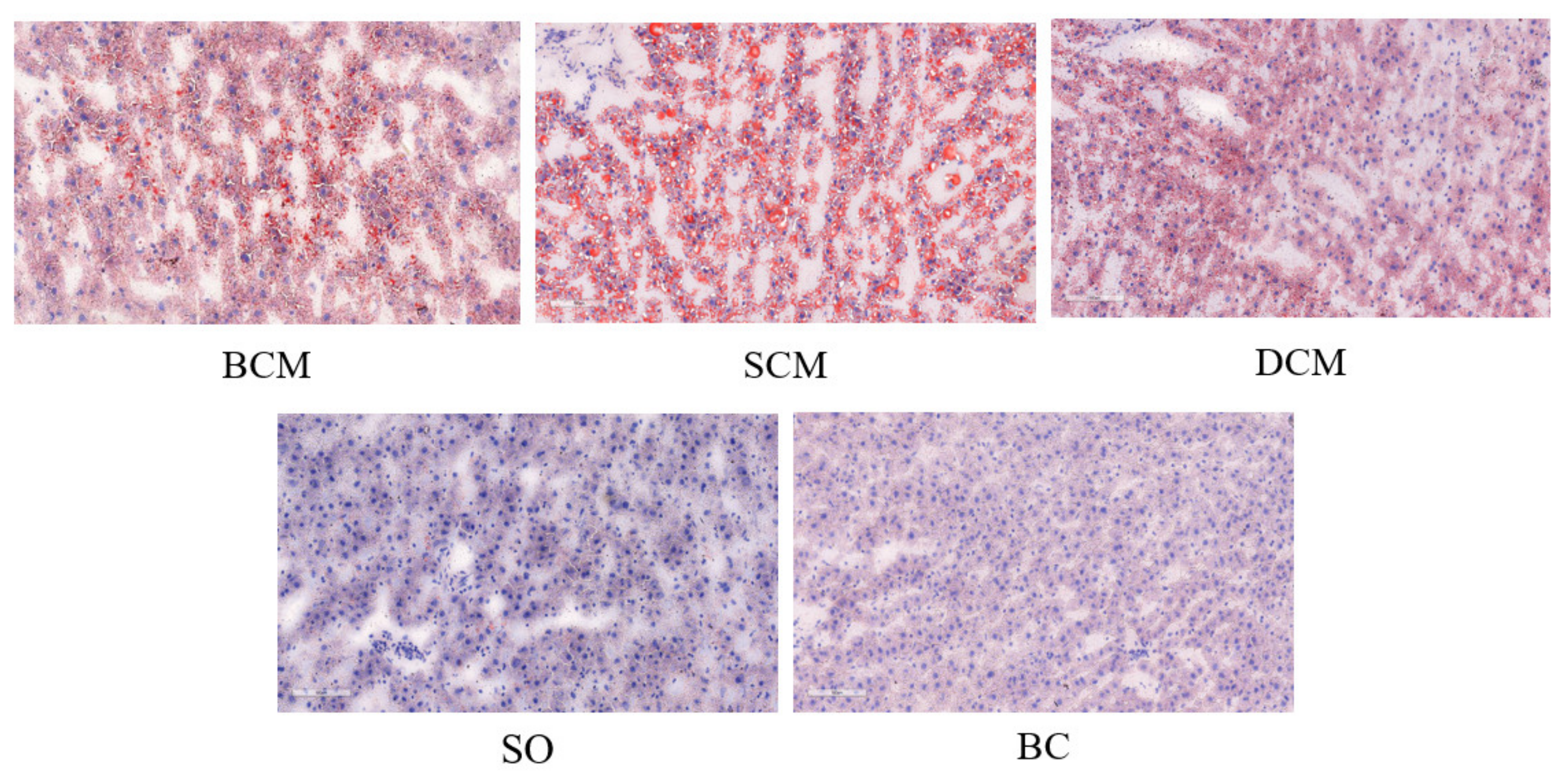
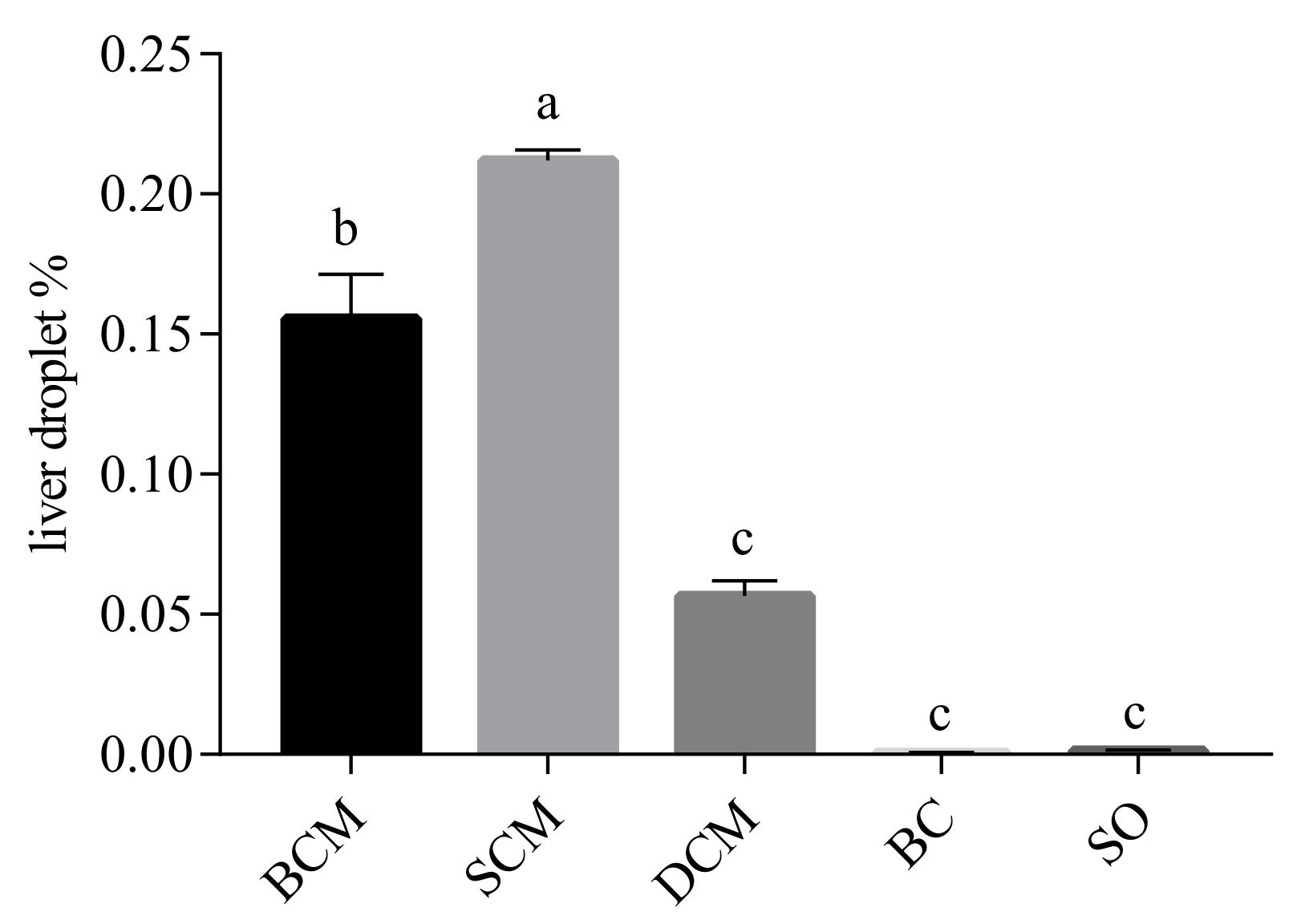
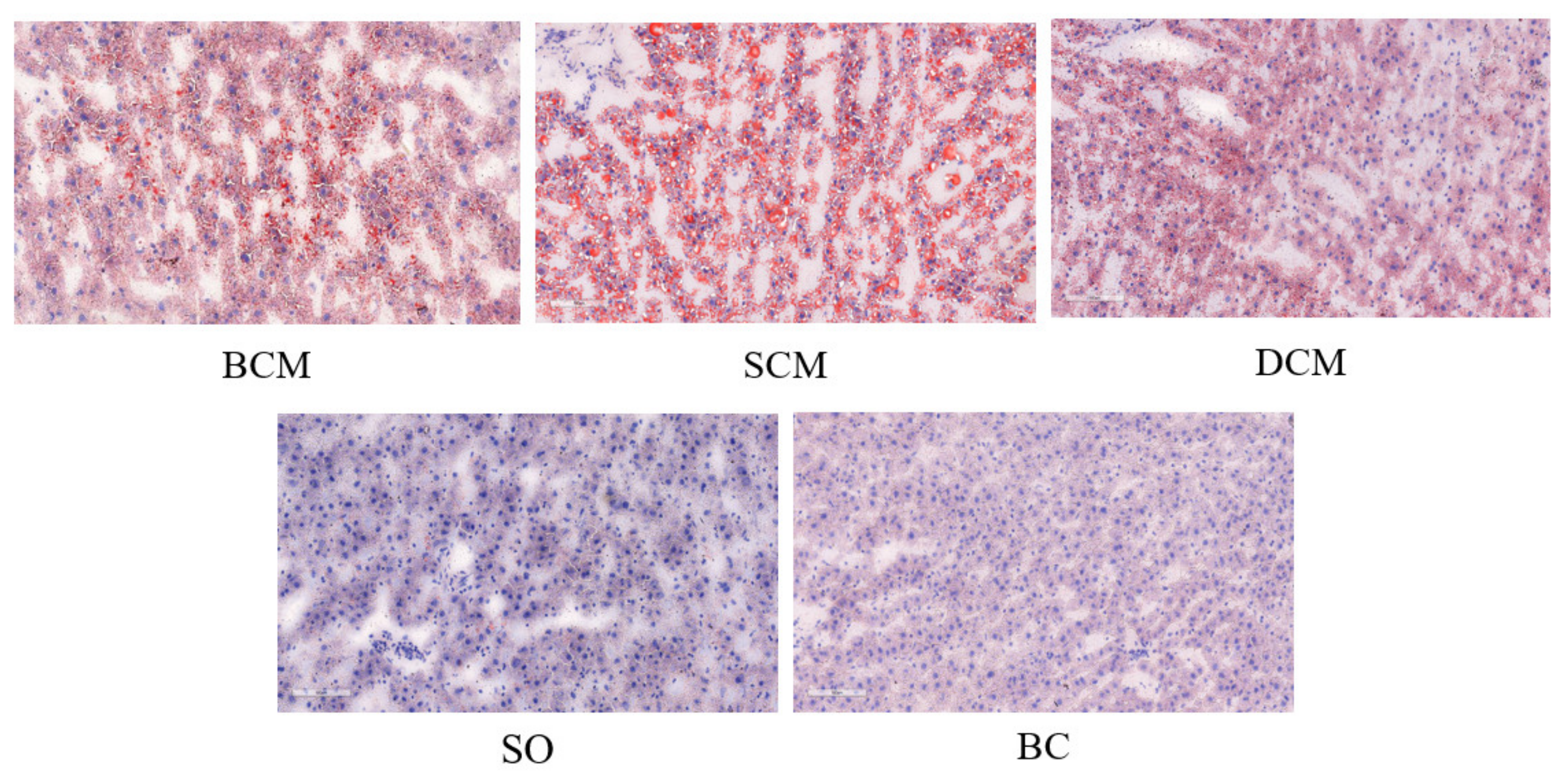
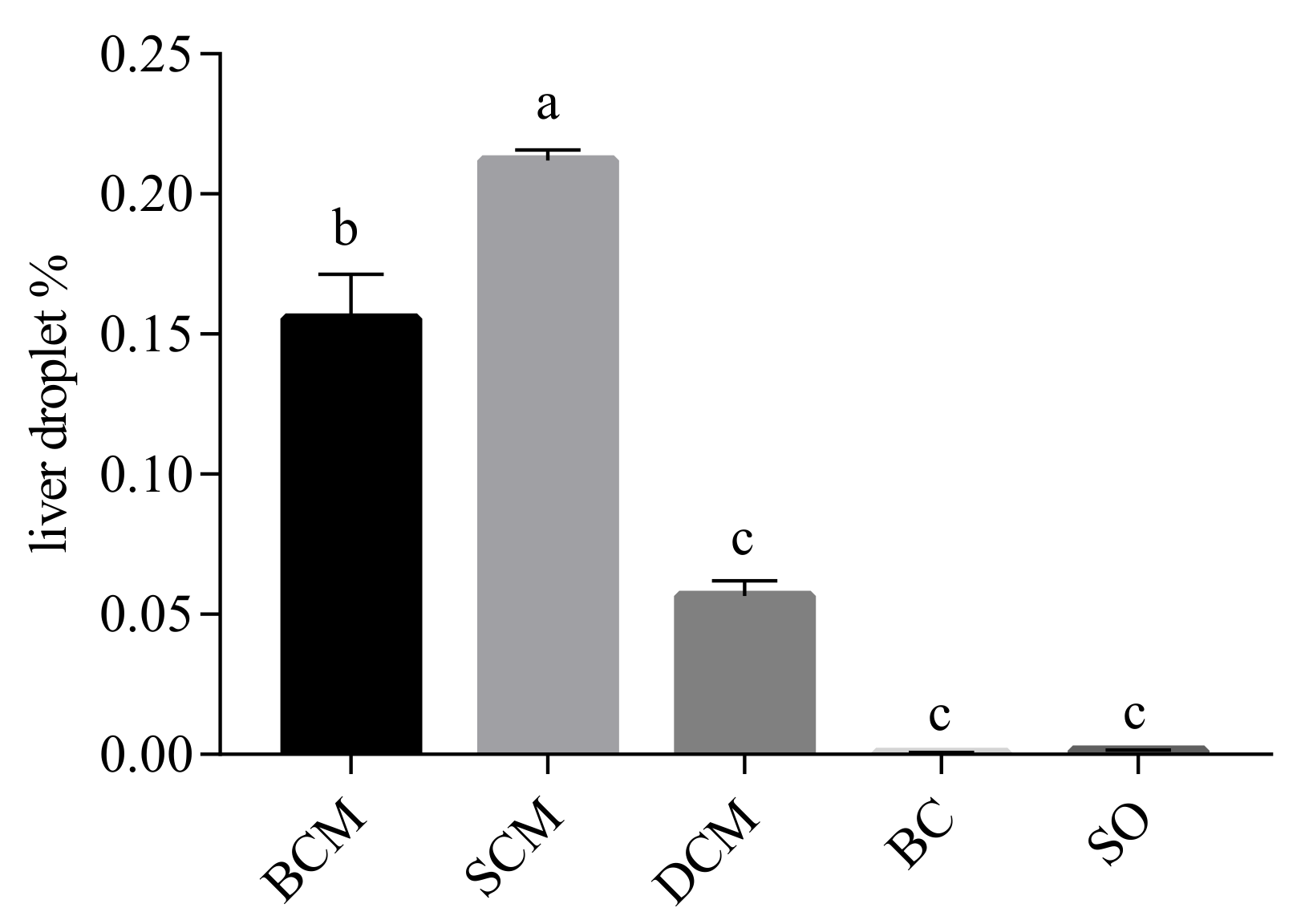
3.4. Effects of Pork Fat Cooked Using Different Methods on Liver Fat Deposition in Rats
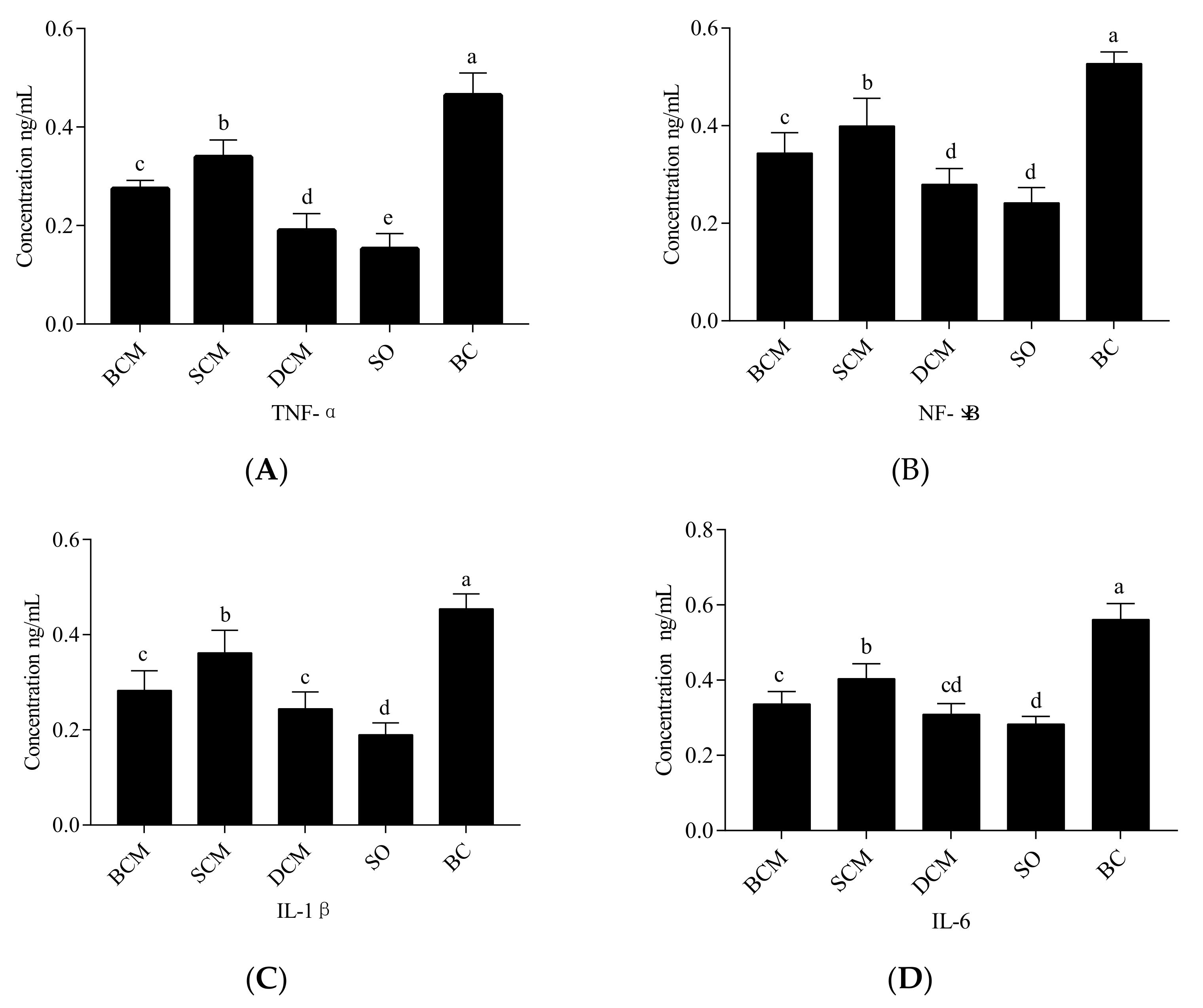
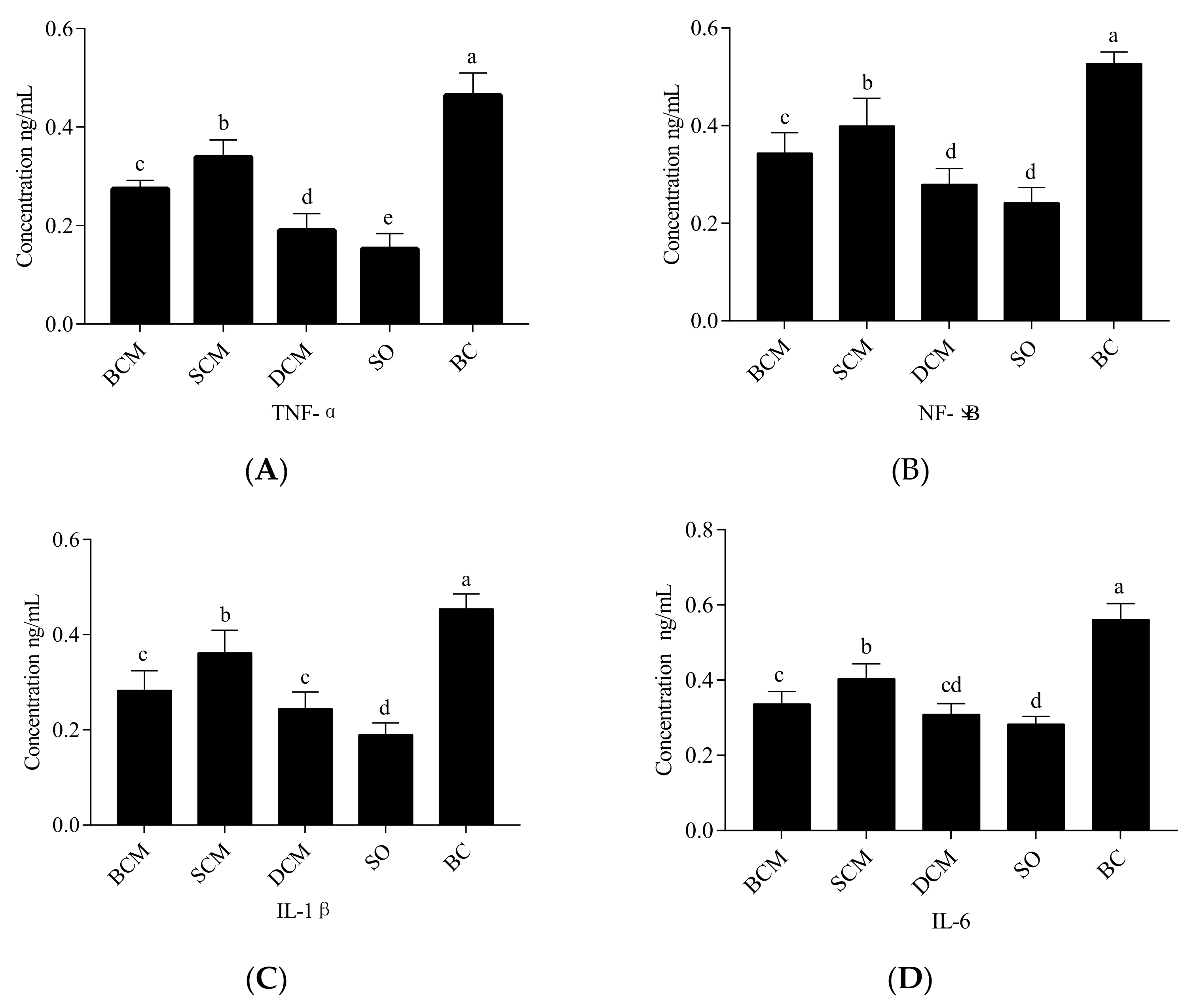
3.5. Effects of Pork Fat Subjected to Different Cooking Methods on Inflammatory Factors in the Liver Tissues of Rats
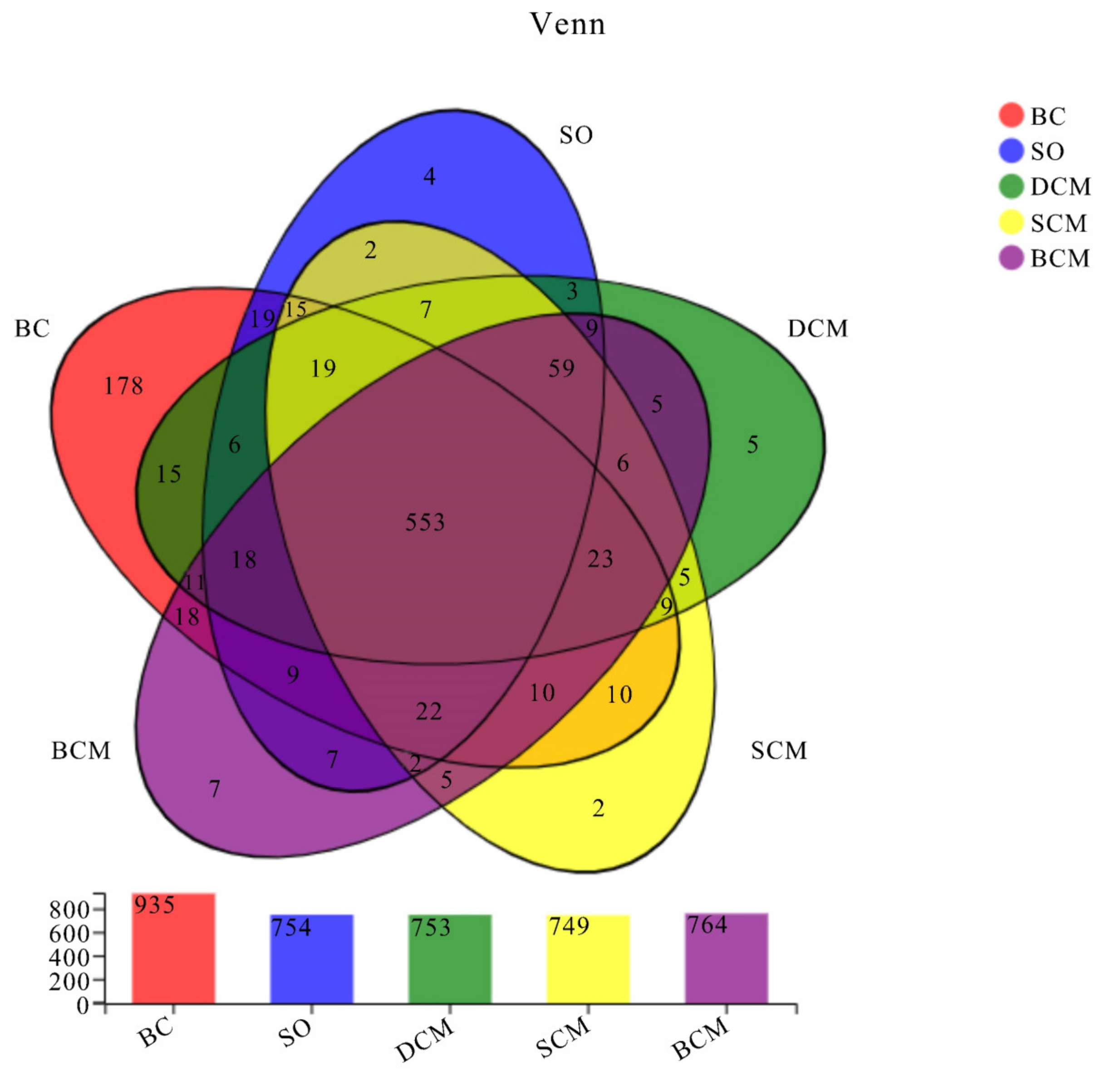
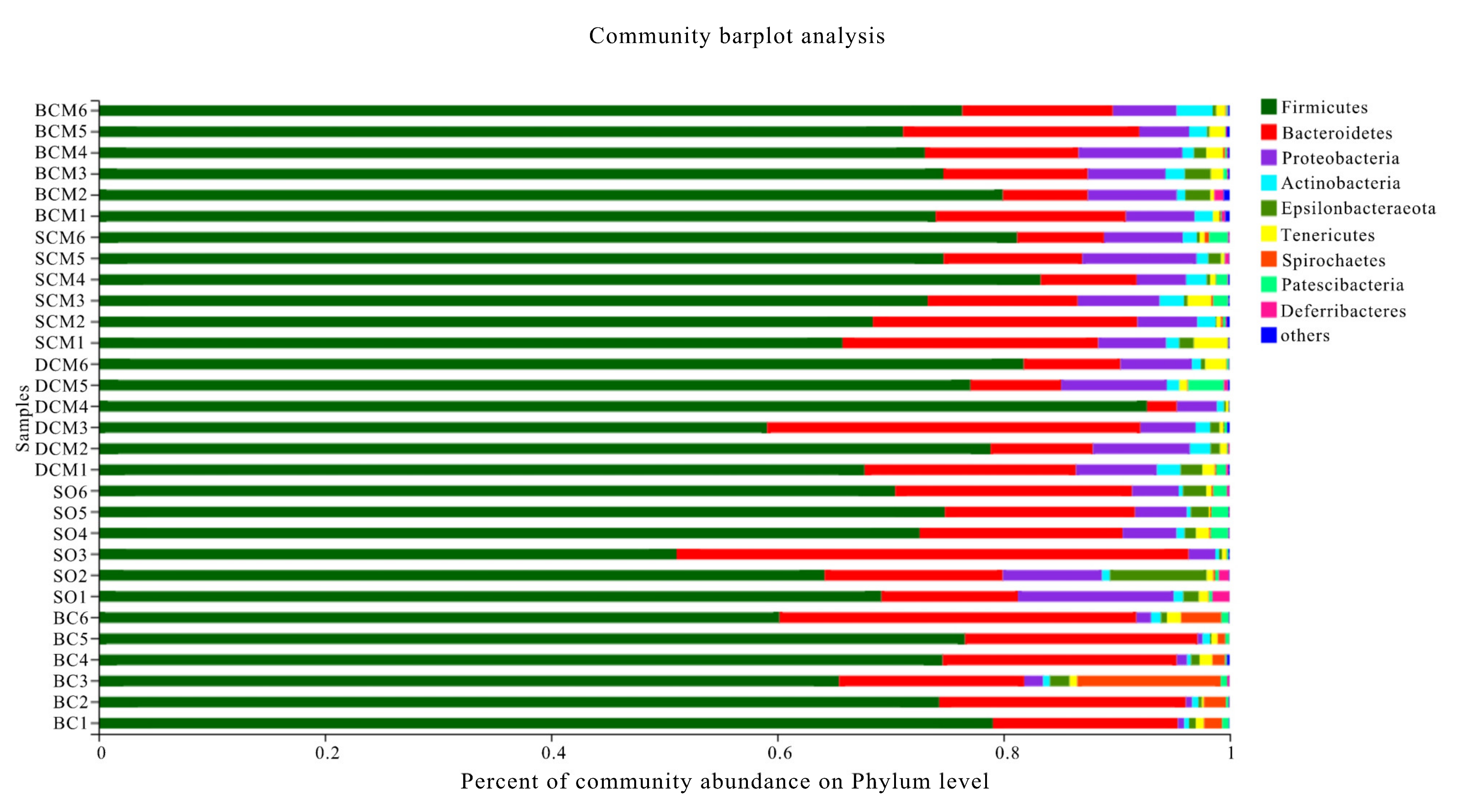
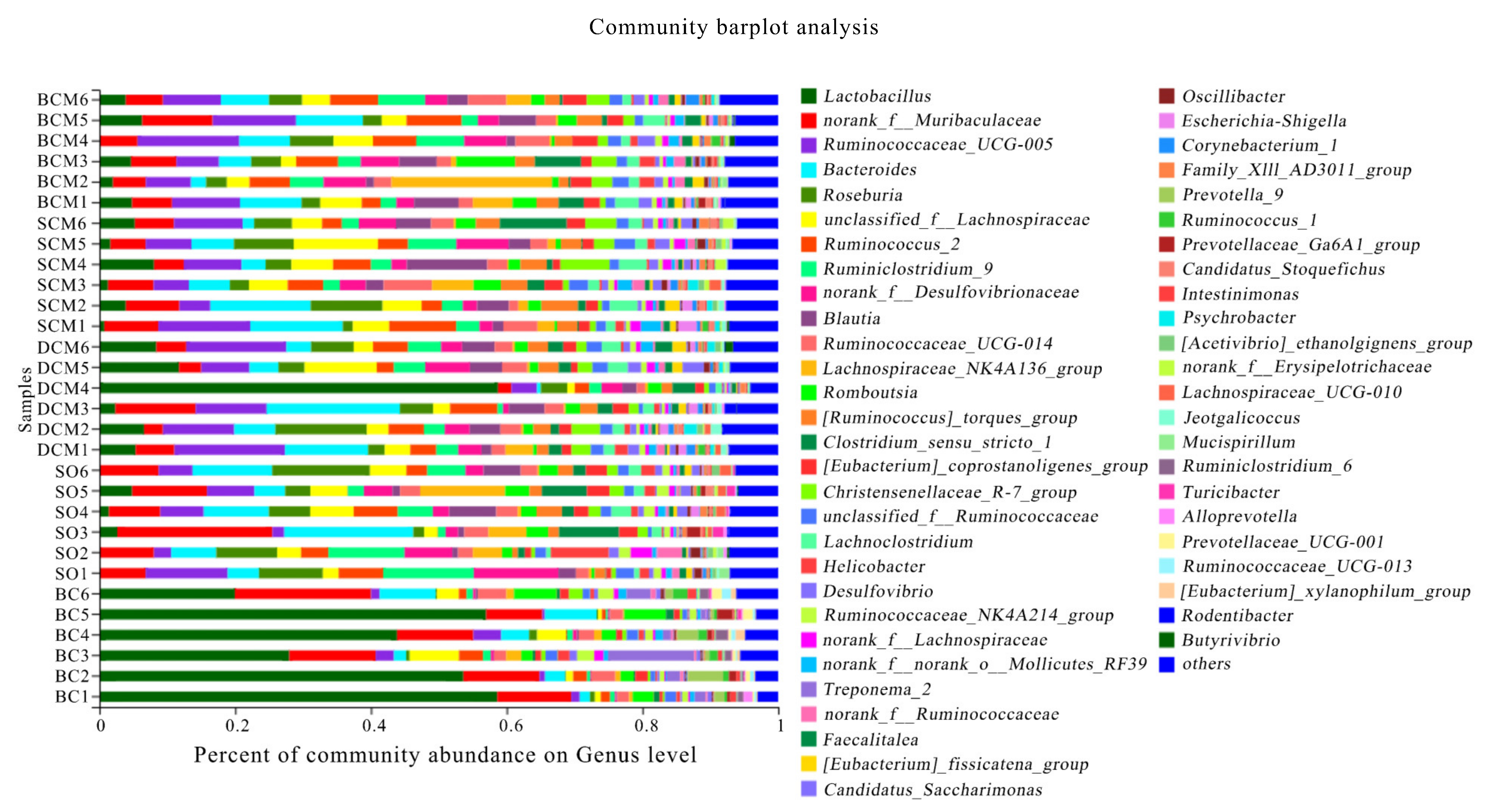
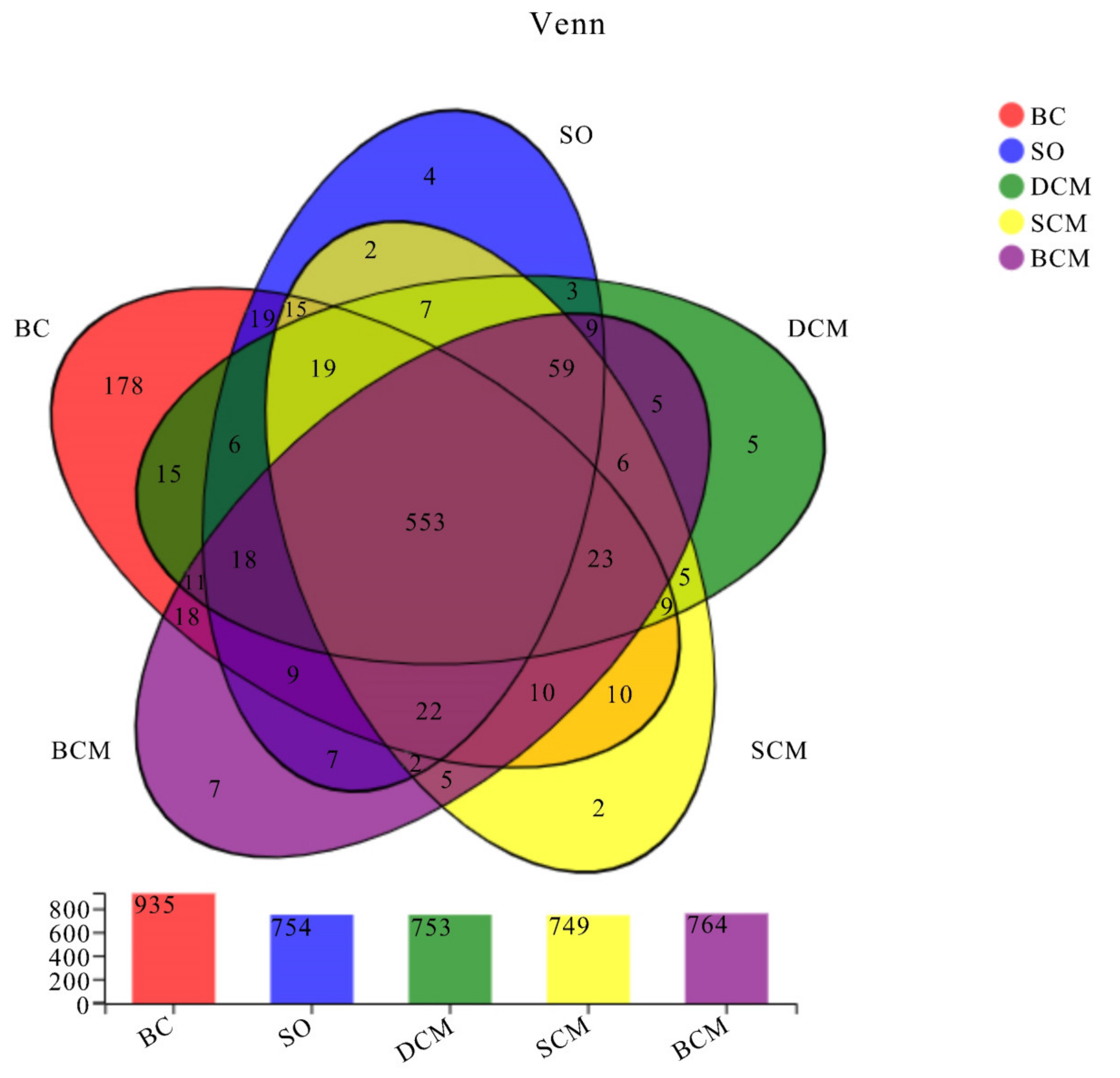
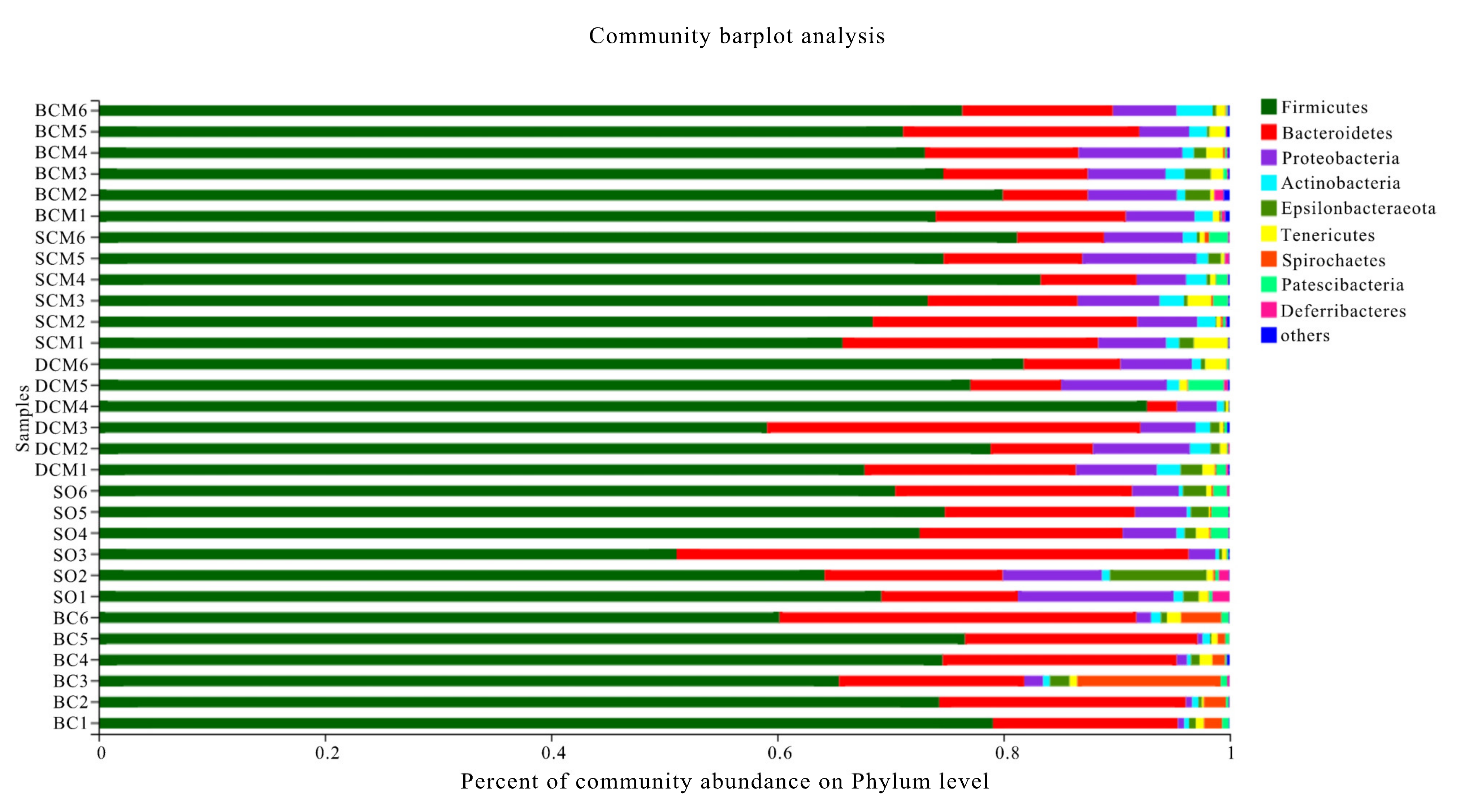
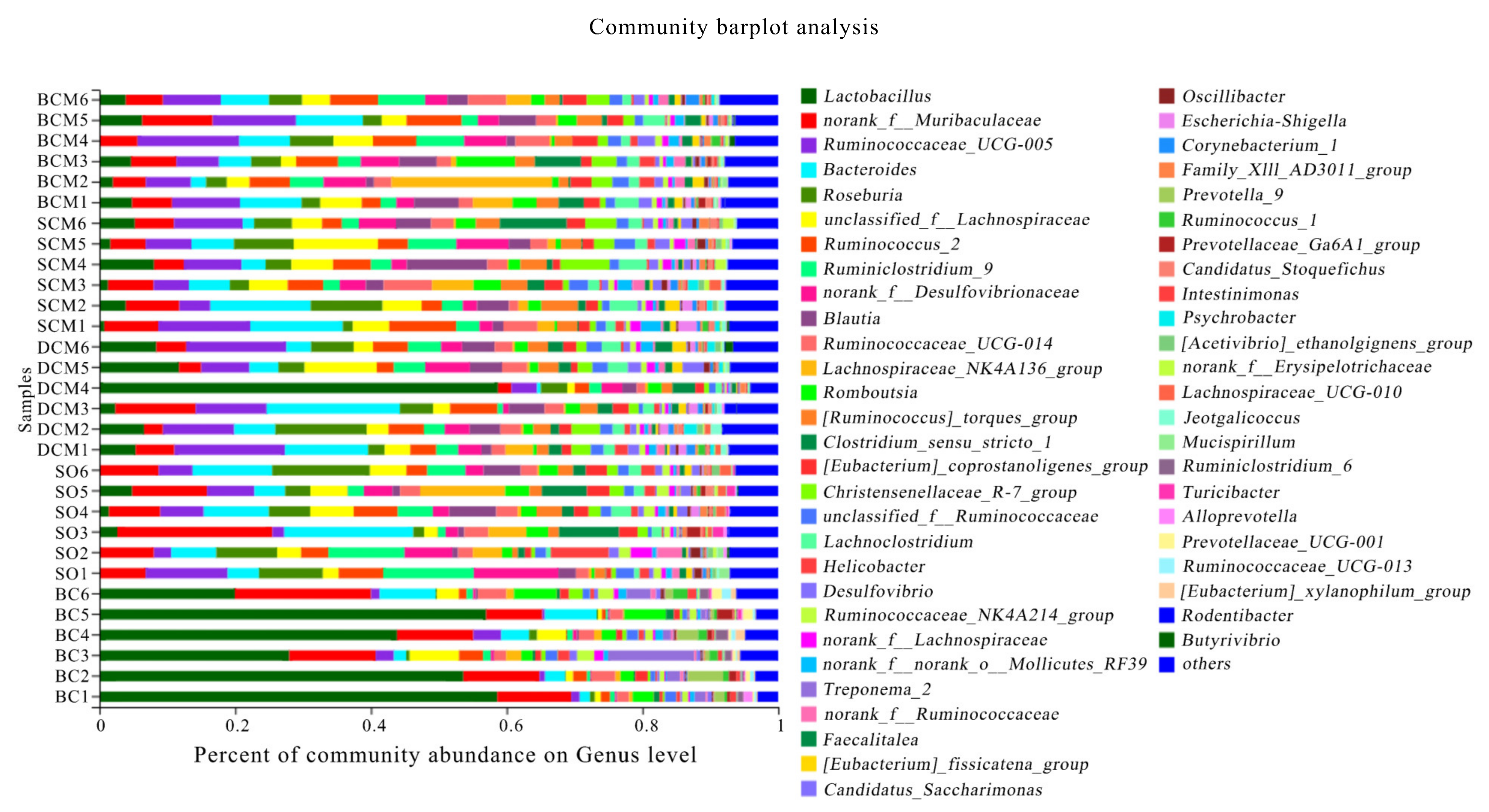
3.6. Effects of Pork Fat Cooked Using Different Methods on the Rat Gut Microbiota Structure and Abundance of Bacteria
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.C.; Li, H.J.; Wu, Z.M.; Li, F. Comparison of fat content and fatty acid composition in different parts of Rongchang pork. Food Sci. 2011, 32, 216–220. [Google Scholar]
- Broncano, J.M.; Petron, M.J.; Parra, V.; Timon, M.L. Effect of different cooking methods on lipid oxidation and formation of free cholesterol oxidation products (COPs) in Latissimus dorsi muscle of Iberian pigs. Meat Sci. 2009, 83, 431–437. [Google Scholar] [CrossRef]
- Huang, Y.C.; Li, H.J.; Qin, G.; Wang, T. Effects of different processing methods and time on fat content and fatty acid composition of pork. Sci. Tech. Food Ind. 2012, 1, 159–163. [Google Scholar]
- Li, Y.; Li, C.; Zhao, F.; Lin, X.; Bai, Y.; Zhou, G. The effects of long-duration stewing combined with different cooking and heating methods on the quality of pork belly. J. Food Process. Pres.. 2016, 40, 94–102. [Google Scholar] [CrossRef]
- Rasinska, E.; Rutkowska, J.; Czarniecka-Skubina, E.; Tambor, K. Effects of cooking methods on changes in fatty acids contents, lipid oxidation and volatile compounds of rabbit meat. Lebensm. Wiss. Technol. 2019, 110, 64–70. [Google Scholar] [CrossRef]
- Hu, L.; Ren, S.; Shen, Q.; Chen, J.; Ye, X.; Ling, J. Proteomic study of the effect of different cooking methods on protein oxidation in fish fillets. RSC Adv. 2017, 7, 27496–27505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; He, H.; Xiong, Z.B. The effect of cooking methods on pork quality and nutritional components. Food Mach. 2018, 6, 21–25. [Google Scholar]
- Russell, F.D.; Burgin-Maunder, C.S. Distinguishing health benefits of eicosapentaenoic and docosahexaenoic acids. Mar. Drugs 2012, 10, 2535–2559. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.H.; Xu, J.X.; Peng, Z.Y. The latest research on the nutrition and function of fatty acids. China Oils Fats 2000, 25, 20–23. [Google Scholar]
- Webb, E.C.; ONeill, H.A. The animal fat paradox and meat quality. Meat Sci. 2008, 80, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cao, J.; Dai, B.; Jiang, W.; Yang, Y.; Wen, D. Changes of fatty acids composition in beef under different thermal treatment. J. Food Nutr. Res. 2015, 3, 303–310. [Google Scholar]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Xu, Y. Lipid Changes during Braised Pork Stew and Its Effect on the Intestinal Flora of Rats. Master’s Thesis, Yangzhou University, Yangzhou, China, 2018. [Google Scholar]
- Escobar, J.S.; Klotz, B.; Valdes, B.E.; Agudelo, G.M. The gut microbiota of Colombians differs from that of Americans, Europeans and Asians. BMC Microbiol. 2014, 14, 311. [Google Scholar] [CrossRef] [Green Version]
- Carmody, R.N.; Bisanz, J.E.; Bowen, B.P.; Maurice, C.F.; Lyalina, S.; Louie, K.B.; Treen, D.; Chadaideh, K.S.; Rekdal, V.M.; Bess, E.N.; et al. Cooking shapes the structure and function of the gut microbiome. Nat. Microbiol. 2019, 4, 2052–2063. [Google Scholar] [CrossRef]
- Caesar, R.; Nygren, H.; Oresic, M.; Backhed, F. Interaction between dietary lipids and gut microbiota regulates hepatic cholesterol metabolism. J. Lipid Res. 2016, 57, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, M.; Pang, X.; Zhao, Y.; Wang, L.; Zhao, L. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 2012, 6, 1848–1857. [Google Scholar] [CrossRef]
- Ghosh, S.; Molcan, E.; Decoffe, D.; Dai, C.; Gibson, D.L. Diets rich in n-6 PUFA induce intestinal microbial dysbiosis in aged mice. Br. J. Nutr. 2013, 110, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmody, R.N.; Dannemann, M.; Briggs, A.W.; Nickel, B.; Groopman, E.E.; Wrangham, R.W.; Kelso, J. Genetic evidence of human adaptation to a cooked diet. Genome Biol. Evol. 2016, 8, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Z.; Yan, S.Y.; Xu, Y.; Wang, Q.Y.; Zhang, M.; Zhang, H.M.; Zhou, X.Y.; Yang, Z.P. Analysis of volatile flavor components of braised pork with different cooking time by headspace solid phase microextraction and gas mass spectrometry. Food Ferment. Ind. 2021, 47, 247–253. [Google Scholar]
- Zhu, W.Z.; Xu, Y.; Liu, W.; Wang, Q.Y.; Sha, W.X.; Zhou, X.Y. The effect of cooking time on the nutritional quality and volatile flavor compounds of lion head. Food Ferment. Ind. 2021, 47, 208–214. [Google Scholar]
- Yu, W.J.; Hou, J.W.; Zhu, B.S. Accurate determination of 14 fatty acids in pork by external standard-gas chromatography-mass spectrometry. Anal. Instrum. 2012, 3, 10–16. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Peng, C.; Gathagan, R.J.; Covell, D.J.; Medellin, C.; Stieber, A.; Robinson, J.L.; Zhang, B.; Pitkin, R.M.; Olufemi, M.F.; Luk, K.C.; et al. Cellular milieu imparts distinct pathological α-synuclein strains in α-synucleinopathies. J. Nat. 2018, 557, 558–563. [Google Scholar] [CrossRef]
- Gu, W.G.; Zhang, J.J.; Yao, Y.J.; Ji, R.; Ye, X.Q.; Chen, J.C. Fat oxidation and fatty acid composition changes during braised pork production. Food Sci. 2011, 17, 76–80. [Google Scholar]
- Liu, D.Y.; Tan, Y.; Gai, S.M.; Feng, N.; Guan, D.X.; Zhang, H.L. Changes of fat and fatty acid during braised pork belly. Food Sci. 2015, 36, 28–32. [Google Scholar]
- Duran-Montge, P.; Realini, C.E.; Barroeta, A.C.; Lizardo, R.G.; Esteve-Garcia, E. De novo fatty acid synthesis and balance of fatty acids of pigs fed different fat sources. J. Livest. Sci. 2010, 132, 157–164. [Google Scholar] [CrossRef]
- Gomes, M.B.; Negrato, C.A. Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. J. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Jiang, F.; Cheng, R.; Luo, Y.; Wang, J.; Luo, Z.; Li, M.; Shen, X.; He, F. A high-fat diet and high-fat and high-cholesterol diet may affect glucose and lipid metabolism differentially through gut microbiota in mice. J. Exp. Anim. 2021, 70, 73–83. [Google Scholar] [CrossRef]
- Qiu, B.F.; Zhao, T.P.; Liu, S.Y.; Dong, D.C. Effects of diets with different fatty acid composition on blood lipids and glucose metabolism in rats. Asia Pac. Tradit. Med. 2008, 4, 18–20. [Google Scholar]
- Campbell, C.L.; Yu, R.; Li, F.; Zhou, Q.; Chen, D.; Qi, C.; Yin, Y.; Sun, J. Modulation of fat metabolism and gut microbiota by resveratrol on high-fat diet-induced obese mice. J. Diabetes Metab. Syndr. Obes. 2019, 2, 97–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Araujo, T.V.; Andrade, E.F.; Lobato, R.V.; Orlando, D.R.; Gomes, N.F.; de Sousa, R.V.; Zangeronimo, M.G.; Pereira, L.J. Effects of beta-glucans ingestion (Saccharomyces cerevisiae) on metabolism of rats receiving high-fat diet. J. Anim. Physiol. Anim. Nutr. 2017, 101, 349–358. [Google Scholar] [CrossRef]
- Yin, C.N.; Shi, Y.H.; Wang, Y.Y.; Wang, S.P.; Le, G.W. Effects of lard, tea oil and oxidized tea oil on antioxidant capacity and blood lipid metabolism in mice. Acta Nutr. Sin. 2008, 30, 624–626. [Google Scholar]
- Zou, W.I. Effects of Different Diets on Intestinal Metabolites and Appetite Regulating Factors in SD rats. Ph.D. Thesis, Central South University, Changsha, China, 2014. [Google Scholar]
- Li, Y.Q. Study on the Quality Changes of Pork Fat During the Stewing Process and the Mechanism of Its Effects on the Physiological Functions of Rats. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Janssen, C.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef]
- Dimova, L.G.; Zlatkov, N.; Verkade, H.J.; Uhlin, B.E.; Tietge, U. High-cholesterol diet does not alter gut microbiota composition in mice. J. Nutr. Metab. 2017, 14, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosqvist, F.; Iggman, D.; Kullberg, J.; Cedernaes, J.; Johansson, H.E.; Larsson, A.; Johansson, L.; Ahlström, H.; Arner, P.; Dahlman, I.; et al. Overfeeding polyunsaturated and saturated fat causes distinct effects on liver and visceral fat accumulation in humans. Diabetes 2014, 63, 2356–2368. [Google Scholar] [CrossRef] [Green Version]
- Charlotte, G.; Leanne, H. The influence of dietary fat on liver fat accumulation. Nutrients 2014, 6, 5018–5033. [Google Scholar]
- Davide, F.A.M.; Di Natale, C.; D’Amico, A. Self-organizing multisensor systems for odour classification: Internal categorization, adaptation and drift rejection. Sens. Actuators B Chem. 1994, 18, 244–258. [Google Scholar] [CrossRef]
- Cheng, M.J. The Relationship between Liver Endoplasmic Reticulum Stress and Diet-Induced Insulin Resistance and Liver Steatosis in Obese Rats. Ph.D. Thesis, Huazhong University of Science and Technology, Wuhan, China, 2012. [Google Scholar]
- Hyotylainen, T.; Oresic, M. Analytical lipidomics in metabolic and clinical research. Trends Endocrinol. Metabol. 2015, 26, 671–673. [Google Scholar] [CrossRef]
- Jordan, S.; Tung, N.; Casanova-Acebes, M.; Chang, C.; Cantoni, C.; Zhang, D.; Wirtz, T.H.; Naik, S.; Rose, S.A.; Brocker, C.N.; et al. Dietary intake regulates the circulating inflammatory monocyte pool. Cell 2019, 178, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Lee, E.K.; Choi, Y.J.; Kim, J.M.; Kim, D.H.; Zou, Y.; Kim, C.H.; Lee, J.; Kim, H.S.; Kim, N.D.; et al. Molecular inflammation as an underlying mechanism of the aging process and age-related diseases. J. Dent. Res. 2011, 90, 830–840. [Google Scholar] [CrossRef]
- Jurk, D.; Wilson, C.; Passos, F.J.; Oakley, F.; Correia-Melo, C. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat. Commun. 2014, 2, 4172. [Google Scholar] [CrossRef]
- Il’Yasova, D.; Ivanova, A.; Morrow, J.D.; Cesari, M.; Pahor, M. Correlation between two markers of inflammation, serum C-reactive protein and interleukin 6, and indices of oxidative stress in patients with high risk of cardiovascular disease. J. Biomark. 2008, 13, 41–51. [Google Scholar] [CrossRef]
- Byrne, C.D. Fatty liver: Role of inflammation and fatty acid nutrition. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 265–271. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.A.; Goodwin, J.S. Immunology of Aging; Springer: New York, NY, USA, 1997. [Google Scholar]
- Enos, R.; Davis, J.M.; Velazquez, K.; McClellan, J.; Carnesville, K.; Murphy, E.A. Influence of dietary saturated fat content on adiposity, macrophage behavior, inflammation, and metabolism: Composition matters. J. Lipid Res. 2013, 54, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, K.; Manabe, I.; Oishi-Tanaka, Y.; Ohsugi, M.; Kono, N.; Ogata, F.; Yagi, N.; Ohto, U.; Kimoto, M.; Miyake, K.; et al. Saturated Fatty Acid and TLR Signaling Link β Cell Dysfunction and Islet Inflammation. J. Cell Metabolism. 2012, 15, 518–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachithanandan, N.; Fam, B.C.; Fynch, S.; Dzamko, N.; Watt, M.J.; Wormald, S.; Honeyman, J.; Galic, S.; Proietto, J.; Andrikopoulos, S.; et al. Liver-specific suppressor of cytokine signaling-3 deletion in mice enhances hepatic insulin sensitivity and lipogenesis resulting in fatty liver and obesity. J. Hepatol. 2010, 52, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Senn, J.J.; Klover, P.J.; Nowak, I.A.; Zimmers, T.A.; Koniaris, L.G.; Furlanetto, R.W.; Mooney, R.A. Suppressor of Cytokine Signaling-3 (SOCS-3), a Potential Mediator of Interleukin-6-dependent Insulin Resistance in Hepatocytes. J. Biol. Chem. 2003, 278, 13740–13746. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.I.; Ijaz, M.U.; Hussain, M.; Haq, I.; Zhao, D.; Li, C.B. High-fat proteins drive dynamic changes in gut microbiota, hepatic metabolome, and endotoxemia-TLR-4-NF kappa B-mediated inflammation in mice. J. Agric. Food Chem. 2020, 68, 11710–11725. [Google Scholar] [CrossRef]
- Xu, P.; Li, M.; Zhang, J.; Zhang, T. Correlation of intestinal microbiota with overweight and obesity in Kazakh school children. BMC Microbiol. 2012, 12, 283. [Google Scholar] [CrossRef] [Green Version]
- Power, S.E.; Toole, P.W.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. Intestinal microbiota, diet and health. Br. J. Nutr. 2014, 111, 387–402. [Google Scholar] [CrossRef]
- Yan, Y.L.; Ha, W.Y.; Campbell, C.R.; Mitchell, A.J.; Dinudom, A.; Oscarsson, J.; Cook, D.I.; Hunt, N.H.; Caterson, I.D.; Holmes, A.J.; et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS ONE 2012, 7, e34233. [Google Scholar]
- Huang, X.Y.; Zeng, Y.Y. Progress in the pathophysiology of intestinal microbiota. Chin. J. Pathophysiol. 2014, 30, 1127–1135. [Google Scholar]
- Zou, F. Study on the Effects of Different Types of Fats on the Growth Performance and Microecological Effects of Rats. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2009. [Google Scholar]
- Fang, Z.G. Effects of Diets with Different Fat Sources on Piglet Intestinal Development and Intestinal Microflora. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2012. [Google Scholar]
- Sefcikova, Z.; Bujnakova, D. Effect of pre- and post-weaning high-fat dietary manipulation on intestinal microflora and alkaline phosphatase activity in male rats. Physiol. Res. 2017, 66, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Brinkworth, G.D.; Noakes, M.; Clifton, P.M.; Bird, A.R. Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br. J. Nutr. 2009, 101, 1493–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wit, N.D.; Derrien, M.; Bosch, V.H.; Oosterink, E.; Meer, V.D. Saturated fat stimulates obesity and hepatic steatosis and affects gut microbiota composition by an enhanced overflow of dietary fat to the distal intestine. Am. J. Physiol. Gastrointest. 2012, 303, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.Y.; Wan, Y.P.; Fang, Q.Y.; Lu, W.; Zhang, H.B.; Cai, W. The effect of high-fat diet on intestinal probiotics and liver fat content in rats. Acta Nutr. Sin. 2011, 4, 367–369. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Miele, L.; Marrone, G.; Lauritano, C.; Cefalo, C.; Gasbarrini, A.; Day, C.; Grieco, A. Gut-liver axis and microbiota in NAFLD: Insight pathophysiology for novel therapeutic target. Curr. Pharm. Des. 2013, 19, 5314–5324. [Google Scholar] [CrossRef]
- Kübeck, R.; Bonet-Ripoll, C.; Hoffmann, C.; Walker, A.; Müller, V.; Schüppel, V.; Lagkouvardos, I.; Scholz, B.; Engel, K.-H.; Daniel, H.; et al. Dietary fat and gut microbiota interactions determine diet-induced obesity in mice. J. Mol. Metab. 2016, 5, 1162–1174. [Google Scholar] [CrossRef]
- Wan, J.; Hu, S.; Jacoby, J.J.; Liu, J.; Zhang, Y.; Yu, L. The impact of dietary sn-2 palmitic triacylglycerols in combination with docosahexaenoic acid or arachidonic acid on lipid metabolism and host faecal microbiota composition in Sprague Dawley rats. Food Funct. 2017, 8, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cui, L.H. The relationship between hyperlipidemia, high-fat diet and intestinal flora. World Chin. J. Digest. 2013, 14, 1273–1277. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | BCM | SCM | DCM | SO | BC |
|---|---|---|---|---|---|
| Casein | 200.0 | 200.0 | 200.0 | 200.0 | 180.3 |
| Corn starch | 397.4 | 397.4 | 397.4 | 397.4 | 519.8 |
| Dextrin | 132.0 | 132.0 | 132.0 | 132.0 | 119.0 |
| Sucrose | 100.0 | 100.0 | 100.0 | 100.0 | 90.1 |
| Pork fat of the BCM | 70.0 | - | - | - | - |
| Pork fat of the SCM | - | 70.0 | - | - | - |
| Pork fat of the DCM | - | - | 70.0 | - | - |
| Soybean oil | - | - | - | 70.0 | - |
| Crude fat | 40.0 | ||||
| Cellulose | 50.0 | 50.0 | 50.0 | 50.0 | 45.0 |
| Minerals | 35.0 | 35.0 | 35.0 | 35.0 | 31.5 |
| Vitamins | 10.0 | 10.0 | 10.0 | 10.0 | 9.0 |
| L-cystine | 3.0 | 3.0 | 3.0 | 3.0 | 2.7 |
| Choline chloride | 2.5 | 2.5 | 2.5 | 2.5 | 2.2 |
| Tert-butyl hydrogen | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| Total (g) | 1000 | 1000 | 1000 | 1000 | 1000 |
| Total energy (kcal/g) | 3.7 | 3.7 | 3.7 | 3.7 | 3.7 |
| Protein (%) | 19.4 | 19.4 | 19.4 | 19.4 | 19.4 |
| Carbohydrate (%) | 63.6 | 63.6 | 63.6 | 63.6 | 63.6 |
| Fat (%) | 16.9 | 16.9 | 16.9 | 16.9 | 9.67 |
| Fatty Acid | BCM | SCM | DCM | SO |
|---|---|---|---|---|
| C10:0 | 0.16 ± 0.03 a | 0.15 ± 0.03 a | 0.21 ± 0.03 a | — |
| C12:0 | 0.07 ± 0.02 b | 0.26 ± 0.04 a | 0.16 ± 0.02 ab | — |
| C16:0 | 69.05 ± 1.38 c | 133.70 ± 0.95 a | 103.24 ± 0.35 b | 11.04 ± 0.15 d |
| C17:0 | 0.64 ± 0.14 b | 2.82 ± 0.26 a | 1.10 ± 0.05 b | 0.05 ± 0.005 c |
| C18:0 | 38.33 ± 4.72 c | 91.64 ± 2.08 a | 54.67 ± 0.25 b | 4.67 ± 0.18 d |
| C20:0 | 0.85 ± 0.15 b | 2.18 ± 0.23 a | 1.59 ± 0.01 a | 0.40 ± 0.005 c |
| C14:1 | 4.03 ± 0.81 b | 12.82 ± 1.05 a | 7.41 ± 0.49 b | 0.01 ± 0.00 d |
| C16:1 | 5.35 ± 1.07 b | 21.15 ± 1.58 a | 17.44 ± 0.17 a | 2.12 ± 0.01 b |
| C17:1 | 0.35 ± 0.08 c | 1.68 ± 0.12 a | 0.87 ± 0.01 b | 0.03 ± 0.00 d |
| C18:1 | 94.66 ± 1.93 b | 174.39 ± 1.09 a | 151.29 ± 10.75 a | 22.21 ± 0.23 d |
| C20:1 | 3.27 ± 0.50 b | 8.84 ± 0.75 a | 4.64 ± 0.02 b | 0.18 ± 0.01 c |
| C18:2 | 58.27 ± 2.81 b | 103.85 ± 1.07 a | 96.48 ± 6.72 a | 50.77 ± 1.83 b |
| C20:2 | 2.98 ± 0.48 b | 7.82 ± 0.68 a | 3.91 ± 0.00 b | — |
| C18:3 | 0.58 ± 0.14 a | 1.37 ± 0.14 a | 0.52 ± 0.39 a | 6.11 ± 0.23 b |
| C20:4 | 1.30 ± 0.23 c | 3.31 ± 0.30 a | 2.30 ± 0.04 b | — |
| C22:4 | 0.08 ± 0.01 b | 0.55 ± 0.05 a | 0.18 ± 0.01 b | — |
| C20:5 | 0.25 ± 0.00 b | 0.52 ± 0.08 a | 0.50 ± 0.03 a | — |
| SFAs | 109.09 ± 6.45 c | 222.74 ± 3.59 a | 160.97 ± 0.54 b | 16.63 ± 0.35 d |
| MUFAs | 107.66 ± 4.40 c | 218.87 ± 4.61 a | 181.66 ± 10.40 b | 24.54 ± 0.21 d |
| PUFAs | 63.46 ± 3.67 b | 117.43 ± 2.33 a | 103.90 ± 7.13 a | 57.91 ± 1.05 c |
| UFAs | 171.12 ± 8.08 b | 336.30 ± 6.93 a | 285.56 ± 17.53 a | 81.45 ± 2.27 d |
| UFAs/SFAs | 1.57 ± 0.03 b | 1.51 ± 0.01 b | 1.77 ± 0.10 a | 5.35 ± 0.21 a |
| Lipid | Day | BCM | DCM | SCM | SO | BC |
|---|---|---|---|---|---|---|
| TG (mmol/L) | 0 | 0.74 ± 0.16 d | 0.84 ± 0.15 c | 0.53 ± 0.09 d | 0.68 ± 0.11 b | 0.81 ± 0.12 d |
| 14 | 1.37 ± 0.16 bc | 1.38 ± 0.31 b | 1.23 ± 0.14 bc | 1.36 ± 0.14 a | 1.19 ± 0.24 ab | |
| 28 | 1.07 ± 0.22 c | 0.86 ± 0.32 c | 0.86 ± 0.22 cd | 1.12 ± 0.14 ab | 0.87 ± 0.14 bc | |
| 42 | 1.65 ± 0.11 b | 1.21 ± 0.33 bc | 1.60 ± 0.29 b | 1.72 ± 0.61 a | 1.14 ± 0.41 abc | |
| 56 | 2.26 ± 0.65 a | 2.22 ± 0.35 a | 2.2 ± 0.85 a | 1.74 ± 0.74 a | 1.35 ± 0.26 a | |
| TC (mmol/L) | 0 | 2.36 ± 0.17 a | 2.42 ± 0.21 a | 2.32 ± 0.17 a | 2.08 ± 0.52 a | 1.63 ± 0.47 a |
| 14 | 1.98 ± 0.14 b | 1.94 ± 0.39 bc | 1.90 ± 0.18 bc | 2.15 ± 0.18 a | 1.84 ± 0.16 a | |
| 28 | 1.46 ± 0.13 c | 1.68 ± 0.48 c | 1.55 ± 0.10 d | 1.37 ± 0.12 b | 1.50 ± 0.16 a | |
| 42 | 1.85 ± 0.14 b | 1.81 ± 0.25 c | 1.72 ± 0.16 cd | 1.64 ± 0.20 b | 1.47 ± 0.12 a | |
| 56 | 2.22 ± 0.23 a | 2.33 ± 0.20 ab | 1.96 ± 0.13 b | 2.12 ± 0.22 a | 1.52 ± 0.21 a | |
| HDL-C (mmol/L) | 0 | 0.94 ± 0.02 a | 0.97 ± 0.05 a | 0.94 ± 0.10 a | 0.92 ± 0.08 a | 0.84 ± 0.06 a |
| 14 | 0.70 ± 0.02 b | 0.73 ± 0.13 b | 0.76 ± 0.07 b | 0.72 ± 0.06 b | 0.69 ± 0.09 b | |
| 28 | 0.57 ± 0.12 c | 0.60 ± 0.06 c | 0.64 ± 0.06 c | 0.57 ± 0.04 c | 0.55 ± 0.07 c | |
| 42 | 0.60 ± 0.05 c | 0.61 ± 0.05 c | 0.62 ± 0.05 c | 0.56 ± 0.08 c | 0.52 ± 0.06 c | |
| 56 | 0.62 ± 0.06 bc | 0.66 ± 0.06 bc | 0.57 ± 0.06 c | 0.62 ± 0.03 c | 0.47 ± 0.04 c | |
| LDL-C (mmol/L) | 0 | 0.43 ± 0.08 c | 0.45 ± 0.09 b | 0.37 ± 0.05 b | 0.42 ± 0.03 c | 0.44 ± 0.06 c |
| 14 | 0.34 ± 0.09 d | 0.27 ± 0.06 c | 0.22 ± 0.01 c | 0.35 ± 0.05 d | 0.40 ± 0.02 c | |
| 28 | 0.77 ± 0.04 b | 0.84 ± 0.09 a | 0.80 ± 0.03 a | 0.76 ± 0.06 b | 0.89 ± 0.04 a | |
| 42 | 0.88 ± 0.02 a | 0.85 ± 0.03 a | 0.79 ± 0.02 a | 0.84 ± 0.06 a | 0.89 ± 0.09 a | |
| 56 | 0.88 ± 0.05 a | 0.89 ± 0.07 a | 0.79 ± 0.03 a | 0.86 ± 0.07 a | 0.78 ± 0.06 b |
| BCM (%) | SCM (%) | DCM (%) | BC (%) | SO (%) | |
|---|---|---|---|---|---|
| Firmicutes | 74.72 ± 2.67 a | 74.40 ± 6.24 a | 76.14 ± 10.63 a | 71.75 ± 6.71 a | 66.84 ± 7.73 b |
| Proteobacteria | 6.71 ± 1.52 a | 6.68 ± 1.82 a | 6.64 ± 1.95 a | 0.92 ± 0.46 b | 6.36 ± 3.77 a |
| Bacteroidetes | 14.13 ± 4.10 b | 14.54 ± 6.16 b | 13.36 ± 9.98 b | 21.10 ± 5.14 a | 21.65 ± 10.87 a |
| Deferribacteres | 0.23 ± 0.27 ab | 0.13 ± 0.13 ab | 0.12 ± 0.13 ab | 0.05 ± 0.07 b | 0.47 ± 0.57 a |
| Epsilonbacteraeota | 1.03 ± 0.93 a | 0.57 ± 0.49 a | 0.70 ± 0.66 a | 0.68 ± 0.52 a | 2.49 ± 2.76 a |
| Tenericutes | 1.05 ± 0.34 a | 1.17 ± 1.10 a | 0.82 ± 0.56 a | 0.84 ± 0.39 a | 0.61 ± 0.34 a |
| Actinobacteria | 1.65 ± 0.75 a | 1.52 ± 0.39 a | 1.31 ± 0.50 a | 0.62 ± 0.23 b | 0.56 ± 0.20 b |
| Patescibacteria | 0.14 ± 0.11 a | 0.74 ± 0.65 a | 0.77 ± 1.10 a | 0.43 ± 0.20 a | 0.83 ± 0.59 a |
| Spirochaetes | 0.06 ± 0.07 b | 0.13 ± 0.15 b | 0.05 ± 0.05 b | 3.53 ± 4.07 a | 0.12 ± 0.06 b |
| BCM (%) | SCM (%) | DCM (%) | BC (%) | SO (%) | |
|---|---|---|---|---|---|
| Ruminococcaceae_UCG-005 | 9.31 ± 1.26 a | 7.67 ± 1.07 a | 10.74 ± 1.57 a | 2.24 ± 0.75 b | 6.52 ± 2.02 a |
| Bacteroides | 7.70 ± 1.14 a | 3.34 ± 2.24 b | 7.27 ± 1.95 a | 2.62 ± 0.60 b | 8.16 ± 1.58 a |
| Roseburia | 3.70 ± 0.55 bc | 4.39 ± 1.09 b | 4.98 ± 0.92 b | 0.64 ± 0.22 c | 9.73 ± 1.73 a |
| Lactobacillus | 4.92 ± 0.50 b | 2.40 ± 0.57 c | 7.14 ± 2.18 b | 46.24 ± 6.75 a | 0.42 ± 0.33 b |
| Lachnospiraceae_NK4A136_group | 2.66 ± 1.29 ab | 4.55 ± 0.60 a | 0.32 ± 0.03 b | 1.18 ± 0.38 b | 1.38 ± 0.98 b |
| Blautia | 5.15 ± 0.74 a | 2.36 ± 0.71 ab | 5.57 ± 2.10 a | 0.11 ± 0.05 b | 3.97 ± 1.38 a |
| Romboutsia | 4.24 ± 1.51 ab | 7.18 ± 1.78 a | 1.80 ± 0.29 b | 2.19 ± 0.31 b | 1.74 ± 0.50 b |
| Clostridium_sensu_stricto_1 | 2.92 ± 1.49 b | 5.51 ± 0.49 a | 1.75 ± 0.37 bc | 0.36 ± 0.17 c | 1.19 ± 0.40 bc |
| Ruminococcus_2 | 6.06 ± 1.17 a | 5.71 ± 2.12 a | 4.84 ± 0.17 ab | 1.78 ± 0.72 b | 5.06 ± 0.88 ab |
| Ruminiclostridium_9 | 3.73 ± 1.08 b | 4.04 ± 1.61 b | 2.78 ± 0.12 b | 0.80 ± 0.15 b | 8.83 ± 2.04 a |
| Ruminococcaceae_UCG-014 | 3.20 ± 0.85 a | 3.99 ± 1.06 a | 4.79 ± 1.95 a | 2.34 ± 0.43 a | 2.43 ± 0.27 a |
| norank_f__Muribaculaceae | 7.11 ± 1.11 b | 1.04 ± 0.29 d | 3.33 ± 0.93 c | 11.56 ± 0.42 a | 7.71 ± 0.41 b |
| unclassified_f__Lachnospiraceae | 4.03 ± 0.80 a | 2.08 ± 0.40 a | 8.42 ± 3.79 a | 3.33 ± 1.54 a | 4.41 ± 0.92 a |
| [Eubacterium]_coprostanoligenes_group | 2.48 ± 0.41 b | 1.08 ± 0.29 b | 1.88 ± 0.23 ab | 1.02 ± 0.29 ab | 1.77 ± 0.55 a |
| Christensenellaceae_R-7_group | 2.38 ± 0.67 c | 1.15 ± 0.27 c | 3.07 ± 0.23 c | 0.48 ± 0.08 b | 1.15 ± 0.31 b |
| norank_f__Desulfovibrionaceae | 3.67 ± 0.68 b | 0.74 ± 0.19 b | 3.56 ± 1.19 ab | 0.36 ± 0.11 ab | 6.14 ± 2.36 a |
| [Ruminococcus]_torques_group | 3.03 ± 0.31 b | 0.81 ± 0.41 b | 2.27 ± 0.35 a | — | 2.11 ± 0.62 a |
| Lachnoclostridium | 2.45 ± 0.52 c | 3.77 ± 1.13 bc | 1.30 ± 0.42 bc | — | 1.55 ± 0.52 a |
| Faecalitalea | 1.29 ± 0.35 d | 0.58 ± 0.11 c | 1.70 ± 0.20 bc | — | 0.73 ± 0.81 a |
| Desulfovibrio | 1.12 ± 0.42 b | — | 2.32 ± 0.65 b | 0.49 ± 0.21 b | 1.03 ± 0.18 a |
| unclassified_f__Ruminococcaceae | 1.53 ± 0.14 b | 4.83 ± 1.16 b | 1.24 ± 0.36 b | 1.41 ± 0.25 b | 1.86 ± 0.28 a |
| [Eubacterium]_fissicatena_group | — | 5.28 ± 2.28 | 0.79 ± 0.57 | — | 0.69 ± 0.30 |
| norank_f__Erysipelotrichaceae | — | 3.93 ± 2.32 | 0.24 ± 0.21 | 0.04 ± 0.01 | 0.17 ± 0.20 |
| norank_f__norank_o__Mollicutes_RF39 | — | 3.61 ± 0.35 | 0.65 ± 0.18 | 0.64 ± 0.34 | 0.69 ± 0.27 |
| Escherichia-Shigella | — | 2.25 ± 0.70 | 1.05 ± 0.59 | — | 0.42 ± 0.51 |
| norank_f__Peptococcaceae | — | 1.96 ± 2.43 | 0.59 ± 0.19 | 0.06 ± 0.02 | 0.32 ± 0.20 |
| Candidatus_Saccharimonas | — | 0.42 ± 0.12 | 1.02 ± 0.60 | 0.39 ± 0.20 | 0.84 ± 0.54 |
| Helicobacter | — | 0.60 ± 0.30 | 0.51 ± 0.04 | 0.85 ± 0.55 | 3.26 ± 3.08 |
| norank_f__Ruminococcaceae | — | 0.63 ± 0.31 | 0.11 ± 0.19 | 0.36 ± 0.07 | 1.38 ± 1.33 |
| Ruminococcaceae_NK4A214_group | — | — | 0.80 ± 0.26 | 1.24 ± 0.73 | 1.39 ± 0.29 |
| norank_f__Lachnospiraceae | — | 0.69 ± 0.10 | — | — | 1.24 ± 1.14 |
| Treponema_2 | — | — | — | 4.35 ± 4.84 | 0.14 ± 0.04 |
| Prevotella_9 | — | — | — | 2.60 ± 1.74 | — |
| Ruminiclostridium_6 | — | — | — | 1.00 ± 0.54 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Xu, Y.; Liu, J.; Chen, D.; Zhang, H.; Yang, Z.; Zhou, X. Effects of Dietary Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism, Liver Inflammation and Gut Microbiota in Rats. Foods 2021, 10, 3030. https://doi.org/10.3390/foods10123030
Zhu W, Xu Y, Liu J, Chen D, Zhang H, Yang Z, Zhou X. Effects of Dietary Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism, Liver Inflammation and Gut Microbiota in Rats. Foods. 2021; 10(12):3030. https://doi.org/10.3390/foods10123030
Chicago/Turabian StyleZhu, Wenzheng, Yan Xu, Jun Liu, Dawei Chen, Huimin Zhang, Zhangping Yang, and Xiaoyan Zhou. 2021. "Effects of Dietary Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism, Liver Inflammation and Gut Microbiota in Rats" Foods 10, no. 12: 3030. https://doi.org/10.3390/foods10123030
APA StyleZhu, W., Xu, Y., Liu, J., Chen, D., Zhang, H., Yang, Z., & Zhou, X. (2021). Effects of Dietary Pork Fat Cooked Using Different Methods on Glucose and Lipid Metabolism, Liver Inflammation and Gut Microbiota in Rats. Foods, 10(12), 3030. https://doi.org/10.3390/foods10123030