Abstract
In recent years, chronic diseases including obesity, diabetes, cancer, cardiovascular, and neurodegenerative disorders have been the leading causes of incapacity and death globally. Increasing evidence suggests that improvements of lifestyle habits and diet is the most commonly adopted strategy for the prevention of chronic disorders. Moreover, many dietary compounds have revealed health-promoting benefits beyond their nutritional effects. It is worth noting that diet plays an important role in shaping the intestinal microbiota. Coarse cereals constitute important sources of nutrients for the gut microbiota and contribute to a healthy gut microbiome. Furthermore, the gut microbiota converts coarse cereals into functional substances and mediates the interaction between the host and these components. In this study, we summarize the recent findings concerning functional components of cereal grains and their potential chemopreventive activity via modulating the gut microbiota.
1. Introduction
Coarse cereals are cereal grains except for rice and wheat, including oats (Avena sativa), buckwheat (Fagopyrum esculentum), barley (Hordeum vulgare), sorghum (Sorghum vulgare), quinoa (Chenopodium quinoa), and some millet species [1]. Due to their rich protein content and balanced amino acid profile, the nutritional value of coarse cereals has attracted wider attention than that of major cereals [2]. In addition to their nutritional value, the main bioactive substances (dietary fiber, starch, flavonoid, polyphenol, saponin, polysaccharide, protein, and peptides) of coarse cereals have been demonstrated to exert various biological functions. For instance, a previous study reported that coarse-cereal fiber had a positive effect on cardiometabolic and obesity-related diseases [3]. Idehen et al. found that barley phytochemicals (phytosterol, flavonoids, phenolic acids, etc.) can exert antioxidant, cholesterol-lowering, and anti-cancer functions to prevent cardiovascular diseases [4]. It has been reported that phytosterols play a crucial role in inhibiting the progression of cancers [5].
Dietary structure is a significant impact factor in intestinal microecology, which demonstrates a trend towards dynamic change along with changes in eating habits and trophic factor intake [6]. The roles that human colonic bacteria or the gut microbiota play in health and disease have received considerable attention over the last decade, supported by recent reports suggesting that the gut microbiota have the potential to treat many diseases that plague modern society, including obesity, diabetes, and colorectal cancer [7]. As previously described, the main microorganisms in the human intestine are closely related to the nutritional components and their proportions in the dietary structure [8]. A diet rich in fiber stimulates an increase in the number of Bifidobacteria in the human intestine [9]. Moreover, a high consumption of protein and fat is responsible for Bacteroides, and high saturated fatty acids (FAs) can promote an abundance of Bilophila wadsworthia [10].
Although the non-digestible components in coarse grains are considered poor for gut microbial fermentation, the bioactive compounds of coarse grains have the potential to modulate the gut microbiota and thereby impact on consumer health [11]. David et al. suggested that increasing the proportion of coarse grains in the dietary structure is an effective way to promote a diversity of probiotics in the intestinal flora and alleviate a variety of chronic diseases [12]. A bifidogenic effect has been observed after intake of whole-grain cereals, and some Bifidobacterium strains have been reported as markers of a healthy gut microbiota [11,13]. In addition, oat, barley husks, and rye bran were reported to increase the growth of Bifidobacteria or Lactobacillus, which exert anti-tumor potential and enhance the formation of short-chain fatty acids (SCFAs) such as acetate and butyrate [14,15]. Butyrate is an important metabolite of gut microbial fermentation of carbohydrates due to its contribution to host colonic epithelial cell energy, its anti-inflammatory properties, and its anti-cancer effects [16]. Whole grains have been shown to decrease protein fermentation by the gut microbiota and may also increase the diversity of the microbiota [17,18]. Recently, the impact of food matrix components including fiber, protein, lipid oxidation products, and emulsions, and their effects on gut and metabolic health, has gained much attention. In this review, how the main components of coarse grains exert their beneficial health characteristics by regulating intestinal flora and the underlying mechanisms involved will be discussed (Figure 1).
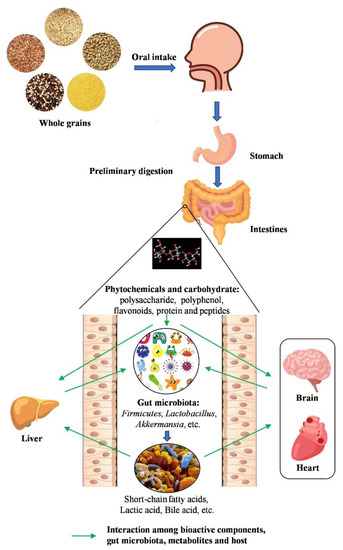
Figure 1.
The interaction between whole grains and gut microbiota. The bioactive components of whole grains, which are produced after oral, gastric, and small intestine digestion, improve gut health by modulating gut microbiota. Phytochemicals and carbohydrates are metabolized by gut microbiota into short-chain fatty acids (SCFAs) and other small functional molecules. These functional components can regulate gut microbiota that contribute to exerting health effects on various diseases.
2. Effects on Gastrointestinal Diseases
The gastrointestinal (GI) tract is the main interface for the interaction of dietary compounds and the organism. This complex system has developed multiple protective mechanisms against pathogens and toxic agents from the external environment, and it maintains a specialized balance of GI homeostasis, which is crucial for the digestive system to absorb nutrients and water [19]. However, poor lifestyle habits and imbalanced diet are the main reasons for the increasing incidence of gastrointestinal diseases such as inflammatory bowel disease (IBD), colorectal cancer (CRC), intestinal barrier dysfunction, etc.
Dietary interventions are not thought of as replacing agents in conventional medical therapies, but may be effective as complementary strategies for GI disorders [20]. Moreover, dietary molecules can influence the mechanisms of nutrient sensing and bioavailability, as well as the composition and metabolism of the gut microbiota, which ultimately might also affect GI homeostasis [21].
2.1. Inflammatory Bowel Disease
Recently, growing evidence points to the potential role of dietary bioactive compounds and microbiota modulation in IBD [22,23]. The effect of whole-grain components on regulating the gut microbiota is summarized in Table 1. For instance, intake of oat β-glucan significantly ameliorated the clinical features of ulcerative colitis in mice, attenuating the severe colonic inflammatory status and improving colonic mucosal barrier function concurrently with regulating gut-derived SCFAs and intestinal microbial metabolic profiles [24]. According to the authors, the reversed expression levels of zonula occludens-1 (ZO-1), occludin, claudin-1, and claudin-4 indicated that oat β-glucan promoted tight junction protein formation and restored intestinal epithelial barrier function. Moreover, they proved that pro-inflammatory cytokine interferon-γ (IFN-γ) and interleukin-6 (IL-6) were inhibited, while the production of SCFAs (especially acetate and propionate) increased, playing an important role in regulating intestinal epithelial tight junctions in UC mice. In a similar experiment, the low-molecular-weight form of β-glucan had a positive effect on the colon tissue of rats with lipopolysaccharide (LPS)-induced enteritis, resulting in changes in the levels of IL-10, IL-12, and tumor necrosis factor-α (TNF-α), as well as the number of intraepithelial lymphocytes (IELs) and lamina propria lymphocytes (LPLs) [25]. Moreover, a statistically significant reduction in the formation of hydroxybutyric acid and enhanced concentrations of lactic acid and propionic acid were observed, which was consistent with the number of lactic acid bacteria. This suggests an improvement in the alimentary tract contents.
Oat bran is one of the major byproducts in the processing of husked oat, and it contains relatively high levels of protein, minerals, vitamins, and soluble β-glucan [26]. He et al. investigated the effects of oat bran on nutrient digestibility, intestinal microbiota, and inflammatory responses in the hindgut of growing pigs. After supplementing with 10% oat bran (OB), the dietary nutrient digestibility showed no differences between the OB group and the control group on day 28. However, the mRNA expression of IL-8, nuclear factor-κB (NF-κB), and TNF-α was decreased in the OB group. Moreover, the abundances of Prevotella, Butyricicoccus, and Catenibacterium were increased in the colonic digesta, which may contribute to ameliorating inflammatory responses in the hindgut via enhancing the fermentation of fiber to produce SCFAs. Similarly, insoluble fiber in oat hulls could alleviate necrotic enteritis, and broilers fed with oat hulls had heavier gizzards compared with those without oat hulls. The gut microbiota composition was also improved by increasing the abundance of Clostridium perfringens and reducing the abundance of Lactobacillus and Salmonellae [27]. However, despite the fact that dietary fiber has been proved to be an effective strategy to prevent and relieve IBD through gut microbiota modulation, the function of soluble dietary fiber was found to be more favorable than that of insoluble dietary fiber. Tian et al. evaluated the bioactivity of insoluble dietary fiber from barley leaf (BLIDF) in inhibiting gut inflammation through regulating the intestinal microbiota in dextran sulfate sodium (DSS)-induced colitis mice. After intake of a BLIDF-supplemented diet for 28 days, they found that BLIDF markedly attenuated the severity of inflammation in DSS-induced acute colitis mice and increased the abundance of Parasutterella, Erysipelatoclostridium, and Alistipes, while the abundance of Akkermansia was significantly decreased. Moreover, BLIDF feeding can reverse the DSS-induced decline of short-chain fatty acids and secondary bile acids in mice feces [28]. Likewise, Li et al. explored the potential modulating roles of gut microbial metabolites of BL (barley leaf) in protecting against colitis and elucidated the underlying molecular mechanisms. They reported that supplementing with BL ameliorated DSS-induced gut microbiota dysbiosis and resulted in the enrichment of the microbiota-derived purine metabolite inosine, which could simulate peroxisome-proliferators-activated receptor γ (PPARγ) signaling to improve intestinal mucosal barrier functions in human colon epithelial cells [29].
It is also noteworthy that consumption of many purified dietary fibers has not been shown to increase the diversity of gut microbiota. The beneficial effects of whole grains on health are likely to be a combined result of many components within the grain rather than one specific component [30,31]. It was demonstrated that consumption of quinoa alleviated clinical symptoms according to the reduced disease activity index and the degree of histological damage. Compared to the control group, the species richness and diversity were increased in the quinoa consumption group, while abnormal expansion of the phylum Proteobacteria was decreased. Moreover, the overgrowth of the genera Escherichia/Shigella and Peptoclostridium was inhibited, whereas the abundances of Firmicutes and Bacteroidetes did not change significantly in the quinoa treatment group [32]. Another experiment investigated the effect of a buckwheat diet in a high-fat diet-induced gastritis mice model. The results showed that gastritis was lessened after intake of buckwheat, and the microbial dysbiosis induced by a high-salt diet could be recovered [33]. In the case of millet whole grain, its consumption by DSS-induced colitis murines alleviated the symptoms of enteritis to varying degrees, and completely alleviated DSS-induced dysbiosis. Moreover, the decreased levels of IL-6 and claudin2 expression in the colon indicated that systemic inflammation and gut barrier function was improved after the consumption of millet whole grain [34].
2.2. Colorectal Cancer
Epidemiological studies have suggested that IBD patients exhibit a higher risk of CRC and that the incidence of cancer is positively correlated with the duration of IBD [35]. A recent study suggested that coarse cereal diets are beneficial for relieving CRC. Zhang et al. reported that intake of foxtail millet ameliorated AOM/DSS-induced colitis-associated CRC in mice via the activation of AHR and GPCRs and the inhibition of STAT3 phosphorylation by the microbial metabolites of the foxtail millet. In line with a previous study, the abundances of Bifidobacterium and Bacteroidales_S24-7 were increased after millet consumption, compared to the control group [36]. Similarly, Yang et al. studied the effect of sorghum in an HFD-induced CRC mice model. The results showed that sorghum exhibited tremendous anti-CRC effects by suppressing the growth and metastasis of cancerous colon epithelial cells, as well as protecting against gut microbiota alterations linked to colitis [37].
Collectively, IBD, CRC, and intestinal barrier dysfunction could be alleviated by intake of a diet supplemented with coarse cereals or their bioactive components, due to the changes in relative gene expression, reduction of inflammatory cytokines (IL-6, IL-8, IL-10, etc.) and increased production of SCFAs demonstrated to be affected by the composition of gut microbiota.

Table 1.
Effect of whole-grain components on gut microbiota of gastrointestinal diseases host.
3. Effects on Cardiovascular Diseases
Hyperlipidemia is the main cause of cardiovascular diseases such as hypertension, atherosclerosis, stroke, etc. [40]. Hyperlipidemia is a chronic lipid disorder that causes abnormally increased levels of total cholesterol (TC), triglycerides (TG), and low-density lipoprotein cholesterol (LDL-C), and a decreased level of high-density lipoprotein cholesterol (HDL-C) [41]. Although hyperlipidemia can be controlled using pharmacological strategies, long-term exposure to drugs may lead to unpleasant side effects [42]. Therefore, natural lipid-lowering drugs without adverse effects have been considered the best alternatives against hyperlipidemia. Increasing evidence has indicated that the gut microbiota, which is involved in lipid absorption and liver cholesterol metabolism, is closely associated with hyperlipidemia [43,44].
3.1. Hyperlipidemia
Recent studies have proved that the consumption of some beneficial whole-grain products is associated with a lower risk of hyperlipidemia and has a close relationship with alterations in the gut microbiota [45]. The composition, content, and functional properties of phytochemicals in grains, including flavonoids, polysaccharides, polyphenols, and polypeptides, have been shown to possess a potential antihyperlipidemic effect by ameliorating lipid profiles and gut dysbiosis in vivo [46]. Here, we present a summary of the bioactive substances of whole grains in the regulation of gut microbiota (Table 2). In a high-fat diet (HFD)-induced hyperlipidemia mice model, flavonoids from whole-grain oat (FO) improved serum lipid profiles and decreased lipid deposition. RT-qPCR and Western blot analysis showed that FO suppressed lipogenesis and promoted lipolysis, BA synthesis, and efflux to feces through the FXR pathway. Moreover, 16s rRNA sequencing revealed that FO significantly increased Akkermansia and significantly decreased Lachnoclostridium, Blautia, Colidextribacter, and Desulfovibrio, which was correlated with hyperlipidemia-related parameters. Thus, FO exhibited hyperlipidemic inhibitory activity by regulating the gut–liver axis (BA metabolism and gut microbiota) [47]. Likewise, intake of embryo-remaining oat rice (EROR) dramatically improved the lipid profile in the serum and liver. Furthermore, EROR supplementation significantly increased the total SCFAs, acetate, and propionate and promoted the abundance of SCFA-producing bacteria. In addition, EROR consumption caused an alteration in the composition of the indigenous flora, with increases in Bifidobacterium and Akkermansia and decreases in Rombutsia, Fusicatenibacter, Holdemanella, and Turicibacter that were correlated with the lipid-metabolism-related indices [48].
Plant-derived polysaccharides are natural polysaccharides and are non-toxic with few side effects [49]. Cao et al. found that oral administration of quinoa polysaccharide mitigated dyslipidemia and reduced hepatic lipid accumulation by decreasing the contents of TG, LDL-C, glutamic oxaloacetic transaminase (AST), glutamic pyruvic transaminase (ALT), and malondialdehyde (MDA). Concurrently, the species richness and gut microbiota community structure were positively regulated by intake of quinoa polysaccharide. Furthermore, the results of 16s rRNA sequencing showed that the ratio of Firmicutes and Bacteroides (F/B ratio) and the abundance of Proteobacteria, Desulfovibrio, and Allobaculum were decreased, which was beneficial in mitigating intestinal inflammation and ameliorating the serum lipid levels. Thus, quinoa polysaccharide could reduce the numbers of potentially pathogenic bacteria to decrease infection, inflammation, and hyperlipidemia [50].
Compared to most cereals, the protein fractions of quinoa and buckwheat have high nutritive value. In the research of Fotschki and co-workers, the effects of quinoa and buckwheat protein-rich flours on the growth parameters, intestinal microbial activity, and plasma lipid profile were studied. The studied flours favorably reduced the plasma total cholesterol and LDL cholesterol, increased the levels of plasma IL-6 and alanine transaminase, and regulated the microbial production of SCFAs in rats [51]. Among these SCFAs, butyric acid was significantly elevated. This is the preferred energy source for colon epithelial cells and could decrease the pH together with other SCFAs [52]. Moreover, butyric acid is able to activate AhR and negatively regulates several enzymes, including fatty-acid synthase and a cholesterol metabolism regulator in the form of sterol regulatory element-binding protein-1c [53]. In a similar experiment, male C57BL/6 mice were fed an HFD with buckwheat protein (BWP) to evaluate the effect of BWP in attenuating dyslipidemia. The levels of TC, TG, LPS, TNF-α, and IL-6 in the mice fed on the HFD with BWP were significantly lower than those on an HFD with casein. In addition, BWP markedly increased the total bile acids and short-chain fatty acids, and also improved the abundances of Lactobacillus, Bifidobacterium, and Enterococcus, and decreased the abundance of Escherichia coli in HFD-fed mice. Notably, the Bifidobacterium population was closely related to the plasma lipids content [54]. A previous study reported that the presence of specific prebiotics in buckwheat may stimulate the growth of probiotics and other beneficial gut bacteria present in the colon and help to maintain healthy gut conditions [55]. Zhou et al. explored the effect of buckwheat fermented milk on the intestinal flora and SCFAs of HFD-fed rats. The buckwheat fermented milk remarkably repressed the increases in LPS levels in the colon and the antioxidant indexes in HFD-fed rats. In addition, buckwheat fermented milk significantly enhanced the abundance of the genus Akkermansia and decreased the abundance of the Lachnospiraceae family [56]. It has been proved that the species belonging to Lachnospiraceae are linked to the development of obesity and type 2 diabetes in ob/ob rats, whereas Akkermansia, belonging to Verrucomicrobia, plays an important role in treating obesity, inflammation, type 2 diabetes, and metabolic syndrome [57,58].
There has been growing evidence suggesting that intake of oat and buckwheat is associated with a reduction in serum cholesterol [59,60]. Zhou and co-workers reported that whole-grain oat (WGO) flour improved insulin sensitivity and the plasma cholesterol profile compared with low-bran oat (LBO) flour, resulting in lower plasma insulin, C-peptide, total cholesterol, and non-HDL cholesterol, associated with the changes in cecal microbiota composition. The relative abundances of Prevotellaceae, Lactobacillaceae, and Alcaligenaceae families were increased in the WGO group, while abundances of Clostridiaceae and Lachnospiraceae families were greater in the LBO group [61]. Subsequent analysis investigated the cholesterol-lowering effect of a whole-grain oat-based breakfast in a cardio-metabolic “at risk” population. It was observed that TC and LDL-C were significantly decreased after WGO consumption compared to the control group. Furthermore, a significant change was also observed in the relative abundances of fecal bifidobacteria, lactobacilli, and total bacterial count, which were all increased after WGO supplementation [62]. In another similar study, the effect of oat- and tartary-buckwheat-based food (OF) on cholesterol reduction and the gut microbiota in hypercholesterolemic hamsters was evaluated. After intake of OF for 30 days, the plasma TC and LDL-C, liver TC, cholesterol ester (CE), and TG were significantly decreased, whereas the concentrations of acetate, propionate, butyrate, and total SCFAs were significantly increased, in the hamsters fed with OF. Moreover, 16s rRNA sequencing results showed that the relative abundances of Erysipelotrichaceae, Ruminococcaceae, and Lachnospiraceae were decreased and the relative abundance of Eubacteriaceae was increased, beneficially influencing the fatty acid metabolism [60]. These results indicated that the increased SCFAs and the improved gut microbiota served their hypocholesterolemic function.
3.2. Hypertension
Hypertension is a major risk factor for cardiovascular disease [63]. Dietary approaches to stop hypertension (DASH) involve diets rich in fruits, vegetables, whole grains, and low-fat dairy products, with a reduced content of saturated and total fat, and are recommended for adults with elevated hypertension or blood pressure [64]. In a randomized controlled trial, the participants who consumed oat bran (30 g/d) had a lower office systolic blood pressure and office diastolic blood pressure, and the use of antihypertensive drugs in the DF group was reduced. Moreover, the abundances of Bifidobacterium and Spirillum were significantly elevated [65]. Similarly, the BP-lowering effect of buckwheat iminosugar d-fagomine was measured in sucrose-induced hypertensive rats. D-fagomine reduced sucrose-induced hypertension, urine uric acid, and steatosis, without impairing glucose tolerance and without affecting perigonadal fat deposition. The intestinal microbiota was also influenced under a high-sucrose (HS) diet with D-fagomine (FG): the Bacteroidetes/Firmicutes ratio was slightly increased compared to the HS diet group, which was consistent with the changes among the standard, HS, and HS+FG groups. Furthermore, the increased numbers of excreted Enterobacteriales in the HS+FG group proved the inhibitory activity of d-fagomine on epithelial adhesion of Escherichia coli [66]. Recently, Guo et al. studied the antihypertensive effect of quinoa protein on spontaneously hypertensive rats (SHRs). During the five weeks of intake of quinoa protein, the blood pressure (BP) was significantly decreased, while the alpha diversity was increased, and the microbial structure was changed. Subsequent analysis revealed that the abundances of Turicibacter and Allobaculum genera were negatively correlated with BP [67].
3.3. Nonalcoholic Fatty Liver Disease
In general, patients with dyslipidemia also suffer from nonalcoholic fatty liver disease (NAFLD), and clinical manifestations include excessive accumulation of lipids, including TG and FAs, in hepatocytes [68]. Previous studies suggested that the gut microbiome is involved in the development of dyslipidemia and NAFLD [69], whereas the intestine and liver are intrinsically linked and strongly interdependent in metabolic functions [70,71]. For example, common buckwheat consumption significantly improved physiological indexes and biochemical parameters related to dyslipidemia and NAFLD in mice fed with an HFD. In addition, the fecal bile acid (BA) abundance was increased and fecal SCFA reductions were recovered after intake of buckwheat. Moreover, it has been found that buckwheat intervention is more beneficial than simvastatin (10 mg/kg/d) in ameliorating the dysbiosis of intestinal microbial populations induced by an HFD. The abundances of Lactobacillus, Blautia, and Akkermansia were markedly increased, whereas the abundances of Allobaculum, Desulfovibrio, and Bacteroidales_S24-7 were decreased under buckwheat intervention [71]. It has been reported that the possible mechanism of the lipid-lowering effect of Lactobacillus may be to produce lactic acid through intestinal glucose metabolism and promote blood lipid metabolism γ-production of aminobutyric acid and other useful substances [72]. The increase in the genus Blautia might occur mainly because of the syntrophic effect [18]. Akkermansia could consume host-derived mucins such as carbon and nitrogen sources and ferment indigestible carbohydrates, while mainly producing propionic acid. In contrast, Allobaculum showed a strong positive association with lipid metabolic phenotypes [73]. These changes were consistent with previous studies. However, the abundance of Bacteroidales_S24-7, reported to be involved in host–microbe interactions that impact gut function and health [74], was considerably increased by the HFD supplement but reduced by the buckwheat intervention. Recent studies have shown that Monascus fermentation can increase the content of polyphenols or flavonoids in the fermentation substrate [75]. Huang and co-workers found that oral administration of Monascus purpureus-fermented common buckwheat (HQ) significantly inhibited the abnormal growth of body weight and epididymal white adipose tissue (eWAT), prevented the hypertrophy of epididymal adipocytes, and alleviated some biochemical parameters of serum and liver related to lipid metabolism in HFD-induced mice. Histological analysis also exhibited that HQ supplementation reduced the excessive accumulation of lipid droplets in the livers. Furthermore, HQ consumption significantly changed the structure of the intestinal microflora. In the comparison of the HFD and HQ groups, intake of HQ markedly increased the relative levels of Ruminiclostridium, Lacobacillus, and Alistipes and decreased the abundances of the Bacteroidales S24-7 group and Clostridiales XIII in HFD-induced mice. Moreover, UPLC-QTOF/MS-based liver metabolomics showed that HQ consumption exerted remarkable effects on the metabolic pathways of primary bile acid biosynthesis, ether lipid metabolism, glycine, serine and threonine metabolism, glutathione metabolism, pyrimidine metabolism, amino sugar and nucleotide sugar metabolism, etc. These results demonstrated that NAFLD could be attenuated by HQ intervention through regulating the hepatic metabolite profile and gut microbiota. [76].
3.4. Atherosclerosis
Atherosclerosis, the major cause of cardiovascular disease, is a chronic inflammatory disease, which can lead to vascular cognitive impairment and dementia via gradual thickening of large artery walls, reducing blood flow [77,78]. In atherosclerosis, inflammation can damage vascular endothelial cells and promote the formation of foam cells, causing lesions. Increased inflammation and lipid accumulation play key roles in the development of atherosclerosis [79,80]. In the classic atherosclerosis model, the anti-atherosclerotic activity of millet shell polyphenols (MSPs) was measured in ApoE-/-mice fed with a high-fat diet. The results showed that the atherosclerotic plaques in the aorta were effectively inhibited by MSPs, and the LPS and inflammatory cytokine levels, including TNF-α and IL-1β, were also decreased. Conversely, the mRNA expression of tight junction proteins such as occludin, zona occludens-1, and claudin1 was significantly increased in the MSPs group. Moreover, the structure of gut microbiota in ApoE-/-mice with a high-fat diet was improved under consumption of MSPs, resulting in increased abundances of Oscillospira and Ruminococcus and a decreased abundance of Allobaculum at the genus level [81]. Recently, the LDL receptor knock-out mice (LDLR-/-) model, along with a high LDL-cholesterol concentration, has been the established model for atherosclerotic-related cognitive dysfunction [82]. Gao et al. evaluated the effects of an oat-fiber intervention on cognitive behavior by targeting the neuroinflammation signal and the microbiome–gut–brain axis in a mouse model of atherosclerosis. They found that dietary oat fiber retarded the progression of cognitive impairment in a mouse model of atherosclerosis, due to the effect of its metabolites and SCFAs on the diversity and abundance of the gut microbiota that produced anti-inflammatory metabolites, leading to repressed neuroinflammation and reduced gut permeability through the microbiome–gut–brain axis [83].
Overall, increasing numbers of bioactive components in coarse cereals have been proved to possess a hypolipidemic effect, leading to decreased TC, TG, HDL-C, LDL-C, AST, ALT, and MDA. The key genes of lipid metabolism and inflammatory factors are inhibited, whereas the composition of gut microbiota may have an important role in hyperlipidemic regulation. Among these changed bacteria, the F/B ratio, Akkermansia, Prevotellaceae, and Lactobacillaceae were thought to contribute to regulating serum lipid, while the abundances of Allobaculum, Lachnospiraceae, and Bacteroidales S24-7 were inversely correlated with levels of plasma lipids.
Table 2.
Effect of whole-grain components on gut microbiota of cardiovascular diseases host.
4. Effects on Obesity
Obesity has become a serious public health problem all over the world [88], and is closely associated with several diseases including hypertension, type 2 diabetes, coronary artery disease, chronic inflammation, and cancer [89,90,91]. Previous studies have proved that the occurrence and development of obesity is related to changes in the composition and metabolism of the gut microbiota [92]. In the above section, coarse cereals and their bioactive substances exhibited lipid-lowering efficacy in cardiovascular disease. Furthermore, they also have potential for the management of obesity and several related metabolic disorders. In an experiment by Ji et al., dietary intake of a mixture of coarse cereals could prevent body weight gain and fat accumulation, improve blood glucose tolerance and serum lipids levels, and reduce systemic inflammation in HFD-fed mice. Moreover, the relative abundances of Lactobacillus and Bifidobacterium could be increased after supplementing with a mixture of coarse cereals, which might contribute to the anti-obesity activity [93].
In addition to mixed coarse cereals, many single grains have also been proved to promote weight reduction. For instance, obese db/db mice were fed commercial diets with and without quinoa supplementation for eight weeks. The results showed that the quinoa-supplemented diet delayed the body weight gain of mice. Moreover, 16s sequencing analysis showed that the relative abundances of Enterococcus, Turicibacter, Akkermansia, and Bacteroidetes returned to normal levels as shown in the lean group [94]. Among these groups, Turicibacter was considered to be a group with several beneficial effects, whereas Akkermansia was a mucin-degrading bacterial taxon that was negatively correlated with beneficial body weight effects [95,96,97]. An opposite result was observed in this study: the abundance of Firmicutes was higher in lean mice compared to db/db mice, which was mainly related to their diets [94]. Likewise, the effect of barley supplementation on the fecal microbiota was also measured in obese db/db mice. However, the fecal microbiota results showed that barley consumption contributed to a phylogenetically unique microbiota distinct from both obese and lean controls. Moreover, the content of cecal butyrate was similar in obese mice, whereas succinic acid was lower in the barley group. Compared to the obese group, barley supplementation was also relevant, with lower plasma insulin and resistin levels. [98]. Furthermore, previous studies reported higher and lower levels of cecal butyrate during intake of whole wheat and quinoa, respectively, compared to obese controls [94,99].
Along the same lines, the effect of a whole barley (WB) supplement on preventing obesity was evaluated in germ-free (GF) C57BL/6J mice and human-flora-associated (HFA) mice. The results showed that WB prevented obesity and hypercholesterolemia in the GF and HFA mice. The postulated mechanism was the inhibition of cholesterol synthesis due to the downregulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase and sterol regulatory element-binding protein-1c, thereby decreasing cholesterol accumulation. In addition, intake of WB enriched a variety of bacterial genera that are negatively associated with obesity, including Bacteroides, Parabacteroides, and Clostridium cluster XIVa, suggesting that WB counteracted gut dysbiosis in obese mice [100]. In an experiment by Zhong et al., whole-grain barley and barley malt consumption improved the structure of cecal microbiota in HFD-induced rats. Compared to the control group, the barley group had lower abundances of Firmicutes and Deferribacteres and higher abundances of Verrucomicrobia and Actinobacteria. At the genus level, the abundances of Akkermansia, Ruminococcus, Blautia, and Bilophila were increased, while Turicibacter and Roseburia were more abundant in the malt group. Most genera correlated with acetic and propionic acids. However, Roseburia and Turicibacter correlated with butyric acid, and Clostridium and Akkermansia correlated with succinic acid [101]. It has also been demonstrated that a synbiotic consisting of Lactobacillus plantarum S58 (LP.S58) and hull-less barley β-glucan (β-G) attenuates lipid accumulation in HFD-induced mice. In addition, LP.S58 and β-G alleviated gut microbiological dysbiosis in HFD-fed mice. The abundances of Helicobacter, Ruminococcaceae, and Bacteroides were markedly reduced after synergistic treatment with LP.S58 and β-G, while the abundances of Lactobacillus, Allobaculum, Turicibacter, Dubosiella, Akkermansia, and Faecalibaculum were increased [102].
Likewise, Hereu et al. investigated the effect of combined d-fagomine and ω-3 polyunsaturated fatty acids (ω-3 PUFAs) on weight gain and beneficial gut bacterial strains. D-fagomine was first isolated from the seeds of buckwheat and could reduce weight gain in the rats fed with ω-3 PUFAs, compared to the controls. Moreover, intake of ω-3 PUFAs promoted the diversity of the gut microbiota by increasing the abundance of Bacteroidetes in healthy rats and mitigated the age-related reduction of Lactobacillus and Bifidobacterium, which are presumed to be beneficial [103,104]. Eicosapentaenoic acid (EPA, 20:5, n-3) and docosahexaenoic acid (DHA, 22:6, n-3) are the major ω-3 PUFAs, and they are considered to be crucial components in preventing cardiovascular diseases [105]. Previous studies reported that EPA and DHA significantly increased the abundances of Firmicutes (Lactobacillus genus) and Bifidobacteria in mice fed an HFD [106,107]. According to the authors, the d-fagomine and ω-3 PUFAs combination group gained less weight compared to the controls and the ω-3 PUFAs group, and the combination helped maintain the relative populations of Bacteroides and Prevotella in the rat intestinal tract, providing the anti-inflammatory and cardiovascular benefits of ω-3 PUFAs [108]. It is well known that tartary buckwheat is rich in flavonoids, and dietary rutin or quercetin in buckwheat can ameliorate lipid metabolism, whereas buckwheat-resistant starch may also have beneficial effects on lipid metabolism. For example, Peng et al. proved that quercetin could significantly reduce body weight, serum TC, LDL-C, insulin, and TNF-α, while buckwheat significantly increased the weight of the rats. Moreover, rutin, quercetin, and buckwheat tended to decrease fat deposition in the liver of rats but had little effect on production of SCFAs. It has also been found that they can shape the specific structure of the gut microbiota [109]. Zhou et al. investigated the regulatory effects of tartary-buckwheat-resistant starch (BRS) on blood lipid levels and the gut microbiota in HFD-induced mice. After treatment, plasma levels of TC, TG, glucose, and cytokines were markedly decreased and the antioxidant capacity in the liver and duodenum was enhanced. Furthermore, the gut microbiota composition was regulated after the intake of BRS, and the populations of Bacteroides, Blautia, Lactobacillus, Bifidobacterium, and Enterococcus were promoted, while the growth of Faecalibaculum, Erysipelatoclostridium, and Escherichia coli was inhibited [110,111].
Like quinoa, millet lacks gluten and is rich in dietary fiber, polyphenols, and minerals, which means that it could be explored as a healthy food. Recently, Sarma and co-workers investigated the protective role of whole-grain kodo millet supplementation in HFD-induced mice. They found that millet consumption ameliorated glucose tolerance and inhibited increases in the serum cholesterol and lipid parameters but did not influence weight gain. Moreover, millet increased the beneficial gut bacterial abundances, including Lactobacillus spp., Bifidobacteria, Akkermansia, and Roseburia spp. [112].
Grain sorghum is the world’s fifth most produced cereal crop, but it is commonly grown only in specific areas. Sorghum contains various types of polyphenols, which are found in the bran [113]. Ashley et al. studied the effect of grain sorghum polyphenols on the gut microbiota of different weight classes. There were no significant changes in total production of SCFAs between normal-weight and overweight/obese groups. In contrast, a combination of sorghum bran polyphenols and fructo-oligosaccharides could increase the populations of Bifidobacterium and Lactobacillus, and sorghum bran polyphenols independently enhanced the abundances of Roseburia and Prevotella [114].
The same clinical parameters were observed in obesity as in cardiovascular diseases. There were some differences in the flora changes, such as for Allobaculum and Blautia, which were increased, but further study is needed on role of the gut microbiota in the regulation of obesity.
5. Effects on Diabetes
Diabetes and its related diseases have reached alarming levels worldwide [115]. Type 2 diabetes (T2D) accounts for the vast majority of cases of diabetes. Unhealthy dietary habits, including excess consumption of sugar, oil, and flour, leads to obesity, which is the main cause of T2D [116]. Growing evidence suggests that T2D can be prevented and treated through intake of whole grains [117]. However, there are few studies about the hypoglycemic effect of grains via regulation of the gut microbiota. Recently, Ren et al. evaluated the hypoglycemic effect of foxtail millet (FM) in high-fat-diet and streptozotocin-induced diabetic rats, and the results exhibited remarkably decreased fasting glucose (FG), glycated serum protein, and areas under the glucose tolerance test. A 16s rRNA sequencing analysis showed that FM consumption significantly increased the relative abundances of Lactobacillus and Ruminococcus_2, which were significantly negatively correlated with FG and 2 h glucose. Subsequently, a Spearman’s correlation analysis indicated a complicated set of interdependences among the gut microbiota and metabolic parameters [118].
Regarding the protein of FM, the hypoglycemic impact of different protein types was investigated by Fu and co-workers. Protein isolates from raw and cooked FM improved glucose intolerance and insulin resistance in diabetic mice. However, only the protein of cooked FM recovered the weight loss trend and alleviated lipid disorders in diabetic mice. The results of the gut flora showed that intake of FM protein isolates simulated the population of Akkermansia and decreased the abundance of Lachnospiraceae [119]. Akkermansia was reported to be negatively associated with obesity, insulin resistance, diabetes, and other metabolic diseases [120]. In this study, the increased population of Akkermansia may be attributed to the fact that the amino acids produced by the hydrolysis of millet protein isolate in the intestine can be metabolized by the intestinal flora, finally producing amines and polyamines through a decarboxylation reaction, which was demonstrated to improve the Akkermansia abundance [121,122]. Moreover, the protein isolated from the raw foxtail millet increased the abundances of the Clostridiaceae_1 family and Faecalibaculum genus, whereas protein isolate from the cooked foxtail millet increased the level of the genus Ruminococcaceae_UCG-014. A previous study proved that the populations of the Clostridiaceae_1 family and Ruminococcaceae_UCG-014 genus were negatively associated with the circulating isoleucine level in plasma [123]. Isoleucine belongs to the branched-chain amino acids (BCAAs) and has been shown to be positively correlated with the development of obesity and insulin resistance [124]. Furthermore, Faecalibaculum was found to produce lactic acid, and its metabolites were reported to have anti-diabetic effects [125]. More recently, these authors investigated the hypoglycemic effect of prolamin from cooked foxtail millet (PCFM) on T2DM mice. PCFM ameliorated glucose metabolism disorders associated with type 2 diabetes, and diabetes-related and gut microbiota dysbiosis in mice. PCFM inhibited the growth of Lachnospiraceae_UCG-006, Anaerotruncus, and Butyricimonas, as well as promoting the populations of Odoribacter, Blautia, and Akkermansia [126]. Lachnospiraceae_UCG-006 was revealed to be positively correlated with TG, TC, HDL-C, and LDL-C [127]. Moreover, Anaerotruncus genus was reported to be positively related to glucose intolerance in HFD mice, whereas a positive correlation between the abundance of Butyricimonas and fasting blood glucose in T2DM patients has been observed [128,129]. In contrast, Odoribacter was negatively related to steady-state plasma glucose in pre-diabetic individuals [130]. Moreover, the gut microbiota composition was markedly altered and glucose metabolism disorders in STZ-induced diabetic rats were also attenuated after intake of Akkermansia muciniphila [131]. Furthermore, enriched Blautia may contribute to the amelioration of inflammation and insulin resistance in diabetes via the production of short-chain fatty acids [132].
In a previous study, D-fagomine was proved to reverse sucrose-induced steatosis and hypertension, presumably by reducing the postprandial levels of fructose in the liver [66]. Lately, Ramos-Romero et al. measured the effect of buckwheat iminosugar D-fagomine on diet-induced pre-diabetes in rats. They found that D-fagomine could reduce fat-induced impaired glucose tolerance, inflammation markers, and mediators. In addition, the changes in the main two bacterial phyla, Bacteroidetes and Firmicutes, in the intestinal tract were measured, and D-fagomine had no impact on modifying the reduction in the Bacteroidetes/Firmicutes ratio that led to body weight gain. Moreover, compared to the effect of D-fagomine on fat-induced changes in Enterobacteriales and Bifidobacteriales, it had a significant effect on glucose tolerance and inflammation [133].
In brief, obesity is recognized as the main cause of diabetes; hence, the pathophysiological parameters and changes in gut microbiota, such as the weight loss trend, insulin resistance, glucose intolerance, and inflammation markers would be expected to be similar. The composition of gut flora also exhibited similar alterations with an increasing abundance of Blautia and Akkermansia and a reduction in the numbers of Lachnospiraceae and Faecalibaculum. However, the relationships among gut microbiota, obesity, and diabetes are still unclear.
6. Prebiotic Effect
The concept of prebiotics was proposed by Gibson and Roberfroid in 1995. They first defined a prebiotic as a “non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon, and thus improves host health” [134]. The functional characteristics of prebiotics include resistance to low pH in the stomach, resistance to hydrolysis by mammalian enzymes, resistance to absorption in the upper gastrointestinal tract, ability to be fermented by intestinal microflora, and selective stimulation of the growth and/or activities of intestinal bacteria related to host health and overall well-being [13,134]. Generally, non-digestible carbohydrates were considered to be the main prebiotics, but other compounds, including phenolics, polyunsaturated fatty acid, linoleic acid, and phytochemicals, were included in the updated prebiotic definition [135]. As our knowledge and understanding of the gut microbiota and the importance of the gut microbiota increases, consumers must understand the key differences between the different forms of prebiotics and their presence in a variety of foods and related products.
Whole grains are rich in dietary fiber, micronutrients, and phytochemicals, which are considered to be natural prebiotics. For instance, the effect of oat bran on the human fecal microbiota composition was measured via in vitro anaerobic batch-culture experiments. The 16s rRNA sequencing results showed that the populations of Proteobacteria and Bacteroidetes were increased after oat supplementation, whereas fermentation of the 1% (w/v) oat bran increased the abundances of Bifidobacterium unassigned at 10 h and Bifidobacterium adolescentis at 10 and 24 h, compared to the control group. The SCFA analysis showed that consumption of oats stimulated the production of acetic and propionic acid, while oat bran fermentation resulted in an elevated SCFAs content. Meanwhile, β-glucan treatment induced an increase in the phylum Bacteroidetes, and the polyphenol mix induced an increase in the Enterobacteriaceae family at 24 h. It was concluded that the prebiotic effect of oats could be attributed to the synergy of all oat compounds within the complex food matrix [136]. Likewise, the prebiotic effect of quinoa and amaranth were evaluated in batch cultures with fecal human inocula. In this study, fluorescence in situ hybridization (FISH) was used to explore the microbiota composition and metabolic products. After treatment, the changes in the SCFAs of the quinoa group and the amaranth group were consistent with the decrease in pH. Moreover, there were significant alterations in the gut flora, including Bifidobacterium spp., Lactobacillus–Enterococcus, Atopobium, Bacteroides–Prevotella, Clostridium coccoides–Eubacterium rectale, Faecalibacterium prausnitzii, and Roseburia intestinalis. Due to the growth of these bacterial groups, the SCFAs, acetate, propionate, and butyrate were markedly increased, which is critical for maintaining a balanced intestinal microbiota [137].
The Maillard reaction (MR) produces several low-mass molecules by reducing sugar and amino acids during food heat treatment and storage; these low-weight compounds are usually recombined by a series of advanced MRs to form melanoidins [138]. Because they have a similar structure to fibers, melanoidins were considered to have prebiotic properties [139]. Aljahdali and co-workers examined the effect of intake of melanoidins of barley malts on gut microbiota in mice. It was found that the melanoidins significantly changed the gut microbiota and stimulated the production of SCFAs. The populations of Lactobacillus, Parasutterella, Akkermansia, Bifidobacterium, and Barnesiella were remarkably increased, while Dorea, Oscillibacter, and Alisitpes were decreased, proving the prebiotic effect of melanoidins [140].
7. Effect on Other Chronic Diseases
7.1. Metabolic Syndrome
Metabolic syndrome is a complex disease with a rapid increase in incidence in recent years, and dietary intervention is an effective approach to ameliorate the metabolic status of patients [141]. Dietary composition can actively change the composition and function of intestinal microflora, so as to improve individual conditions related to metabolic disorders [142]. Barley β-glucans have been demonstrated to exert a hypoglycemic effect and a cholesterol-lowering effect. In an experiment by Velikonja et al., the impact of a dietary supplement of barley β-glucans on metabolic syndrome patients was investigated. After treatment, TC in plasma was decreased compared to the control group. Moreover, the fecal SCFAs were markedly changed, with increased propionic acid, while the acetic acid was decreased in the control group. Furthermore, 16s sequencing results showed that the populations of health-associated Bifidobacterium spp. and Akkermansia municiphila were increased after barley β-glucans consumption [143].
Growing evidence suggests that circadian disruption may increase the risk of metabolic diseases, causing a series of metabolic abnormalities including obesity, dyslipidemia, and glucose intolerance that are regarded as circadian-disruption-induced metabolic syndrome (CDIMS). Moreover, circadian disruption can increase the gut permeability, leading to dysfunction of the gut barrier and alteration of the gut microbiota [144]. In an induced CDIMS mice model, intake of oat β-glucans mitigated CDIMS, resulting in an increase in body weight, liver-weight-to-body-weight ratio, and plasma leptin concentration, and restored glucose tolerance. The analysis of gut microbiota showed that oat β-glucans increased the species richness and improved the abundance of seven bacterial genera and increased butyrate producers such as Ruminococcaceae and Lachnospiraceae, which contribute to glucose homeostasis and gut barrier protection [145]. On the other hand, antibiotic overuse can also destroy the gut microbiota by changing the community structure and reducing the diversity, resulting in gut barrier dysfunction [146,147]. Zhu and co-workers assessed the effect of a water-soluble polysaccharide from Fagopyrum esculentum Moench bee pollen (WFPP) on gut barrier integrity in antibiotic-treated mice. WFPP reversed the growth retardation, atrophy of the thymus and spleen, increased gut permeability, and intestinal barrier damage under ceftriaxone (a broad-spectrum antibiotic) treatment. Furthermore, WFPP was correlated with reduced inflammatory response and enhanced intestinal sIgA secretion. In addition, 16S rDNA sequencing results showed that WFPP ameliorated gut microbiota dysbiosis induced by ceftriaxone, and especially that it regulated the populations of Proteobacteria (sIgA-secretion-related bacteria) and Enterococcus (inflammation-related bacteria) [148].
7.2. Neurodegenerative Disorders
Neurodegenerative disorders such as Alzheimer’s disease (AD) and dementias are major causes of incidence rates, reduced quality of life and medical costs in elderly people [149]. There is evidence that microbial changes are related to neuroinflammation and cognitive impairment, which are two critical features of AD pathogenesis and progression [150]. Although neurodegenerative diseases are currently incurable, one third of dementia cases can be prevented by addressing lifestyle factors, including diet [149]. As an example, β-glucan prevents cognitive impairment caused by a high-fat and fiber-deficient diet (HFFD), as shown by behavioral appraisement through object positioning, new object recognition and nesting tests. The results showed that β-glucan reversed the cognitive function loss through more optimized synaptic and signaling transduction pathways in crucial brain areas, while intake of β-glucan also improved gut barrier dysfunction and reduced bacterial endotoxin translocation. Moreover, β-glucan alleviated gut microbiota dysbiosis caused by HFFD, especially Bacteroidetes at phylum level and its lower taxa [151]. A previous study demonstrated that the phylum Bacteroidetes was intimately related to neurodegenerative diseases. There was a lower abundance of Bacteroides at genus level in the gut microbiota of dementia patients, whereas the population of Bacteroides fragilis at species level was lower in patients with cognitive impairment [152,153]. According to the authors, oat β-glucan increased satiety in diet-induced obese mice by activating the gut–hypothalamic (PYY-NPY) axis [154]. SCFAs that were produced from β-glucan fermentation efficiently increased PYY production in human entero-endocrine cells [155]. In addition, excessive dietary energy intake and insulin resistance have adverse effects on human cognition, but energy restriction could enhance neural plasticity by reducing inflammation and increasing the expression of synaptic-plasticity-related proteins [156]. Therefore, β-glucan consumption is important for preventing cognitive impairment induced by HFFD by maintaining the gut microbiota–brain axis.
7.3. Malnutrition
Malnutrition is usually caused by insufficient protein and micronutrients due to lack of food [157]. Due to the poorly developed immune system and delayed puberty, children are the group most vulnerable to malnutrition. Recently, Li and co-workers evaluated the effect of a millet supplement on malnutrition and gut microbiota dysbiosis in a malnourished pig model. Compared to the control group, the millet supplement improved the growth status, gastrointestinal pathology, and content of total protein and globulin in the blood. Moreover, intake of millet increased the abundance of Bacteroidetes, Firmicutes, and Lachnospira spp., while the population of Proteobacteria was decreased. Furthermore, a Pearson’s correlation analysis showed a strong positive correlation between the abundances of Faecalibacterium and Lachnospira spp. and body weight, crown–rump length and total serum protein [158]. Previously, a low abundance of Faecalibacterium prausnitzii was found in the intestines of malnourished children and malnourished pigs in Bangladesh [159]. Moreover, the abundance of Lachnospira spp. was positively associated with an anti-inflammatory effect in weaner pigs fed with an HFD [160]. Therefore, millet exhibited a promising benefit in regulating gut microbiota dysbiosis in malnourished pigs.
8. Prospects
In this study, we reviewed the functional components of coarse grains in alleviating chronic diseases through modulating the gut microbiota. A correct understanding of the critical role of coarse grains in gut health and microbial regulation could help ameliorate chronic diseases and improve quality of life. The complexity of mechanisms among the components of coarse grains, the digestive system and the gut microbiota is an important factor restricting development and utilization of coarse grains. Despite the fact that many bioactive substances in coarse grains have been proved to serve various functions in both in vitro and in vivo experiments, there is still only limited research on accurate metabolic processes of bioactive substances and their functional signal pathways. Moreover, the intensity and the targeting effects of these substances in the treatment of chronic diseases are far less than those of drugs. At present, there are a number of mature plant-derived drug evaluation methods, including in vitro high-throughput screening, animal model experiments, and human trials. However, single-substance experiments are not suitable for assessing the nutritional characteristics of coarse grains.
Generally, coarse grains are considered to be food rather than drugs. Therefore, coarse grains play a preventive and regulatory role via the synergistic action of various substances. Moreover, gut dysbiosis usually occurs in the early stage of disease. In order to ensure that the active substances of coarse cereals play a more effective role in improving chronic diseases, several aspects should be further studied in detail, including: (1) improving the bioactivity of these substances, which is effective and important for further research and application; (2) exploring more precise and persuasive experiments to clarify the interaction mechanisms between gut microbiota and bioactive substances; (3) combining the in vitro bioreactor research model and the in vivo gut reaction model to establish rapid screening and comprehensive evaluation of the bioactivity of coarse cereals. The outcome of this research would be helpful to further clarify the beneficial effects of cereal bioactive components on chronic diseases.
Author Contributions
Conceptualization, G.R., X.F., and C.T.; writing—original draft preparation, X.F., G.R., and Y.L.; writing—review and editing, C.T., C.B., N.E., and G.R.; funding acquisition, G.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (Grant No. CAAS-ASTIP-2017-ICS).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fu, J.; Zhang, Y.; Hu, Y.C.; Zhao, G.; Tang, Y.; Zou, L. Concise review: Coarse cereals exert multiple beneficial effects on human health. Food Chem. 2020, 325, 126761. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.B.; Wang, Y.Y.; Shabani, K.I.; Qin, X.L.; Liu, X. Comparison of structural features of reconstituted doughs affected by starches from different cereals and other botanical sources. J. Cereal Sci. 2020, 93, 102937. [Google Scholar] [CrossRef]
- Lie, L.; Brown, L.; Forrester, T.E.; Plange-Rhule, J.; Bovet, P.; Lambert, E.V.; Layden, B.T.; Luke, A.; Dugas, L.R. The Association of Dietary Fiber Intake with Cardiometabolic Risk in Four Countries across the Epidemiologic Transition. Nutrients 2018, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Idehen, E.; Tang, Y.; Sang, S.M. Bioactive phytochemicals in barley. J. Food Drug Anal. 2017, 25, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Llaverias, G.; Escola-Gil, J.C.; Lerma, E.; Julve, J.; Pons, C.; Cabre, A.; Cofan, M.; Ros, E.; Sanchez-Quesada, J.L.; Blanco-Vaca, F. Phytosterols inhibit the tumor growth and lipoprotein oxidizability induced by a high-fat diet in mice with inherited breast cancer. J. Nutr. Biochem. 2013, 24, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M.; de Vos, E.A.J. Role of the intestinal microbiome in health and disease: From correlation to causation. Nutr. Rev. 2012, 70, S45–S56. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Devkota, S.; Wang, Y.W.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10(-/-) mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henao-Mejia, J.; Elinav, E.; Jin, C.C.; Hao, L.M.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Fogliano, V.; Leonard, C.; Gibson, G.R.; Tuohy, K.M. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2008, 99, 110–120. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Xing, X.; Zhen, H. Enzyme deactivation treatments did not decrease the beneficial role of oat food in intestinal microbiota and short-chain fatty acids: An in vivo study. J. Sci. Food Agric. 2013, 93, 504–508. [Google Scholar] [CrossRef]
- Berger, K.; Falck, P.; Linninge, C.; Nilsson, U.; Axling, U.; Grey, C.; Stalbrand, H.; Karlsson, E.N.; Nyman, M.; Holm, C.; et al. Cereal Byproducts Have Prebiotic Potential in Mice Fed a High-Fat Diet. J. Agric. Food Chem. 2014, 62, 8169–8178. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharm. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Pere-Trepat, E.; Montoliu, I.; Martin, F.P.J.; Collino, S.; Moco, S.; Godin, J.P.; Cleroux, M.; Guy, P.A.; Breton, I.; et al. A Whole-Grain-Rich Diet Reduces Urinary Excretion of Markers of Protein Catabolism and Gut Microbiota Metabolism in Healthy Men after One Week. J. Nutr. 2013, 143, 766–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.Y.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2013, 7, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Tome, S.; Hernandez-Ledesma, B.; Chaparro, M.; Indiano-Romacho, P.; Bernardo, D.; Gisbert, J.P. Role of food proteins and bioactive peptides in inflammatory bowel disease. Trends Food Sci. Technol. 2019, 88, 194–206. [Google Scholar] [CrossRef]
- Gassull, M.A. Review article: The intestinal lumen as a therapeutic target in inflammatory bowel disease. Aliment. Pharm. Ther. 2006, 24, 90–95. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Larussa, T.; Imeneo, M.; Luzza, F. Potential role of nutraceutical compounds in inflammatory bowel disease. World J. Gastroenterol. 2017, 23, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Albenberg, L.; Compher, C.; Baldassano, R.; Piccoli, D.; Lewis, J.D.; Wu, G.D. Diet in the Pathogenesis and Treatment of Inflammatory Bowel Diseases. Gastroenterology 2015, 148, 1087–1106. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.Y.; Zhao, J.J.; AL-Ansi, W.; Wang, J.; Xue, L.M.; Liu, J.X.; Wang, Y.; Fan, M.C.; Qian, H.F.; Li, Y.; et al. Oat beta-glucan alleviates DSS-induced colitis via regulating gut microbiota metabolism in mice. Food Funct. 2021, 12, 8976–8993. [Google Scholar] [CrossRef] [PubMed]
- Wilczak, J.; Blaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jalosinska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The effect of low or high molecular weight oat beta-glucans on the inflammatory and oxidative stress status in the colon of rats with LPS-induced enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef]
- Menon, R.; Gonzalez, T.; Ferruzzi, M.; Jackson, E.; Winderl, D.; Watson, J. Oats-From Farm to Fork. Adv. Food Nutr. Res. 2016, 77, 1–55. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.B. Effect of oat hulls as a free choice feeding on broiler performance, short chain fatty acids and microflora under a mild necrotic enteritis challenge. Anim. Nutr. 2018, 4, 65–72. [Google Scholar] [CrossRef]
- Tian, M.L.; Li, D.T.; Ma, C.; Feng, Y.; Hu, X.S.; Chen, F. Barley Leaf Insoluble Dietary Fiber Alleviated Dextran Sulfate Sodium-Induced Mice Colitis by Modulating Gut Microbiota. Nutrients 2021, 13, 846. [Google Scholar] [CrossRef] [PubMed]
- Li, D.T.; Feng, Y.; Tian, M.L.; Ji, J.F.; Hu, X.S.; Chen, F. Gut microbiota-derived inosine from dietary barley leaf supplementation attenuates colitis through PPAR gamma signaling activation. Microbiome 2021, 9, 1–22. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [Green Version]
- Rose, D.J. Impact of whole grains on the gut microbiota: The next frontier for oats? Br. J. Nutr. 2014, 112, S44–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhang, Y.; Qiu, B.; Fan, S.J.; Ding, H.F.; Liu, Z.H. Quinoa whole grain diet compromises the changes of gut microbiota and colonic colitis induced by dextran Sulfate sodium in C57BL/6 mice. Sci. Rep. 2018, 8, 14916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, W.; Wang, X.; Ding, C.; Liu, J.Z.; Li, Y.; Li, W.J.; Sun, Y.D. High-Salt Diet-Induced Gastritis in C57BL/6 Mice is Associated with Microbial Dysbiosis and Alleviated by a Buckwheat Diet. Mol. Nutr. Food Res. 2020, 64, e1900965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, W.; Zhang, D.; Yang, Y.; Wang, X.; Li, L. Fermented and Germinated Processing Improved the Protective Effects of Foxtail Millet Whole Grain Against Dextran Sulfate Sodium-Induced Acute Ulcerative Colitis and Gut Microbiota Dysbiosis in C57BL/6 Mice. Front. Nutr. 2021, 8, 694936. [Google Scholar] [CrossRef]
- Nadeem, M.S.; Kumar, V.; Al-Abbasi, F.A.; Kamal, M.A.; Anwar, F. Risk of colorectal cancer in inflammatory bowel diseases. Semin. Cancer Biol. 2020, 64, 51–60. [Google Scholar] [CrossRef]
- Zhang, B.W.; Xu, Y.C.; Liu, S.; Lv, H.; Hu, Y.Z.; Wang, Y.Y.; Li, Z.; Wang, J.; Ji, X.M.; Ma, H.; et al. Dietary Supplementation of Foxtail Millet Ameliorates Colitis-Associated Colorectal Cancer in Mice via Activation of Gut Receptors and Suppression of the STAT3 Pathway. Nutrients 2020, 12, 2367. [Google Scholar] [CrossRef]
- Yang, J.; McDowell, A.; Kim, E.K.; Seo, H.; Lee, W.H.; Moon, C.M.; Kym, S.M.; Lee, D.H.; Park, Y.S.; Jee, Y.K.; et al. Development of a colorectal cancer diagnostic model and dietary risk assessment through gut microbiome analysis. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.P.; Shan, S.H.; Zhang, C.; Shi, J.Y.; Li, H.Q.; Li, Z.Y. Inhibitory Effects of Bound Polyphenol from Foxtail Millet Bran on Colitis-Associated Carcinogenesis by the Restoration of Gut Microbiota in a Mice Model. J. Agric. Food Chem. 2020, 68, 3506–3517. [Google Scholar] [CrossRef]
- He, B.B.; Bai, Y.; Jiang, L.L.; Wang, W.; Li, T.T.; Liu, P.; Tao, S.Y.; Zhao, J.C.; Han, D.D.; Wang, J.J. Effects of Oat Bran on Nutrient Digestibility, Intestinal Microbiota, and Inflammatory Responses in the Hindgut of Growing Pigs. Int. J. Mol. Sci. 2018, 19, 2407. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Miyoshi, T.; Yunoki, K.; Ito, H. Postprandial hyperlipidemia as a potential residual risk factor. J. Cardiol. 2016, 67, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.; Shi, Z.; Yang, X.L.; Cai, Z.W.; Wang, L.X.; Wu, X.M.; Ye, A.Q.; Jiang, J.P. Hypolipidemic effect of pure total flavonoids from peel of Citrus (PTFC) on hamsters of hyperlipidemia and its potential mechanism. Exp. Gerontol. 2020, 130, 110786. [Google Scholar] [CrossRef] [PubMed]
- Magni, P.; Macchi, C.; Morlotti, B.; Sirtori, C.R.; Ruscica, M. Risk identification and possible countermeasures for muscle adverse effects during statin therapy. Eur. J. Intern. Med. 2015, 26, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.P.; Tang, K.R.; Deng, Y.J.; Chen, R.S.; Liang, S.; Xie, H.J.; He, Y.F.; Chen, Y.N.; Yang, Q.H. Effects of shenling baizhu powder herbal formula on intestinal microbiota in high-fat diet-induced NAFLD rats. Biomed. Pharm. 2018, 102, 1025–1036. [Google Scholar] [CrossRef]
- Han, F.; Wang, Y.; Han, Y.Y.; Zhao, J.X.; Han, F.L.; Song, G.; Jiang, P.; Miao, H.J. Effects of Whole-Grain Rice and Wheat on Composition of Gut Microbiota and Short-Chain Fatty Acids in Rats. J. Agric. Food Chem. 2018, 66, 6326–6335. [Google Scholar] [CrossRef]
- Marshall, S.; Petocz, P.; Duve, E.; Abbott, K.; Cassettari, T.; Blumfield, M.; Fayet-Moore, F. The Effect of Replacing Refined Grains with Whole Grains on Cardiovascular Risk Factors: A Systematic Review and Meta-Analysis of Randomized Controlled Trials with GRADE Clinical Recommendation. J. Acad. Nutr. Diet 2020, 120, 1859–1883.e31. [Google Scholar] [CrossRef]
- Duan, R.Q.; Guan, X.; Huang, K.; Zhang, Y.; Li, S.; Xia, J.A.; Shen, M. Flavonoids from Whole-Grain Oat Alleviated High-Fat Diet-Induced Hyperlipidemia via Regulating Bile Acid Metabolism and Gut Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 7629–7640. [Google Scholar] [CrossRef]
- Huang, K.; Yu, W.W.; Li, S.; Guan, X.; Liu, J.; Song, H.D.; Liu, D.D.; Duan, R.Q. Effect of embryo-remaining oat rice on the lipid profile and intestinal microbiota in high-fat diet fed rats. Food Res. Int. 2020, 129, 108816. [Google Scholar] [CrossRef]
- Zhang, L.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Smith, P.T.; Ananthan, R.; Longvah, T. Antioxidant and immunomodulatory activities of polysaccharides from the roots of Sanguisorba officinalis. Int. J. Biol. Macromol. 2012, 51, 1057–1062. [Google Scholar] [CrossRef]
- Cao, Y.N.; Zou, L.; Li, W.; Song, Y.; Zhao, G.; Hu, Y.C. Dietary quinoa (Chenopodium quinoa Willd.) polysaccharides ameliorate high-fat diet-induced hyperlipidemia and modulate gut microbiota. Int. J. Biol. Macromol. 2020, 163, 55–65. [Google Scholar] [CrossRef]
- Fotschki, B.; Juskiewicz, J.; Jurgonski, A.; Amarowicz, R.; Opyd, P.; Bez, J.; Muranyi, I.; Petersen, I.L.; Llopis, M.L. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients 2020, 12, 2781. [Google Scholar] [CrossRef]
- Riviere, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Marinelli, L.; Martin-Gallausiaux, C.; Bourhis, J.M.; Beguet-Crespel, F.; Blottiere, H.M.; Lapaque, N. Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells. Sci. Rep.-UK 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.L.; Yan, B.B.; Xiao, Y.; Zhou, Y.M.; Liu, T.Y. Tartary buckwheat protein prevented dyslipidemia in high-fat diet-fed mice associated with gut microbiota changes. Food Chem. Toxicol. 2018, 119, 296–301. [Google Scholar] [CrossRef]
- Rivera-Espinoza, Y.; Gallardo-Navarro, Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, Q.; Zhao, S.; Yan, B.; Zhou, X. Impact of Buckwheat Fermented Milk Combined with High-Fat Diet on Rats’ Gut Microbiota and Short-Chain Fatty Acids. J. Food Sci. 2019, 84, 3833–3842. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Itoh, K. Intestinal colonization by a Lachnospiraceae bacterium contributes to the development of diabetes in obese mice. Microbes Environ. 2014, 29, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, S.A.; Kamil, A.; Fleige, L.; Gahan, C.G.M. The Cholesterol-Lowering Effect of Oats and Oat Beta Glucan: Modes of Action and Potential Role of Bile Acids and the Microbiome. Front. Nutr. 2019, 6, 171. [Google Scholar] [CrossRef]
- Sun, N.X.; Tong, L.T.; Liang, T.T.; Wang, L.L.; Liu, L.Y.; Zhou, X.R.; Zhou, S.M. Effect of Oat and Tartary Buckwheat-Based Food on Cholesterol—Lowering and Gut Microbiota in Hypercholesterolemic Hamsters. J. Oleo Sci. 2019, 68, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.L.; Hergert, N.; Rompato, G.; Lefevre, M. Whole grain oats improve insulin sensitivity and plasma cholesterol profile and modify gut microbiota composition in C57BL/6J mice. J. Nutr. 2015, 145, 222–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, M.L.; Tzounis, X.; Tuohy, K.M.; Lovegrove, J.A. Hypocholesterolemic and Prebiotic Effects of a Whole-Grain Oat-Based Granola Breakfast Cereal in a Cardio-Metabolic “At Risk” Population. Front. Microbiol. 2016, 7, 1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Chen, C.A.; Tsai, J.S.; Chen, G.W. Preparation and Identification of Novel Antihypertensive Peptides from the In Vitro Gastrointestinal Digestion of Marine Cobia Skin Hydrolysates. Nutrients 2019, 11, 1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Himmelfarb, D.C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, 1269–1324. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The effect of dietary fiber (oat bran) supplement on blood pressure in patients with essential hypertension: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2458–2470. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Amezqueta, S.; Casas, J.; Munoz, S.; Medina, I.; Miralles-Perez, B.; Romeu, M.; Torres, J.L. The Buckwheat Iminosugar d-Fagomine Attenuates Sucrose-Induced Steatosis and Hypertension in Rats. Mol. Nutr. Food Res. 2020, 64, e1900564. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Hao, Y.; Fan, X.; Richel, A.; Everaert, N.; Yang, X.; Ren, G. Administration with Quinoa Protein Reduces the Blood Pressure in Spontaneously Hypertensive Rats and Modifies the Fecal Microbiota. Nutrients 2021, 13, 2446. [Google Scholar] [CrossRef]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Sakanoi, Y.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K.; Tsuduki, T.; et al. Simultaneous Intake of Euglena Gracilis and Vegetables Synergistically Exerts an Anti-Inflammatory Effect and Attenuates Visceral Fat Accumulation by Affecting Gut Microbiota in Mice. Nutrients 2018, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Guo, W.L.; Li, Q.Y.; Xu, J.X.; Cao, Y.J.; Liu, B.; Yu, X.D.; Rao, P.F.; Ni, L.; Lv, X.C. The protective mechanism of Lactobacillus plantarum FZU3013 against non-alcoholic fatty liver associated with hyperlipidemia in mice fed a high-fat diet. Food Funct. 2020, 11, 3316–3331. [Google Scholar] [CrossRef]
- Huang, Z.R.; Deng, J.C.; Li, Q.Y.; Cao, Y.J.; Lin, Y.C.; Bai, W.D.; Liu, B.; Rao, P.F.; Ni, L.; Lv, X.C. Protective Mechanism of Common Buckwheat (Fagopyrum esculentum Moench.) against Nonalcoholic Fatty Liver Disease Associated with Dyslipidemia in Mice Fed a High-Fat and High-Cholesterol Diet. J. Agric. Food Chem. 2020, 68, 6530–6543. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Jeong, M.H.; Ahn, C.B.; Je, J.Y. Antioxidant activity and gamma-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Z.; Guo, H.; He, D.; Zhao, H.; Wang, Z.; Zhang, W.; Liao, L.; Zhang, C.; Ni, L. The modulatory effect of infusions of green tea, oolong tea, and black tea on gut microbiota in high-fat-induced obese mice. Food Funct. 2016, 7, 4869–4879. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, K.L.; Wood, D.L.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bei, Q.; Liu, Y.; Wang, L.; Chen, G.; Wu, Z.Q. Improving free, conjugated, and bound phenolic fractions in fermented oats (Avena sativa L.) with Monascus anka and their antioxidant activity. J. Funct. Foods 2017, 32, 185–194. [Google Scholar] [CrossRef]
- Huang, Z.R.; Chen, M.; Guo, W.L.; Li, T.T.; Liu, B.; Bai, W.D.; Ai, L.Z.; Rao, P.F.; Ni, L.; Lv, X.C. Monascus purpureus-fermented common buckwheat protects against dyslipidemia and non-alcoholic fatty liver disease through the regulation of liver metabolome and intestinal microbiome. Food Res. Int. 2020, 136, 109511. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef]
- Kasikara, C.; Doran, A.C.; Cai, B.S.; Tabas, I. The role of non-resolving inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 2713–2723. [Google Scholar] [CrossRef] [Green Version]
- Lutgens, E.; Atzler, D.; Doring, Y.; Duchene, J.; Steffens, S.; Weber, C. Immunotherapy for cardiovascular disease. Eur. Heart J. 2019, 40, 3937–3946. [Google Scholar] [CrossRef]
- Liu, F.M.; Shan, S.H.; Li, H.Q.; Shi, J.Y.; Hao, R.L.; Yang, R.P.; Li, Z.Y. Millet shell polyphenols prevent atherosclerosis by protecting the gut barrier and remodeling the gut microbiota in ApoE(-/-) mice. Food Funct. 2021, 12, 7298–7309. [Google Scholar] [CrossRef]
- De Oliveira, J.; Hort, M.A.; Moreira, E.L.G.; Glaser, V.; Ribeiro-do-Valle, R.M.; Prediger, R.D.; Farina, M.; Latini, A.; de Bem, A.F. Positive Correlation between Elevated Plasma Cholesterol Levels and Cognitive Impairments in Ldl Receptor Knockout Mice: Relevance of Cortico-Cerebral Mitochondrial Dysfunction and Oxidative Stress. Neuroscience 2011, 197, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Song, R.; Li, Y.; Zhang, W.; Wan, Z.; Wang, Y.; Zhang, H.; Han, S. Effects of Oat Fiber Intervention on Cognitive Behavior in LDLR(-/-) Mice Modeling Atherosclerosis by Targeting the Microbiome-Gut-Brain Axis. J. Agric. Food Chem. 2020, 68, 14480–14491. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.H.; Wu, S.S.; Xia, Y.; Huang, J.Z.; Ye, J.F.; Xuan, Z.N.; Li, P.; Du, B. Probiotic-fermented black tartary buckwheat alleviates hyperlipidemia and gut microbiota dysbiosis in rats fed with a high-fat diet. Food Funct. 2021, 12, 6045–6057. [Google Scholar] [CrossRef]
- Sarma, S.M.; Singh, D.P.; Singh, P.; Khare, P.; Mangal, P.; Singh, S.; Bijalwan, V.; Kaur, J.; Mantri, S.; Boparai, R.K.; et al. Finger millet arabinoxylan protects mice from high-fat diet induced lipid derangements, inflammation, endotoxemia and gut bacterial dysbiosis. Int. J. Biol. Macromol. 2018, 106, 994–1003. [Google Scholar] [CrossRef]
- Li, S.; Liang, T.; Zhang, Y.; Huang, K.; Yang, S.Y.; Lv, H.Y.; Chen, Y.; Zhang, C.H.; Guan, X. Vitexin alleviates high-fat diet induced brain oxidative stress and inflammation via anti-oxidant, anti-inflammatory and gut microbiota modulating properties. Free Radic. Biol. Med. 2021, 171, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.M.; London, L.E.E.; Bjorndahl, T.C.; Mandal, R.; Murphy, K.; Fitzgerald, G.F.; Shanahan, F.; Ross, R.P.; Wishart, D.S.; Caplice, N.M.; et al. Microbiome and metabolome modifying effects of several cardiovascular disease interventions in apo-E-/- mice. Microbiome 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, H.; Lim, J.; Lee, H.; Yoon, S.; Shin, S.S.; Yoon, M. The lemon balm extract ALS-L1023 inhibits obesity and nonalcoholic fatty liver disease in female ovariectomized mice. Food Chem. Toxicol. 2017, 106 Pt A, 292–305. [Google Scholar] [CrossRef]
- Xiao, X.; Clark, J.M.; Park, Y. Potential contribution of insecticide exposure and development of obesity and type 2 diabetes. Food Chem. Toxicol. 2017, 105, 456–474. [Google Scholar] [CrossRef]
- Gasparotto, J.; Chaves, P.R.; da Boit Martinello, K.; Silva Oliveira, L.F.; Gelain, D.P.; Fonseca Moreira, J.C. Obesity associated with coal ash inhalation triggers systemic inflammation and oxidative damage in the hippocampus of rats. Food Chem. Toxicol. 2019, 133, 110766. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-induced gut microbiota reduces obesity in high-fat diet-fed mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar] [CrossRef]
- Ji, Y.; Ma, N.; Zhang, J.M.; Wang, H.T.; Tao, T.Y.; Pei, F.; Hu, Q.H. Dietary intake of mixture coarse cereals prevents obesity by altering the gut microbiota in high-fat diet fed mice. Food Chem. Toxicol. 2021, 147, 111901. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Mills, D.; Noratto, G. Molecular exploration of fecal microbiome in quinoa-supplemented obese mice. FEMS Microbiol. Ecol. 2016, 92, fiw089. [Google Scholar] [CrossRef] [Green Version]
- Presley, L.L.; Wei, B.; Braun, J.; Borneman, J. Bacteria Associated with Immunoregulatory Cells in Mice. Appl. Environ. Microbol. 2010, 76, 936–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, J.; Massart, S.; Vandamme, P.; De Brandt, E.; Pot, B.; Foligne, B. Ecotoxicology inside the gut: Impact of heavy metals on the mouse microbiome. BMC Pharm. Toxicol. 2013, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mazcorro, J.F.; Mills, D.A.; Murphy, K.; Noratto, G. Effect of barley supplementation on the fecal microbiota, caecal biochemistry, and key biomarkers of obesity and inflammation in obese db/db mice. Eur. J. Nutr. 2018, 57, 2513–2528. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Ivanov, I.; Mills, D.A.; Noratto, G. Influence of whole-wheat consumption on fecal microbial community structure of obese diabetic mice. PeerJ 2016, 4, e1702. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.X.; Wang, T.X.; Sun, C.; Wang, J.; Sun, B.G. Whole barley prevents obesity and dyslipidemia without the involvement of the gut microbiota in germ free C57BL/6J obese mice. Food Funct. 2019, 10, 7498–7508. [Google Scholar] [CrossRef]
- Zhong, Y.; Nyman, M.; Fak, F. Modulation of gut microbiota in rats fed high-fat diets by processing whole-grain barley to barley malt. Mol. Nutr. Food Res. 2015, 59, 2066–2076. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Song, J.; Li, J.; Wang, H.; Zhang, Y.; Suo, H. A synbiotic consisting of Lactobacillus plantarum S58 and hull-less barley beta-glucan ameliorates lipid accumulation in mice fed with a high-fat diet by activating AMPK signaling and modulating the gut microbiota. Carbohydr. Polym. 2020, 243, 116398. [Google Scholar] [CrossRef] [PubMed]
- Przybylski, P.; Pyta, K.; Ratajczak-Sitarz, M.; Katrusiak, A.; Brzezinski, B. Structure of a New Schiff Base of Gossypol with Ethyl 4-Amino-1-piperidine Carboxylate in the Solid and in the Solution. Pol. J. Chem. 2009, 83, 747–759. [Google Scholar]
- Hereu, M.; Ramos-Romero, S.; Garcia-Gonzalez, N.; Amezqueta, S.; Torres, J.L. Eubiotic effect of buckwheat D-fagomine in healthy rats. J. Funct. Foods 2018, 50, 120–126. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Diwan, V.; Brown, L. Omega-3 fatty acids and metabolic syndrome: Effects and emerging mechanisms of action. Prog. Lipid Res. 2011, 50, 372–387. [Google Scholar] [CrossRef]
- Mujico, J.R.; Baccan, G.C.; Gheorghe, A.; Diaz, L.E.; Marcos, A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br. J. Nutr. 2013, 110, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.C.; Oriach, C.S.; Murphy, K.; Moloney, G.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Stanton, C. Omega-3 polyunsaturated fatty acids critically regulate behaviour and gut microbiota development in adolescence and adulthood. Brain Behav. Immun. 2017, 59, 21–37. [Google Scholar] [CrossRef]
- Hereu, M.; Ramos-Romero, S.; Marin-Valls, R.; Amezqueta, S.; Miralles-Perez, B.; Romeu, M.; Mendez, L.; Medina, I.; Torres, J.L. Combined Buckwheat D-Fagomine and Fish Omega-3 PUFAs Stabilize the Populations of Gut Prevotella and Bacteroides While Reducing Weight Gain in Rats. Nutrients 2019, 11, 2606. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Zhang, Q.; Zhang, Y.; Yao, Z.; Song, P.; Wei, L.; Zhao, G.; Yan, Z. Effect of tartary buckwheat, rutin, and quercetin on lipid metabolism in rats during high dietary fat intake. Food Sci. Nutr. 2020, 8, 199–213. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, S.; Jiang, Y.; Wei, Y.; Zhou, X. Regulatory Function of Buckwheat-Resistant Starch Supplementation on Lipid Profile and Gut Microbiota in Mice Fed with a High-Fat Diet. J. Food Sci. 2019, 84, 2674–2681. [Google Scholar] [CrossRef]
- Zhou, Y.; Wei, Y.; Yan, B.; Zhao, S.; Zhou, X. Regulation of tartary buckwheat-resistant starch on intestinal microflora in mice fed with high-fat diet. Food Sci. Nutr. 2020, 8, 3243–3251. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.M.; Khare, P.; Jagtap, S.; Singh, D.P.; Baboota, R.K.; Podili, K.; Boparai, R.K.; Kaur, J.; Bhutani, K.K.; Bishnoi, M.; et al. Kodo millet whole grain and bran supplementation prevents high-fat diet induced derangements in a lipid profile, inflammatory status and gut bacteria in mice. Food Funct. 2017, 8, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Awika, J.M.; McDonough, C.M.; Rooney, L.W. Decorticating sorghum to concentrate healthy phytochemicals. J. Agric. Food Chem. 2005, 53, 6230–6234. [Google Scholar] [CrossRef]
- Ashley, D.; Marasini, D.; Brownmiller, C.; Lee, J.A.; Carbonero, F.; Lee, S.O. Impact of Grain Sorghum Polyphenols on Microbiota of Normal Weight and Overweight/Obese Subjects during In Vitro Fecal Fermentation. Nutrients 2019, 11, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Yang, C.; Wai, S.T.C.; Zhang, Y.; Portillo, M.P.; Paoli, P.; Wu, Y.; San Cheang, W.; Liu, B.; Carpene, C.; et al. Regulation of glucose metabolism by bioactive phytochemicals for the management of type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2019, 59, 830–847. [Google Scholar] [CrossRef]
- Della Pepa, G.; Vetrani, C.; Vitale, M.; Riccardi, G. Wholegrain Intake and Risk of Type 2 Diabetes: Evidence from Epidemiological and Intervention Studies. Nutrients 2018, 10, 1288. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Wang, L.X.; Chen, Z.L.; Hou, D.A.Z.; Xue, Y.; Diao, X.M.; Shen, Q. Foxtail Millet Improves Blood Glucose Metabolism in Diabetic Rats through PI3K/AKT and NF-kappa B Signaling Pathways Mediated by Gut Microbiota. Nutrients 2021, 13, 1837. [Google Scholar] [CrossRef]
- Fu, Y.X.; Yin, R.Y.; Guo, E.H.; Cheng, R.H.; Diao, X.M.; Xue, Y.; Shen, Q. Protein Isolates from Raw and Cooked Foxtail Millet Attenuate Development of Type 2 Diabetes in Streptozotocin-Induced Diabetic Mice. Mol. Nutr. Food Res. 2021, 65, 2000365. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Govoni, S.; Coppola, A.; Gazzaruso, C. The role of gut microbiota in obesity, diabetes mellitus, and effect of metformin: New insights into old diseases. Curr. Opin. Pharm. 2019, 49, 1–5. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Gallego, C.; Collado, M.C.; Perez, G.; Ilo, T.; Jaakkola, U.M.; Bernal, M.J.; Periago, M.J.; Frias, R.; Ros, G.; Salminen, S. Resembling breast milk: Influence of polyamine-supplemented formula on neonatal BALB/cOlaHsd mouse microbiota. Br. J. Nutr. 2014, 111, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vojinovic, D.; Radjabzadeh, D.; Kurilshikov, A.; Amin, N.; Wijmenga, C.; Franke, L.; Ikram, M.A.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Relationship between gut microbiota and circulating metabolites in population-based cohorts. Nat. Commun. 2019, 10, 5813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenberg, C.; Savage, D.B. An amino acid profile to predict diabetes? Nat. Med. 2011, 17, 418–420. [Google Scholar] [CrossRef]
- Chang, D.H.; Rhee, M.S.; Ahn, S.; Bang, B.H.; Oh, J.E.; Lee, H.K.; Kim, B.C. Faecalibaculum rodentium gen. nov. sp. nov. isolated from the faeces of a laboratory mouse. Antonie Van Leeuwenhoek 2015, 108, 1309–1318. [Google Scholar] [CrossRef]
- Fu, Y.X.; Yin, R.Y.; Liu, Z.Y.; Niu, Y.; Guo, E.H.; Cheng, R.H.; Diao, X.M.; Xue, Y.; Shen, Q. Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice. Nutrients 2020, 12, 3452. [Google Scholar] [CrossRef]
- Zheng, J.P.; Zhu, L.; Hu, B.F.; Zou, X.J.; Hu, H.M.; Zhang, Z.G.; Jiang, N.; Ma, J.; Yang, H.B.; Liu, H.T. 1-Deoxynojirimycin improves high fat diet-induced nonalcoholic steatohepatitis by restoring gut dysbiosis. J. Nutr. Biochem. 2019, 71, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; Francois, P.; de Vos, W.M.; et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and Diet-Induced Leptin-Resistant Mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lian, F.M.; Zhao, L.H.; Zhao, Y.F.; Chen, X.Y.; Zhang, X.; Guo, Y.; Zhang, C.H.; Zhou, Q.; Xue, Z.S.; et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J. 2015, 9, 552–562. [Google Scholar] [CrossRef]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al. Longitudinal multi-omics of host-microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Q.; Liu, M.; Zhang, X.; He, F.; Wang, G. Akkermansia muciniphila can reduce the damage of gluco/lipotoxicity, oxidative stress and inflammation, and normalize intestine microbiota in streptozotocin-induced diabetic rats. Pathog. Dis. 2018, 76, fty028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, Y.F.; Xu, J.; Xue, Z.S.; Zhang, M.H.; Pang, X.Y.; Zhang, X.J.; Zhao, L.P. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5, 14405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Casas, J.; Taltavull, N.; Romeu, M.; Amezqueta, S.; Dasilva, G.; Medina, I.; Torres, J.L. Functional Effects of the Buckwheat Iminosugar D-Fagomine on Rats with Diet-Induced Prediabetes. Mol. Nutr. Food Res. 2018, 62, e1800373. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Kristek, A.; Wiese, M.; Heuer, P.; Kosik, O.; Schar, M.Y.; Soycan, G.; Alsharif, S.; Kuhnle, G.G.C.; Walton, G.; Spencer, J.P.E. Oat bran, but not its isolated bioactive beta-glucans or polyphenols, have a bifidogenic effect in an in vitro fermentation model of the gut microbiota. Br. J. Nutr. 2019, 121, 549–559. [Google Scholar] [CrossRef]
- Gullon, B.; Gullon, P.; Tavaria, F.K.; Yanez, R. Assessment of the prebiotic effect of quinoa and amaranth in the human intestinal ecosystem. Food Funct. 2016, 7, 3782–3788. [Google Scholar] [CrossRef]
- Bekedam, E.K.; Roos, E.; Schols, H.A.; van Boekel, M.A.J.S.; Smit, G. Low molecular weight melanoidins in coffee brew. J. Agric. Food Chem. 2008, 56, 4060–4067. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Bellesia, A. The gastro-intestinal tract as the major site of biological action of dietary melanoidins. Amino Acids 2015, 47, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Aljahdali, N.; Gadonna-Widehem, P.; Anton, P.M.; Carbonero, F. Gut Microbiota Modulation by Dietary Barley Malt Melanoidins. Nutrients 2020, 12, 241. [Google Scholar] [CrossRef] [Green Version]
- Voreades, N.; Kozil, A.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velikonja, A.; Lipoglavsek, L.; Zorec, M.; Orel, R.; Avgustin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front. Microbiol. 2018, 9, 737. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Lam, K.L.; Li, X.J.; Kong, A.P.S.; Cheung, P.C.K. Circadian disruption-induced metabolic syndrome in mice is ameliorated by oat beta-glucan mediated by gut microbiota. Carbohydr. Polym. 2021, 267, 118216. [Google Scholar] [CrossRef]
- Dahlhausen, K.E.; Doroud, L.; Firl, A.J.; Polkinghorne, A.; Eisen, J.A. Characterization of shifts of koala (Phascolarctos cinereus) intestinal microbial communities associated with antibiotic treatment. PeerJ 2018, 6, e4452. [Google Scholar] [CrossRef]
- Sun, L.J.; Ma, L.J.; Ma, Y.B.; Zhang, F.M.; Zhao, C.H.; Nie, Y.Z. Insights into the role of gut microbiota in obesity: Pathogenesis, mechanisms, and therapeutic perspectives. Protein Cell 2018, 9, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.Y.; Li, J.; Wei, C.H.; Luo, T.; Deng, Z.Y.; Fan, Y.W.; Zheng, L.F. A polysaccharide from Fagopyrum esculentum Moench bee pollen alleviates microbiota dysbiosis to improve intestinal barrier function in antibiotic-treated mice. Food Funct. 2020, 11, 10519–10533. [Google Scholar] [CrossRef]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Salbaum, J.M.; Luo, M.; Blanchard, E.T.; Taylor, C.M.; Welsh, D.A.; Berthoud, H.R. Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol. Psychiatry 2015, 77, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.L.; Yu, Y.H.; Lin, D.H.; Zheng, P.; Zhang, P.; Hu, M.M.; Wang, Q.; Pan, W.; Yang, X.Y.; Hu, T.; et al. beta-glucan attenuates cognitive impairment via the gut-brain axis in diet-induced obese mice. Microbiome 2020, 8, 1–21. [Google Scholar] [CrossRef]
- Saji, N.; Niida, S.; Murotani, K.; Hisada, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Toba, K.; Sakurai, T. Analysis of the relationship between the gut microbiome and dementia: A cross-sectional study conducted in Japan. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.F.; Yu, Y.H.; Beck, E.J.; South, T.; Li, Y.L.; Batterham, M.J.; Tapsell, L.C.; Chen, J.Z. Diet high in oat beta-glucan activates the gut-hypothalamic (PYY3-36-NPY) axis and increases satiety in diet-induced obesity in mice. Mol. Nutr. Food Res. 2011, 55, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Mattson, M.P. The impact of dietary energy intake on cognitive aging. Front. Aging Neurosci. 2010, 2. [Google Scholar] [CrossRef]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Maternal and Child Nutrition Study Group. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 396. [Google Scholar] [CrossRef]
- Li, X.J.; Hui, Y.; Leng, B.F.; Ren, J.L.; Song, Y.N.; Che, L.Q.; Peng, X.; Huang, B.J.; Liu, S.L.; Li, L.; et al. Millet-based supplement restored gut microbial diversity of acute malnourished pigs. PLoS ONE 2021, 16, e0250423. [Google Scholar] [CrossRef]
- Gehrig, J.L.; Venkatesh, S.; Chang, H.W.; Hibberd, M.C.; Kung, V.L.; Cheng, J.Y.; Chen, R.Y.; Subramanian, S.; Cowardin, C.A.; Meier, M.F.; et al. Effects of microbiota-directed foods in gnotobiotic animals and undernourished children. Science 2019, 365, eaau4732. [Google Scholar] [CrossRef] [Green Version]
- Umu, O.C.O.; Fauske, A.K.; Akesson, C.P.; de Nanclares, M.P.; Sorby, R.; Press, C.M.; Overland, M.; Sorum, H. Gut microbiota profiling in Norwegian weaner pigs reveals potentially beneficial effects of a high-fiber rapeseed diet. PLoS ONE 2018, 13, e0209439. [Google Scholar] [CrossRef] [Green Version]
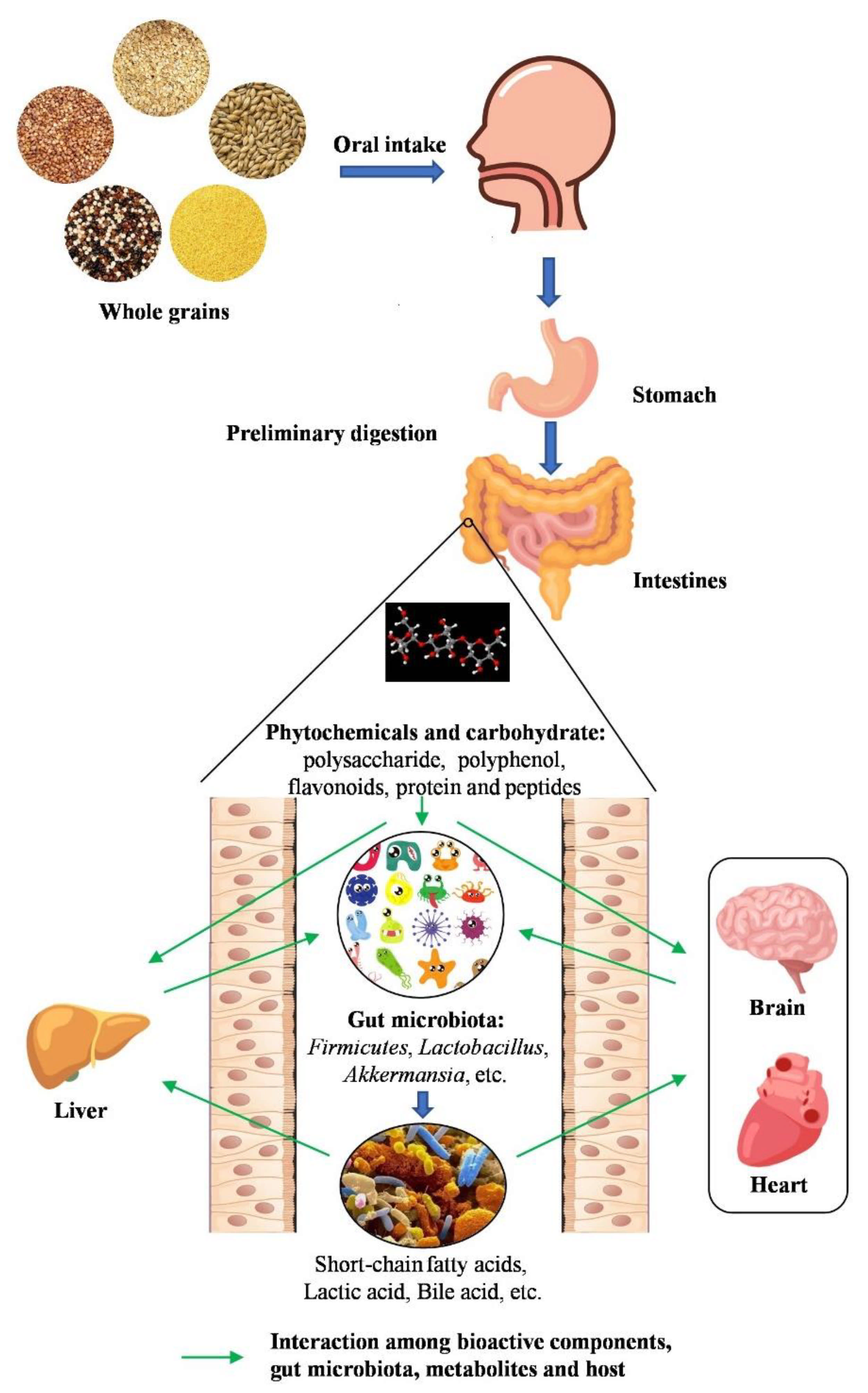
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).