Abstract
The objectives of this study are to investigate the cytotoxic effect of different Citrullus colocynthis extracts on breast and lung cancer cell lines using flow cytometry to gain mechanistic insights. C. colocynthis was extracted sequentially using the Soxhlet method. We first tested the plant extracts’ cytotoxicity on non-malignant L929 cells and cancerous breast (MCF-7) and lung (A549) cell lines. We observed that the IC50 of the methanol extract on the viability of MCF-7 and A549 cell lines was 81.08 µg/mL and 17.84 µg/mL, respectively, using the MTT assay. The aqueous and methanol extracts were less toxic when tested against the non-cancerous L929 cell line, with IC50 values of 235.48 µg/mL and 222.29 µg/mL, respectively. Then, using flow cytometry, we investigated the underlying molecular pathways with Annexin-V, Anti-Bcl-2, Caspase-3, and DNA fragmentation (TUNEL) assays. Flow cytometric and molecular marker analyses revealed that the methanol extract activated caspase-3 and inhibited Bcl-2 protein, causing early and late apoptosis, as well as cell death via DNA damage in breast and lung cancer cells. These findings indicate that the methanol extract of C. colocynthis is cytotoxic to breast and lung cancer cell lines. The total phenolic and flavonoid content analysis results showed the methanolic extract of C. colocynthis has a concentration of 326.25 μg GAE/g dwt and 274.61 μg QE/g dwt, respectively. GC-MS analysis of the methanol extract revealed phytochemicals relevant to its cytotoxicity.
1. Introduction
According to WHO statistical data from 185 countries for the year 2019; it was reported that cancer and cardiovascular disease are among the leading causes of death for people of age 30–70 years [1,2]. It is estimated that approximately 3 out of 10 deaths in the population are due to cancer [1]. According to the recent data from global cancer statistics ‘GLOBOCAN 2020’, it was estimated that around 19.3 million new cancer cases and nearly 9.9 million deaths were reported (excluding non-melanoma skin cancer) [2]. The number of deaths due to cancer is expected to reach about 21.4 million by 2030 [3].
Among the different cancer types, the majority of cases were found to be breast cancer in females, which accounts for about 11.7% of total newly diagnosed cases (i.e., 2,261,419 cases); and is followed by lung cancer, 11.4% (2,206,771). It was reported that lung cancer is one of the cancer types that has a high mortality rate i.e., 18% of cancer cases (approximately one in five deaths) [2].
According to a WHO report in the year 2017, it was recorded that the Kingdom of Saudi Arabia ranked second among all Gulf countries in mortality rate due to cancer [4]. According to the “Saudi Cancer Registry”, 27,885 new cases of cancer and 13,069 deaths from cancer were recorded in 2020. Among all cancer types, the incidence of colorectal cancer (14.4%) is higher in all populations (including men and women) followed by breast cancer (14.25%). However, the incidence of breast cancer in the female population in the Kingdom was found to be higher (about 29%) when compared to other types of cancer [5].
Meanwhile, the incidence of lung cancer in the Kingdom of Saudi Arabia was about 4.1% of newly diagnosed cancer cases in the year 2020. However, the mortality rate of lung cancer in the Kingdom was found to be high; about 1157 new cases and 1001 fatal cases due to lung cancer were registered [5].
Since the ancient period, mankind has used several plant materials for treating many ailments due to their low cost and their effectiveness against various diseases [6]. Hippocrates, the father of modern medicine used plants for treating diseases [7]. The Prophet Mohammed (PBUH) and his companions regularly used plant-based materials for the treatment of many diseases, as is well documented in the book known as “The Prophetic Medicine (Al- Tibb al-Nabawi)” [8,9]. Today, most modern medicines are prepared/designed based on a lead compound obtained from natural resources [10]. For example, atropine, chloroquine, quinidine, etc. However, some drugs, such as vincristine and vinblastine, are, to this day, only obtained from the Vinca plant [7]. Hence, due to the growing number of cancer cases and a number of serious side effects associated with the chemotherapy and radiotherapy of cancer, the current world’s focus is shifted toward natural resources [9,11].
The kingdom of Saudi Arabia is very well known for its bio-diversity in natural flora, due to its diversity in environmental and geographical conditions [12].
Several plant species having medicinal value were identified in different regions of the country [6,12,13].
Citrullus colocynthis L. (F. Curcurbitaceae) is a perennial herbaceous vine commonly grown in desert regions, referred as Colocynth/Bitter-apple/Bitter-gourd (in English), Hanzal (in Arabic and Urdu), and Indrayan (in Hindi) [14].
It is a widely grown medicinal plant, commonly available in Asia, North Africa, and also in Europe [15], and it is indigenously grown in countries such as Saudi Arabia, Syria, Jordan Lebanon, Kuwait, Turkey, Iran, India, Pakistan, Afghanistan, Cyprus, Egypt, North African countries, and also in Mediterranean Europe [16].
It is traditionally used for its medicinal value against various diseases, such as diabetes [17,18], cancer, gastrointestinal disorders such as indigestion, constipation, gastro-enteritis, dysentery, colic pain; cardiovascular disorders, such as hypertension and hypercholesterolemia; as an analgesic and anti-inflammatory agent [19] against various musculoskeletal disorders, such as joint pain and rheumatism; various pulmonary diseases, such as asthma and bronchitis; against various infectious diseases [20] caused by bacteria, parasites (plasmodium), fungi [21], mycobacteria, mastitis, and gynecological infections, jaundice, dermatological conditions, and polycystic ovarian syndrome [22].
It has been shown that roots, stems, leaves, and fruit have several active principals, such as triterpene, alkaloids, glycosides, flavonoids, essential oils, and fatty acids. Cucurbitacins, which are a tetracyclic triterpene, are a major active constituent present in fruit. Different types of cucurbitacins were found in fruit, including A, B, C, D, E, I, J, K, and L [14]. However, the major types reported are cucurbitacin E and I from the pulp of the fruit. Whereas, cucurbitacin B and I were found in the rind of the fruit [21]. Some of this triterpene was found to exist in glycoside form i.e., colocynthosides A and colocynthosides B [23]. In another study, the major active constituent obtained from fruit was reported to be 2-O-β-D-glucopyranosyl cucurbitacin E [24]. Other constituents found in fruit were khekadaengoside E, cucurbitacin L 2-O-β-D glucopyranoside, cucurbitacin J 2-O-β-D-glucopyranoside, cucurbitacin I 2-O-β-D-glucopyranoside, cucurbitacin K 2-O-β-D glucopyranoside, and cucurbitacin I 2-O-β-D glucopyranoside [23].
Flavonoid and polyphenol content were also reported from the fruit of C. colocynthis; however, the percentage of total flavonoid and polyphenol content from fruits was low when compared to leaves and roots [25].
The main active principle found in C. colocynthis is cucurbitacin, and there were nearly 20 different types of cucurbitacins isolated from fruit [26]. It is very well known that cucurbitacins have very strong anticancer activity, as well as a synergistic effect with many anticancer drugs [26,27,28,29,30]. Cucurbitacin glucosides B and E isolated from leaves of C. colocynthis have been reported to have anticancer activity against breast cancer cell-lines MCF-7 [28]. In our study, the title plant, C. colocynthis, was selected for its well-known antitumor capabilities. However, there is scarcity of data on the molecular pathways that underpin its anticancer properties. As a result, our primary objective was to reevaluate the mechanistic pathways of the indigenously cultivated C. colocynthis fruit extracts on MCF-7 and A549 cell lines utilizing Annexin-V, Caspase-3, anti-Bcl-2, and DNA degradation by TUNEL assay.
Several research papers have reported the activity of C. colocynthis in various lung cancer cell-lines [31,32,33,34,35,36,37]. The cytotoxic activity of medicinal plants can be attributed to the presence of phytochemicals, such as polyphenols, flavonoids, terpenes, and alkaloids [38,39,40,41,42,43].
The main aim of this study was to perform a cytotoxicity evaluation of the different extracts of the whole fruit of C. colocynthis, followed by a mechanistic study of its cytotoxic activity on breast cancer (MCF-7) and lung cancer (A549) cell lines. Metabolite profiling of the non-polar extract and quantification of the polar extract’s total phenolic content were also accomplished.
2. Materials and Methods
2.1. Collection and Crude Extraction
C. colocynthis whole fruits were collected from the Najran University Campus in Saudi Arabia and identified by an expert pharmacognosist, Prof. Mohammed A. A. Orabi, Department of Pharmacognosy, College of Pharmacy, Najran University, Najran, Saudi Arabia, and a herbarium specimen (CC-102021) was deposited in the Department of Pharmacognosy. After being washed, the collected plant was sliced thinly and dried in the shade, then powdered. For the Soxhlet extraction, about 100 g of dried C. colocynthis powder was used. Hexane, ethyl acetate, methanol, and distilled water were used to perform the solvent extraction process in order of increasing polarity. The extracted plant material was always dried before being extracted using subsequent solvents. By using a rotary vacuum evaporator to concentrate the extracts, they were all evaporated to dryness and stored for further use.
2.2. In-Vitro Cytotoxicity of C. colocynthis Extracts
MTT assay was used to evaluate the cytotoxicity of various C. colocynthis solvent extracts, as previously mentioned [41]. In summary, in 96-well plates, non-cancerous (L929) and cancerous (MCF-7 and A549) cell lines were initially plated at a density of 1 × 105 cells per mL. Cells were treated with a range of concentrations (50, 100, 150, 200, and 250 µg/mL) of various fractions and then incubated for 24 h at 37 °C. Each well received 20 μL of the MTT solution (5 mg/mL) before being further incubated at 37 °C for 4 h. At 570 nm, an ELISA reader (Bio-Tek Instruments, Winooski, VT, USA) was used to read the optical density. Each concentration was assayed in triplicate. In order to determine the IC50 values, a linear regression curve was plotted.
Inhibition Percentage = OD of rest sample/OD of control × 100
2.3. Flow Cytometry Based Mechanistic Study
2.3.1. Detection of Early and Late Apoptosis through Binding of Annexin-V to Phosphatidylserine
Flow cytometry was used to investigate the detection of early and late apoptosis by binding of Annexin V to phosphatidylserine (PS). The overall cell apoptosis detection procedures were similar to those described earlier [44]. The cells were seeded in a 6-well flat bottom micro plate with cover slips and incubated overnight at 37 °C in a CO2 incubator. MCF-7 and A549 cancer cell lines were treated for 24 h with the IC50 concentration of methanol extract. After incubation, cells were washed twice with PBS, centrifuged at 500× g for 5 min at 4 °C, and the supernatant was removed. The cell pellets were resuspended (1 × 106 per mL) in binding buffer, then gently mixed in 5 µL of AbFlour 488 Annexin V and 2 µL PI. Tubes were incubated on ice for 15 min in the dark, and finally, gently mixed in 400 µL of ice-cold 1× binding buffer. Flow cytometry was used to analyze cell preparations within 30 min. The data was then analyzed using the FlowJoX 10.0.7 software (2).
2.3.2. Caspase Activation Assay
Following the manufacturer’s guidelines, caspase-3 activity was measured using a commercially available kit (TACS kit). Briefly, MCF-7 and A549 Cancer cell lines were treated with a methanol extract of C. colocynthis for 24 h after being pre-treated with the caspase-3 inhibitor Z-DEVD-FMK (20 M, BD Biosciences) for 2 h. A flow cytometer was used to measure the levels of caspase 3 activity (Acquisition: 7. Cytomics FC500 Flow cytometer, Beckman Coulter, CA, USA).
2.3.3. Anti Bcl-2 Activity
According to the method outlined by Moraes et al., 2013 [45], we used flow cytometry to determine the amount of Bcl-2 protein present in cancer cell lines MCF-7 and A549 that had been treated with methanolic extract of C. colocynthis. Following treatment with the test specimen at its IC50 concentrations, the cell preparations were analyzed flow cytometrically using FlowJo X 10.0.7 software within thirty minutes of the end of the treatment.
2.3.4. DNA Fragmentation Analysis
The effects of C. colocynthis methanol extract on DNA damage in breast cancer MCF-7 and lung cancer A549 cells were studied using a TUNEL assay kit (The APO-DIRECTTM Kit) and flow cytometry (Cytomics FC500 Flow cytometer, Beckman Coulter, CA, USA). Test sample cells were fixed in 2% formaldehyde on ice for 15 min, then washed in PBS. The cells were then fixed in ice-cold 70% ethanol for 30 min, washed once with PBS, and incubated in 50 μL of terminal deoxynucleotidyl transferase (TdT) reaction solution at 37 °C for 1 h. After the reaction, the cells were stained with 2.5 μg mL−1 FITC-avidin, 0.1% Triton X-100, and 5% dried low-fat milk in 4 SSC (0.6 M sodium chloride, 60 mM sodium citrate, pH 7) for 1 h. Cells were counterstained with Propidium iodide and RNase A in PBS. Cytomics FC500 Flowcytometry, Beckman Coulter, USA, was used to analyze the FITC signal [46].
2.4. Phytochemical Analysis
In the current investigation, an initial phytochemical analysis of crude solvent extracts of C. colocynthis was carried out as follows:
2.4.1. Estimation of Total Phenols and Flavonoid Content
The total amount of phenolic compounds in the methanolic extract was measured by using spectrophotometer, as previously described [39]. Gallic acid (GA) calibration curves were developed for concentrations between 20 and 100 μg/mL. Lastly, the phenolic concentrations were normalized to gallic acid equivalents (GAE): mg GAE/g of dry weight (dw) of the extract. From the calibration curve (y = 0.001x + 0.113), we were able to determine the total phenolic content.
The total flavonoid content was also calculated using a colorimetric assay with the aluminum chloride method [40]. Standard curves for quercetin concentrations between 20 and 100 µg/mL were created. In order to compare the quercetin concentrations in methanol, 1 mL of each quercetin concentration was added to a 10 m: volumetric flask containing 4 mL of double-distilled water. Sodium nitrite (0.3 mL, 5%), aluminum chloride (0.3 mL, 10%), and sodium hydroxide (2 mL, 1 M) were added at t = 0. After quick stirring in 2.4 mL of double-distilled water, the mixture was ready for use. The absorbance of the pink color mixture was measured at 510 nm, with a quercetin-free blank serving as a reference. The typical quercetin absorbance values were used to plot the calibration curve. Extract flavonoid concentrations were expressed as quercetin equivalents (Y = 0.265 × 0.152), which are expressed as mg QE/g dry weight.
2.4.2. Gas Chromatography-Mass Spectrometry (GC-MS) Profiling
The method of GC-MS analysis of different solvent extracts of C. colocynthis was performed as previously described [42]. GC-MS model GCMS-QP2010S with an ELITE-5MS column (30 m × 0.25 mm, i.d.) was used in the analysis that employs fused silica column and the components were separated using helium as a carrier gas at a constant flow of 1 mL/min. The 1 μL methanol extract of C. colocynthis was injected into the instrument through split injector. The initial temperature of column was set at 80 °C, whereas the injector temperature was set at 260 °C, and throughout the process, temperature flow was set at the speed of increasing 10 °C/min, with specifications of Linear Velocity, Pressure: 65.0 kPa, Total Flow: 24.0 mL/min, Column Flow: 1.00 mL/min, Linear Velocity: 36.8 cm/sec, Purge Flow: 3.0 mL/min and final temperature was adjusted to 280 °C and run for 6 min. Identification of the major components of plant extracts were confirmed by comparison with their relative retention time and mass spectra with the standard compounds.
2.5. Statistical Analysis
Using GraphPad Prism 6, a one-way analysis of variance (ANOVA) and Dunnett’s test were performed to determine statistical significance at a p value of less than 0.05. Three independent tests (n = 3) were used to calculate the mean and standard deviation.
3. Results
3.1. Cytotoxicity Study of C. colocynthis Extracts
As the present study focused on the anticancer applications of phytochemicals from C. colocynthis, the different solvent extracts were subjected to cytotoxicity study on normal fibroblast cell line L929 using MTT cell viability assay. In all treated extracts, there was dose-dependent activity, that is, with an increase of the concentrations of extracts, the percentage of cell viability was decreased. Almost all tested extracts showed a toxic nature at higher concentrations. The results are shown in Table 1. Whereas the morphological effects of each solvent extract are shown in Figure S1 (Supplementary file).

Table 1.
Cytotoxicity of different solvent extracts of C. colocynthis against L929 cell line.
The cytotoxicity studies of the breast cancer MCF-7 and lung cancer A549 cell lines using MTT viability assay, along with cisplatin as a standard drug, showed significant activity by suppressing the cells’ growth through a decrease of the cell viability in both cell lines. Similar to the toxicity assay, increasing the concentration of extracts reduced the cell viability significantly. At the initial concentration of hexane, ethyl acetate, methanol, and aqueous extracts, the cell viability in lung cancer was observed to be 46.99 ± 0.002, 37.81 ± 0.022, 43.57 ± 0.003, and 49.39 ± 0.012, (Table 2 and Figure S2), respectively, and in the case of breast cancer, it was found to be 61.36 ± 0.005, 65.21 ± 0.020, 55.00 ± 0.022, and 66.99 ± 0.005 (Table 3 and Figure S3). In comparison, the selected solvent extracts were more effective against lung cancer than against breast cancer. Based on overall cell viability and preliminary observations, it was evident that the methanol extract was the most effective against both the breast cancer MCF-7 and lung cancer A549 cell lines of all tested extracts.
Table 2.
Anticancer activity of different solvent extracts of C. colocynthis against the lung cancer A549 cell line.
Table 3.
Anticancer activity of different solvent extracts of C. colocynthis against the breast cancer MCF-7 cell line.
Further, the standard calibration curve was used to determine the IC50 value. For the A549 cell line, the hexane, ethyl acetate, methanol, and aqueous extracts showed an IC50 of 36.80, 39.48, 17.84 and 56.44 µg/mL, respectively. In the case of breast cancer MCF-7, it was found to be 148.01, 108.15, 81.08, and 150.43 µg/mL, respectively. The IC50 values are shown in Table 4.
Table 4.
IC50 value (µg/mL) of solvent extracts of C. colocynthis against different cell lines.
The MTT findings are consistent with the morphological findings, as well. Cellular morphology was seen to be altered in test samples from the treated MCF-7 and A549 cell lines when compared to untreated cells (Figures S2 and S3). There was a greater number of cells in the untreated group and their morphology was more distinct. When the concentration was increased, however, the number of cells in the test samples began to decrease, and intracellular gaps were observed. Cell shrinkage, apoptotic bodies, membrane blabbing, cell turgidity, etc. were seen, among other distinct morphological traits with abnormalities. All of these characteristics were typical of cells in the apoptotic process. Examination under a microscope and with an MTT assay suggest that the test samples may induce apoptosis in the MCF-7 and A549 cell lines.
3.2. Flow Cytometry-Based Mechanistic Pathway Studies
3.2.1. Detection of Apoptosis Based on Annexin-V Assay
In the present study, both treated breast cancer MCF-7 and lung cancer A549 cells were subjected for the detection of early apoptosis and late apoptosis. The results showed that in the case of lung cancer, A549 untreated cells reached around 0.129% of apoptosis, whereas standard drug cisplatin showed 41.7% of apoptotic cells, and test sample methanol extract showed 42.20% of apoptotic cells. Similarly, the breast cancer MCF-7 untreated group showed around 0.167% apoptotic cells, while the standard drug was about 33% of apoptosis cells and methanol extract showed 20.77% apoptotic cells (Figure 1 and Figure 2).
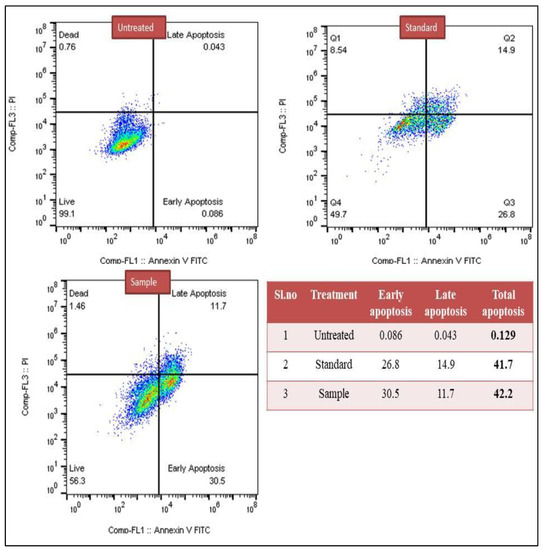
Figure 1.
Detection of early and late apoptosis induced by methanol extract of C. colocynthis in the lung cancer A549 cell line.
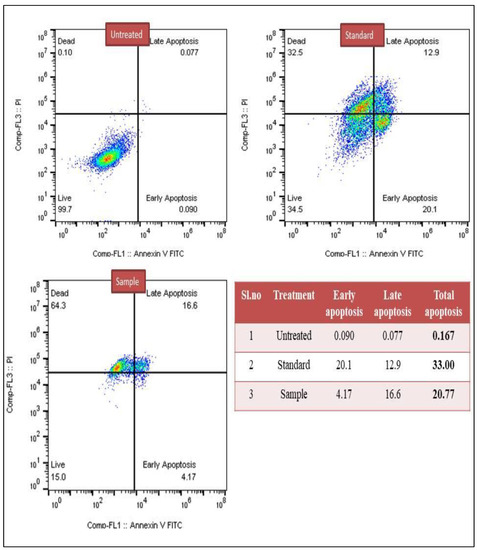
Figure 2.
Detection of early and late apoptosis induced by methanol extract of C. colocynthis in the breast cancer MCF-7 cell line.
3.2.2. Caspase-3 Marker Assay
The caspase-3 assay findings revealed that there was evidence for the induction of apoptosis in both cancer cell lines when treated with the the methanol extract of C. colocynthis. The data shows that in the case of lung cancer A549 cells, the positive group cisplatin drug showed 16.2% of cells positive for caspase-3, and for methanol extract, it was observed to be 8.39%. Further, in the case of breast cancer MCF-7, the standard drug showed around 51.3% of positive cells for caspase-3 and methanol extract showed 13.8%. Among the untreated lung cancer and breast cancer cells, it was found to be 4.94% and 6.24%, respectively. The results are shown in Figure 3 and Figure 4.

Figure 3.
Confirmation of apoptosis induced by methanol extract of C. colocynthis in the lung cancer A549 cell line through caspase-3 assay.
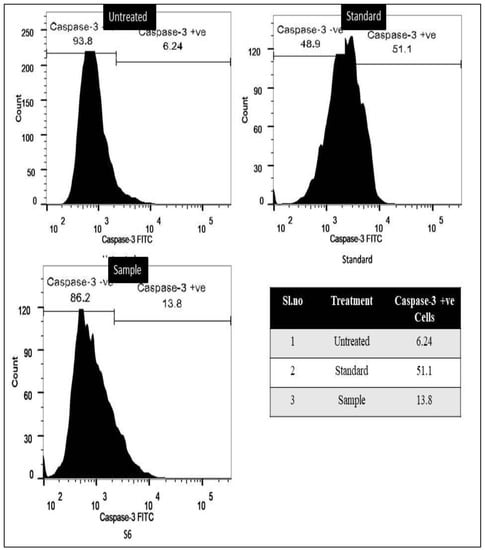
Figure 4.
Confirmation of apoptosis induced by methanol extract of C. colocynthis in the breast cancer MCF-7 cell line through caspase-3 assay.
3.2.3. Bcl-2 Marker Assay
The results indicate that for untreated cells, there was reduced apoptosis, with a high level of Bcl-2 positive cells; 92.5% for A549 and 98.5% for MCF-7. In both the test sample and the standard medication cisplatin treated cells, there was a decrease of the quantity of Bcl-2 positive cells. In the A549 lung cancer test sample, low Bcl-2 positive cells were 21.5% and high Bcl-2 positive cells were 78.5%. For the standard drug, 45.3% of cells showed low levels of Bcl-2, whereas 54.7% of cells showed high levels of Bcl-2. Similarly, in the case of MCF-7, low Bcl-2 positive cells were 46.3% and high Bcl-2 positive cells were 53.7% for the test drug. However, significant variations in the Bcl-2 level were detected in the standard medication cisplatin treated group of cells, i.e., for breast cancer MCF-7, low Bcl-2 was seen in 51.5% of cells and high level Bcl-2 was seen in 48.5% of cells. (Figure 5 and Figure 6).
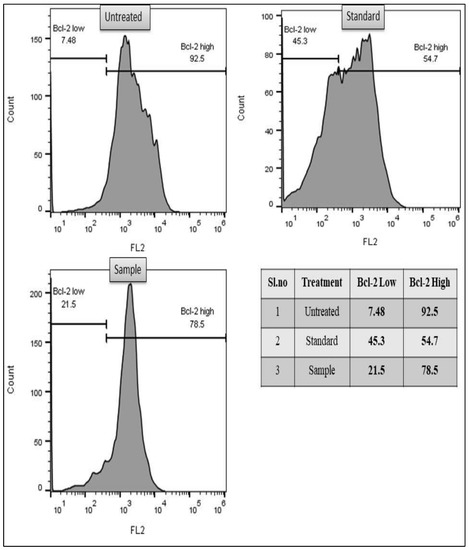
Figure 5.
Flow cytometry-based study of inhibition of anti-apoptotic protein BCL-2 through methanol extract of C. colocynthis in the lung cancer A549 cell line.

Figure 6.
Flow cytometry-based study of inhibition of anti-apoptotic protein BCL-2 through methanol extract of C. colocynthis in the breast cancer MCF-7 cell line.
3.2.4. TUNEL Assay for DNA Damage
Flow cytometry TUNEL assay results showed that 8.26% and 3.87% of cells in the untreated groups of breast cancer MCF-7 and lung cancer A549 exhibited DNA damage, respectively. This could be because just a small percentage of cells died by apoptosis. DNA damage was observed in approximately 53.2% and 56.00% of cells in the test sample treated for breast cancer MCF-7 and lung cancer A549. In comparison, DNA damage was observed in approximately 85.7% and 83.9% of cells in the sample group treated with the gold standard medication cisplatin. (Figure 7 and Figure 8).
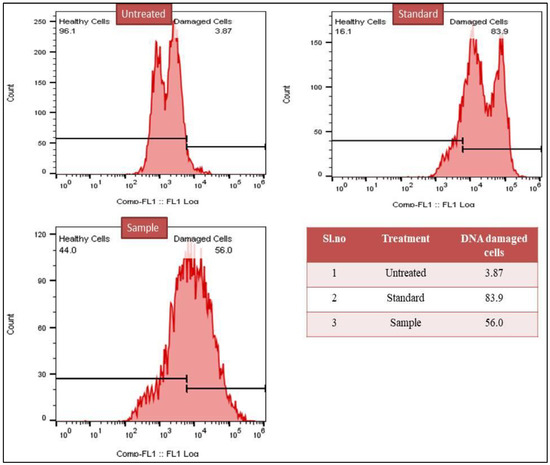
Figure 7.
Flow cytometry-based study of DNA damage induced by methanol extract of C. colocynthis in the lung cancer A549 cell line.

Figure 8.
Flow cytometry-based study of DNA damage induced by methanol extract of C. colocynthis in breast cancer MCF-7 cell line.
3.3. Total Phenolic and Total Flavonoid Contents
The various solvent extracts of C. colocynthis were examined for the qualitative presence of various secondary metabolites. The results revealed that the hexane extract demonstrated the presence of steroids, the ethyl acetate extracts demonstrated the presence of saponins and steroids, the methanol extract demonstrated the presence of alkaloids, flavonoids, glycosides, phenols, and terpenoids as major phytochemical constituents, and the aqueous extract demonstrated the presence of flavonoids, phenols, and terpenoids.
Based on phytochemical results, the methanol and aqueous extracts of C. colocynthis were further subjected for quantification to evaluate total phenol and flavonoid content. The quantification results showed that the methanol extract exhibited the highest phenolic content i.e., 326.25 mg GAE/g dwt, whereas the aqueous extract exhibited 230.14 mg GAE/g dwt. The total phenolic content was calculated from a standard calibration curve (Y = 0.001X + 0.113) (Figure S4). Similarly, for flavonoids, the methanolic extract showed 274.61 mg QE/g dwt, and the aqueous extract exhibited 154.42 mg QE/g dwt.
3.4. Chromatography and Spectrometry GC–MS Analysis
The hexane extract of C. colocynthis was subjected to GC-MS analysis to identify the phytochemical compounds. The GC-MS results show the presence of 7 sharp peaks in a gas chromatogram (Figure S5). Each peak was subjected to a similarity search by using the NIST library database to identify the compounds listed in Table 5. These compounds are known to have several applications, such as pharmaceutical, industrial applications, organic synthesis, and biological activities (Table 5).
Table 5.
GC-MS identified compounds present in methanol extract of C. colocynthis.
4. Discussion
Cancer is the most common disease worldwide, affecting several organs in the human body and having a high mortality rate [60]. There is strong evidence that antioxidant supplementation during cancer chemotherapy may help to minimize toxicity that is dose-limiting. Plant derived phytochemicals have been found to exhibit a wide range of anticancer actions, including modulating the activity of enzymes that scavenge reactive oxygen species (ROS), participating in cell cycle arrest, inducing apoptosis and autophagy, and reducing the proliferation and invasiveness of cancer cells [61,62,63,64,65].
The current study’s objectives were to investigate the cytotoxic potential of whole fruit extracts of C. colocynthis in the MCF-7 and A549 cell lines, and to investigate the potential mechanistic pathways for its cytotoxic action.
The MTT assay results in the present study revealed that the methanol extract of C. colocynthis has shown the most promise in significantly (p < 0.001) inhibiting the proliferation of breast and lung cancer cell lines, with an IC50 value that is significantly lower compared to other solvent extracts. In addition, noncancerous cells demonstrated decreased toxicity, thereby reducing the possible toxicity to normal cells. Microscopic examination revealed morphological abnormalities such as cell shrinkage, apoptotic bodies, membrane blabbing, and cell turgidity, which are indicative of apoptotic cells. The results of the MTT assay and the microscopic examination showed that both the MCF-7 and the A549 cell lines underwent apoptosis. This indicates that the test samples were successful in their ability to induce apoptosis. In addition, flow cytometry was utilized in order to determine and confirm the mechanistic pathways for its cytotoxic action.
The characteristics of cancers, such as unchecked proliferation, angiogenesis, and ability to elude apoptosis [66,67], are shared by all cancer cells, regardless of their origin or subtype. One of apoptosis’ primary roles is cancer prevention [68]. Although many methods exist to prevent apoptosis in cancer, the intrinsic route is the most common target. By preventing cancer cells from undergoing apoptosis, tumors are able to grow and spread more aggressively, increase angiogenesis, manifest uncontrolled cell proliferation, and hinder differentiation. Gaining control over, or perhaps arresting, the unchecked proliferation of cancer cells is one strategy employed in cancer treatment. A successful tool is to employ the cells’ own programmed cell death. Further, apoptosis targeting is the most effective non-surgical treatment. Apoptosis evasion is a universal feature of cancer, regardless of its origin or subtype; therefore, therapies that target this phenomenon are appropriate to all cancers. Different points in the intrinsic and the extrinsic pathways are the focus of numerous anticancer medicines [69,70,71]. Stimulating proapoptotic molecules and blocking antiapoptotic molecules are two frequent therapeutic targeted techniques [72]. Inhibitors for BCL-2 [73], ligands for death-receptors [68], XIAP inhibition [68], and alkyl phospholipid analogs (APL), which operate as apoptotic signals [70] are only a few of the targets that have been examined. Treatment can be directed at any stage along the pathways; however, it is unclear which point is the most efficient. As more drugs that induce apoptosis in cancer cells are discovered, their most efficient targets will become evident.
In the current study, early and late apoptosis were detected in breast cancer MCF-7 and lung cancer A549 cells, after treatment with C. colocynthis methanol extract. Untreated lung cancer A549 cells showed 0.129% apoptosis, while cisplatin showed 41.7%, and methanol extract showed 42.2%. In breast cancer MCF-7, untreated cells showed 0.167% apoptosis, cisplatin 33%, and methanol extract 20.7%. Thus, the findings of the flow cytometry imply that the phytochemicals found in C. colocynthis work against cancer cells by inducing and facilitating apoptosis.
These findings are consistent with earlier studies which reported that phytochemicals like methyl palmitate (which was one of the active principles of C. colocynthis detected in GC-MS in this study) enhanced the anti-cancer effects of Sorafenib therapy by raising cytotoxicity, triggering apoptosis, and preventing Hep-G2 cell migration [49]. Another study concluded that luteolin promoted apoptosis and autophagy in ANA-1 cells most likely by influencing the p38, JNK, and Akt pathways, decreasing Bcl-2 and Beclin-1 activity, and upregulating caspase 8 and caspase 3 expression [74]. Similarly, kaempferol induced apoptosis in colorectal cancer cells via ROS and p53 mediated activation of p38 and caspases [75].
Caspases are key mediators of apoptosis, their activation is tightly regulated by a phylogenetically conserved death program, which is essential for higher organisms’ growth and development. Caspases are proteases that are normally found in healthy cells as inactive zymogens but undergo autolytic cleavage upon stimulation to become fully active. Caspase-3 is primarily involved in proteolysis during apoptosis, and the presence of cleaved caspase-3 is regarded as a credible marker of apoptotic or dying cells [76]. Understanding the function of caspases and the mechanisms by which they can be regulated is crucial to the development of new therapeutics for cancer and inflammatory disorders, two of the most lethal diseases afflicting humans [77]. In the current study, induced apoptosis was confirmed using a highly precise and targeted caspase-3 marker assay. Caspase-3 assay results showed C. colocynthis methanolic extract induced apoptosis in both cancer cells. In lung cancer A549 cells, cisplatin showed 16.2% caspase-3 positive cells, while methanol extract showed 8.39%. MCF-7 breast cancer standard drug exhibited 51.3% caspase-3 positive cells, whereas methanol extract showed 13.8%. These data showed that C. colocynthis induced apoptosis through caspase-3-mediated pathways in MCF-7 and A549 cells. Evidence from previous literature indicates that phytochemicals, such as lysicamine, podophyllotoxin, and majoranolide, are responsible for caspase-3-mediated apoptosis. The first orphan medicine to be approved by the FDA, procaspase-activating compound-1 (PAC-1), and its synthetic derivative, WF-208, also demonstrated intriguing caspase-3 mediated anticancer efficacy. To this day, a huge number of compounds have been reported and patented for their caspase-3-mediated cytotoxicity. At the same time, scientists are working to bring new compounds to the market that mediate their anticancer action through the regulation of caspases [78].
Furthermore, apoptosis is controlled by proteins in the Bcl-2 family. They regulate the intrinsic apoptosis pathway, which is a method by which mitochondria help kill cells. Bcl-2 stops the oligomerization of BAX/BAK, which would cause the mitochondria to release several molecules that cause cells to undergo apoptosis. It is also known that Bcl-2 binds to BAX and other pro-apoptotic proteins and inactivates them, thus inhibiting apoptosis [79]. In the current investigation, the level of Bcl-2 protein that was detected in the cells was used as a marker to determine the cytotoxic potential of the test sample in terms of its ability to induce apoptosis in cancer cells. In comparison to untreated, test samples and standard drugs, both demonstrated anti-Bcl-2 protein activity, which promotes apoptosis. These findings corroborate with the results of previous studies, which have demonstrated the anti-Bcl-2 activity of several phytochemicals, for instance, morin [80] and Malvidin [81] showed anticancer activity against Human colorectal cancer cell line (HCT-116). Similarly, Catechin [82], Kuarinone [83], Apigenin [84], Quercetin [85], Acacetin [86], and Genistein [87], to name a few, demonstrated significant cytotoxicity against Human breast cancer cell line (MCF-7) and Human non-small-cell lung cancer cell line (A549).
In the current study, the extent of DNA damage was assessed by the TUNEL assay. The free 3-hydroxyl termini are labelled in the TUNEL assay, which uses terminal deoxynucleotidyl transferase to detect DNA breakage. TUNEL staining is still frequently used as a measure of cell death during apoptosis because genomic DNA breaks appear during the early and late stages of apoptosis [88,89]. The assay results confirmed the DNA damage in cancer cells treated with standard and the test drug. The untreated MCF-7 and A549 cells showed 8.26% and 3.87% DNA damage, which may be attributed to the low apoptosis rate. In test sample-treated MCF-7 and A549, 53.2% and 56.0% of cells showed DNA damage, whereas, in standard drug cisplatin-treated samples, 85.7% and 83.9% of cells showed DNA damage. Thus, the mechanistic study results reaffirm that the cytotoxic potential of C. colocynthis is mediated through cell apoptosis via anti-Bcl-2, pro-caspase-3 activity and DNA damage.
In the current study, qualitative phytochemical investigation of C. colocynthis extracts revealed the presence of several phytochemicals, including flavonoids, alkaloids, phenols, glycosides, and terpenes. The presence of flavonoids and phenols was further confirmed by the quantification results. These findings are consistent with previous studies, which have reported the presence of similar phytochemicals in C. colocynthis [14,21,22,23,24]. Polyphenols and flavonoids’ antioxidant properties are thought to protect against a variety of diseases associated with reactive oxygen species (ROS), including cancer. In the current study, all four extracts of C. colocynthis were evaluated for cytotoxic potential against non-cancerous L929 and cancerous MCF-7 and A549 cell lines. The preliminary investigation revealed the methanol extract to be effective against the tested cancer cell lines, and was further subjected to GCMS analysis, which revealed the presence of phytochemicals with known cytotoxic and antioxidant potentials (Table 5).
Several studies conducted around the world have suggested that phytochemicals can inhibit tumor generation, induce apoptosis in cancer cells, and interfere with tumor progression [62,63]. Oxidative stress has been linked to cancer incidence and carcinogenesis. Reactive oxygen species levels rise, and antioxidant levels fall during the carcinogenesis process in cancer cells. These cells’ ROS levels can rise as a result of intrinsic or extrinsic factors, which can change transcriptional processes, signaling pathways, and induction gene mutations, all of which contribute to the development of cancer [64]. Adjuvant chemotherapy benefits from the use of antioxidants because they neutralize and remove oxidizing free radicals before they can cause cellular damage. Estimates place the percentage of cancer patients taking antioxidant supplements anywhere from 13% to 87%. Additionally, the use of antioxidant supplements during chemotherapy has been shown to improve therapeutic efficacy and prolong patient survival [65]. Thus, C. colocynthis methanolic extract, which is rich in flavonoid and phenolics, as shown from the quantitative assay, may also operate by reducing oxidative stress [66], and thus, has the potential to be used as a beneficial anticancer therapy.
5. Conclusions
The C. colocynthis methanol extract was found to have significant cytotoxic activity against both the MCF-7 breast cancer and A549 lung cancer cell lines, which is comparable to that of the gold standard drug cisplatin. The signaling pathways of the test drug were confirmed to induce apoptosis and DNA damage. Therefore, C. colocynthis is a valuable source of new anticancer therapeutics and should be further investigated to identify lead compounds with cytotoxic potential. Further research (clinical trials) is needed to determine if the study’s findings can be applied to human beings.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/separations9120411/s1, Figure S1. Cytotoxicity of C. colocynthis on L929 cell line. Figure S2. Cytotoxic activity of C. colocynthis on Lung cancer A549 cell line. Figure S3. Cytotoxic activity C. colocynthis on Breast cancer MCF-7 cell line. Figure S4. Calibration curve for standard gallic acid. Figure S5. GC chromatogram and mass spectra of methanol extract of C. colocynthis.
Author Contributions
Conceptualization, A.M.A., I.A.S., M.A.A.O., B.A.A.-W. and M.S.H.; Funding acquisition, A.M.A.; Investigation, I.A.S., M.M.K. and A.K.S.; Methodology, M.A.A.O., B.A.A.-W., M.S.H. and A.K.S.; Project administration, S.A.A. and J.H.H.; Resources, I.A.S. and I.A.W.; Software, M.M.K.; Supervision, A.M.A. and I.A.W.; Visualization, S.A.A.; Writing—original draft, I.A.S., M.S.H. and A.K.S.; Writing—review & editing, A.M.A., S.A.A., M.A.A.O., B.A.A.-W., I.A.W., M.M.K. and J.H.H. All authors have read and agreed to the published version of the manuscript.
Funding
The authors would like to acknowledge the support of the Deputy for Research and Innovation-Ministry of Education, Kingdom of Saudi Arabia, for a grant (NU/IFC/ENT/01/006) under the Institutional Funding Committee at Najran University, Kingdom of Saudi Arabia.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data has been included in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer Treatment and Survivorship Statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, W.S.; Almufareh, N.A.; Domiaty, D.M.; Albasher, G.; Alduwish, M.A.; Alkhalaf, H.; Almuzzaini, B.; Al-Marshidy, S.S.; Alfraihi, R.; Elasbali, A.M.; et al. Epidemiology of Cancer in Saudi Arabia Thru 2010–2019: A Systematic Review with Constrained Meta-Analysis. AIMS Public Health 2020, 7, 679–696. [Google Scholar] [CrossRef]
- Asiri, S.; Asiri, A.; Ulahannan, S.; Alanazi, M.; Humran, A.; Hummadi, A. Incidence Rates of Breast Cancer by Age and Tumor Characteristics among Saudi Women: Recent Trends. Cureus 2020, 12, e6664. [Google Scholar] [CrossRef]
- Yonbawi, A.R.; Abdallah, H.M.; Alkhilaiwi, F.A.; Koshak, A.E.; Heard, C.M. Anti-Proliferative, Cytotoxic and Antioxidant Properties of the Methanolic Extracts of Five Saudi Arabian Flora with Folkloric Medicinal Use: Aizoon Canariense, Citrullus Colocynthis, Maerua Crassifolia, Rhazya Stricta and Tribulus Macropterus. Plants 2021, 10, 2073. [Google Scholar] [CrossRef]
- Innocent, E. Trends and Challenges toward Integration of Traditional Medicine in Formal Health-Care System: Historical Perspectives and Appraisal of Education Curricula in Sub-Sahara Africa. J. Intercult. Ethnopharmacol. 2016, 5, 312–316. [Google Scholar] [CrossRef]
- Ahmad, R.; Ahmad, N.; Abbas, A.; Shehzad, A. Journal of Traditional and Complementary Medicine Role of Traditional Islamic and Arabic Plants in Cancer Therapy. J. Tradit. Chin. Med. Sci. 2017, 7, 195–204. [Google Scholar]
- Al-Jauziyah, Q. Healing with the Medicine of the Prophet, 2nd ed.; Manderola, R.J., Ed.; Dar-us-Salam for Publishing & Distribution: Riyadh, Saudi Arabia, 2003; ISBN 9789960892917. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Zaid, H.; Rayan, A.; Said, O.; Saad, B. Cancer Treatment by Greco-Arab and Islamic Herbal Medicine. Open Nutraceuticals J. 2010, 3, 203–212. [Google Scholar] [CrossRef][Green Version]
- Yusuf, M.; Al-Oqail, M.M.; Al-Sheddr, E.S.; Al-Rehaily, A.J.; Rahman, M.A. Diversity of Medicinal Plants in the Flora of Saudi Arabia 3: An Inventory of 15 Plant Families and Their Conservation Management. Int. J. Environ. 2014, 3, 312–320. [Google Scholar] [CrossRef]
- Al-Namazi, A.A.; Algarni, S.M.; Wan, J.S.H.; Al Mosallam, M.S.; Alotaibi, F. Floristic Composition of Jandaf Mountain as Biodiversity Hotspot Area in Southwestern Saudi Arabia. Saudi J. Biol. Sci. 2022, 29, 3654–3660. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.I.; Rathore, H.A. Abdur-Sattar, “Citrullus Colocynthis (L.) Schrad (Bitter Apple Fruit): A Review of Its Phytochemistry, Pharmacology, Traditional Uses and Nutritional Potential. J. Ethnopharmacol. 2014, 155, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Eid, M.S.A.; Ghassan, M.A.; Sheikha, A. Evaluation of the Anticancer Activity and Fatty Acids Composition of ‘Handal’ (Citrullus Colocynthis L.) Seed Oil, a Desert Plant from South Jordan. Food Sci. Nutr. 2020, 9, 282–289. [Google Scholar]
- Mariod, A.A.; Saeed Mirghani, M.E.; Hussein, I. Citrullus Colocynthis Colocynth, Bitter Apple, Bitter Gourd. In Unconventional Oilseeds and Oil Sources; Elsevier: Amsterdam, The Netherlands, 2017; pp. 99–105. ISBN 9780128094358. [Google Scholar]
- Karimabad, M.N.; Niknia, S.; Golnabadi, M.B.; Poor, S.F.; Hajizadeh, M.R.; Mahmoodi, M. Effect of Citrullus Colocynthis Extract on Glycated Hemoglobin Formation (in Vitro). Eurasian J. Med. 2020, 52, 47–51. [Google Scholar] [CrossRef]
- Shi, C.; Karim, S.; Wang, C.; Zhao, M.; Murtaza, G. A Review on Antidiabetic Activity of Citrullus Colocynthis Schrad. Acta Pol. Pharm. 2014, 71, 363–367. [Google Scholar]
- Pashmforosh, M.; Rajabi Vardanjani, H.; Rajabi Vardanjani, H.; Pashmforosh, M.; Khodayar, M.J. Topical Anti-Inflammatory and Analgesic Activities of Citrullus Colocynthis Extract Cream in Rats. Medicina 2018, 54, 51. [Google Scholar] [CrossRef]
- Hameed, B.; Ali, Q.; Hafeez, M.M.; Malik, A. Antibacterial and Antifungal Activity of Fruit, Seed and Root Extracts of Citrullus Colocynthis Plant. Biol. Clin. Sci. Res. J. 2020, 2020, 1–5. [Google Scholar] [CrossRef]
- Bourhia, M.; Bouothmany, K.; Bakrim, H.; Hadrach, S.; Salamatullah, A.M.; Alzahrani, A.; Khalil Alyahya, H.; Albadr, N.A.; Gmouh, S.; Laglaoui, A.; et al. Chemical Profiling, Antioxidant, Antiproliferative, and Antibacterial Potentials of Chemically Characterized Extract of Citrullus Colocynthis L. Seeds. Separations 2021, 8, 114. [Google Scholar] [CrossRef]
- Barzegar, M.H.; Khazali, H.; Kalantar, S.M.; Khoradmehr, A. Effect of Citrullus Colocynthis Hydro-Alcoholic Extract on Hormonal and Folliculogenesis Process in Estradiol Valerate-Induced PCOs Rats Model: An Experimental Study. Int. J. Reprod. Biomed. 2017, 15, 661–668. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Morikawa, T.; Kobayashi, H.; Nakamura, A.; Matsuhira, K.; Nakamura, S.; Matsuda, H. Bioactive Saponins and Glycosides. XXVII. Structures of New Cucurbitane-Type Triterpene Glycosides and Antiallergic Constituents from Citrullus Colocynthis. Chem. Pharm. Bull. 2007, 55, 428–434. [Google Scholar] [CrossRef]
- Torkey, H.M.; Abou-Yousef, H.M.; Azeiz, A.; Farid, H.E.A. Insecticidal Effect of Cucurbitacin E Glycoside Isolated from Citrullus Colocynthis against Aphis Craccivora. Aust. J. Basic Appl. Sci. 2009, 3, 4060–4066. [Google Scholar]
- Hussain, A.I.; Rathore, H.A.; Sattar, M.Z.A.; Chatha, S.A.S.; ud din Ahmad, F.; Ahmad, A.; Johns, E.J. Phenolic Profile and Antioxidant Activity of Various Extracts from Citrullus colocynthis (L.) from the Pakistani Flora. Ind. Crops Prod. 2013, 45, 416–422. [Google Scholar] [CrossRef]
- Chawech, R.; Jarraya, R.; Girardi, C.; Vansteelandt, M.; Marti, G.; Nasri, I.; Racaud-Sultan, C.; Fabre, N. Cucurbitacins from the Leaves of Citrullus colocynthis (L.) Schrad. Molecules 2015, 20, 18001–18015. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Fang, X.; He, C.; Li, P.; Xiao, F.; Wang, Y.; Chen, M. Cucurbitacins: A Systematic Review of the Phytochemistry and Anticancer Activity. Am. J. Chin. Med. 2015, 43, 1331–1350. [Google Scholar] [CrossRef]
- Tannin-Spitz, T.; Grossman, S.; Dovrat, S.; Gottlieb, H.E.; Bergman, M. Growth Inhibitory Activity of Cucurbitacin Glucosides Isolated from Citrullus Colocynthis on Human Breast Cancer Cells. Biochem. Pharmacol. 2007, 73, 56–67. [Google Scholar] [CrossRef]
- Hajjar, D.; Kremb, S.; Sioud, S.; Emwas, A.H.; Voolstra, C.R.; Ravasi, T. Anti-Cancer Agents in Saudi Arabian Herbals Revealed by Automated High-Content Imaging. PLoS ONE 2017, 12, e0177316. [Google Scholar] [CrossRef]
- Chowdhury, K.; Sharma, A.; Kumar, S.; Gunjan, G.K.; Nag, A.; Mandal, C.C. Colocynth Extracts Prevent Epithelial to Mesenchymal Transition and Stemness of Breast Cancer Cells. Front. Pharmacol. 2017, 8, 593. [Google Scholar] [CrossRef]
- Zhang, M.; Bian, Z.G.; Zhang, Y.; Wang, J.H.; Kan, L.; Wang, X.; Niu, H.Y.; He, P. Cucurbitacin B Inhibits Proliferation and Induces Apoptosis via STAT3 Pathway Inhibition in A549 Lung Cancer Cells. Mol. Med. Rep. 2014, 10, 2905–2911. [Google Scholar] [CrossRef]
- Khan, N.; Jajeh, F.; Khan, M.I.; Mukhtar, E.; Shabana, S.M.; Mukhtar, H. Sestrin-3 Modulation Is Essential for Therapeutic Efficacy of Cucurbitacin B in Lung Cancer Cells. Carcinogenesis 2016, 38, 184–195. [Google Scholar] [CrossRef]
- Ni, Y.; Wu, S.; Wang, X. Cucurbitacin I Induces pro-Death Autophagy in A549 Cells via the ERK-MTOR-STAT3 Signaling Pathway. J. Cell. Biochem. 2018, 119, 6104–6112. [Google Scholar] [CrossRef]
- Liu, P.; Xiang, Y.; Liu, X.; Zhang, T.; Yang, R.; Chen, S.; Xu, L.; Yu, Q.; Zhao, H.; Zhang, L.; et al. Cucurbitacin B Induces the Lysosomal Degradation of EGFR and Suppresses the CIP2A/PP2A/Akt Signaling Axis in Gefitinib-Resistant Non-Small Cell Lung Cancer. Molecules 2019, 24, 647. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Khan, S.; Kumar, S. Cucurbitacin B Alters the Expression of Tumor-Related Genes by Epigenetic Modifications in NSCLC and Inhibits NNK-Induced Lung Tumorigenesis. Cancer Prev. Res. 2015, 8, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Tian, B.; Yang, Y.L.; Wang, Y.C.; Liu, S.; Urisman, A.; Yang, C.T.; Xu, Z.; Jablons, D.M.; You, L. Cucurbitacin E Inhibits the Yes associated Protein Signaling Pathway and Suppresses Brain Metastasis of Human Non small Cell Lung Cancer in a Murine Model. Oncol. Rep. 2019, 42, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yan, Q.; Peng, B. Use of Cucurbitacins for Lung Cancer Research and Therapy. Cancer Chemother. Pharmacol. 2021, 88, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gul, R.; Jan, S.U.; Faridullah, S.; Sherani, S.; Jahan, N. Preliminary Phytochemical Screening, Quantitative Analysis of Alkaloids, and Antioxidant Activity of Crude Plant Extracts from Ephedra Intermedia Indigenous to Balochistan. Sci. World J. 2017, 2017, 5873648. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Oxidants and Antioxidants Part A; Elsevier: Amsterdam, The Netherlands, 1999; pp. 152–178. ISBN 9780121822002. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- van Meerloo, J.; Kaspers, G.J.L.; Cloos, J. Cell Sensitivity Assays: The MTT Assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar] [CrossRef]
- Davies, N.W. Gas Chromatographic Retention Indices of Monoterpenes and Sesquiterpenes on Methyl Silicon and Carbowax 20M Phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Irawan, C.; Rochaeni, H.; Sulistiawaty, L.; Roziafanto, A.N. Phytochemical Screening, LC-MS Studies and Antidiabetic Potential of Methanol Extracts of Seed Shells of Archidendron Bubalinum (Jack) I.C. Nielson (Julang Jaling) from Lampung, Indonesia. Pharmacogn. J. 2018, 10, s77–s82. [Google Scholar] [CrossRef]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, db.prot087288. [Google Scholar] [CrossRef] [PubMed]
- Moraes, V.W.R.; Caires, A.C.F.; Paredes-Gamero, E.J.; Rodrigues, T. Organopalladium Compound 7b Targets Mitochondrial Thiols and Induces Caspase-Dependent Apoptosis in Human Myeloid Leukemia Cells. Cell Death Dis. 2013, 4, e658. [Google Scholar] [CrossRef]
- Kyrylkova, K.; Kyryachenko, S.; Leid, M.; Kioussi, C. Detection of Apoptosis by TUNEL Assay. Methods Mol. Biol. 2012, 887, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, M.; Sali, V.K.; Mani, S.; Vasanthi, H.R. Neophytadiene from Turbinaria Ornata Suppresses LPS-Induced Inflammatory Response in RAW 264.7 Macrophages and Sprague Dawley Rats. Inflammation 2020, 43, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Hamed, A.; Mantawy, E.; El-Bakly, W.; Abdel-Mottaleb, Y.; Azab, S. Methyl Palmitate: The Naturally Occurring Cardioprotective Agent. Arch. Pharm. Sci. Ain Shams Univ. 2020, 9, 47–62. [Google Scholar] [CrossRef]
- Breeta, R.D.I.E.; Grace, V.M.B.; Wilson, D.D. Methyl Palmitate-A Suitable Adjuvant for Sorafenib Therapy to Reduce in Vivo Toxicity and to Enhance Anti-Cancer Effects on Hepatocellular Carcinoma Cells. Basic Clin. Pharmacol. Toxicol. 2021, 128, 366–378. [Google Scholar] [CrossRef]
- Ahamed, A.; Panneerselvam, A.; Alaklabi, A.; Arif, I.A.; Ambikapathy, V.; Thajuddin, N. Molecular Perspective and Anticancer Activity of Medicinal Plants. Saudi J. Biol. Sci. 2020, 27, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Lendzion, K.; Gornowicz, A.; Strawa, J.W.; Bielawska, K.; Czarnomysy, R.; Popławska, B.; Bielawski, K.; Tomczyk, M.; Miltyk, W.; Bielawska, A. LC-PDA-MS and GC-MS Analysis of Scorzonera Hispanica Seeds and Their Effects on Human Breast Cancer Cell Lines. Int. J. Mol. Sci. 2022, 23, 11584. [Google Scholar] [CrossRef]
- Atolani, O.; Olatunji, G.A.; Fabiyi, O.A.; Adeniji, A.J.; Ogbole, O.O. Phytochemicals from Kigelia Pinnata Leaves Show Antioxidant and Anticancer Potential on Human Cancer Cell Line. J. Med. Food 2013, 16, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Win, D.T. Oleic Acid-The Anti-Breast Cancer Component in Olive Oil. AU Jt 2005, 9, 75–78. [Google Scholar]
- Rao, A.; Srivastava, S.J.; Kannaujiya, A.K.; Singh, V.K. Structural Information of Natural Product Metabolites in Bryophytes. In Evolutionary Diversity as a Source for Anticancer Molecules; Singh, D., Ed.; Elsevier: San Diego, CA, USA, 2021; pp. 209–231. [Google Scholar]
- Yang, Y.; Fu, C.; Zhou, F.; Luo, X.; Li, J.; Zhao, J.; He, J.; Li, X.; Li, J. Chemical Composition, Antioxidant and Antitumor Activities of Sub-Fractions of Wild and Cultivated Pleurotus Ferulae Ethanol Extracts. PeerJ 2018, 6, e6097. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hady, H.; El-Wakil, E.A.; Abdel-Gawad, M. GC-MS Analysis, Antioxidant and Cytotoxic Activities of Mentha Spicata. Eur. J. Med. Plants 2018, 26, 1–12. [Google Scholar] [CrossRef]
- Nakagawa, M.; Yamaguchi, T.; Fukawa, H.; Ogata, J.; Komiyama, S.; Akiyama, S.; Kuwano, M. Potentiation by Squalene of the Cytotoxicity of Anticancer Agents against Cultured Mammalian Cells and Murine Tumor. Jpn. J. Cancer Res. 1985, 76, 315–320. [Google Scholar]
- Huang, Z.R.; Lin, Y.K.; Fang, J.Y. Biological and Pharmacological Activities of Squalene and Related Compounds: Potential Uses in Cosmetic Dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Ma, X.; Sun, J.; Ding, G.; Cui, Q.; Miao, Y.; Hou, Y.; Jiang, M.; Bai, G. Active Fragments-Guided Drug Discovery and Design of Selective Tropane Alkaloids Using Ultra-High Performance Liquid Chromatography-Quadrupole Time-of-Flight Tandem Mass Spectrometry Coupled with Virtual Calculation and Biological Evaluation. Anal. Bioanal. Chem. 2017, 409, 1145–1157. [Google Scholar] [CrossRef]
- Dai, X.; Wang, X.; Yang, C.; Huang, M.; Zhou, Z.; Qu, Y.; Cui, X.; Liu, R.; Chen, C. Human Fibroblasts Facilitate the Generation of IPSCs-Derived Mammary-like Organoids. Stem Cell Res. Ther. 2022, 13, 377. [Google Scholar] [CrossRef]
- Bharadvaja, N. Medicinal Plants in the Management of Cancer: A Review. Int. J. Complement. Altern. Med. 2017, 9, 00291. [Google Scholar] [CrossRef][Green Version]
- Chiang, E.P.I.; Tsai, S.Y.; Kuo, Y.H.; Pai, M.H.; Chiu, H.L.; Rodriguez, R.L.; Tang, F.Y. Caffeic Acid Derivatives Inhibit the Growth of Colon Cancer: Involvement of the PI3-K/Akt and AMPK Signaling Pathways. PLoS ONE 2014, 9, e99631. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
- Nourazarian, A.R.; Kangari, P.; Salmaninejad, A. Roles of Oxidative Stress in the Development and Progression of Breast Cancer. Asian Pac. J. Cancer Prev. 2014, 15, 4745–4751. [Google Scholar] [CrossRef]
- Block, K.I.; Koch, A.C.; Mead, M.N.; Tothy, P.K.; Newman, R.A.; Gyllenhaal, C. Impact of Antioxidant Supplementation on Chemotherapeutic Toxicity: A Systematic Review of the Evidence from Randomized Controlled Trials. Int. J. Cancer 2008, 123, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kumar, D.; Manjusha; Saroha, K.; Singh, N.; Vashishta, B. Antioxidant and Free Radical Scavenging Potential of Citrullus colocynthis (L.) Schrad. Methanolic Fruit Extract. Acta Pharm. 2008, 58, 215–220. [Google Scholar] [CrossRef]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and Molecular Targeting Therapy in Cancer. Biomed Res. Int. 2014, 2014, 150845. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Tait, S.W.G. Mitochondrial Apoptosis: Killing Cancer Using the Enemy Within. Br. J. Cancer 2015, 112, 957–962. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, X. Endoplasmic Reticulum-Mitochondria Tethering in Neurodegenerative Diseases. Transl. Neurodegener. 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Villa-Pulgarín, J.A.; Gajate, C.; Botet, J.; Jimenez, A.; Justies, N.; Varela-M, R.E.; Cuesta-Marbán, Á.; Müller, I.; Modolell, M.; Revuelta, J.L.; et al. Mitochondria and Lipid Raft-Located FOF1-ATP Synthase as Major Therapeutic Targets in the Antileishmanial and Anticancer Activities of Ether Lipid Edelfosine. PLoS Negl. Trop. Dis. 2017, 11, e0005805. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, Q.; Zhu, Z.; Xu, H.; Ding, F.; Wang, M.; Du, S.; Du, Y.; Yan, Z. BHX, a Novel Pyrazoline Derivative, Inhibits Breast Cancer Cell Invasion by Reversing the Epithelial-Mesenchymal Transition and down-Regulating Wnt/β-Catenin Signalling. Sci. Rep. 2017, 7, 9153. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell Death. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef]
- Zaman, S.; Wang, R.; Gandhi, V. Targeting the Apoptosis Pathway in Hematologic Malignancies. Leuk. Lymphoma 2014, 55, 1980–1992. [Google Scholar] [CrossRef]
- Liao, Y.; Xu, Y.; Cao, M.; Huan, Y.; Zhu, L.; Jiang, Y.; Shen, W.; Zhu, G. Luteolin Induces Apoptosis and Autophagy in Mouse Macrophage ANA-1 Cells via the Bcl-2 Pathway. J. Immunol. Res. 2018, 2018, 4623919. [Google Scholar] [CrossRef]
- Choi, J.B.; Kim, J.H.; Lee, H.; Pak, J.N.; Shim, B.S.; Kim, S.H. Reactive Oxygen Species and P53 Mediated Activation of P38 and Caspases Is Critically Involved in Kaempferol Induced Apoptosis in Colorectal Cancer Cells. J. Agric. Food Chem. 2018, 66, 9960–9967. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Waterhouse, N.J. Detecting Cleaved Caspase-3 in Apoptotic Cells by Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, db.prot087312. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Yadav, P.; Yadav, R.; Jain, S.; Vaidya, A. Caspase-3: A Primary Target for Natural and Synthetic Compounds for Cancer Therapy. Chem. Biol. Drug Des. 2021, 98, 144–165. [Google Scholar] [CrossRef] [PubMed]
- Tzifi, F.; Economopoulou, C.; Gourgiotis, D.; Ardavanis, A.; Papageorgiou, S.; Scorilas, A. The Role of BCL2 Family of Apoptosis Regulator Proteins in Acute and Chronic Leukemias. Adv. Hematol. 2012, 2012, 524308. [Google Scholar] [CrossRef]
- Hyun, H.B.; Lee, W.S.; Go, S.I.; Nagappan, A.; Park, C.; Han, M.H.; Hong, S.H.; Kim, G.; Kim, G.Y.; Cheong, J.; et al. The Flavonoid Morin from Moraceae Induces Apoptosis by Modulation of Bcl-2 Family Members and Fas Receptor in HCT 116 Cells. Int. J. Oncol. 2015, 46, 2670–2678. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, J.; Huang, H.; Liu, L.; Sun, Y. Malvidin Induced Anticancer Activity in Human Colorectal HCT-116 Cancer Cells Involves Apoptosis, G2/M Cell Cycle Arrest and Upregulation of P21WAFI. Int. J. Clin. Exp. Med. 2018, 11, 1734–1741. [Google Scholar]
- Safari, F.; Rabieepor, M.; Jamalomidi, F.; Baghaeifar, Z.; Khodaei, L. Evaluation of Anti-Cancer and pro-Apoptotic Activities of Iranian Green Tea Extract against A549, PC3, and MCF-7 Cancer Cell Lines. Int. J. Basic Sci. Med. 2019, 4, 113–118. [Google Scholar] [CrossRef]
- Yang, J.; Chen, H.; Wang, Q.; Deng, S.; Huang, M.; Ma, X.; Song, P.; Du, J.; Huang, Y.; Wen, Y.; et al. Inhibitory Effect of Kurarinone on Growth of Human Non-Small Cell Lung Cancer: An Experimental Study Both in Vitro and in Vivo Studies. Front. Pharmacol. 2018, 9, 252. [Google Scholar] [CrossRef]
- Lu, H.F.; Chie, Y.J.; Yang, M.S.; Lee, C.S.; Fu, J.J.; Yang, J.S.; Tan, T.W.; Wu, S.H.; Ma, Y.S.; Ip, S.W.; et al. Apigenin Induces Caspase-Dependent Apoptosis in Human Lung Cancer A549 Cells through Bax- and Bcl-2-Triggered Mitochondrial Pathway. Int. J. Oncol. 2010, 36, 1477–1484. [Google Scholar] [CrossRef]
- Sharmila, G.; Bhat, F.A.; Arunkumar, R.; Elumalai, P.; Raja Singh, P.; Senthilkumar, K.; Arunakaran, J. Chemopreventive Effect of Quercetin, a Natural Dietary Flavonoid on Prostate Cancer in in Vivo Model. Clin. Nutr. 2014, 33, 718–726. [Google Scholar] [CrossRef]
- Shim, H.Y.; Park, J.H.; Paik, H.D.; Nah, S.Y.; Kim, D.S.H.L.; Han, Y.S. Acacetin-Induced Apoptosis of Human Breast Cancer MCF-7 Cells Involves Caspase Cascade, Mitochondria-Mediated Death Signaling and SAPK/JNK1/2-c-Jun Activation. Mol. Cells 2007, 24, 95–104. [Google Scholar] [PubMed]
- Ferenc, P.; Solár, P.; Kleban, J.; Mikes, J.; Fedorocko, P. Down-Regulation of Bcl-2 and Akt Induced by Combination of Photoactivated Hypericin and Genistein in Human Breast Cancer Cells. J. Photochem. Photobiol. B 2010, 98, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Murray, D. Do TUNEL and Other Apoptosis Assays Detect Cell Death in Preclinical Studies? Int. J. Mol. Sci. 2020, 21, 9090. [Google Scholar] [CrossRef] [PubMed]
- Alshabi, A.M.; Vastrad, B.; Shaikh, I.A.; Vastrad, C. Exploring the Molecular Mechanism of the Drug-Treated Breast Cancer Based on Gene Expression Microarray. Biomolecules 2019, 9, 282. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).