
Investigation of the Molecular Profile of Granular Cell Tumours and Schwannomas of the Oral Cavity


Abstract
1. Introduction
2. Results
2.1. Demographics
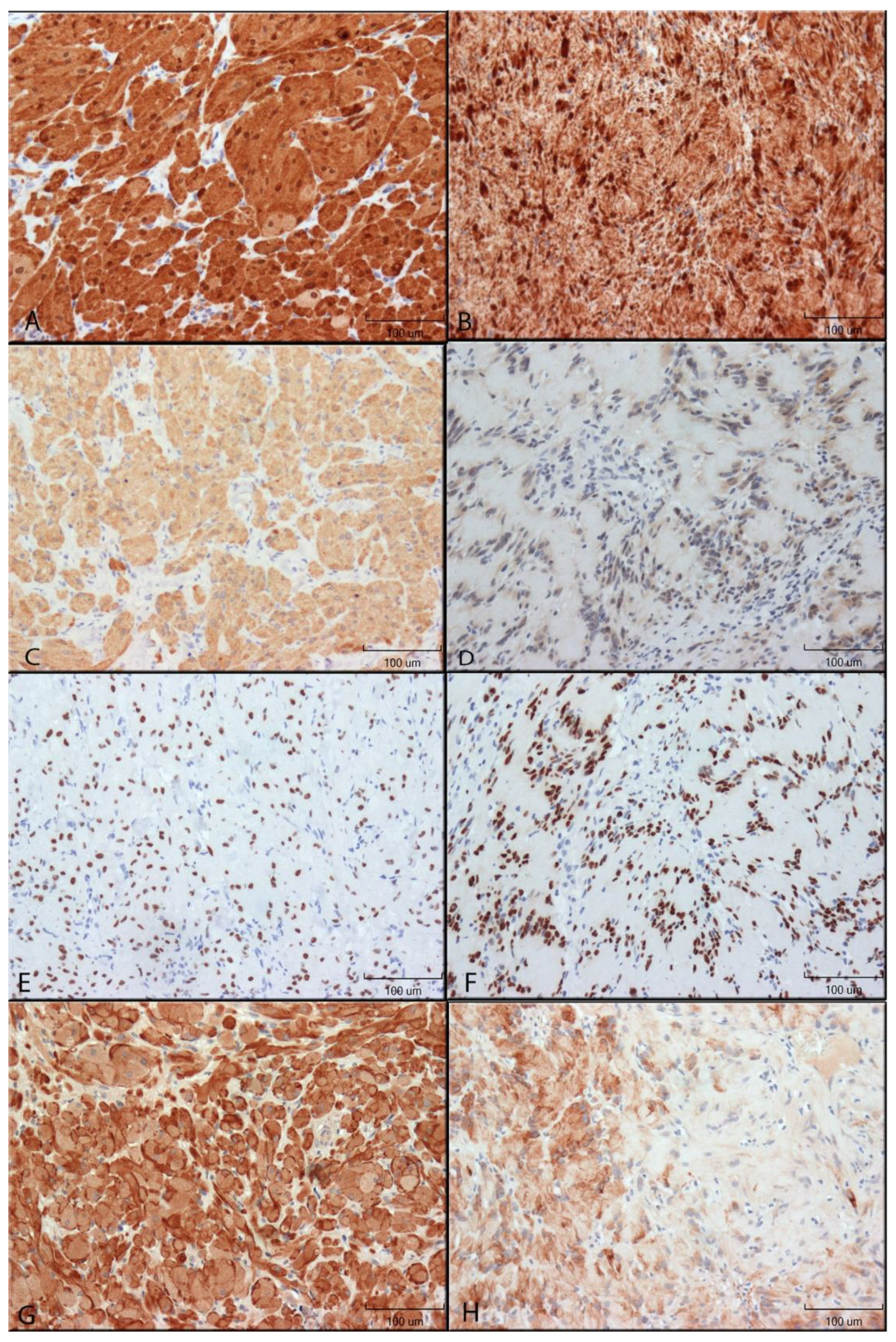
2.2. Immunohistochemistry
2.3. Summary of Cq Value Obtained Using RT-qPCR
3. Discussion
3.1. Protein Expression Supporting an NCC Origin of GCTs
3.2. Protein Expression Supporting an APC Phenotype
3.3. Interpretation of RT-qPCR Data
3.4. Limitations of This Study
3.5. Future Work
4. Material and Methods
4.1. Case Selection
4.2. Immunohistochemistry (IHC)
4.3. Evaluation of Immunostaining
4.3.1. Manual IHC Scoring
4.3.2. IHC Digital Analysis
4.4. Protocol for RT-qPCR
4.4.1. Tissue Preparation
4.4.2. RNA Isolation
4.4.3. RNA Quantification
4.4.4. cDNA Synthesis and RT-qPCR
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ordóñez, N.G.; Mackay, B. Granular cell tumor: A review of the pathology and histogenesis. Ultrastruct. Pathol. 1999, 23, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Allon, I.; Kaplan, I.; Gal, G.; Chaushu, G.; Allon, D.M. The clinical characteristics of benign oral mucosal tumors. Med. Oral Patol. Oral Cir. Bucal. 2014, 19, e438. [Google Scholar] [CrossRef] [PubMed]
- Speight, P.M. World Health Organization Classification of Tumors: Pathology and Genetics of Head and Neck Tumors; Barnes, L., Everson, J., Reichart, P.S.D., Eds.; IARC Press: Lyon, France, 2005. [Google Scholar]
- Goodstein, M.L.; Eisele, D.W.; Hyams, V.J.; Kashima, H.K. Multiple synchronous granular cell tumors of the upper aerodigestive tract. Otolaryngol. Head Neck Surg. 1990, 103, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Barakat, M.; Kar, A.A.; Pourshahid, S.; Ainechi, S.; Lee, H.J.; Othman, M.; Tadros, M. Gastrointestinal and biliary granular cell tumor: Diagnosis and management. Ann. Gastroenterol. 2018, 31, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Porta, N.; Mazzitelli, R.; Cacciotti, J.; Cirenza, M.; Labate, A.; Lo Schiavo, M.G.; Laghi, A.; Petrozza, V.; Della Rocca, C. A case report of a rare intramuscular granular cell tumor. Diagn. Pathol. 2015, 10, 162. [Google Scholar] [CrossRef]
- Curtis, R.; Stewart, H.J.S.; Hall, S.M.; Wilkin, G.P.; Mirsky, R.; Jessen, K.R. GAP-43 is expressed by nonmyelin-forming Schwann cells of the peripheral nervous system. J. Cell Biol. 1992, 116, 1455–1464. [Google Scholar] [CrossRef]
- Rivlin, M.E.; Meeks, G.R.; Ghafar, M.A.; Lewin, J.R. Vulvar granular cell tumor. World J. Clin. Cases 2013, 1, 149–151. [Google Scholar] [CrossRef]
- Gurzu, S.; Ciortea, D.; Tamasi, A.; Golea, M.; Bodi, A.; Sahlean, D.I.; Kovecsi, A.; Jung, I. The immunohistochemical profile of granular cell (Abrikossoff) tumor suggests an endomesenchymal origin. Arch. Dermatol. Res. 2015, 307, 151–157. [Google Scholar] [CrossRef]
- Lack, E.E.; Worsham, R.G.F.; Callihan, M.D.; Crawford, B.E.; Klappenbach, S.; Rowden, G.; Chun, B. Granular cell tumor: A clinicopathologic study of 110 patients. J. Surg. Oncol. 1980, 13, 301–316. [Google Scholar] [CrossRef]
- Worsaae, N.; Schwartz, O.; Pindborg, J.J. Follow-up study of 14 oral granular cell tumors. Int. J. Oral Surg. 1979, 8, 133–139. [Google Scholar] [CrossRef]
- Ferreira, J.C.B.; Oton-Leite, A.F.; Guidi, R.; Mendonça, E.F. Granular cell tumor mimicking a squamous cell carcinoma of the tongue: A case report. BMC Res. Notes 2017, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.S.; Leifer, C.; Chen, S.Y.; Harwick, R.D. Oral granular-cell tumors. Report of twenty-five cases with electron microscopy. Oral Surg. Oral Med. Oral Pathol. 1977, 44, 227–237. [Google Scholar] [CrossRef]
- Sobel, H.J.; Marquet, E.; Avrin, E.; Schwarz, R. Granular cell myoblastoma. An electron microscopic and cytochemical study illustrating the genesis of granules and aging of myoblastoma cells. Am. J. Pathol. 1971, 65, 59–78. [Google Scholar] [PubMed]
- Manara, G.C.; De Panfilis, G.; Bacchi, A.B.; Ferrari, C.; Tedeschi, F.; Brusati, R.; Scandroglio, R.; Allegra, F. Fine structure of granular cell tumor of abrikossoff. J. Cutan. Pathol. 1981, 8, 277–282. [Google Scholar] [CrossRef]
- Stewart, C.M.; Watson, R.E.; Eversole, L.R.; Fischlschweiger, W.; Leider, A.S. Oral granular cell tumors: A clinicopathologic and immunocytochemical study. Oral Surg. Oral Med. Oral Pathol. 1988, 65, 427–435. [Google Scholar] [CrossRef]
- Mukai, M. Immunohistochemical localization of S-100 protein and peripheral nerve myelin proteins (P2 protein, P0 protein) in granular cell tumors. Am. J. Pathol. 1983, 112, 139. [Google Scholar]
- Musha, A.; Ogawa, M.; Yokoo, S. Granular cell tumors of the tongue: Fibroma or schwannoma. Head Face Med. 2018, 14. [Google Scholar] [CrossRef]
- Campbell, L.K.; Thomas, J.R.; Lamps, L.W.; Smoller, B.R.; Folpe, A.L. Protein Gene Product 9.5 (PGP 9.5) Is Not a Specific Marker of Neural and Nerve Sheath Tumors: An Immunohistochemical Study of 95 Mesenchymal Neoplasms. Mod. Pathol. 2003, 16, 963–969. [Google Scholar] [CrossRef]
- Martins, M.D.; De Jesus, L.A.; Fernandes, K.P.S.; Bussadori, S.K.; Taghloubi, S.A.; Martins, M.A.T. Intra-oral schwannoma: Case report and literature review. Indian J. Dent. Res. 2009, 20, 121–125. [Google Scholar] [CrossRef]
- Shintaku, M. Immunohistochemical localization of autophagosomal membrane-associated protein LC3 in granular cell tumor and schwannoma. Virchows Arch. 2011, 459, 315–319. [Google Scholar] [CrossRef][Green Version]
- Regezi, J.A.; Zarbo, R.J.; Courtney, R.M.; Crissman, J.D. Immunoreactivity of granular cell lesions of skin, mucosa, and jaw. Cancer 1989, 64, 1455–1460. [Google Scholar] [CrossRef]
- Pareja, F.; Brandes, A.H.; Basili, T.; Selenica, P.; Geyer, F.C.; Fan, D.; Da Cruz Paula, A.; Kumar, R.; Brown, D.N.; Gularte-Mérida, R.; et al. Loss-of-function mutations in ATP6AP1 and ATP6AP2 in granular cell tumors. Nat. Commun. 2018, 9, 3533. [Google Scholar] [CrossRef]
- Choo, S.Y. The HLA system: Genetics, immunology, clinical testing, and clinical implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef]
- Bergstrom, R.T.; Silverman, D.A.; Chambers, K.; Kim, J.A. CD40 monoclonal antibody activation of antigen-presenting cells improves therapeutic efficacy of tumor-specific T cells. Otolaryngol. Head Neck Surg. 2004, 130, 94–103. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Killingsworth, M.C.; Myasoedova, V.A.; Orekhov, A.N.; Bobryshev, Y.V. CD68/macrosialin: Not just a histochemical marker. Lab. Investig. 2017, 97, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, E.; Kunz-Schughart, L.A.; Weber, A.; Rehli, M.; Peuker, A.; Müller, A.; Kastenberger, M.; Brockhoff, G.; Andreesen, R.; Kreutz, M. Expression of CD68 in non-myeloid cell types. Scand. J. Immunol. 2008, 67, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Etzerodt, A.; Moestrup, S.K. CD163 and inflammation: Biological, diagnostic, and therapeutic aspects. Antioxid. Redox Signal. 2013, 18, 2352–2363. [Google Scholar] [CrossRef] [PubMed]
- Chatzigeorgiou, A.; Lyberi, M.; Chatzilymperis, G.; Nezos, A.; Kamper, E. CD40/CD40L signaling and its implication in health and disease. Biofactors 2009, 35, 474–483. [Google Scholar] [CrossRef]
- Wu, J.; Wu, H.; An, J.; Ballantyne, C.M.; Cyster, J.G. Critical role of integrin CD11c in splenic dendritic cell capture of missing-self CD47 cells to induce adaptive immunity. Proc. Natl. Acad. Sci. USA 2018, 115, 6786–6791. [Google Scholar] [CrossRef]
- Male, D.K. Immunology; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 978-0-7020-5028-2. [Google Scholar]
- Surbhi, S.; Metgud, R.; Naik, S.; Patel, S. Spindle cell lesions: A review on immunohistochemical markers. J. Cancer Res. Ther. 2017, 13, 412–418. [Google Scholar] [CrossRef]
- Karamchandani, J.R.; Nielsen, T.O.; Van De Rijn, M.; West, R.B. Sox10 and s100 in the diagnosis of soft-tissue neoplasms. Appl. Immunohistochem. Mol. Morphol. 2012, 20, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Isgrò, M.A.; Bottoni, P.; Scatena, R. Neuron-specifi c enolase as a biomarker: Biochemical and clinical aspects. Adv. Exp. Med. Biol. 2015, 867, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Perrone-Bizzozero, N.I.; Neve, R.L.; Irwin, N.; Lewis, S.; Fischer, I.; Benowitz, L.I. Post-transcriptional regulation of GAP-43 rnRNA levels during neuronal differentiation and nerve regeneration. Mol. Cell. Neurosci. 1991, 2, 402–409. [Google Scholar] [CrossRef]
- Denny, J. Molecular Mechanisms, Biological Actions, and Neuropharmacology of the Growth-Associated Protein GAP-43. Curr. Neuropharmacol. 2006, 4, 293–304. [Google Scholar] [CrossRef]
- Rizzino, A. Sox2 and Oct-3/4: A versatile pair of master regulators that orchestrate the self-renewal and pluripotency of embryonic stem cells. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 228–236. [Google Scholar] [CrossRef]
- Jo, A.; Denduluri, S.; Zhang, B.; Wang, Z.; Yin, L.; Yan, Z.; Kang, R.; Shi, L.L.; Mok, J.; Lee, M.J.; et al. The versatile functions of Sox9 in development, stem cells, and human diseases. Genes Dis. 2014, 1, 149–161. [Google Scholar] [CrossRef]
- Michalczyk, K.; Ziman, M. Nestin structure and predicted function in cellular cytoskeletal organisation. Histol. Histopathol. 2005, 20, 665–671. [Google Scholar] [CrossRef]
- Erinanç, H.; Göktürk, H.S.; Kanat Ünler, G.; Karagülle, E. Utility of Nestin immunohistochemistry in the diagnosis of granular cell tumor. Arch. Clin. Exp. Med. 2018, 3, 160–164. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, J.; Lee, D.Y.; Kim, Y.D.; Kim, J.Y.; Lim, H.J.; Lim, S.; Cho, Y.S. Schwann Cell Precursors from Human Pluripotent Stem Cells as a Potential Therapeutic Target for Myelin Repair. Stem. Cell Rep. 2017, 8, 1714–1726. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Del Valle, L.; Geddes, J.F.; Aldape, K.; Boyd, J.C.; Legido, A.; Khalili, K.; Perentes, E.; Mörk, S.J. Localization of the neuronal class III β-tubulin in oligodendrogliomas: Comparison with Ki-67 proliferative index and 1p/19q status. J. Neuropathol. Exp. Neurol. 2002, 61, 307–320. [Google Scholar] [CrossRef]
- Lebok, P.; Öztürk, M.; Heilenkötter, U.; Jaenicke, F.; Müller, V.; Paluchowski, P.; Geist, S.; Wilke, C.; Burandt, E.; Lebeau, A.; et al. High levels of class III β-tubulin expression are associated with aggressive tumor features in breast cancer. Oncol. Lett. 2016, 11, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Kuberappa, P.H.; Bagalad, B.S.; Ananthaneni, A.; Kiresur, M.A.; Srinivas, G.V. Certainty of S100 from Physiology to Pathology. J. Clin. Diagn. Res. 2016, 10, ZE10-5. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 Proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Pekmezci, M.; Folpe, A.L.; Ersen, A.; Horvai, A.E. Diagnostic utility of SOX10 to distinguish malignant peripheral nerve sheath tumor from synovial sarcoma, including intraneural synovial sarcoma. Mod. Pathol. 2014, 27, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wisell, J.; Kleinschmidt-DeMasters, B.K. Sox10 nuclear immunostaining lacks diagnostic utility for CNS granular cell tumors. J. Neuropathol. Exp. Neurol. 2014, 73, 98–100. [Google Scholar] [CrossRef][Green Version]
- Solomon, L.W. S-100 Negative Granular Cell Tumor of the Oral Cavity. Head Neck Pathol. 2018, 10, 367–373. [Google Scholar] [CrossRef]
- Roberts, S.L.; Dun, X.P.; Doddrell, R.D.S.; Mindos, T.; Drake, L.K.; Onaitis, M.W.; Florio, F.; Quattrini, A.; Lloyd, A.C.; D’Antonio, M.; et al. Sox2 expression in schwann cells inhibits myelination in vivo and induces influx of macrophages to the nerve. Development 2017, 144, 3114–3125. [Google Scholar] [CrossRef]
- Vinores, S.A.; Herman, M.M.; Rubinstein, L.J. Electron-immunocytochemical localization of neuron-specific enolase in cytoplasm and on membranes of primary and metastatic cerebral tumours and on glial filaments of glioma cells. Histopathology 1986, 10, 891–908. [Google Scholar] [CrossRef]
- Mjønes, P.; Sagatun, L.; Nordrum, I.S.; Waldum, H.L. Neuron-Specific Enolase as an Immunohistochemical Marker Is Better Than Its Reputation. J. Histochem. Cytochem. 2017, 65, 687–703. [Google Scholar] [CrossRef]
- Chen, W.; Chen, P.; Lu, D.; Lind, A.C.; Dehner, L.P. Growth-associated protein 43 in differentiating peripheral nerve sheath tumors from other non-neural spindle cell neoplasms. Mod. Pathol. 2014, 1, 184–193. [Google Scholar] [CrossRef]
- Zheng, C.; Quan, R.-D.; Wu, C.-Y.; Hu, J.; Lin, B.-Y.; Dong, X.-B.; Xia, E.-J.; Bhandari, A.; Zhang, X.-H.; Wang, O.-C. Growth-associated protein 43 promotes thyroid cancer cell lines progression via epithelial-mesenchymal transition. J. Cell. Mol. Med. 2019, 23, 7974–7984. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jin, J.; Wu, J.; Chen, K.; Huang, M.; Yu, H.; Ying, L.; Su, D. Abstract 1083: GAP43, a novel potential metastases promoter in non-small cell lung cancer. Cancer Res. 2018, 1083. [Google Scholar] [CrossRef]
- Klein, A.; Olendrowitz, C.; Schmutzler, R.; Hampl, J.; Schlag, P.M.; Maass, N.; Arnold, N.; Wessel, R.; Ramser, J.; Meindl, A.; et al. Identification of brain- and bone-specific breast cancer metastasis genes. Cancer Lett. 2009, 276, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Wippold, F.J.; Lubner, M.; Perrin, R.J.; Lämmle, M.; Perry, A. Neuropathology for the neuroradiologist: Antoni A and Antoni B tissue patterns. Am. J. Neuroradiol. 2007, 28, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Papiez, J.; Rojiani, M.V.; Rojiani, A.M. Vascular alterations in schwannoma. Int. J. Clin. Exp. Pathol. 2014, 7, 4032. [Google Scholar]
- Kurtin, P.J.; Bonin, D.M. Immunohistochemical demonstration of the lysosome-associated glycoprotein CD68 (KP-1) in granular cell tumors and schwannomas. Hum. Pathol. 1994, 25, 1172–1178. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Chang, W.-A.; Lin, E.-S.; Chen, Y.-J.; Kuo, P.-L. Expressions of HLA Class II Genes in Cutaneous Melanoma Were Associated with Clinical Outcome: Bioinformatics Approaches and Systematic Analysis of Public Microarray and RNA-Seq Datasets. Diagnostics 2019, 9, 59. [Google Scholar] [CrossRef]
- Mehrfeld, C.; Zenner, S.; Kornek, M.; Lukacs-kornek, V.; Lukacs-Kornek, V. The Contribution of Non-Professional Antigen-Presenting Cells to immunity and Tolerance in the Liver. Front. Immunol. 2018, 9, 635. [Google Scholar] [CrossRef]
- Wosen, J.E.; Mukhopadhyay, D.; MacAubas, C.; Mellins, E.D. Epithelial MHC class II expression and its role in antigen presentation in the gastrointestinal and respiratory tracts. Front. Immunol. 2018, 9, 2144. [Google Scholar] [CrossRef]
- Epstein, S.P.; Gadaria-Rathod, N.; Wei, Y.; Maguire, M.G.; Asbell, P.A. HLA-DR expression as a biomarker of inflammation for multicenter clinical trials of ocular surface disease. Exp. Eye Res. 2013, 111, 95–104. [Google Scholar] [CrossRef]
- de Muralt, B.; de Tribolet, N.; Diserens, A.C.; Stavrou, D.; Mach, J.P.; Carrel, S. Phenotyping of 60 cultured human gliomas and 34 other neuroectodermal tumors by means of monoclonal antibodies against glioma, melanoma and HLA-DR antigens. Eur. J. Cancer Clin. Oncol. 1985, 21, 204–216. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed]
- Loughrey, M.B.; Bankhead, P.; Coleman, H.G.; Hagan, R.S.; Craig, S.; McCorry, A.M.B.; Gray, R.T.; McQuaid, S.; Dunne, P.D.; Hamilton, P.W.; et al. Validation of the systematic scoring of immunohistochemically stained tumour tissue microarrays using QuPath digital image analysis. Histopathology 2018, 73, 327–338. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Granular Cell Tumours | |||
|---|---|---|---|
| Case No. | Age | Sex | Site |
| 1 | 41 | Female | midright dorsum of tongue |
| 2 | 33 | Female | dorsum of tongue |
| 3 | 15 | Female | dorsum of tongue |
| 4 | 43 | Male | dorsum of tongue |
| 5 | 20 | female | right tongue |
| 6 | 36 | Male | dorsum of tongue |
| 7 | 48 | female | left posterior hard palate |
| 8 | 50 | Male | dorsum of tongue |
| 9 | 37 | female | left lateral border of tongue |
| 10 | 55 | female | right palatal mucosa |
| 11 | 39 | female | dorsum of tongue |
| 12 | 21 | female | right dorsum of tongue |
| 13 | 52 | male | right lateral border of tongue |
| 14 | 48 | male | right dorsum of tongue |
| 15 | 46 | male | right dorsum of tongue |
| 16 | 35 | female | right dorsum of tongue |
| 17 | 63 | female | left dorsum of tongue |
| 18 | 55 | female | left dorsum of tongue |
| 19 | 37 | male | dorsum of tongue |
| 20 | 21 | female | right lateral border of tongue |
| 21 | 56 | male | dorsum of tongue |
| 22 | 58 | male | right lateral ventral tongue |
| 23 | 43 | Female | left dorsal tongue |
| Schwannomas | |||
| Case No. | Age | Sex | Site |
| 1 | 28 | Male | buccal mucosa 37 |
| 2 | 21 | Male | left dorsum of tongue |
| 3 | 23 | Male | right tip of tongue |
| 4 | 41 | Male | left buccal mucosa |
| 5 | 16 | Female | *** oral cavity |
| 6 | 37 | Female | lower lip mucosa |
| 7 | 22 | Male | lower lip mucosa |
| 8 | 15 | Male | left buccal vestibule |
| 9 | 16 | Male | dorsum of tongue |
| 10 | 31 | Male | Upper lip |
| IHC Antibody | Absent | Low | Medium | High | 0 (0%) | 1 (>0≤50%) | 2 (>50%) | |
|---|---|---|---|---|---|---|---|---|
| S100 | GCT | - | - | - | 23 | - | - | 23 |
| Schwannoma | - | - | - | 10 | - | - | 10 | |
| NSE | GCT | - | - | 3 | 20 | - | - | 23 |
| Schwannoma | - | 4 | 6 | - | - | - | 10 | |
| SOX10 | GCT | 1 | - | - | 22 | 1 | - | 22 |
| Schwannoma | - | - | - | 10 | - | - | 10 | |
| GAP43 | GCT | - | - | 2 | 21 | - | - | 23 |
| Schwannoma | - | - | 4 | 6 | - | 3 | 7 | |
| HLA-DR | GCT | - | - | 6 | 17 | - | - | 23 |
| Schwannoma | - | - | 4 | 6 | - | - | 10 | |
| CD68 | GCT | - | - | 17 | 6 | - | - | 23 |
| Schwannoma | 1 | 6 | 3 | - | 1 | 6 | 3 | |
| CD163 | GCT * | 22 * | - | - | - | 22 * | - | - |
| Schwannoma | - | - | 1 | 9 | - | - | 10 | |
| CD40 | GCT | 23 | - | - | 23 | - | - | |
| Schwannoma | 10 | - | - | 10 | - | - | ||
| CD11c | GCT | 23 | - | - | 23 | - | - | |
| Schwannoma | 10 | - | - | 10 | - | - |
| Antibody | Correlation (r) | 95% CI | p-Value |
|---|---|---|---|
| All tumours | 0.906 | 0.827–0.949 | 0.000000000000001 |
| GCTs only | 0.903 | 0.767–0.961 | 0.00000005 |
| Schwannomas only | 0.9 | 0.759–0.960 | 0.00000007 |
| GAP43 | 0.92 | 0.689–0.981 | 0.0002 |
| HLA-DR | 0.961 | 0.841–0.991 | 0.000009 |
| CD68 | 0.989 | 0.951–0.997 | 0.00000007 |
| CD163 | 0.979 | 0.911–0.979 | 0.0000008 |
| Case | RNA Concentration (µg/mL) | B-Actin Reactivity (Cq Value) | CD68 Reactivity (Cq Value) |
|---|---|---|---|
| 1 | 6.5 | 34.70 | - |
| 3 | 17.4 | 37.38 | 37.15 |
| 6 | 4.24 * | ||
| 7 | 2.88 * | ||
| 8 | 5.7 | 36.94 | 35.96 |
| 15 | 7.4 | 34.53 | 37.12 |
| 16 | Low * | ||
| 17 | 2.2 * | ||
| 18 | 10 | 29.71 | 32.53 |
| 22 | 4.16 * | ||
| 23 | 6.5 | 28.89 | 25.60 |
| NTC | - | - |
| Gene | Cq Value |
|---|---|
| CD68 | 29.60 |
| Nestin | 39.02 |
| SOX2 | - |
| SOX9 | 36.18 |
| SOX10 | 38.01 |
| NF2 | 37.14 |
| GFAP | - |
| Beta Actin | 28.89 |
| Beta TUB3 | 36.72 |
| Antibody | Manufacture/Cat Number | Antibody Type | Constituent Tissue and Labelling Targets | Dilution |
|---|---|---|---|---|
| S100 | Dako, Santa Clara CA IR50461-2 | Rabbit polyclonal | Neural tissues, S100B (strong) S100A1 and S100A6 (weak) | Automated |
| NSE | Dako, Santa Clara CA IR61261-2 | Mouse monoclonal | Neural tissues, γ-enolase subunit | Automated |
| SOX10 | Santa Cruz Biotech, Dallas TX, sc-365692 | Mouse monoclonal | Neural tissues, SOX10 transcription factor | Automated |
| GAP43 | Bio-Techne Canada, Oakville On, NB300-143 | Rabbit polyclonal | Regenerating neural tissues/growth cones, GAP43 intracellular growth protein/membrane protein | 1/5000 |
| HLA-DR | Dako, Santa Clara CA MO74601 | Mouse monoclonal | APCs and lymphocytes, Alpha-chain of HLA-DR cell surface receptor | Automated |
| CD68 | Dako, Santa Clara CA GA61361-2 | Mouse monoclonal | Macrophage, lysosomal-associated membrane proteins | Automated |
| CD163 | Vector laboratories, Burlingame CA VP-C374 | Mouse monoclonal | Macrophage, hemoglobin-scavenger receptor | Automated |
| CD40 | Abcam, Toronto ON, ab13545 | Rabbit polyclonal | APCs, cell surface innate immune response costimulatory protein | 1/1000 |
| CD11c | Abcam, Toronto ON, ab52632 | Rabbit monoclonal | APCs, cell surface fibrinogen receptor | 1/300 |
| Gene Assay | Vendor | Cat Number | Amplicon Length | Exon-Spanning |
|---|---|---|---|---|
| CD68 | QIAGEN | QT00037184 | 73 bp | Y |
| NESTIN | QIAGEN | QT01015301 | 75 bp | Y |
| SOX2 | QIAGEN | QT00237601 | 64 bp | N |
| SOX9 | QIAGEN | QT00001498 | 111 bp | Y |
| SOX10 | QIAGEN | QT01670326 | 145 bp | Y |
| NF2 | QIAGEN | QT00030191 | 148 bp | Y |
| GFAP | QIAGEN | QT00081151 | 96 bp | Y |
| Beta-Actin | QIAGEN | QT01680476 | 104 bp | Y |
| Beta-TUB3 | QIAGEN | QT00083713 | 78 bp | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogala, B.; Khan, Z.A.; Jackson-Boeters, L.; Darling, M.R. Investigation of the Molecular Profile of Granular Cell Tumours and Schwannomas of the Oral Cavity. Dent. J. 2022, 10, 38. https://doi.org/10.3390/dj10030038
Rogala B, Khan ZA, Jackson-Boeters L, Darling MR. Investigation of the Molecular Profile of Granular Cell Tumours and Schwannomas of the Oral Cavity. Dentistry Journal. 2022; 10(3):38. https://doi.org/10.3390/dj10030038
Chicago/Turabian StyleRogala, Benjamin, Zia Ali Khan, Linda Jackson-Boeters, and Mark Roger Darling. 2022. "Investigation of the Molecular Profile of Granular Cell Tumours and Schwannomas of the Oral Cavity" Dentistry Journal 10, no. 3: 38. https://doi.org/10.3390/dj10030038
APA StyleRogala, B., Khan, Z. A., Jackson-Boeters, L., & Darling, M. R. (2022). Investigation of the Molecular Profile of Granular Cell Tumours and Schwannomas of the Oral Cavity. Dentistry Journal, 10(3), 38. https://doi.org/10.3390/dj10030038