

Amelioration of Alcoholic Liver Disease by Activating PXR-Cytochrome P450s Axis with Blackberry Extract

 ,
,
Abstract
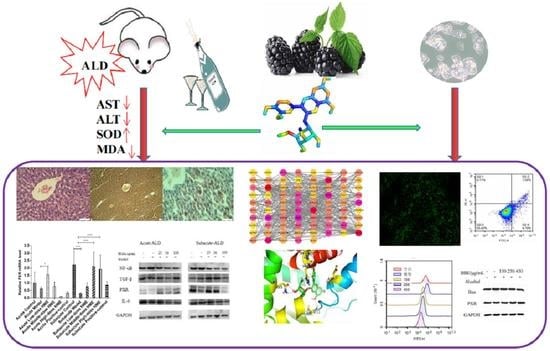
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of the Blackberry Extract
2.3. Polyphenol, Flavonoids, and Anthocyanin Contents
2.4. Antioxidant Activities
2.5. Animals and Treatment
2.5.1. Animals
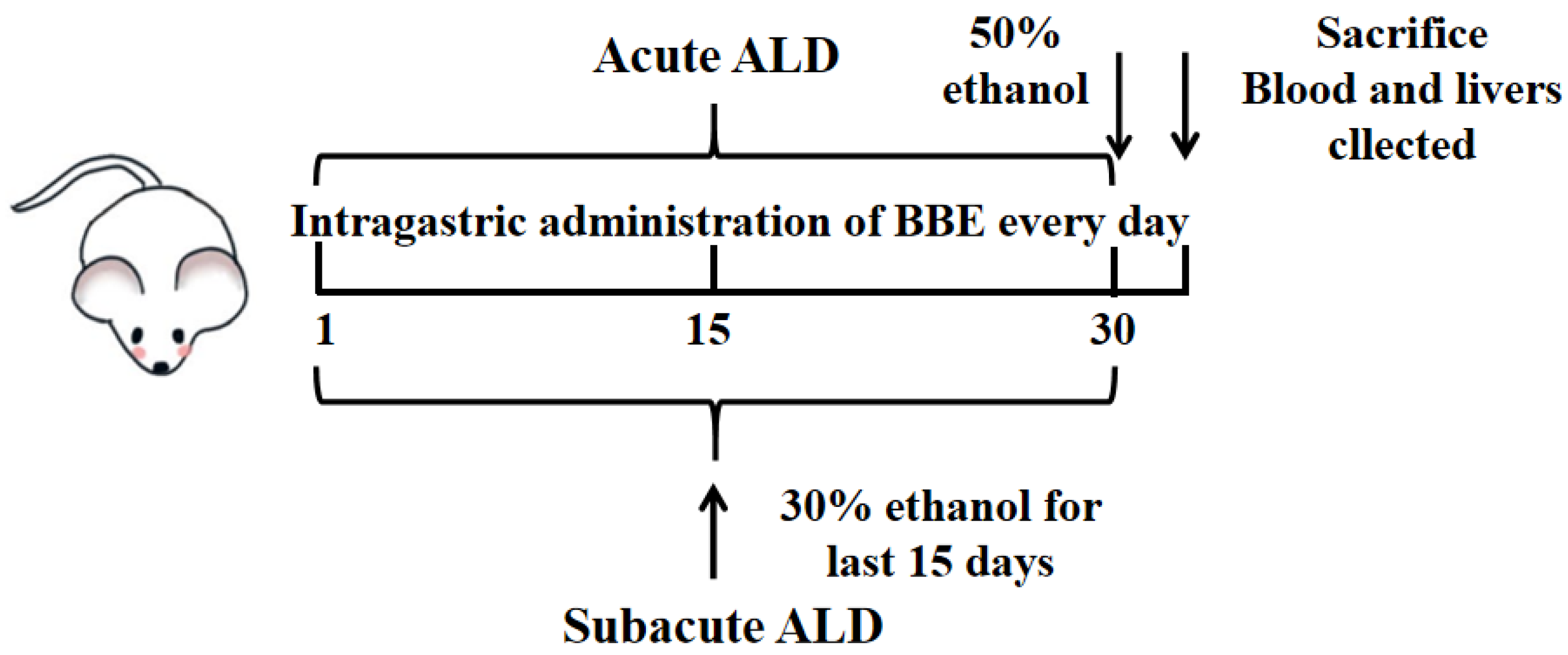
2.5.2. Experimental Design
2.5.3. Measurement of Related Indices
2.5.4. Histopathological Observation
2.5.5. Network Pharmacological Analysis of Cyanidin-3-O-glucoside
2.5.6. Establishment of PPI Network
2.5.7. Analysis of Target Pathways
2.5.8. Construction of Regulatory Network for Cyanidin-3-O-glucoside-Targets-Pathways-Disease
2.5.9. Analysis of Gene Ontology and KEGG
2.5.10. In Silico Molecular Docking Study
2.5.11. qRT-PCR Analysis
2.6. Potential Mechanisms of BBE against HepG2 Cells
2.6.1. Cell Viability Assay
2.6.2. Detection of Apoptosis by Flow Cytometry in HepG2 Cells
2.6.3. Measurement of Mitochondrial Membrane Potential in HepG2 Cells
2.6.4. Detection of ROS Levels in HepG2 Cells
2.7. Western Blotting
2.8. Data and Statistical Analysis
3. Results
3.1. The Contents of Polyphenols, Flavonoids, and Anthocyanins
3.2. Antioxidant Activities
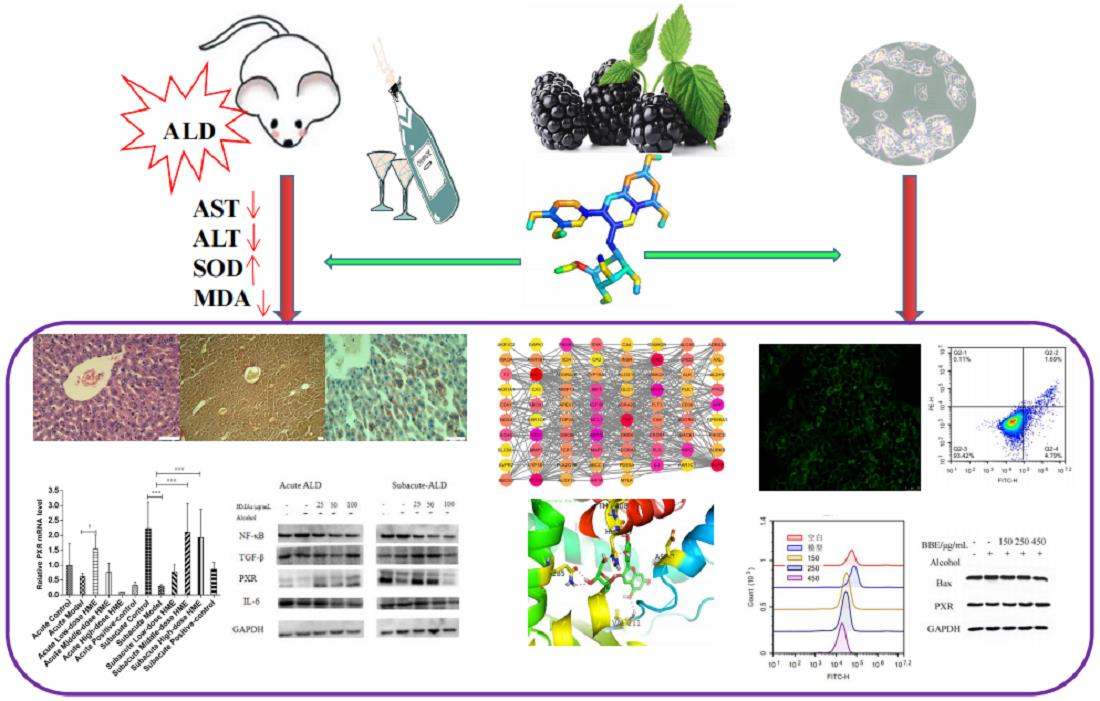
3.3. Measurement of Related Indices
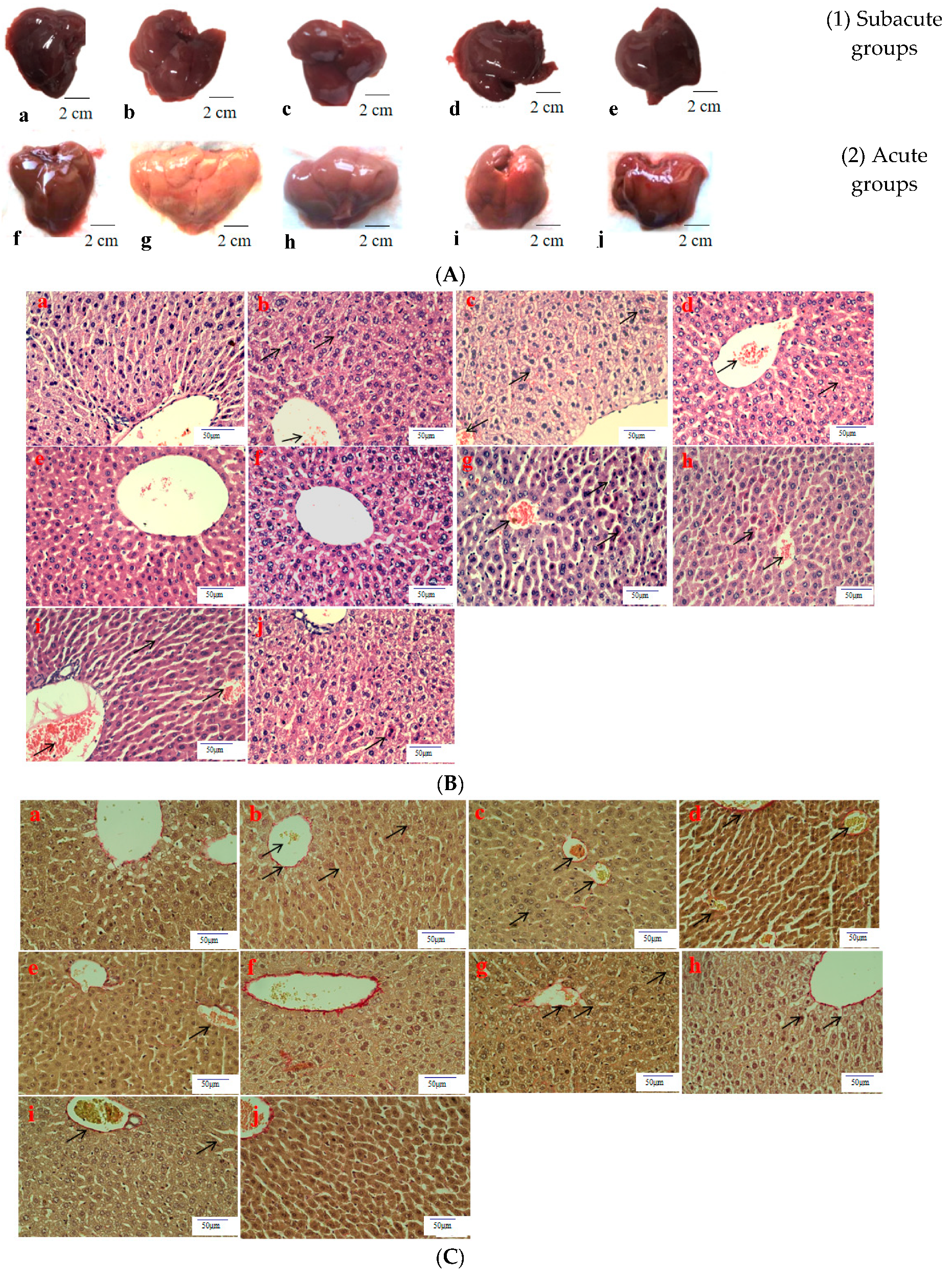
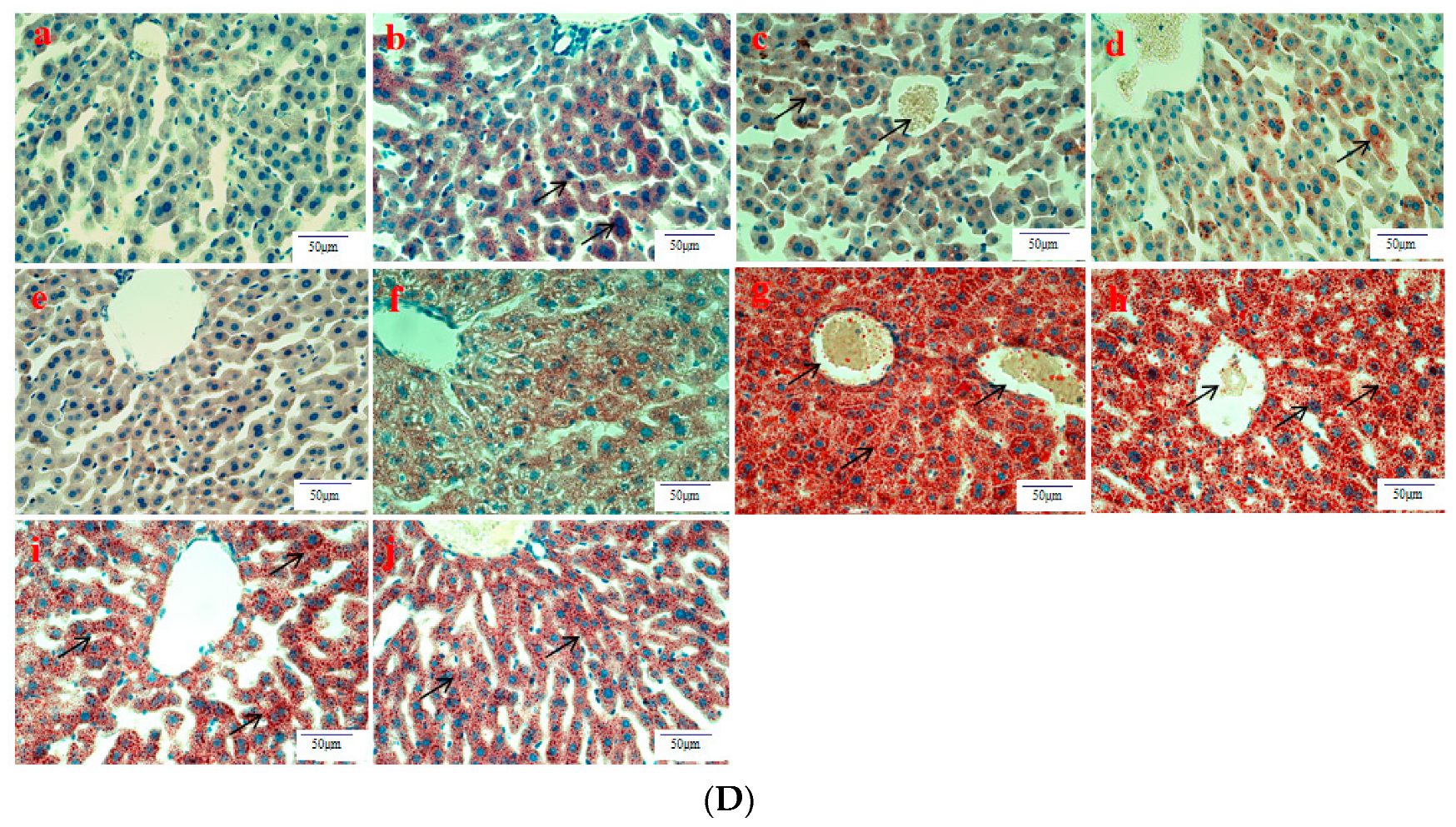
3.4. Histopathological Observation
3.5. Network Pharmacological Analysis of Cyanidin-3-O-glucoside
3.6. Establishment of PPI Network
3.7. Analysis of Target Pathways
3.8. Construction of Regulatory Network for Cyanidin-3-O-glucoside-Targets-Pathways-Disease
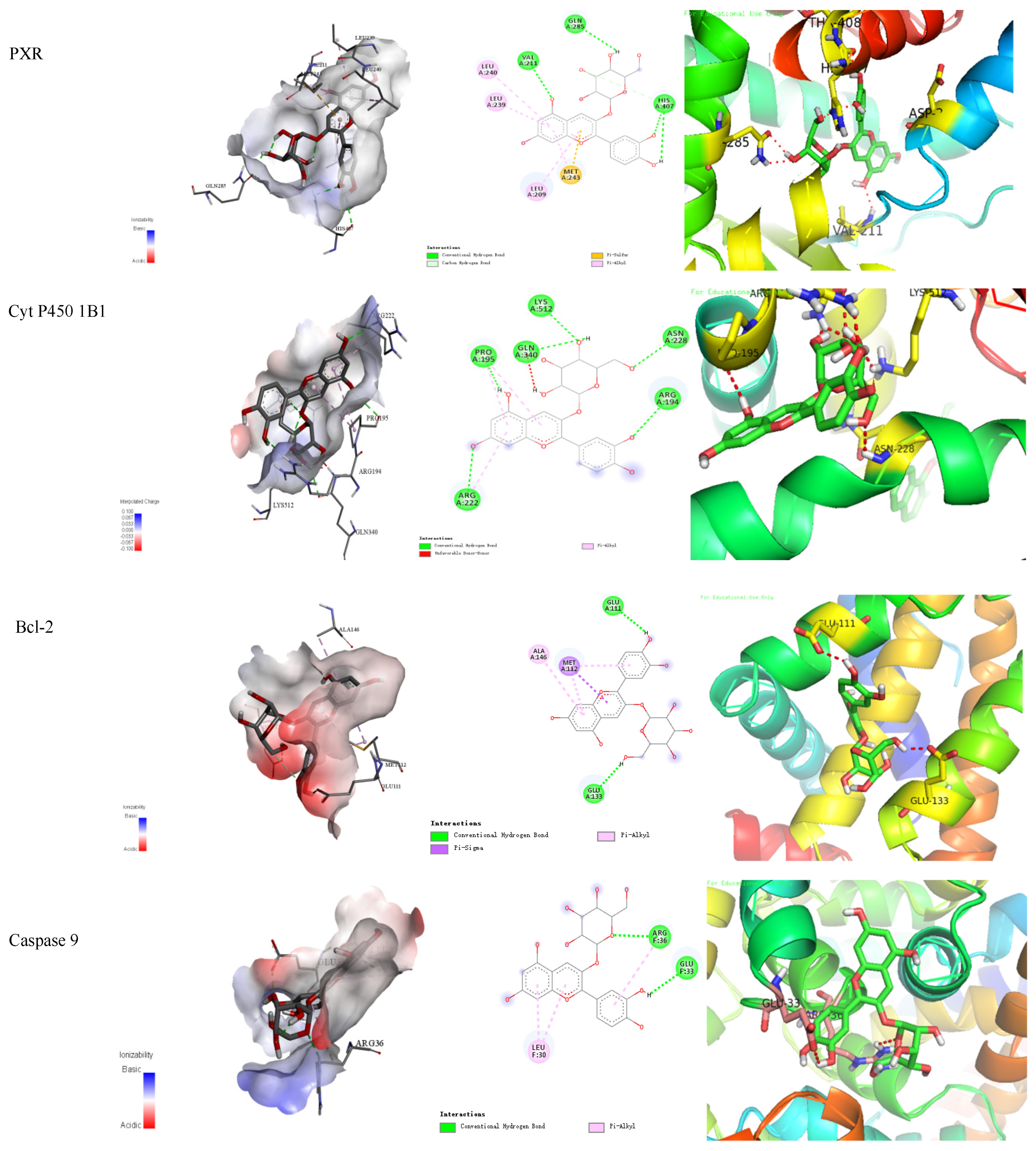
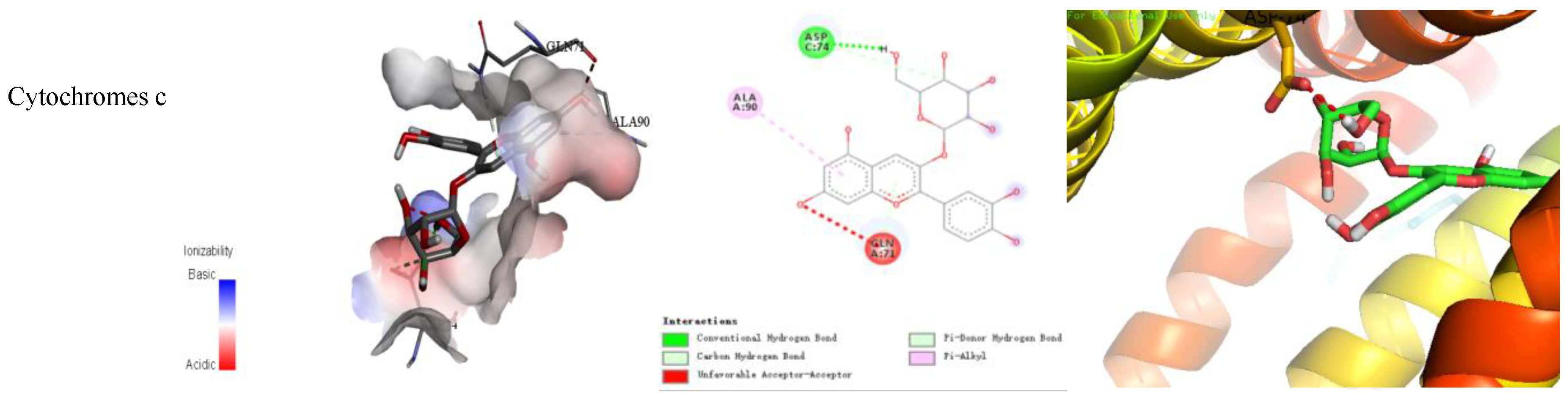
3.9. In Silico Molecular Docking Study
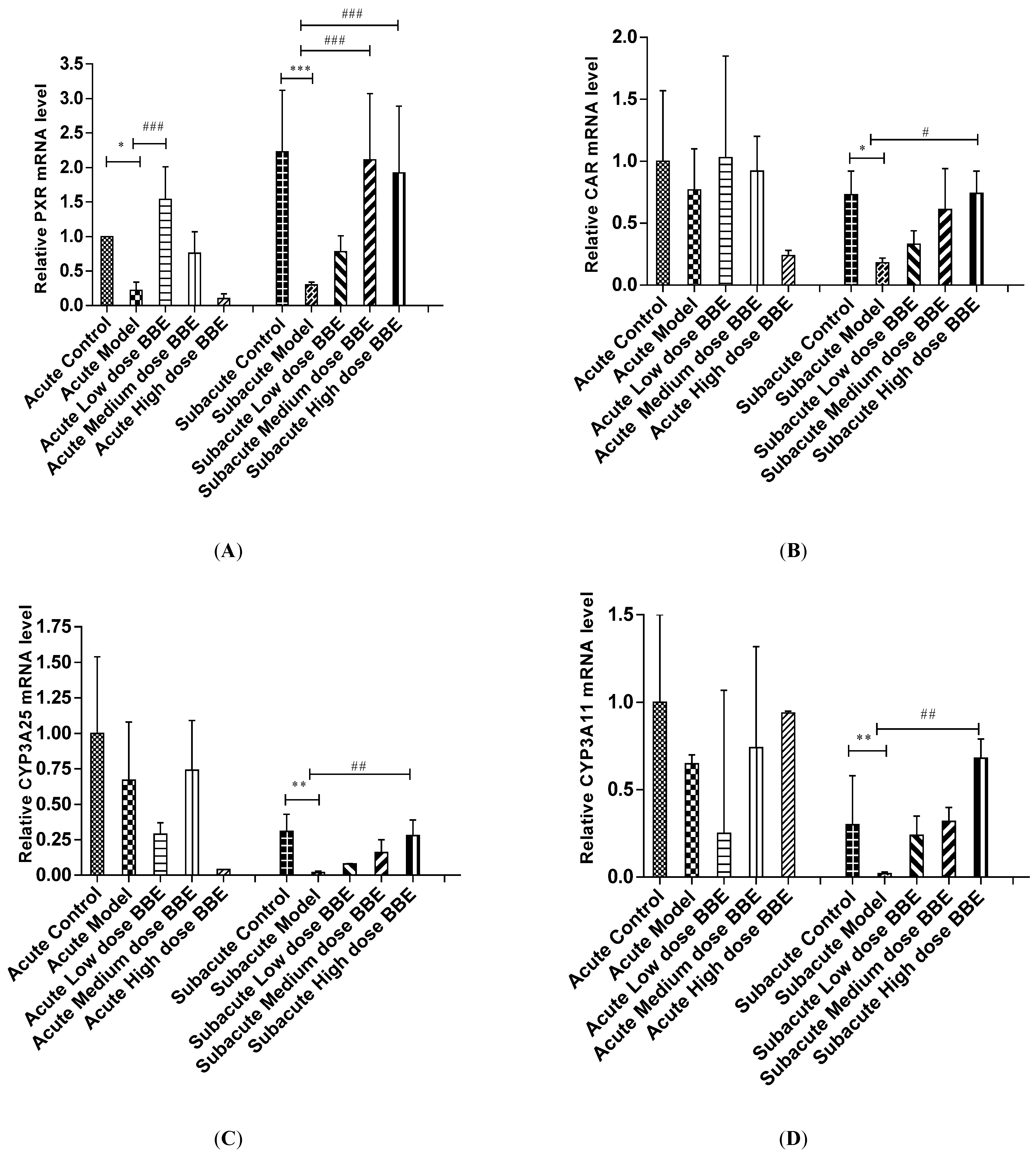
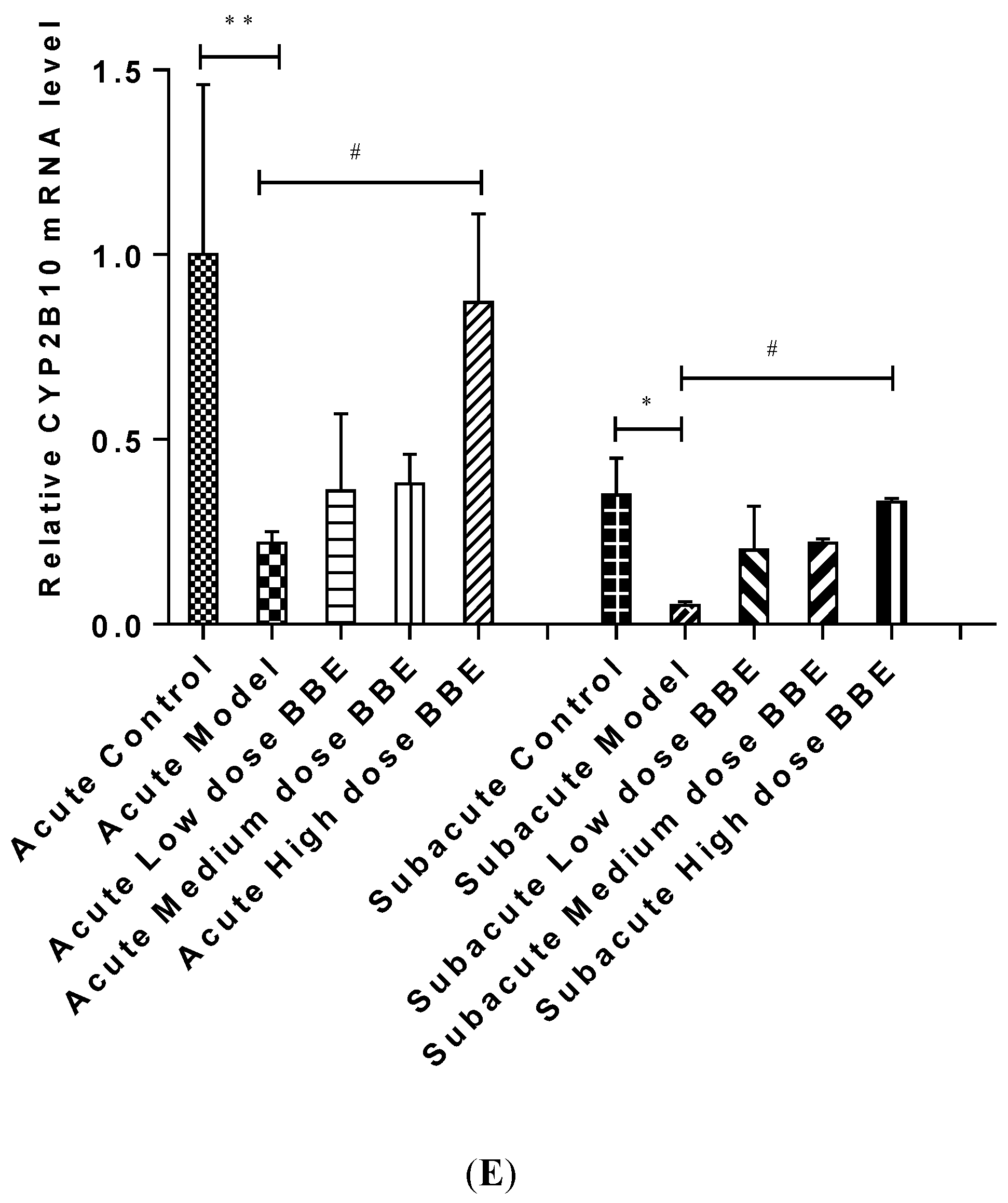
3.10. qRT-PCR Analysis
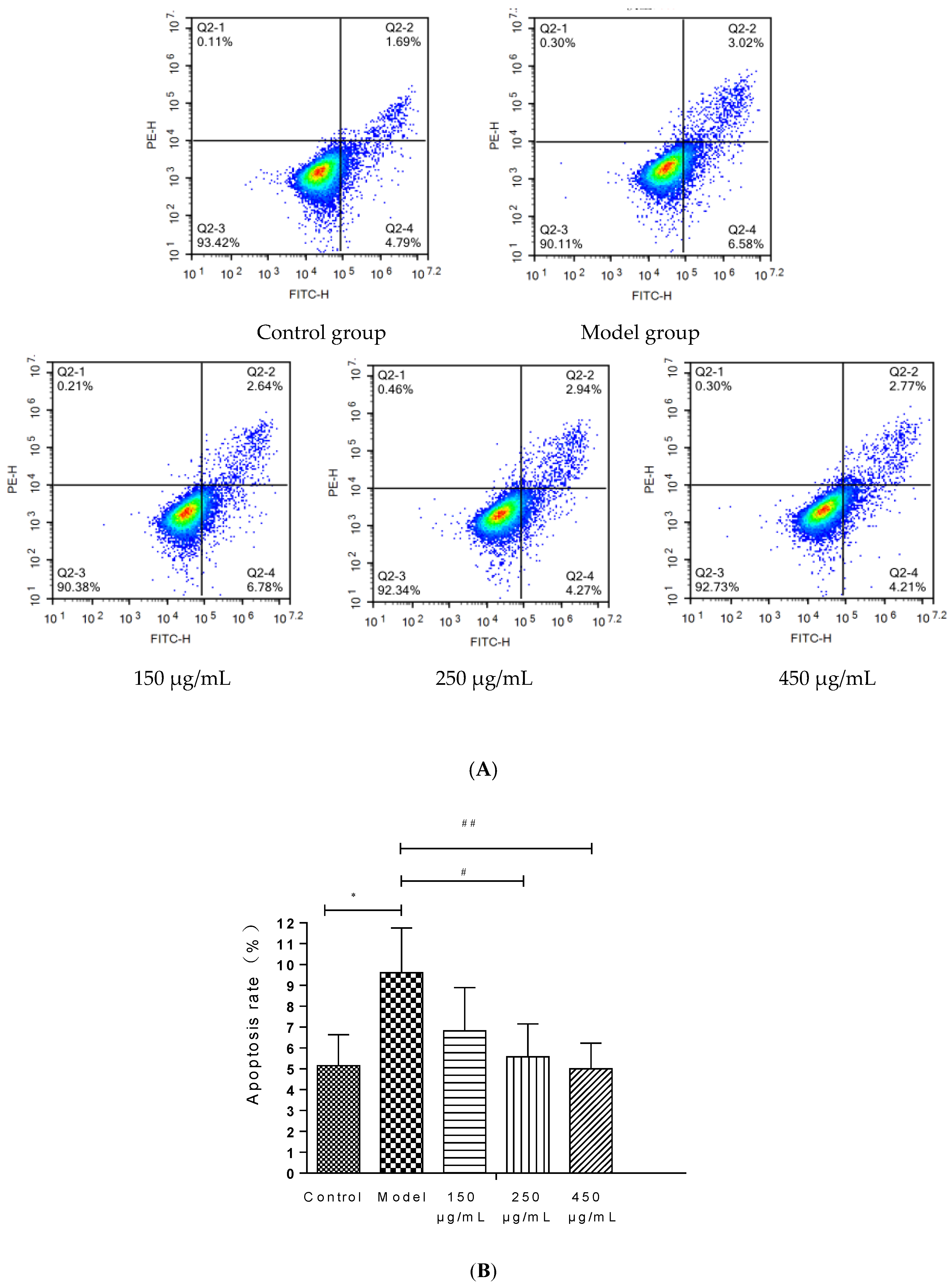
3.11. Detection of Apoptosis by Flow Cytometry in HepG2 Cells
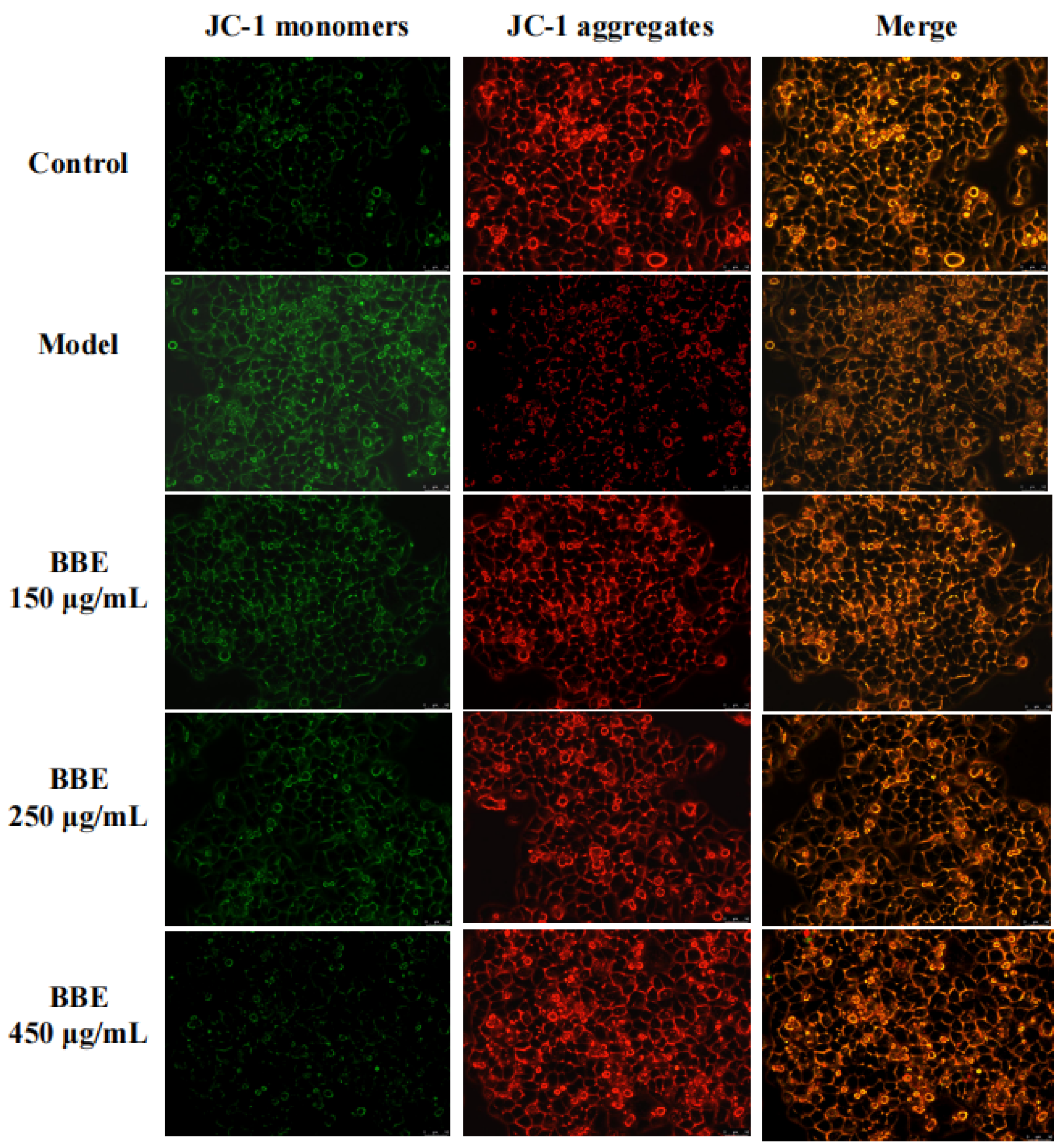
3.12. Measurement of Mitochondrial Membrane Potential in HepG2 Cells
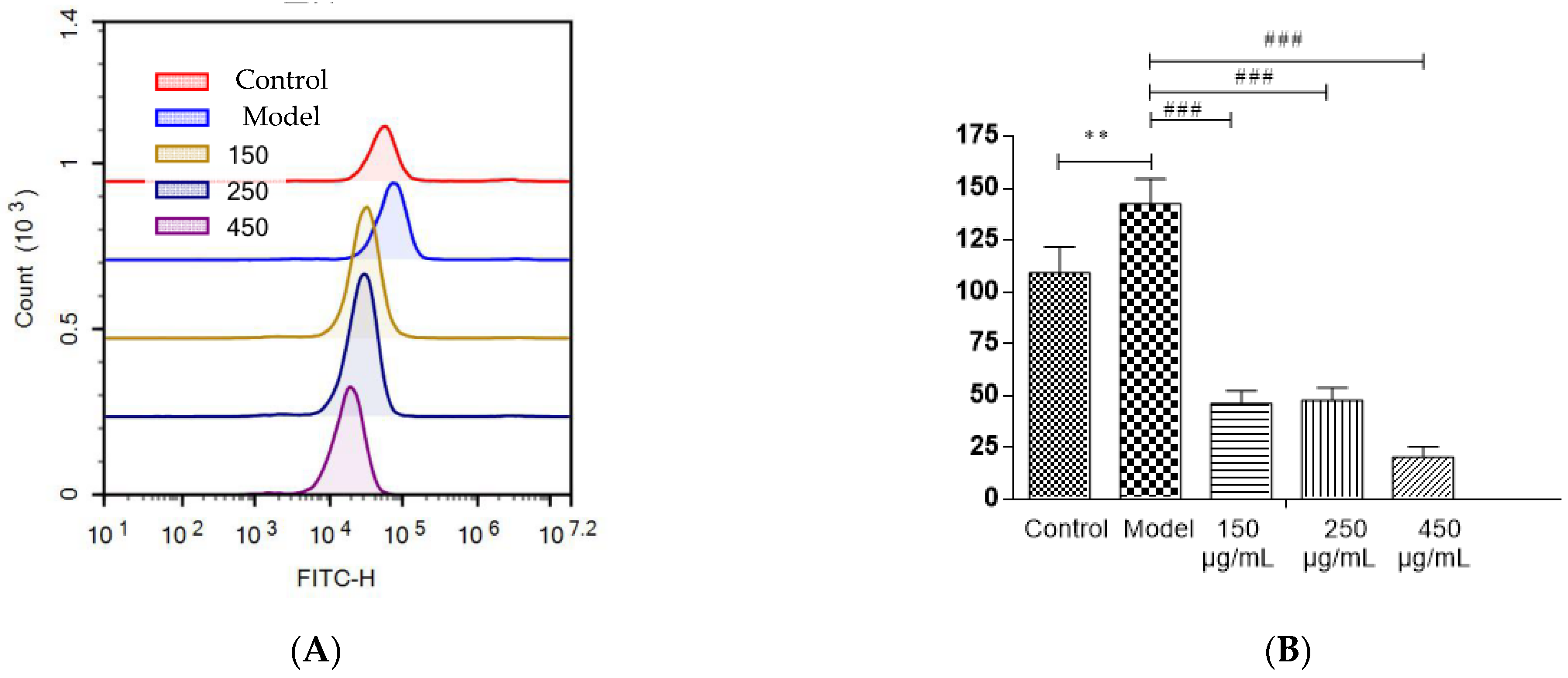
3.13. Detection of ROS Levels in HepG2 cells
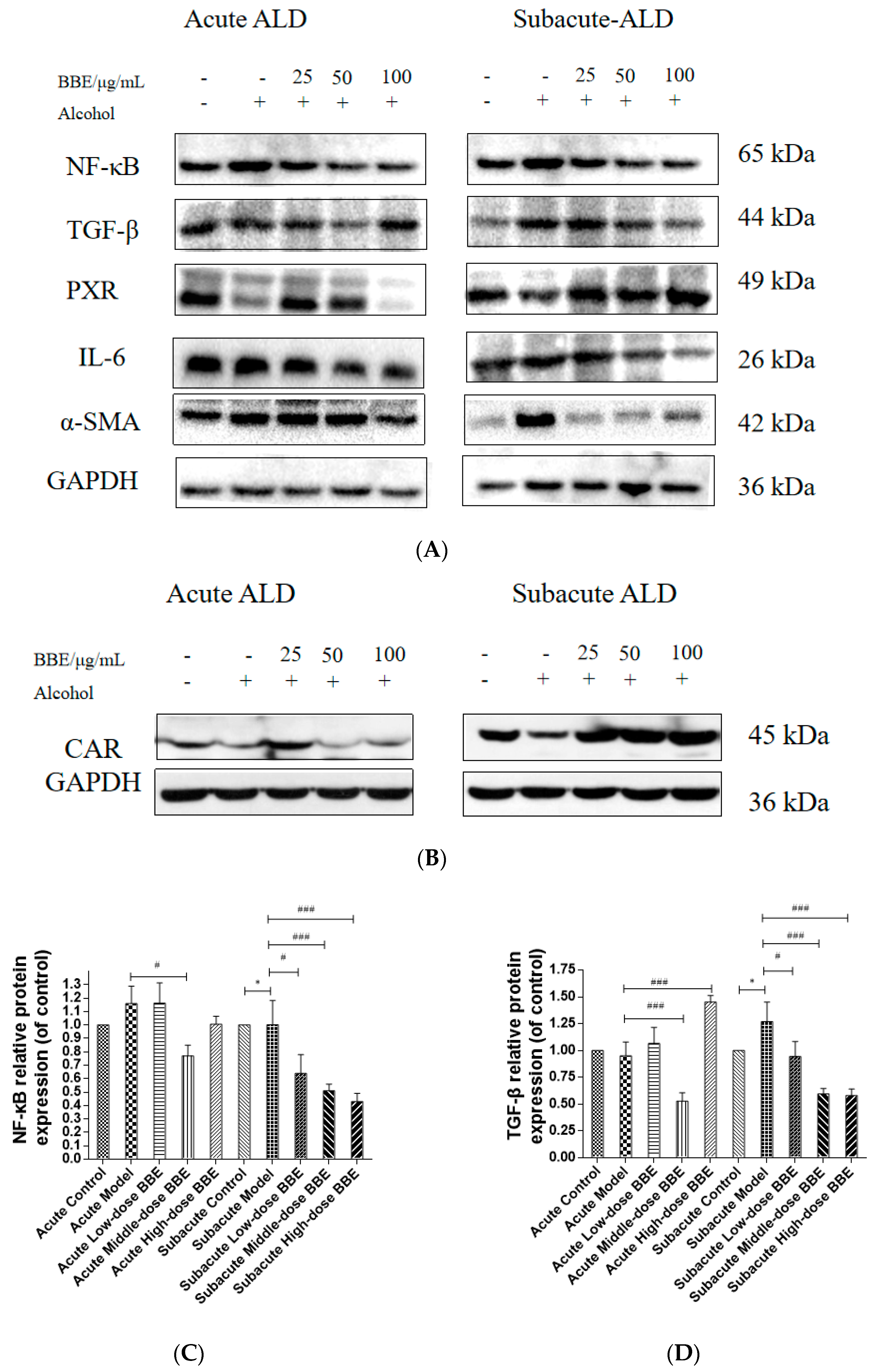
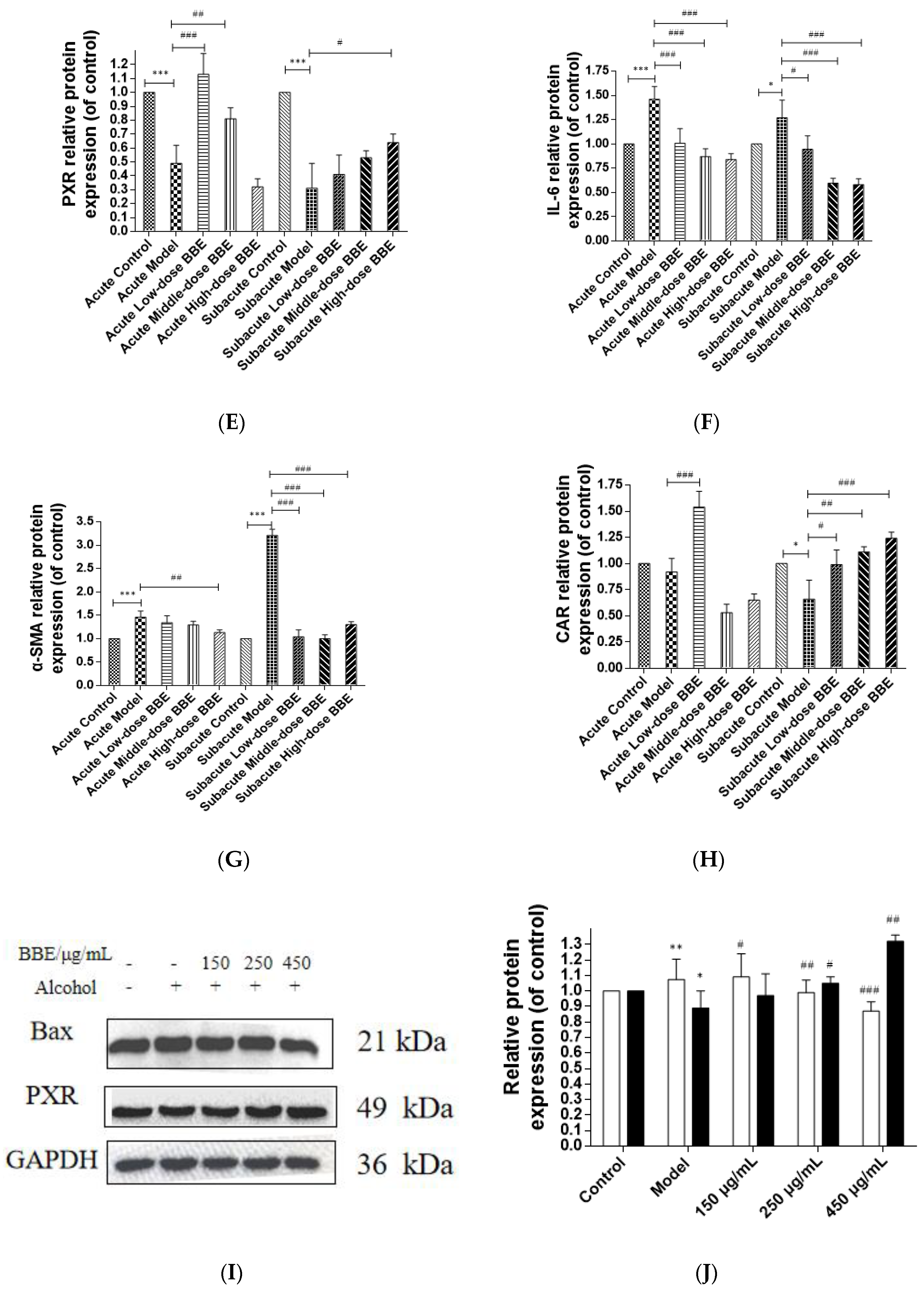
3.14. Western Blotting
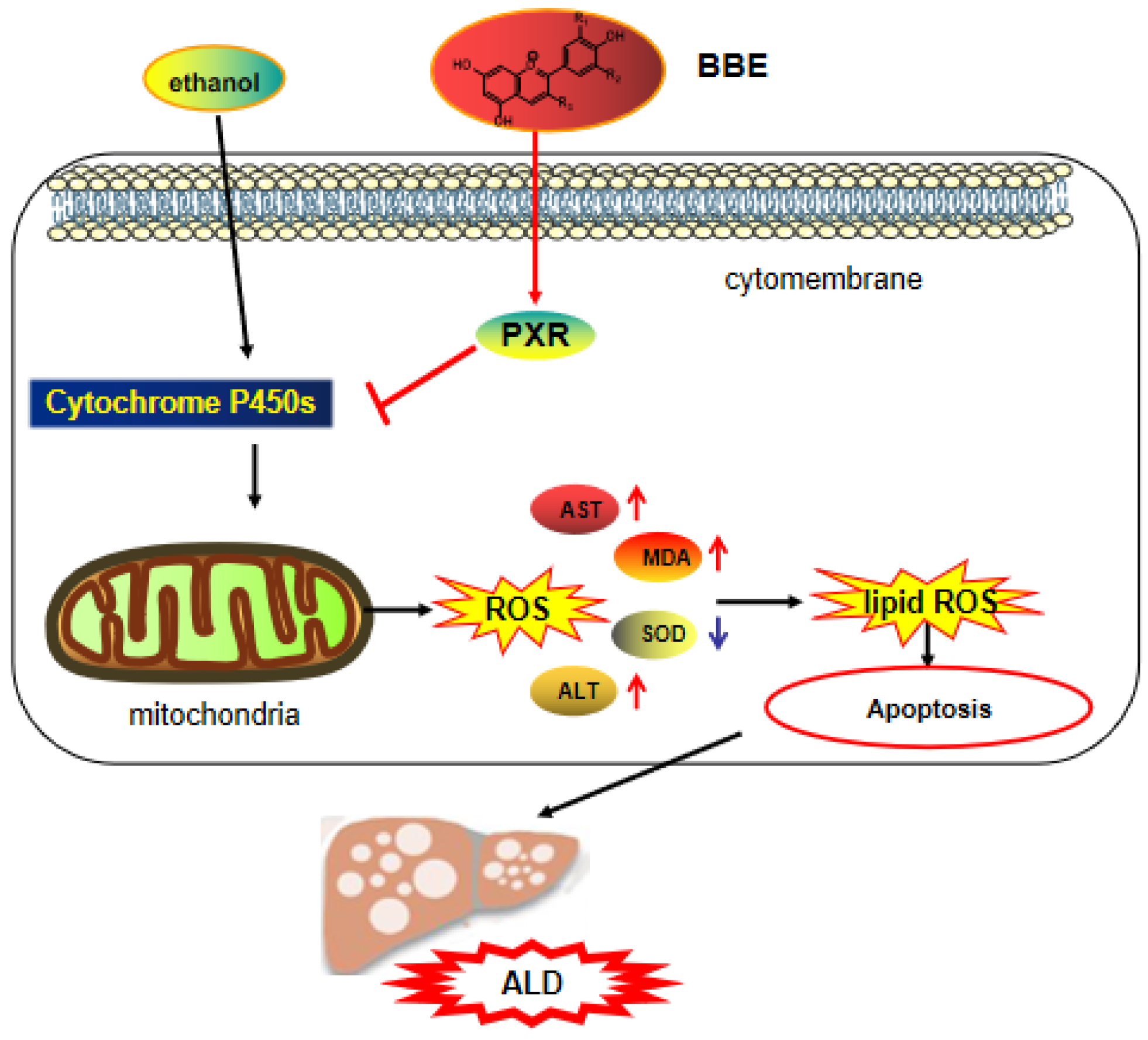
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Justice, A.C. Alcohol and the global burden of cancer: What are we missing? Lancet Oncol. 2021, 22, 1048–1049. [Google Scholar] [CrossRef]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Shi, L.G.; Hu, G.Y.; Chen, S. Saponins from Panax japonicus Protect against Alcohol-Induced Hepatic Injury in Mice by Up-regulating the Expression of GPX3, SOD1 and SOD3. Alcohol Alcohol. 2010, 45, 320–331. [Google Scholar] [CrossRef]
- Rattan, P.; Shah, V.H. Review article: Current and emerging therapies for acute alcohol-associated hepatitis. Aliment. Pharmacol. Ther. 2022, 56, 28–40. [Google Scholar] [CrossRef]
- Lee, D.U.; Chou, H.; Wang, E.; Fan, G.H.; Han, J.; Chang, K.; Kwon, J.; Lee, K.J.; Blanchard, J.; Urrunaga, N.H. The clinical implication of psychiatric illnesses in patients with alcoholic liver disease: An analysis of US hospitals. Expert Rev. Gastroenterol. Hepatol. 2022, 16, 689–697. [Google Scholar] [CrossRef]
- Singal, A.K.; Arsalan, A.; Dunn, W.; Arab, J.P.; Wong, R.J.; Kuo, Y.F.; Kamath, P.S.; Shah, V.H. Alcohol- associated liver disease in the United States is associated with severe forms of disease among young, females and Hispanics. Aliment. Pharmacol. Ther. 2021, 54, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yue, Y.; Tian, H.; Li, X.; He, W.; Ding, H. Carthamus red from Carthamus tinctorius L. exerts antioxidant and hepatoprotective effect against CCl4-induced liver damage in rats via the Nrf2 pathway. J. Ethnopharmacol. 2013, 148, 570–578. [Google Scholar] [CrossRef]
- Potts, J.R.; Goubet, S.; Heneghan, M.A.; Verma, S. Determinants of long-term outcome in severe alcoholic hepatitis. Aliment. Pharmacol. Ther. 2013, 38, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Thursz, M.R.; Richardson, P.; Allison, M.; Austin, A.; Bowers, M.; Day, C.P.; Downs, N.; Gleeson, D.; Macgilchrist, A.; Grant, A.; et al. Prednisolone or pentoxifylline for alcoholic hepatitis. New Engl. J. Med. 2015, 372, 1619–1628. [Google Scholar] [CrossRef]
- Plauth, M.; Bernal, W.; Dasarathy, S.; Merli, M.; Plank, L.D.; Schütz, T.; Bischoff, S. ESPEN guideline on clinical nutrition in liver disease. Clin. Nutr. 2019, 38, 485–521. [Google Scholar] [CrossRef]
- Petrasek, J.; Bala, S.; Csak, T.; Lippai, D.; Kodys, K.; Menashy, V.; Barrieau, M.; Min, S.; Kurt-Jones, E.A.; Szabo, G. IL-1 receptor antagonist ameliorates inflammasome- dependent alcoholic steatohepatitis in mice. J. Clin. Investig. 2012, 122, 3476–3489. [Google Scholar] [CrossRef] [PubMed]
- Aghemo, A.; Alekseeva, O.P.; Angelico, F.; Bakulin, I.G.; Bakulina, N.V.; Bordin, D.; Bueverov, A.O.; Drapkina, O.M.; Gillessen, A.; Kagarmanova, E.M.; et al. Role of silymarin as antioxidant in clinical management of chronic liver diseases: A narrative review. Ann. Med. 2022, 54, 1548–1560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xie, W.; Krasowski, M.D. PXR: A xenobiotic receptor of diverse function implicated in pharmacogenetics. Pharmacogenomics 2008, 9, 1695–1709. [Google Scholar] [CrossRef] [PubMed]
- Oladimeji, P.O.; Chen, T. PXR: More than just a master xenobiotic receptor. Mol. Pharmacol. 2018, 93, 119–127. [Google Scholar] [CrossRef]
- Ghareaghajlou, N.; Hallaj-Nezhadi, S.; Ghasempour, Z. Red cabbage anthocyanins: Stability, extraction, biological activities and applications in food systems. Food Chem. 2021, 365, 130482. [Google Scholar] [CrossRef]
- Fernández-Demeneghi, R.; Rodríguez-Landa, J.; Guzmán-Gerónimo, R.; Acosta-Mesa, H.; Meza-Alvarado, E.; Vargas-Moreno, I.; Herrera-Meza, S. Effect of blackberry juice (Rubus fruticosus L.) on anxiety-like behaviour in Wistar rats. Int. J. Food Sci. Nutr. 2019, 70, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jeong, D.; Kim, S.; Lee, S.; Sin, H.; Yu, K.; Jeong, S.; Kim, S. Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling. Nutrients 2019, 11, 2429. [Google Scholar] [CrossRef] [PubMed]
- Figueira, I.; Tavares, L.; Jardim, C.; Costa, I.; Terrasso, A.P.; Almeida, A.F.; Santo, C.N. Blood-brain barrier transport and neuroprotective potential of blackberry-digested polyphenols: An in vitro study. Eur. J. Nutr. 2019, 58, 113–130. [Google Scholar] [CrossRef]
- Cenk, E.; Schmutz, C.; Pahlke, G.; Oertel, A.; Kollarova, J.; Mock, H.P.; Matros, A.; Marko, D. Immunomodulatory Properties of blackberry anthocyanins in THP-1 derived macrophages. Int. J. Mol. Sci. 2021, 22, 10483. [Google Scholar] [CrossRef]
- Tatar, M.; Bagheri, Z.; Varedi, M.; Naghibalhossaini, F. Blackberry extract inhibits telomerase activity in human colorectal cancer cells. Nutrtion Cancer Int. J. 2019, 71, 461–471. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Wang, D.; Huo, Y.; Ji, B. Anthocyanin-rich extracts from blackberry, wild blueberry, strawberry, and chokeberry: Antioxidant activity and inhibitory effect on oleic acid-induced hepatic steatosis in vitro. Int. J. Food Sci. Nutr. 2016, 96, 2494–2503. [Google Scholar] [CrossRef]
- Xiao, T.; Luo, Z.; Guo, Z.; Wang, X.; Ding, M.; Wang, W.; Shen, X.; Zhao, Y. Multiple Roles of Black Raspberry Anthocyanins Protecting against Alcoholic Liver Disease. Molecules 2021, 26, 2313. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Guo, Z.; Sun, B.; Zhao, Y. Identification of Anthocyanins from Four Kinds of Berries and Their Inhibition Activity to α-Glycosidase and Protein Tyrosine Phosphatase 1B by HPLC FT-ICR MS/MS. J. Agric. Food Chem. 2017, 65, 6211–6221. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zheng, X.; Zhao, L.; Yi, J.; Cai, S. Effects and interaction mechanism of soybean 7S and 11S globulins on anthocyanin stability and antioxidant activity during in vitro simulated digestion. Curr. Res. Food Sci. 2021, 4, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, H.; Zhu, K.; Yang, Y.; Li, J.; Jiang, H.; Liu, Z. Investigation of candidate molecular biomarkers for expression profile analysis of the Gene expression omnibus (GEO) in acute lymphocytic leukemia (ALL). Biomed. Pharm. 2019, 120, 109530. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L.; Seruga, M.; Medvidovic-Kosanovic, M.; Novak, I. Anthocyanin content and antioxidant activity of various red fruit juices. Dtsch. Lebensm. Rundsch. 2007, 103, 58–64. [Google Scholar]
- Cho, M.J.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid glycosides and antioxidant capacity of various blackberry, blueberry and red grape genotypes determined by high-performance liquid chromatography/mass spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, W.; Lü, L.; Li, W. Change Regularity of Polyphenol Content in Blackberry Fruit during Growth and Development. J. Jilin Agric. Univ. 2013, 35, 457–462. [Google Scholar]
- Xiao, T.; Guo, Z.; Bi, X.; Zhao, Y. Polyphenolic profile as well as anti-oxidant and anti-diabetes effects of extracts from freeze-dried black raspberries. J. Funct. Foods 2017, 31, 179–187. [Google Scholar] [CrossRef]
- Sozio, M.; Crabb, D.W. Alcohol and lipid metabolism. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E10–E16. [Google Scholar] [CrossRef]
- Li, S.; Fan, T.P.; Jia, W.; Lu, A.; Zhang, W. Network Pharmacology in Traditional Chinese Medicine. Evid. Based Complement. Altern. Med. 2014, 2014, 138460. [Google Scholar] [CrossRef]
- Jiang, X.; Tang, X.; Zhang, P.; Liu, G.; Guo, H. Cyanidin-3-O-b-glucoside protects primary mouse hepatocytes against high glucose-induced apoptosis by modulating mitochondrial dysfunction and the PI3K/Akt pathway. Biochem. Pharmacol. 2014, 90, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X. Anthocyanins: Structural characteristics that result in unique metabolic patterns and biological activities. Free Radic. Res. 2006, 40, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Lipton, S.A. Preventing Ca2+-mediated nitrosative stress in neurodegenerative diseases: Possible pharmacological strategies. Cell Calcium 2010, 47, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Xiang, J.; Zhu, W.; Li, Z.; Ling, S. Effect of juice and fermented vinegar from Hovenia dulcis peduncles on chronically alcohol induced liver damage in mice. Food Funct. 2012, 3, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S. Alcoholic fatty liver: Its pathogenesis and mechanism of progression to inflammation and fibrosis. Alcohol 2004, 34, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, J.; Liu, C.; Kong, P.; Yan, J.; Cheng, H. Protective Effect of Green Tea Polyphenols against Chronic Alcoholic Hepatic Injury in Mice. Food Sci. 2011, 32, 310–313. [Google Scholar]
- He, Z.; Wang, Y.; Liu, L.; Lou, Z. Effects of Panax Japonicus Extractions on Serum Biochemical Indices and Inflammatory Factors in Mice with Alcoholic Liver Injury. Zhejiang J. Integr. Tradit. Chin. West. Med. 2018, 28, 21–24. [Google Scholar]
- Baldi, E.; Burra, P.; Plebani, M.; Salvagnini, M. Serum malondialdehyde and mitochondrial aspartate aminotransferase activity as markers of chronic alcohol intake and alcoholic liver disease. Ital. J. Gastroenterol. Hepatol. 1993, 25, 429–432. [Google Scholar]
- Sun, H.; Mu, T.; Liu, X.; Zhang, M.; Chen, J. Purple sweet potato (Lpomoea batatas L.) anthocyanins: Preventive effect on acute and subacute alcoholic liver damage and dealcoholic effect. J. Agric. Food Chem. 2014, 62, 2364–2373. [Google Scholar] [CrossRef]
- Yang, C.; Liao, A.; Cui, Y.; Yu, G.; Hou, Y.; Pan, L.; Chen, W.; Zheng, S.; Li, X.; Ma, J.; et al. Wheat embryo globulin protects against acute alcohol-induced liver injury in mice. Food Chem. Toxicol. 2021, 153, 112240. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Jiang, Y.; Chen, P.; Fan, X.; Li, D.; Liu, A.; Ma, X.; Xie, W.; Liu, P.; Gonzalez, F.J.; et al. Schisandrol B protects against cholestatic liver injury through pregnane X receptors. Br. J. Pharmacol. 2017, 174, 672–688. [Google Scholar] [CrossRef] [PubMed]
- Ostberg, T.; Bertilsson, G.; Jendeberg, L.; Berkenstam, A.; Uppenberg, J. Identification of residues in the PXR ligand binding domain critical for species specific and constitutive activation. Eur. J. Biochem. 2002, 269, 4896–4904. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Viswakarma, N.; Reddy, J.K. Med1 subunit of the mediator complex in nuclear receptor-regulated energy metabolism, liver regeneration, and hepatocarcinogenesis. Gene Expr. 2014, 16, 63–75. [Google Scholar] [CrossRef]
- Wilkinson, G.R. Cytochrome P4503A (CYP3A) metabolism: Prediction of in vivo activity in humans. J. Pharmacokinet. Biopharm. 1996, 24, 475–490. [Google Scholar] [CrossRef]
- Jones, B.C.; Rollison, H.; Johansson, S.; Kanebratt, K.P.; Lambert, C.; Vishwanathan, K.; Andersson, T.B. Managing the risk of CYP3A induction in drug development: A strategic approach. Drug Metab. Dispos. 2017, 45, 35–41. [Google Scholar] [CrossRef]
- Komura, H.; Iwaki, M. Species differences in in vitro and in vivo small intestinal metabolism of CYP3A substrates. J. Pharm. Sci. 2008, 97, 1775–1800. [Google Scholar] [CrossRef] [PubMed]
- Christidis, C.; Karatayli, E.; Hall, R.A.; Weber, S.N.; Reichert, M.C.; Hohl, M.; Qiao, S.; Boehm, U.; Lütjohann, D.; Lammert, F.; et al. Fibroblast Growth Factor 21 Response in a Preclinical Alcohol Model of Acute-on-Chronic Liver Injury. Int. J. Mol. Sci. 2021, 22, 7898. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, H.; Liao, C.; Chou, A.; Lee, H. The Timing and Effects of Low-Dose Ethanol T reatment on Acetaminophen-Induced Liver Injury. Life 2021, 11, 1094. [Google Scholar] [CrossRef]
- Larsen, B.D.; Sørensen, C.S. The caspase-activated DNase: Apoptosis and beyond. FEBS J. 2017, 284, 1160–1170. [Google Scholar] [CrossRef]
- Sumida, Y.; Niki, E.; Naito, Y.; Yoshikawa, T. Involvement of free radicals and oxidative stress in NAFLD/NASH. Free Radic. Res. 2013, 47, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.C.; Cassanye, A.; Martín-Gari, M.; Granado-Serrano, A.B.; Portero-Otín, M. Effect of dietary bioactive compounds on mitochondrial and metabolic flexibility. Diseases 2016, 4, 14. [Google Scholar] [CrossRef]
- Fulda, S. Modulation of mitochondrial apoptosis by PI3K inhibitors. Mitochondrion 2013, 13, 195–198. [Google Scholar] [CrossRef]
- Handschin, C.; Podvinec, M.; Meyer, U.A. CXR, a chicken xenobiotic-sensing orphan nuclear receptor, is related to both mammalian pregnane X receptor (PXR) and constitutive androstane receptor (CAR). Proc. Natl. Acad. Sci. USA 2000, 97, 10769–10774. [Google Scholar] [CrossRef]
- Garcia, M.; Thirouard, L.; Sedes, L.; Monrose, M.; Holota, H.; Caira, F.; Volle, D.H.; Beaudoin, C. Nuclear receptor metabolism of bile acids and xenobiotics: A coordinated detoxification system with impact on health and diseases. Int. J. Mol. Sci. 2018, 19, 3630. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Wei, Y.; Huo, X.; Sun, X.; Zhang, C.; Ming, W.; Luo, Z.; Du, C.; Li, Y.; Xu, H.; et al. Pregnane X receptor (PXR) protects against cisplatin-induced acute kidney injury in mice. BBA-Mol. Basis Dis. 2021, 1867, 165996. [Google Scholar] [CrossRef]
- Yan, W.; Ma, X.; Zhao, X.; Zhang, S. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des. Dev. Ther. 2018, 12, 3961–3972. [Google Scholar] [CrossRef]
- Feng, L.-X.; Sun, P.; Mi, T.; Liu, M.; Liu, W.; Yao, S.; Cao, Y.-M.; Yu, X.-L.; Wu, W.-Y.; Jiang, B.-H. Agglutinin isolated from Arisema heterophyllum Blume induces apoptosis and autophagy in A549 cells through inhibiting PI3K/Akt pathway and inducing ER stress. Chin. J. Nat. Med. 2016, 14, 856–864. [Google Scholar] [CrossRef]
- Gu, X.; Yao, Y.; Cheng, R.; Zhang, Y.; Dai, Z.; Wan, G.; Yang, Z.; Cai, W.; Gao, G.; Yang, X. Plasminogen K5 activates mitochondrial apoptosis pathway in endothelial cells by regulating Bak and Bcl-xL subcellular distribution. Apoptosis 2011, 16, 846–855. [Google Scholar] [CrossRef]
- Zhao, N.; Xia, G.; Cai, J.; Li, Z.; Lv, X. Adenosine receptor A2B mediates alcoholic hepatitis by regulating cAMP levels and the NF-KB pathway. Toxicol. Lett. 2022, 359, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Kartasheva-Ebertz, D.; Gaston, J.; Lair-Mehiri, L.; Massault, P.; Scatton, O.; Vaillant, J.; Morozov, V.-A.; Pol, S.; Lagaye, S. Adult human liver slice cultures: Modelling of liver fibrosis and evaluation of new anti-fibrotic drugs. World J. Hepatol. 2021, 13, 187–217. [Google Scholar] [CrossRef] [PubMed]
- Al-Humadi, H.-W.; Al-Saigh, R.; Sahib, A. The impact of low alcohol consumption on the liver and inflammatory cytokines in diabetic rats. Adv. Clin. Exp. Med. 2019, 28, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Unny, A.K.; Vij, M.; Safwan, M.; Balaji, M.S.; Rela, M. α-Smooth muscle actin expression predicts the outcome of Kasai portoenterostomy in biliary atresia. Saudi J. Gastroenterol. 2019, 25, 101–105. [Google Scholar] [CrossRef]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver fibrosis: Mechanistic concepts and therapeutic perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| PXR | AGACGGCAGCATCTGGAACTAC | GTTTCTGGAAGCCACCATTAGG |
| CAR | CCACAGGCTATCATTTCCACG | CCTTCCAGCAAACGGACAGAT |
| CYP3A25 | GGAGGCCTGAACTGCTAAAG | GTAGTTGAAAATGGTGCCAAGTAAC |
| CYP3A11 | GGTGCTCCTAGCAATCAGCTT | AAGGAGAGGCGTTTGACCATC |
| CYP2B10 | TCTGCCCTTCTCAACAGGAAAG | AAGGAGAGGCGTTTGACCATC |
| GAPDH | TTGGAGCCCTGGAGATTTGGA | TGAGGTCAATGAAGGGGTCGT |
| Species | TP a | TF b | TMA c | DPPH d | ABTS d | FRAP e |
|---|---|---|---|---|---|---|
| Blackberry | 5.70 ± 1.34 | 105.70 ± 12.11 | 24.50 ± 3.49 | 22.08 ± 7.13 | 51.69 ± 7.84 | 2.93 |
| Vc | 4.76 ± 1.15 | 36.23 ± 5.78 | 1 |
| Treatment Group | Final Weight (g) | Liver Index (%) | AST (U/L) | ALT (U/L) | SOD (U/mL) | MDA (U/mL) |
|---|---|---|---|---|---|---|
| (a) Subacute ALD mice | ||||||
| Control | 41.45 ± 2.78 | 4.23 ± 0.24 | 135.60 ± 8.75 | 55.07 ± 7.03 | 260.13 ± 17.52 | 5.76 ± 0.24 |
| Model | 34.48 ± 3.45 | 3.76 ± 0.31 | 189.92 ± 15.87 *** | 94.08 ± 1.55 *** | 177.88 ± 3.24 *** | 11.37 ± 0.14 |
| Low-dose BBE | 36.69 ± 4.05 | 3.81 ± 0.55 | 169.60 ± 11.35 ### | 82.01 ± 1.09 ### | 191.01 ± 2.89 ### | 8.79 ± 0.24 ### |
| Medium-dose BBE | 35.87 ± 3.61 | 3.71 ± 0.39 | 163.73 ± 10.24 ### | 72.28 ± 2.23 ### | 229.02 ± 6.96 ### | 8.19 ± 0.65 ### |
| High-dose BBE | 36.12 ± 3.34 | 3.69 ± 0.25 | 151.18 ± 5.27 ### | 61.57 ± 3.97 ### | 235.07 ± 3.15 ### | 8.03 ± 0.42 ### |
| (b) Acute ALD mice | ||||||
| Control | 40.90 ± 0.45 | 3.29 ± 0.30 | 121.14 ± 9.95 | 44.71 ± 7.13 | 306.80 ± 17.14 | 6.37 ± 0.42 |
| Model | 41.00 ± 0.31 | 4.30 ± 0.21 | 210.14 ± 22.60 | 72.65 ± 0.55 | 231.62 ± 4.29 | 8.94 ± 0.21 |
| Low-dose BBE | 41.31 ± 0.62 | 4.04 ± 0.57 | 176.69 ± 14.77 | 70.82 ± 1.19 | 245.44 ± 3.58 | 8.49 ± 0.08 |
| Medium-dose BBE | 41.58 ± 0.45 | 3.33 ± 0.22 | 158.99 ± 10.16 | 60.01 ± 1.23 | 253.22 ± 7.68 | 7.28 ± 0.16 |
| High-dose BBE | 41.69 ± 0.56 | 3.10 ± 1.27 | 151.37 ± 3.76 | 53.94 ± 4.97 | 266.19 ± 2.55 | 7.12 ± 0.21 |
| Targets | Binding Free Energy (kcal/mol) | Numbers of Bonds |
|---|---|---|
| PXR | −8.7 | 10 |
| Cyt P450 1B1 | −8.2 | 14 |
| Bcl-2 | −7.3 | 6 |
| Caspase 9 | −7.2 | 5 |
| Cytochrome c | −7.6 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, T.; Guo, Z.; Fu, M.; Huang, J.; Wang, X.; Zhao, Y.; Tao, L.; Shen, X. Amelioration of Alcoholic Liver Disease by Activating PXR-Cytochrome P450s Axis with Blackberry Extract. Separations 2022, 9, 321. https://doi.org/10.3390/separations9100321
Xiao T, Guo Z, Fu M, Huang J, Wang X, Zhao Y, Tao L, Shen X. Amelioration of Alcoholic Liver Disease by Activating PXR-Cytochrome P450s Axis with Blackberry Extract. Separations. 2022; 9(10):321. https://doi.org/10.3390/separations9100321
Chicago/Turabian StyleXiao, Ting, Zhenghong Guo, Min Fu, Jiaoyan Huang, Xiaowei Wang, Yuqing Zhao, Ling Tao, and Xiangchun Shen. 2022. "Amelioration of Alcoholic Liver Disease by Activating PXR-Cytochrome P450s Axis with Blackberry Extract" Separations 9, no. 10: 321. https://doi.org/10.3390/separations9100321
APA StyleXiao, T., Guo, Z., Fu, M., Huang, J., Wang, X., Zhao, Y., Tao, L., & Shen, X. (2022). Amelioration of Alcoholic Liver Disease by Activating PXR-Cytochrome P450s Axis with Blackberry Extract. Separations, 9(10), 321. https://doi.org/10.3390/separations9100321