Abstract
Cordycepin (3′-deoxyadenosine) is a potent bioactive metabolite of the medicinal fungus Cordyceps militaris, which has been increasingly exploited to treat various chronic diseases in humans. However, the current synthesis and purification procedures of cordycepin are principally laborious and complicated. This study provides a simple protocol approach to isolate and purify cordycepin from C. militaris by normal phase column chromatography at room temperature. Besides, this is the first to investigate the potential of cordycepin and cordycepin-included extracts from C. militaris for making Kombucha functional products. By a repeated column chromatography, an amount of 1.16 g of cordycepin is isolated from 2.8 kg of fruiting bodies of C. militaris, which obtained an efficiency of 83.26% compared to that estimated by high-performance liquid chromatography (HPLC). The purity of cordycepin is confirmed by thin-layer chromatography (TLC), HPLC, and proton nuclear magnetic resonance (1H NMR). In addition, kombucha-fermented extracts from cordycepin and cordycepin-included fractions show potential biological activities in terms of antioxidant, anti-diabetes via α-glucosidase inhibitory assay, and cytotoxicity via MTT assay on Meg-01 and HL-60 cell lines. Further studies on optimization of extraction protocol and verification of health benefits of kombucha products from cordycepin should be conducted prior to the official mass production.
1. Introduction
Cordyceps is a medical mushroom that has been used as a traditional medicine in China since ancient times. The genus Cordyceps is classified in the phylum Ascomycota, the class Claviciptiaceae, the order Hypocreales, and has more than 400 species [1,2]. There are 12 Cordyceps species used for medical purposes, including C. sinensis, C. militaris, C. liangshanesis, C. taishanesis, C. shanxiensis, C. gansuensis, C. grasspara, C. kangdingensis, C. guizhouensis, C. nutans, C. tricentric, and C. gunnii [3]. Among them, C. sinensis has been associated with human society for more than several hundred years, with various recognized benefits for human health [1]. As people become more health-conscious, the demand for dietary supplements, nutraceuticals, and functional foods produced from C. sinensis is growing, but the supply of natural C. sinensis is not enough to meet the demand [4]. This is because C. sinensis is distributed only in limited areas, such as the Tibetan Plateau, where the altitude is 3500–5000 m above sea level [1]. This imbalance between supply and demand not only led to the overexploitation of C. sinensis but also to the price being hiked [2,4]. In order to overcome the situation of strained natural resources, the research has struggled to find alternative resources.
Among the Cordyceps species, C. militaris stood out as the most likely candidate. The major advantages of C. militaris cultivation can be mentioned as (i) the artificial culture method of C. militaris is simpler and less costly than that of C. sinensis [2] and (ii) the artificial culture of C. militaris contains quantitatively and qualitatively comparable bioactive substances to those involved in C. sinensis, and exhibits the same level of bioactivity [5,6]. Similar to C. sinensis, C. militaris shows a variety of biological activities, including anti-inflammatory, antioxidant, anticancer, and antidiabetic properties. The tonic constituents of C. militaris have been studied comprehensively, of which, cordycepin (3′-deoxyadenosine) is the most potent active compound. Structurally, cordycepin (C10H13N5O3, 251.24 g/mol) is an adenosine analog, in which the oxygen bound to the carbon at position 3 of adenosine is deprived (Figure 1). Previous studies indicated numerous pharmacological properties of cordycepin comprising anti-tumor, induction of apoptosis of various human cancer cell lines, anti-malaria, anti-fungal, anti-bacterial, and anti-aging [7]. Among a wide range of biological activities, cordycepin has a particularly prominent activity against cancer. The compound has attracted cancerologists because of its highly selective toxicity to cancer cell lines [8].
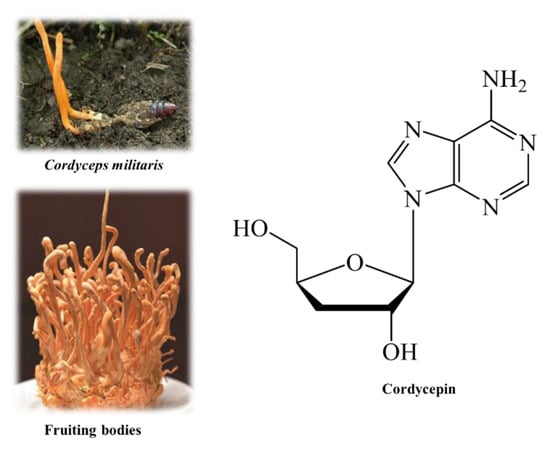
Figure 1.
Cordyceps militaris, fruiting bodies, and cordycepin.
Though possessing valuable health benefits, the isolation and purification of cordycepin from biomass as well as in cultural media has never been simple so far. Presently, most of the protocols have reported complicated processes, which might be costly and laborious. Though a variety of approaches for cordycepin synthesis have been reported, some shortcomings are inevitable [9], such as applicability and feasibility on larger scales of production compared to the laboratory scale. Therefore, cordycepin derived from natural sources has remained an important position in the current demand. As a result, simplified isolation and purification methods are required to shorten the time and cost of production of cordycepin and cordycepin-included purities from the Cordyceps fungi.
On the other hand, kombucha, a fermented functional beverage, has rapidly gained popularity since 1990 due to increasing health demands. In 2019, its market size in the world exceeded 1.8 billion dollars [10]. Many studies have confirmed the biological activities of kombucha products as antioxidant, antibacterial, antifungal, hepatoprotective, and anticancer [11,12,13]. Along with traditional black and green teas, most kombucha products have been made from popular food materials, including edible herbs, fruits, and soybeans [14,15]. To date, there has been no study on kombucha fermentation using Cordyceps species as a raw material or substrate.
This research aims to simplify and optimize the isolation and purification of cordycepin from the C. militaris fruiting body. In addition, the biological activities comprising antioxidant, anti-α-glucosidase, and cytotoxicity on Meg-01 and HL-60 cell lines of kombucha fermented C. militaris extracts including cordycepin have been investigated.
2. Materials and Methods
2.1. Chemicals and Instrumentations
Extraction solvents including methanol, hexane, chloroform, ethyl acetate, and butanol were gained from Junsei Chemical Co., Ltd., Tokyo, Japan. α-Glucosidase from Saccharomyces cerevisiae, p-nitrophenyl-α-D-glucopyranoside (pNPG), constituents for potassium phosphate buffer, α-amylase from porcine pancreas, soluble starch, DMSO, vanillin and silica gel, were purchased from Sigma-Aldrich, St. Louis, MO, USA. The 2,2-Diphenyl-1-picrylhydrazine (DPPH) and components for phosphate saline buffer and acetate buffer were obtained from Kanto Chemical Co., Inc., Tokyo, Japan. Sucrose, iodine solution, hydrochloric acid, and HPLC grade methanol were supplied by FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan. HPLC grade water and acetonitrile were acquired from NACALAI TESQUE, INC, Kyoto, Japan and Thermo Fisher Scientific Co., Ltd., Osaka, Japan, respectively. Cordycepin standard was provided by Cayman chemical, Ann Arbort, MI, USA.
The vacuum evaporator (Rotavapor®® R-300) was purchased from Nihon Buchi K.K., Tokyo, Japan. Freeze dryer (VD-250R) was procured from TAITEC Corporation, Saitama, Japan. Microplate reader (MultiskanTM Microplate Spectrophotometer) was obtained from Thermo Fisher Scientifific, Osaka, Japan. Thin-layer chromatography (TLC) plates (pore size 60 Å, layer thickness 140–160 µm) was gained from Merck, Darmstadt, Germany. HPLC system comprising a PU-4180 RHPLC pump, LC-Net II/ADC controller, and UV-4075 UV/VIS detector was supplied by Jasco Corporation, Tokyo, Japan. Bruker Ascend 400 MHz NMR spectrometer was from BRUKER BioSpin, Faellanden, Switzerland. The growth chamber (NKsysem BIOTRON) was provided by Nippon Medical & Chemical Instruments Co., Ltd., Osaka, Japan. The fume hood KFS 150–60 Hz was procured from Yamato Scientific Co., Ltd., Tokyo, Japan.
2.2. Cordyceps Millitaris Sample
C. militaris fruiting bodies were kindly provided by Truc Anh Company, Bac Lieu city, Vietnam. After harvesting, the fruiting bodies were subjected to a freeze-drying process in a lyophilizer (Mactech MSL1000, Mactech, Hanoi, Vietnam) in Hanoi, Vietnam. The dried samples were then sterilized, shipped in airtight containers, and stored at 4 °C until analysis.
2.3. Sample Preparation and Extraction
The dry C. militaris (2.8 kg) was pulverized into fine powder and then immersed in 6 L of pure methanol (MeOH). After 3 days, the extract was collected, filtered by filter paper, and evaporated at 50 °C by an evaporator. Afterward, the obtained recycled MeOH was used to re-extract the sample residue for other three extraction cycles. The methanolic extracts were combined and concentrated to yield 383.19 g (Figure 2).
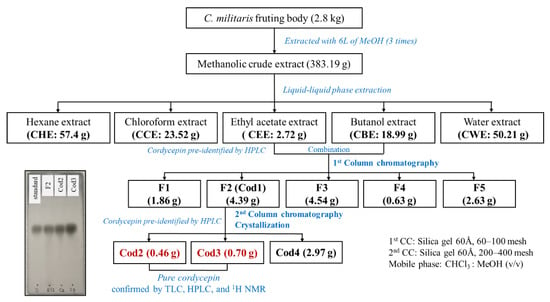
Figure 2.
Extraction and isolation processes of cordycepin from Cordyceps militaris.
The crude methanolic extract was then homogenized in 500 mL of distilled water and subjected to a 1 L separatory funnel. The liquid-liquid phase extraction was carried out by using in turn hexane, chloroform, ethyl acetate, butanol, and water, respectively. For each extraction phase, the corresponding solvent was mixed well with the aqueous crude extract at a ratio of 1:1 (v/v). After vigorous shaking, the mixture was stabilized until the separation appeared between liquid phases owing to their density differences. The organic phase was then collected, and the residue (aqueous phase) was used for the next extraction. Three replications (3 × 500 mL) were conducted for each phase of the liquid-liquid phase extraction. Subsequently, the resulting extracts were separately combined according to the solvent type, which was then filtered and evaporated. Consequently, 57.4 g of hexane extract (CHE), 23.52 g of chloroform extract (CCE), 2.72 g of ethyl acetate extract (CEE), 18.99 of butanol extract (CBE), and 50.21 g of water extract (CWE) were obtained (Figure 2).
2.4. Fractionation, Isolation, Purification, and Crystalization of Cordycepin
The preliminary tests used thin-layer chromatography (TLC) and high-performance liquid chromatography [16] indicated that CEE and CBE extracts contained cordycepin, therefore, these extracts were combined and subjected to a column chromatography. Initially, CEE and CBE extracts were loaded into a normal phase column chromatography that was filled with 200 g of silica gel (60 Å pore size, 60–100 mesh; Sigma-Aldrich, Tokyo, Japan) and the base solvent of chloroform. The column chromatography attempted to fractionate from low-polar compounds to high-polar compounds by gradually increasing the polarity of the eluent with an increasing proportion of MeOH. Each fraction was collected from 100 mL of elute. The component of elution is described in Figure S1.
As shown in Figure S1b, five fractions were collected after recombining 150 initial fractions collected from the first column chromatography (Figure S1a). The re-combination was based on the result of TLC and HPLC tests. Among five fractions, only F2 (Cod1) contained cordycepin, therefore, this fraction was chosen and subjected the second column chromatography. Accordingly, F2 was successively separated in a normal phase column with 30 g of silica gel (60 Å pore size, 60–100 mesh; Sigma-Aldrich, Tokyo, Japan). The eluate was 20 mL per fraction. The obtained fractions and corresponding elution were E1–E5 (C:M, 98:2), E5–E10 (C:M, 95:5), E11–E25 (C:M, 90:10), and E26–E30 (C:M, 80:20). Subsequently, TLC and HPLC tests displayed that the combinations of E13-E17 (Cod2) and E18-E28 (Cod3) contained the purest cordycepin.
In the next step, Cod2 and Cod3 were diluted by chloroform and MeOH (1:1, v/v) prior to being filtered by filter papers (Advantec No. 2, Toyo Kaisha, Ltd., Tokyo, Japan). The resulting filtrates were separately evaporated under vacuum at 40 °C by an evaporator. The dry extracts were then transferred to a 100 mL beaker and dissolved in 50 mL of ethyl acetate and MeOH (3:7, v/v). Afterward, the beaker was wrapped with perforated aluminum foil with small holes on the surface. Subsequently, wrapped beaker was placed in a fume hood, and the evaporation was conducted under room temperature (25 ± 1 °C). After 24 h, the pale-yellow crystals appeared and were filtered with several elutions by the solvent containing chloroform, ethyl acetate, and MeOH (6:2:2, v/v/v). Consequently, 0.46 and 0.70 g of pure crystals were obtained from Cod2 and Cod3, respectively. The filtrate was evaporated to yield a residue (Cod4, 2.97 g), see Figure 2 and Figure 3.
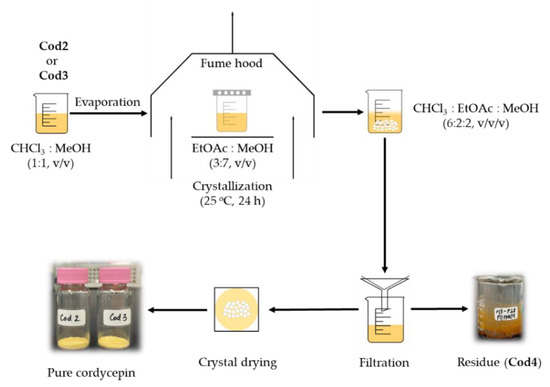
Figure 3.
Crystallization and purification of cordycepin from fractions Cod2 and Cod3.
2.5. Thin-Layer Chromatography and Spectroscopic Analyses
Fractions obtained from column chromatography were separately analyzed by TLC, of which, the stationary phase was TLC Silica gel 60 RP-18 F254S plate (Merck KGaA, Darmstadt, Germany) and the mobile phase was the mixture of CHCl3 and MeOH (v/v). Briefly, samples were spotted on the TLC plate and placed into a chamber with corresponding mobile solvent. After drying, the separation of constituents was observed by a UV lamp under wavelengths of 254 and 360 nm. Besides, the compound separation on TLC plate was determined by a staining-heated method, in which, 1% vanillin-sulfuric acid in pure ethanol was the stain and 100 °C in 2 min was the heating condition [17].
Identification and quantification of cordycepin from samples were conducted by an HPLC method described by Quy et al. [16] with some modifications. Accordingly, the mobile program was adjusted as follows: 0–10 min, 100 % solvent A (10% acetonitrile); 10–12 min, linearly increased to 100% solvent B (100% acetonitrile); 12–20 min, 100% solvent B; 20–22 min, returned to initial condition (100% solvent A); 22–30 min, re-equilibrated the column with 100% solvent A. Cordycepin was detected at 260 nm. Other conditions were maintained as described in the previous study [16]. Limits of detection (LOD) and limits of quantitation (LOQ) for HPLC measurement of cordycepin were calculated from the linear regression analysis of peak areas against concentrations of the standard cordycepin. Quantity of cordycepin in samples was estimated based on the linear equation achieved from standard curve of authentic cordycepin.
Pure crystals from Cod2 and Cod3 were dissolved in DMSO-d6 before analyzed by 1H NMR at 400 MHz. The obtained spectra were analysis by Bruker Topspin software version 3.2 and compared with reported data [18,19,20].
2.6. Kombucha Fermentation
Samples including fraction F2 (Cod1), isolated cordycepin from Cod2 and Cod3 (Cod23), and black tea (BT) were used for Kombucha fermentation. The standard method [21] used a starter KIT (World Kombucha, Bellingham, WA, USA) applied in this study. Initially, 40 g of sample were boiled with 100 mL of 5% sucrose solution for 10 min. After cooling to room temperature (25 ± 1 °C), 20 g of Symbiotic Culture of Bacteria and Yeast (SCOBY) were added to the solution. The container was then covered by a paper cloth for air circulation and prevention of inflow of external pollutants. The fermentation was carried out in a growth chamber at 30 °C in the darkness for 5 days.
The fermented liquid was extracted by adding 80% ethanol (1:1, v/v) and the mixture was vigorously shaken and centrifuged at 10.000 rpm for 10 min at 4 °C. The supernatant was collected and filtered before evaporation. In the next step, the dry fermented sample was dissolved in 20 mL of ethanol (98%) and homogenized in a bath at 60 °C for 2 min. The solution was then cooled down until sucrose precipitated. The liquid phase was carefully separated from the mixture and filtered through a 1.2 μm filter disc (Sartorius Stedim Biotech, Goettingen, Germany). The removal of sucrose was replicated five times. Finally, the extracts for each sample were combined and dry with a freeze dryer. The resulting dry extracts were stored in screw cap vials in darkness at 4 °C for further analyses.
2.7. Biological Acitivities
2.7.1. Antioxidant
The DPPH free radical-scavenging activity of samples was investigated by the method described by Quan et al. [17]. As a specific experimental procedure, 40 µL of sample (dissolved in MeOH), 40 µL of DPPH solution (0.2 mg/mL in MeOH), and 100 µL of 0.1 M acetic acid buffer (pH = 5.5) were homogenized. Subsequently, the reaction solution was incubated at room temperature in the darkness for 15 min. Radical scavenging ability was explored by measuring absorbance of the solution at 517 nm by a microplate reader. Methanol instead of sample was used as a negative control. Results are presented as IC50 values, which represent the concentration at which 50% of the DPPH radical is eliminated. A lower value indicates a stronger DPPH radical scavenging ability.
The inhibition percentage was calculated as follows:
where Ac is the absorbance of the negative control (MeOH), As is the absorbance of the sample, and Acb and Asb are the absorbance of the blank solution (without DPPH) of control and sample, respectively.
DPPH radical-scavenging activity (%) = [(Ac − Acb) − (As − Asb)]/(Ac − Acb) × 100
2.7.2. α-Glucosidase Inhibitory Assay
The inhibitory effect of samples on α-glucosidase was evaluated by the previous method [22]. First, 40 μL of buffer (0.1 M potassium phosphate, pH = 7.0) were mixed with 20 μL of sample (in DMSO). Subsequently, 20 μL of α-glucosidase solution (0.5 units/mL in buffer) were slightly mixed with the above sample solution, and preincubated for 5 min at 25 °C. Afterward, 20 μL of substrate solution (5 mM pNPG in buffer) was supplied and the mixture was incubated under the same conditions for 10 min. Finally, 100 μL of 0.1 M sodium carbonate solution was pipetted to stop the reaction. DMSO instead of the sample was used as a negative control. The reaction solution was detected at a wavelength of 405 nm and the absorbance was determined. The α-glucosidase inhibition percentage was calculated by the following formula:
where A sample is absorbance of samples and A control is absorbance of negative control. The results of anti-α-glucosidase activities are expressed as IC50 values.
Inhibition percentage (%) = (1 − A sample/A control) × 100
2.7.3. Cytotoxic Activity against HL-60 and Meg-01 Cell Lines
MTT assay was used to examine the cytotoxicity of the sample on cancerous cells, which was carried out following the method described by Anh et al. [23]. Iscove’s Modified Dulbecco’s Medium (IMDM) contains fetal bovine serum (10%), L-glutamine (5 mM), penicillin (100 IU/mL), and streptomycin (100 μg/mL). Samples were prepared in 0.1% DMSO in IMDM. Cells were seeded at a density of 5 × 103 in 90 μL on cell culture medium and grown in an incubator (Thermo Fisher Scientific, Waltham, MA, USA) at 37 °C with 5% CO2. After 24 h, 10 µL of sample was pipetted into the cell culture. After 72 h of incubation, the cells were incubated with MTT solution (5 mg/mL) for another 4 h in the same conditions. The cells were then supplemented with cell lysate (10% SDS in 0.01 M HCl), and cell growth was checked by an inverted microscope (LabX, Midland, ON, Canada). Finally, the cell viability was measured by the absorbance recorded at 595 nm. Cell culture medium instead of sample was used as a negative control. The cytotoxic effect of sample on blood cancer cells was determined as follows:
where Abs control is an absorbance of the reaction with negative control, and Abs sample is an absorbance of the reaction with sample.
Inhibition (%) = (Abs control − Abs sample)/Abs control × 100
2.8. Statistical Analysis
Minitab software version 16.0 (Minitab Inc., State College, PA, USA) was used for statistical analyses in this study. Data are expressed as mean ± standard deviation (n = 3). One-way ANOVA using Tukey’s test at p < 0.05 was used to determine significant differences among tested samples.
3. Results
3.1. Isolation and Purification of Cordycepin
3.1.1. Identification of Cordycepin by HPLC
The HPLC chromatogram of crystals purified from Cod2 and Cod3 and the standard cordycepin demonstrates that Cod2 and Cod3 contained highly purified cordycepin (Figure 4).
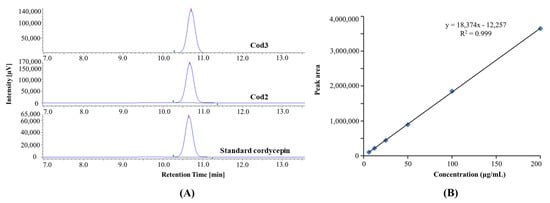
Figure 4.
HPLC chromatogram of purified and standard cordycepin (A), and linear curve of standard cordycepin (B).
Besides, the linearity of the standard curve for cordycepin was established with a correlation coefficient (r2) of 0.9992. The limits of detection (LOD) and quantification (LOQ) were calculated as 0.04 and 0.12 µg/mL, respectively.
3.1.2. Confirmation of Cordycepin by 1H NMR
After pre-identifying by HPLC, crystals of Cod23 were analyzed and confirmed by 1H NMR. The obtained NMR data are as follows: 1H NMR (400 MHz, DMSO-d6) δ 8.35 (s, 1H), 8.14 (s, 1H), 7.28 (s, 2H), 5.86 (d, J = 2.4 Hz, 1H), 5.66 (d, J = 4.3 Hz, 1H), 5.16 (t, J = 5.6 Hz, 1H), 4.61–4.52 (m, 1H), 4.35 (tt, J = 6.9, 3.6 Hz, 1H), 3.69 (ddd, J = 12.0, 50, 3.4 Hz, 1H), 3.51 (ddd, J = 11.9, 6.0, 4.0 Hz, 1H), 2.30–2.18 (m, 1H), 1.92 (ddd, J = 13.0, 6.4, 3.3 Hz, 1H). The comparison of the NMR data with those reported by previous studies [18,19,20] confirmed that Cod23 is cordycepin. The NMR spectrum of Cod23 is presented in Figure 5.
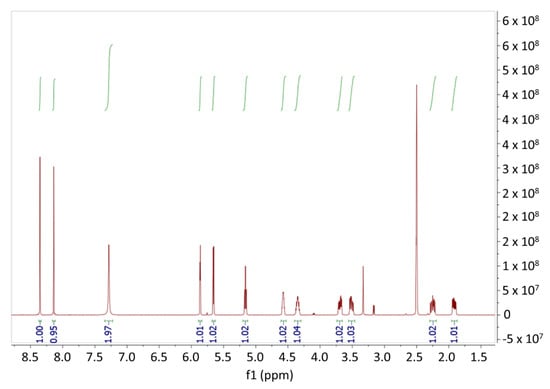
Figure 5.
1H-NMR Spectrum of Cod23.
3.1.3. Isolation and Purification Efficiency of Cordycepin
By HPLC, cordycepin is determined and quantified with an estimated amount of 0.50 g/kg, which is equal to 1.39 g per total dry weight of the sample. On the other hand, by dual-normal phase column chromatography, cordycepin is isolated and purified with an amount of 1.15 g. Therefore, the column chromatography is highly optimal with an efficiency of 82.73 % in a comparison with the estimated quantity by HPLC. Besides, the isolation yield is 0.04 %.
3.2. Biological Activities
Biological activities including antioxidant, anti-α-glucosidase, and cytotoxicity of cordycepin-included fraction (Cod1) and purified cordycepin (Cod23) before and after kombucha fermentation are presented as follows:
3.2.1. Antioxidant Activity
The DPPH assay reveals that BT (IC50 = 0.06 mg/mL) and Cod1 (IC50 = 0.88 mg/mL) exhibit strong radical scavenging activity, while Cod23 shows no effect. However, after kombucha fermentation, Cod23 shows the most potent antioxidant ability (IC50 = 4.76 mg/mL) compared to Cod1 (IC50 = 9.96 mg/mL) and BT (IC50 = 11.60 mg/mL) (Table 1).

Table 1.
Effect of kombucha fermentation on antioxidant activity.
3.2.2. α-Glucosidase Inhibitory Activity
In terms of the α-glucosidase assay, all samples have no effect on the enzymatic reaction at the maximum tested concentration of 10 mg/mL. However, remarkably, after kombucha fermentation, samples show potential inhibition of α-glucosidase (Table 2).
Table 2.
Effect of kombucha fermentation on α-glucosidase inhibitory activity.
In particular, Cod1 exerts the strongest activity with an IC50 value of 5.66 mg/mL, whereas Cod23 and BT showed negligible effects on α-glucosidase, of which Cod23 and BT present 46.17% and 22.15% inhibitions, respectively, at a concentration of 10 mg/mL.
3.2.3. Cytotoxic Activity
The cytotoxicity of tested samples on Meg-01 is maximal at a concentration of 50 μg/mL; therefore, it is expressed as an inhibition percentage (Table 3). Of these, Cod1 (37.89%) and Cod23 (30.60%) perform significant cytotoxicity on the Meg-01 cell line. Nevertheless, the cytotoxicity decreased 2.54 and 3.95-fold for Cod1 and Cod23, respectively, after kombucha fermentation. Besides, the inhibitory activity of BT on Meg-01 growth slightly increases after fermentation, but still has a weak effect (8.64% at 50 μg/mL) (Table 3).
Table 3.
Effect of kombucha fermentation on cytotoxic activity.
On the other hand, both Cod1 and Cod23 exhibit potent cytotoxicity on the HL-60 cell line, with IC50 values of 12.50 and 14.99 μg/mL, respectively. Nevertheless, the effect significantly declines after kombucha fermentation. While fermented Cod1 shows a weak activity (a maximum inhibition of 39.64% at a concentration of 250 μg/mL), fermented Cod23 decreases its cytotoxicity on HL-60 growth by 21.38-fold (IC50 = 320.49 μg/mL) (Table 1). In the case of the BT sample, the maximum inhibition percentage is recorded at a concentration of 250 μg/mL. Accordingly, 51.13% and 11.32% are the inhibition percentages of BT and fermented BT, respectively.
3.3. Effect of Kombucha Fermentation on Cordycepin Content
Kombucha fermentation causes a significant reduction in the content of cordycepin. The quantification based on HPLC indicates that the cordycepin content in Cod1 and Cod23 samples decreased 57.32 and 33.55-fold, respectively, after kombucha fermentation (Table 4).
Table 4.
Change of cordycepin quantity in samples after kombucha fermentation.
4. Discussion
To date, there have been a number of studies on the isolation and purification of cordycepin from every part of the fungus, including mycelia, fruiting bodies, and cultural medium. However, reported protocols seem not to be proportionate to the pharmaceutical values of cordycepin since most isolation and purification procedures are not feasible in industrial and commercial production. After the discovery of cordycepin from the liquid medium of C. militaris in 1951 [24], the isolation methods of cordycepin from the liquid medium of C. militaris by ion column chromatography using Dowex-I (Cl) and Dowex-50 (NH4+) were reported in 1960 and 1965 [25,26]. In later studies, Ahn et al. developed a purification method consisting of column chromatography using Amberlite XAD-2 resin and silica gel (230–400 mesh) followed by preparatory HPLC and two-step column chromatography [8]; however, this procedure was hampered by its low yield. Recently, researchers proposed several advanced techniques for a high-purity and high-yield isolation and purification method to achieve commercial use of cordycepin [6,20,27,28]. Among the reports, the adsorption of cordycepin using macroporous resin is widely used [29], and this method has been incorporated into large-scale purification processes. Nevertheless, most of the reported methods require complicated steps, special instruments, and multi-step solvent systems, which lead to the isolation and purification of cordycepin from C. militaris becoming inaccessible. In this study, we introduced a simple protocol to isolate and purify cordycepin with high purity and an affordable yield. The method consists only of dual-normal phase column chromatography using silica gel, popular solvent systems, and crystallization under room temperature. To the best of our knowledge, there has been no report on the isolation of cordycepin by normal phase column chromatography and crystallization alone.
Table 5 displays some recent column chromatographic methods that are commonly used for the isolation and purification of cordycepin. Accordingly, with a simpler protocol, cordycepin was purified with a comparative yield of 0.04% of the total 2.8 kg of C. militaris fruiting bodies in the present study. Whereas, though the isolation yield is higher, the previously reported procedures are more complicated, time-consuming, and costly. For instance, molecularly imprinted polymer (MIP) chromatography requires a specific synthetic procedure for MIPs column, which is not easily accessible. In fact, column chromatography over silica gel is not a novelty to isolate cordycepin from C. militaris. However, this study simplified the isolation procedure in order to satisfy cost and time, which are the prerequisites in industrial production. Although it needs more practical models to confirm the feasibility of the present method, the information about chromatographic conditions and solvent systems in this study is useful for the development, adjustment, and improvement of larger-scale purification models.
Table 5.
Recent column chromatographic methods for isolation and purification of cordycepin from Cordyceps militaris.
The present study is the first to investigate the potential bioactivities of kombucha made from cordycepin and C. militaris extracts. Results suggested that kombucha fermentation significantly increased the antioxidant and anti-α-glucosidase activities of purified cordycepin (Cod23). Previous studies have shown that SCOBY produces antioxidants such as vitamins, organic acids, and phenolic substances during kombucha fermentation [33]. There is a high possibility that the post-fermentation fractions of C. militaris may contain SCOBY-derived antioxidants or the presence of newly produced active compounds. Remarkably, all samples exhibited potential inhibition on α-glucosidase after kombucha fermentation but did not show any effect before the fermentation. The pH changes and the actions of enzymes from SCOBY during the fermentation might be the major factors that could create more bioactive compounds. In kombucha, the pH is lowered by acetic acid produced by acetic acid bacteria, which degrades polyphenol complexes to generate more phenolics [14,34]. Xia et al. revealed that there was a strong correlation between total phenolic content and α-glucosidase inhibitory activity in soymilk-based kombucha [35]. Phenolic compounds inhibit the activity of α-glucosidase owing to its chelating capacity, causing enzymes to lose their biological activities through structural alteration [36]. Furthermore, it was reported that diabetic mice orally taking kombucha for 30 days lowered blood glucose levels and slowed the absorption of HLD-cholesterol and triglycerides [12]. However, this study has not yet identified the substances responsible for α-glucosidase inhibition in kombucha fermented products. Hence, further assays should be performed to identify bioactive compounds and to confirm the antidiabetic potential of C. militaris kombucha.
In addition, Cod1 and Cod23 exhibited the strongest cytotoxicity on HL-60 and Meg-01 cell lines, while fermented Cod1 and Cod23 showed a decrease in the activity, which was in line with the same decline of cordycepin content involved in samples before and after fermentation. The amount of cordycepin had a significant effect on cytotoxicity, which is consistent with previous results [37]. Hot water extracts of C. militaris fruiting bodies with higher levels of cordycepin showed not only stronger cytotoxicity against human colorectal HCT116, prostatic PC-4, hepatic Huh7, and leukemic HL-60 cancer cell lines, but also induced apoptosis on those cell lines, compared to hot water extract of mycelia [37]. On the other hand, Cod23 inhibited Meg-01 cells by 30.60% at a concentration of 50 μg/mL, but there has been no investigation into the cytotoxicity of cordycepin on the Meg-01 cell line so far. Cytotoxic results suggest that cordycepin is more potent against HL-60 cells than Meg 01 cells. However, earlier studies reported that cordycepin is sensitive to the digestive systems of organisms [38], of which, experiments indicated that cordycepin was greatly affected by the enzyme system of SCOBY. For that reason, one of the most potential solutions is nanotechnology. In which, nanoparticles could cover active ingredients within secondary materials, which might help protect cordycepin from the hydrolytic enzyme system of SCOBY. Therefore, for future study, nanotechnology should be applied to enhance the bioaccessibility of cordycepin during kombucha fermentation.
5. Conclusions
This study provides a simple procedure for isolating and purifying cordycepin from the fruiting bodies of Cordyceps militaris. The simplified dual-normal phase column chromatography using silica gel, filtration, and typical crystallization in the present study is expected to be prospective for the development of a feasible production model of cordycepin on larger scales. Additionally, for the first time, we investigated the potential pharmacological activities of kombucha made from C. militaris-derived bioactive components. The results suggested that purified cordycepin and its kombucha fermented extracts could be potentially used for producing functional supplements to support the treatment of some chronic diseases in humans. Further in vivo studies and clinical tests should be carried out to evaluate kombucha products made from C. militaris and the effectiveness of cordycepin.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/separations9100290/s1, Figure S1: Elution composition and fractions from the first column chromatography. (a) Initial fractions; (b) new fractions after confirming by TLC and HPLC; C, chloroform; M, methanol.
Author Contributions
Conceptualization, N.V.Q. and T.D.X.; methodology, N.V.Q., Y.I., M.H. and L.H.A.; validation, N.V.Q. and T.D.X.; formal analysis, N.V.Q., M.H. and Y.I.; investigation, N.V.Q., Y.I., M.H. and L.H.A.; data curation, N.V.Q. and T.D.X.; writing—original draft preparation, N.V.Q. and Y.I.; writing—review and editing, N.V.Q., Y.I. and T.D.X.; visualization, N.V.Q. and T.D.X.; supervision, N.V.Q. and T.D.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Akiyoshi Takami and Vu Quang Lam for their support in screening cytotoxicity. Truc Anh Manufacturing and Trading Co., Ltd. (Bac Lieu province, Vietnam) is appreciated for providing fruiting bodies of Cordyceps militaris. Abe Manabu and Nguyen Hai Dang are appreciated for their assistance in operating and analyzing NMR spectra.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paterson, R.R.M. Cordyceps—A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef]
- Zhou, X.; Gong, Z.; Su, Y.; Lin, J.; Tang, K. Cordyceps fungi: Natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis part I. J. Altern. Complement. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An overview of its chemical constituents in relation to biological activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical composition and nutritional and medicinal value of fruit bodies and submerged cultured mycelia of culinary-medicinal higher Basidiomycetes mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Zhou, X.-H.; Li, H.-H.; Huang, W.-F. Column chromatographic extraction and preparation of cordycepin from Cordyceps militaris waster medium. J. Chromatogr. B 2009, 877, 2135–2141. [Google Scholar] [CrossRef]
- Tuli, H.S.; Kashyap, D.; Sharma, A.K. Cordycepin: A cordyceps metabolite with promising therapeutic potential. In Fungal Metabolites; Merillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2015; pp. 1–22. [Google Scholar]
- Ahn, Y.-J.; Park, S.-J.; Lee, S.-G.; Shin, S.-C.; Choi, D.-H. Cordycepin: Selective growth inhibitor derived from liquid culture of Cordyceps militaris against Clostridium spp. J. Agric. Food Chem. 2000, 48, 2744–2748. [Google Scholar] [CrossRef]
- Luo, Q.Y.; Cao, H.F.; Liu, S.K.; Wu, M.; Li, S.S.; Zhang, Z.Q.; Chen, A.J.; Shen, G.H.; Wu, H.J.; Li, M.L.; et al. Novel liquid fermentation medium of Cordyceps militaris and optimization of hydrothermal reflux extraction of cordycepin. J. Asian Nat. Prod. Res. 2020, 22, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Fortune Business Insights. Available online: https://www.fortunebusinessinsights.com/industry-reports/kombucha-market-100230 (accessed on 14 October 2021).
- Mousavi, S.M.; Hashemi, S.A.; Zarei, M.; Gholami, A.; Lai, C.W.; Chiang, W.H.; Omidifar, N.; Bahrani, S.; Mazraedoost, S. Recent progress in chemical composition, production, and pharmaceutical effects of kombucha beverage: A complementary and alternative medicine. Evid. Based Complement. Alternat. Med. 2020, 2020, 4397543. [Google Scholar] [CrossRef]
- Aloulou, A.; Hamden, K.; Elloumi, D.; Ali, M.B.; Hargafi, K.; Jaouadi, B.; Ayadi, F.; Elfeki, A.; Ammar, E. Hypoglycemic and antilipidemic properties of kombucha tea in alloxan-induced diabetic rats. BMC Complement. Altern. Med. 2012, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, C.; Farnworth, E. Tea, kombucha, and health: A review. Food Res. Int. 2000, 33, 409–421. [Google Scholar] [CrossRef]
- Coelho, R.M.D.; de Almeida, A.L.; do Amaral, R.Q.G.; da Mota, R.N.; de Sousa, P.H.M. Kombucha: Review. Int. J. Gastron. Food Sci. 2020, 22, 100272. [Google Scholar] [CrossRef]
- Shahbazi, H.; Gahruie, H.H.; Golmakani, M.-T.; Eskandari, M.H.; Movahedi, M. Effect of medicinal plant type and concentration on physicochemical, antioxidant, antimicrobial, and sensorial properties of kombucha. Food Sci. Nutr. 2018, 6, 2568–2577. [Google Scholar] [CrossRef]
- Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.-D.; Khanh, T.D.; Teschke, R. Cordycepin isolated from Cordyceps militaris: Its newly discovered herbicidal property and potential plant-based novel alternative to glyphosate. Molecules 2019, 24, 2901. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.-D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Andriana, Y.; Tuyen, P.T. Antioxidant, α-amylase and α-glucosidase inhibitory activities and potential constituents of Canarium tramdenum Bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I.; et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Hyun, S.H.; Sung, G.H.; Choi, H.K. Simple and rapid determination of cordycepin in Cordyceps militaris fruiting bodies by quantitative nuclear magnetic resonance spectroscopy. Anal. Lett. 2014, 47, 1031–1042. [Google Scholar] [CrossRef]
- Masuda, M.; Hatashita, M.; Fujihara, S.; Suzuki, Y.; Sakurai, A. Simple and efficient isolation of cordycepin from culture broth of a Cordyceps militaris mutant. J. Biosci. Bioeng. 2015, 120, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Tang, S.; Azi, F.; Hu, W.; Dong, M. Use of kombucha consortium to transform soy whey into a novel functional beverage. J. Funct. Foods 2019, 52, 81–89. [Google Scholar] [CrossRef]
- Quan, N.V.; Tran, H.-D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef]
- Anh, L.H.; Quan, N.V.; Lam, V.Q.; Iuchi, Y.; Takami, A.; Teschke, R.; Xuan, T.D. Antioxidant, anti-tyrosinase, anti-α-amylase, and cytotoxic potentials of the invasive weed Andropogon virginicus. Plants 2020, 10, 69. [Google Scholar] [CrossRef]
- Radhi, M.; Ashraf, S.; Lawrence, S.; Tranholm, A.A.; Wellham, P.A.D.; Hafeez, A.; Khamis, A.S.; Thomas, R.; McWilliams, D.; de Moor, C.H. A systematic review of the biological effects of cordycepin. Molecules 2021, 26, 5886. [Google Scholar] [CrossRef]
- Kredich, N.M.; Guarino, A.J. An improved method of isolation and determination of cordycepin. Biochim. Biophys. Acta 1960, 41, 363–365. [Google Scholar] [CrossRef]
- Frederiksen, S.; Malling, H.; Klenow, H. Isolation of 3′-deoxyadenosine (cordycepin) from the liquid medium of Cordyceps militaris (L. ex Fr.) Link. Biochim. Biophys. Acta 1965, 95, 189–193. [Google Scholar] [CrossRef]
- Zhou, X.; Cai, G.; He, Y.; Tong, G. Separation of cordycepin from Cordyceps militaris fermentation supernatant using preparative HPLC and evaluation of its antibacterial activity as an NAD+-dependent DNA ligase inhibitor. Exp. Ther. Med. 2016, 12, 1812–1816. [Google Scholar] [CrossRef]
- Zhang, Z.; Tudi, T.; Liu, Y.; Zhou, S.; Feng, N.; Yang, Y.; Tang, C.; Tang, Q.; Zhang, J. Preparative isolation of cordycepin, N6 -(2-hydroxyethyl)-adenosine and adenosine from Cordyceps militaris by macroporous resin and purification by recycling high-speed counter-current chromatography. J. Chromatogr. B 2016, 1033–1034, 218–225. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, L.; Zhang, Z.; Guan, Y.; Li, B.; Yang, J.; Fan, H.; Yang, G.; Chen, X.; Zhang, J.; et al. Enhanced cordycepin production in caterpillar medicinal mushroom, Cordyceps militaris (Ascomycetes), mutated by a multifunctional plasma mutagenesis system. Int. J. Med. Mushrooms 2020, 22, 1147–1159. [Google Scholar] [CrossRef]
- Zhu, L.; Liang, Y.; Lao, D.; Zhang, T.; Ito, Y. Preparative separation of high-purity cordycepin from Cordyceps militaris (L.) link by high-speed countercurrent chromatography. J. Liq. Chromatogr. Relat. Technol. 2011, 34, 491–499. [Google Scholar]
- Zhang, Y.; Wan, J.; Cao, X. Synthesis of surface molecularly imprinting polymers for cordycepin and its application in separating cordycepin. Process Biochem. 2016, 51, 517–527. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, J.; Choi, J.H.; Hirai, H.; Nishida, H.; Kawagishi, H. Cytotoxic compounds against cancer cells from Bombyx mori inoculated with Cordyceps militaris. Biosci. Biotechnol. Biochem. 2017, 81, 1224–1226. [Google Scholar] [CrossRef]
- Essawet, N.A.; Cvetkovic, D.; Velićanski, A.; Canadanovic-Brunet, J.; Vulić, J.; Maksimović, V.; Sinisa, M.; Sinisa, M. Polyphenols and antioxidant activities of Kombucha beverage enriched with Coffeeberry® extract. Chem. Ind. Chem. Eng. Q. 2015, 21, 399–409. [Google Scholar] [CrossRef]
- Gamboa-Gómez, C.I.; González-Laredo, R.F.; Gallegos-Infante, J.A.; Pérez, M.D.; Moreno-Jiménez, M.R.; Flores-Rueda, A.G.; Rocha-Guzmán, N.E. Antioxidant and angiotensin-converting enzyme inhibitory activity of Eucalyptus camaldulensis and Litsea glaucescens infusions fermented with Kombucha consortium. Food Technol. Biotechnol. 2016, 54, 367–374. [Google Scholar] [CrossRef]
- Xia, X.; Dai, Y.; Wu, H.; Liu, X.; Wang, Y.; Yin, L.; Wang, Z.; Li, X.; Zhou, J. Kombucha fermentation enhances the health-promoting properties of soymilk beverage. J. Funct. Foods 2019, 62, 103549. [Google Scholar] [CrossRef]
- Nagappan, H.; Pee, P.P.; Kee, S.H.Y.; Ow, J.T.; Yan, S.W.; Chew, L.Y.; Kong, K.W. Malaysian brown seaweeds Sargassum siliquosum and Sargassum polycystum: Low density lipoprotein (LDL) oxidation, angiotensin converting enzyme (ACE), α-amylase, and α-glucosidase inhibition activities. Food Res. Int. 2017, 99, 950–958. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yao, Q.M.; Zhou, J.; Chang, Y.; Li, J.L.; Wang, Y.Z.; Wu, H.P.; Chen, Y.H.; Liu, Y.R.; Huang, X.J.; et al. Synergistic antitumoral efficacy of a novel replicative adenovirus SG611-PDCD5 and daunorubicin in human leukemic cells. Onco. Targets Ther. 2018, 11, 5121–5132. [Google Scholar] [CrossRef]
- Wei, H.P.; Ye, X.L.; Chen, Z.; Zhong, Y.J.; Li, P.M.; Pu, S.C.; Li, X.G. Synthesis and pharmacokinetic evaluation of novel N-acyl-cordycepin derivatives with a normal alkyl chain. Eur. J. Med. Chem. 2009, 44, 665–669. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).