
Plasma Short-Chain Fatty Acids and Their Derivatives in Women with Gestational Diabetes Mellitus

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Stock and Standard Solution Preparation
2.3. Biological Samples
2.4. Sample Preparation
2.5. LC-MS/MS Conditions
2.6. Data Treatment and Statistical Analysis
3. Results
3.1. Analytical Characteristics of the LC-MS/MS Method
3.2. General Characteristics of the Participants
3.3. Differences in the Plasma SCFA and Their Derivatives
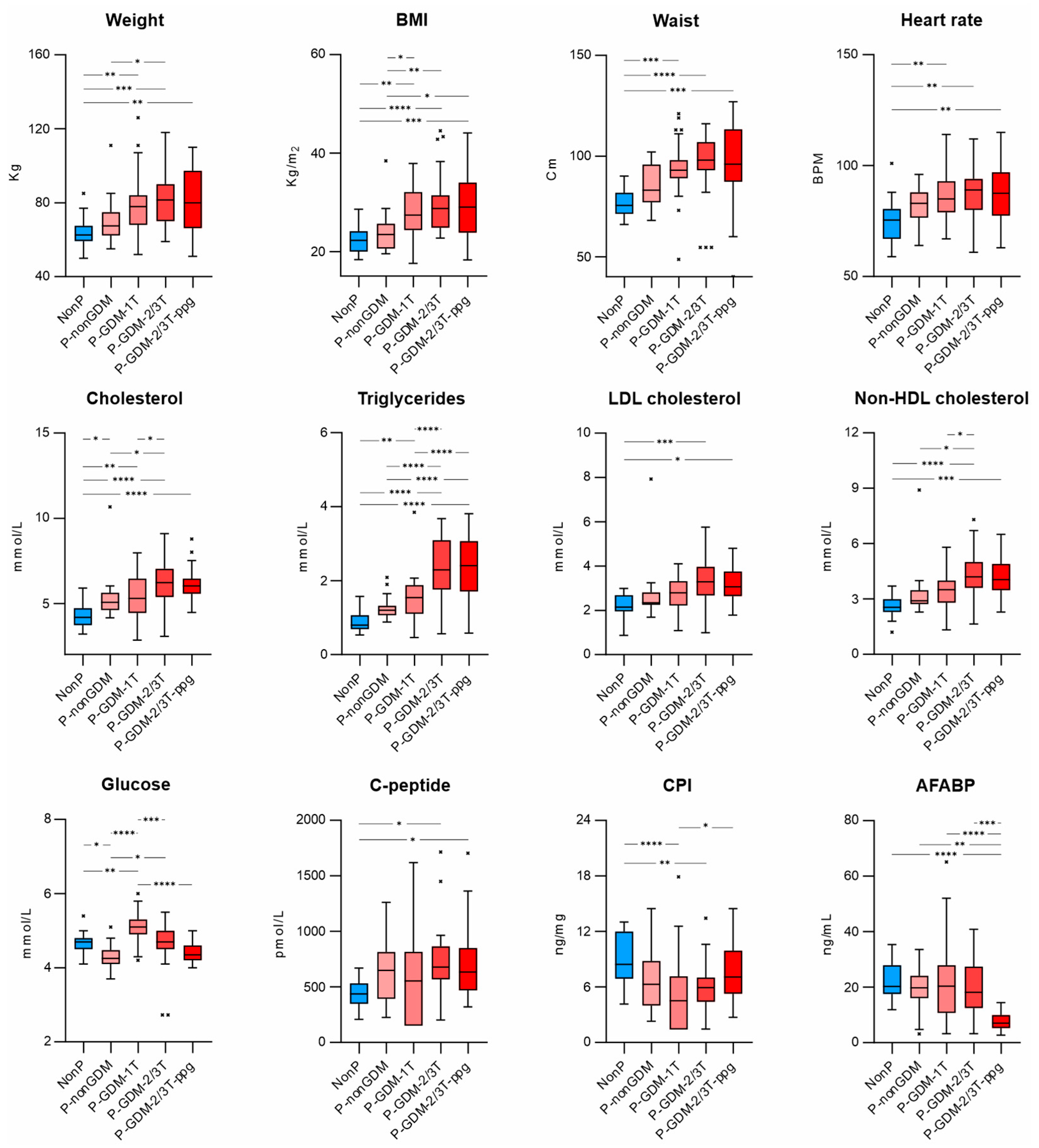
3.4. Differences in the Baseline Body Composition Characteristics and Biochemical Parameters in All Measurement Groups
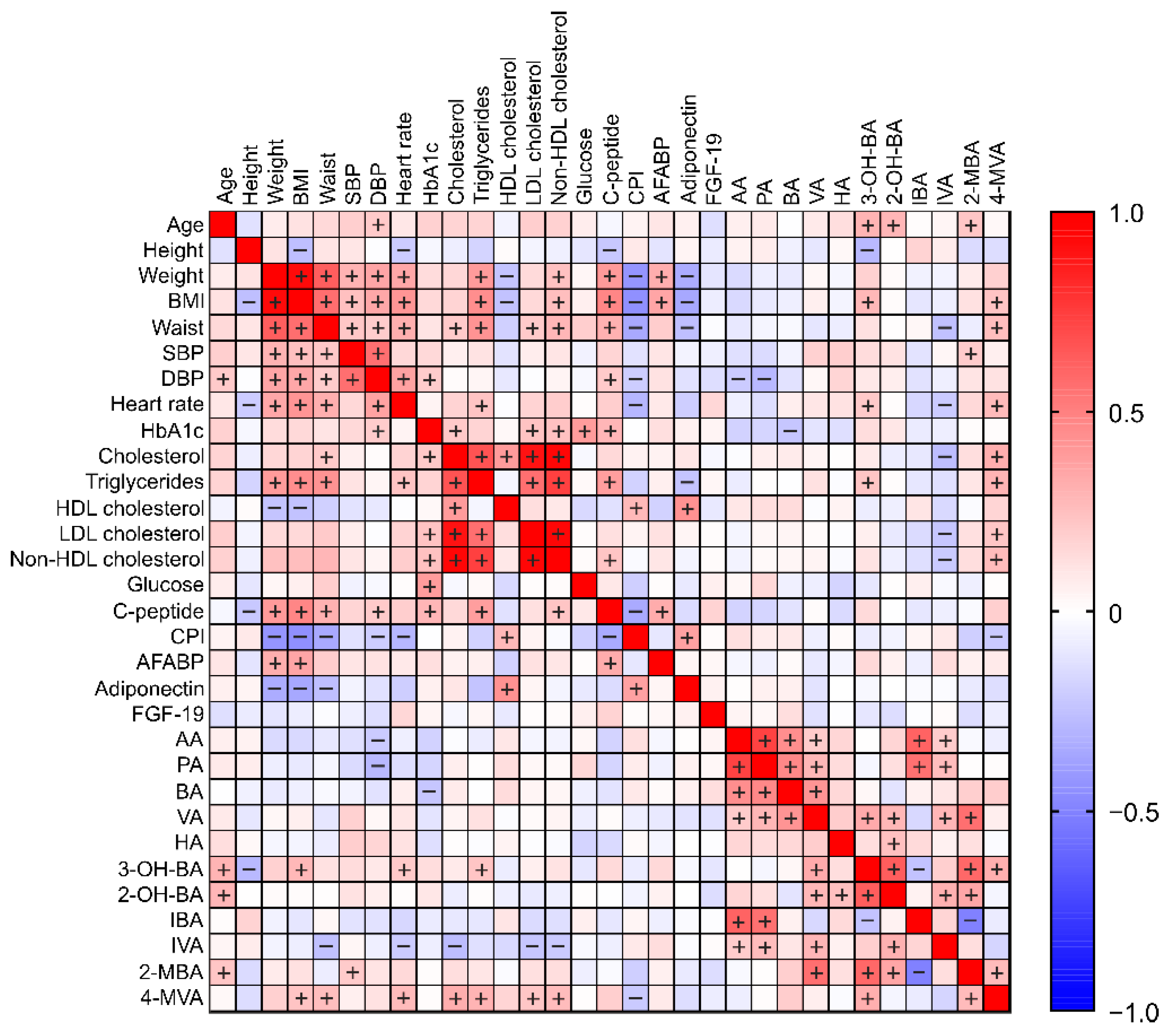
3.5. Pearson Correlations between All Measurement Parameters
3.5.1. Correlations between SCFA (and Their Derivatives) and Parameters of Baseline Body Composition Characteristics and Biochemical Markers
3.5.2. Correlations between SCFA and Their Derivatives
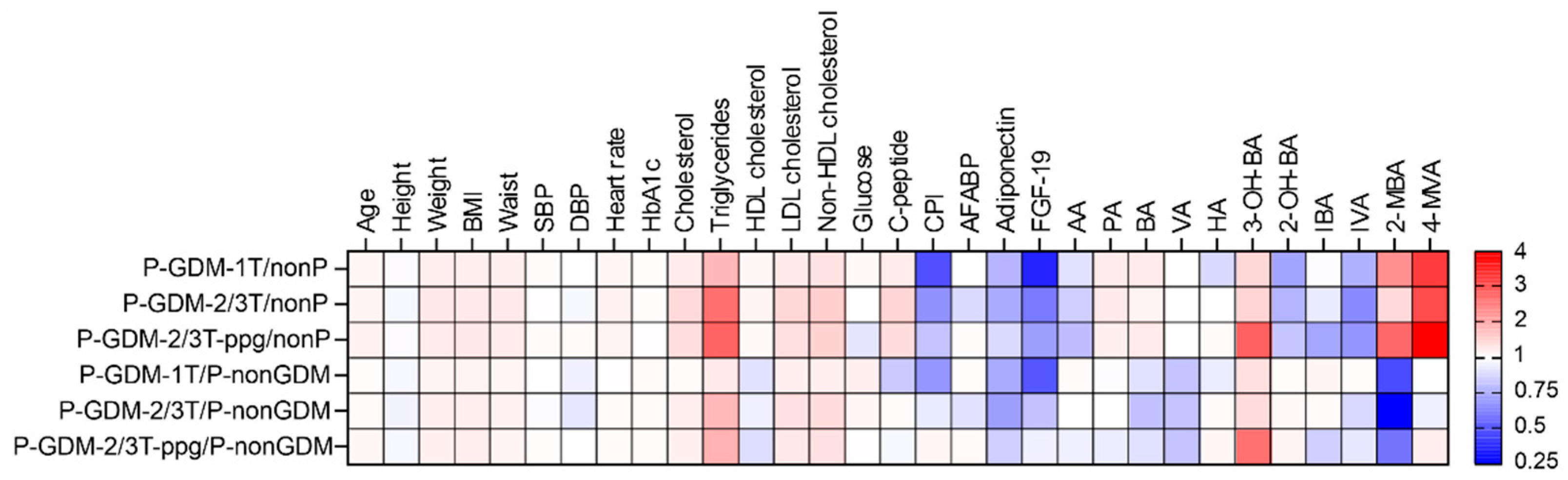
3.6. Biomarkers Associated with the Development of GDM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alesi, S.; Ghelani, D.; Rassie, K.; Mousa, A. Metabolomic Biomarkers in Gestational Diabetes Mellitus: A Review of the Evidence. Int. J. Mol. Sci. 2021, 22, 5512. [Google Scholar] [CrossRef]
- Metzger, B.E.; Gabbe, S.G.; Persson, B.; Buchanan, T.A.; Catalano, P.A.; Damm, P.; Dyer, A.R.; de Leiva, A.; Hod, M.; Kitzmiler, J.L.; et al. International Association of Diabetes and Pregnancy Study Groups Recommendations on the Diagnosis and Classification of Hyperglycemia in Pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Cosson, E.; Carbillon, L.; Valensi, P. High Fasting Plasma Glucose during Early Pregnancy: A Review about Early Gestational Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 8921712. [Google Scholar] [CrossRef] [PubMed]
- Immanuel, J.; Simmons, D.; Harreiter, J.; Desoye, G.; Corcoy, R.; Adelantado, J.M.; Devlieger, R.; Lapolla, A.; Dalfra, M.G.; Bertolotto, A.; et al. Metabolic Phenotypes of Early Gestational Diabetes Mellitus and Their Association with Adverse Pregnancy Outcomes. Diabet. Med. 2021, 38, e14413. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Pan, L.-L.; Lv, S.; Yang, Q.; Zhang, H.; Chen, W.; Lv, Z.; Sun, J. Alterations of Gut Microbiota and Blood Lipidome in Gestational Diabetes Mellitus With Hyperlipidemia. Front. Physiol. 2019, 10, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulven, T. Short-Chain Free Fatty Acid Receptors FFA2/GPR43 and FFA3/GPR41 as New Potential Therapeutic Targets. Front. Endocrinol. 2012, 3, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet–Induced Obesity via a PPARγ-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [Green Version]
- Nishina, P.M.; Freedland, R.A. Effects of Propionate on Lipid Biosynthesis in Isolated Rat Hepatocytes. J. Nutr. 1990, 120, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary Fiber and Bacterial SCFA Enhance Oral Tolerance and Protect against Food Allergy through Diverse Cellular Pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.; Engevik, M.A.; Spinler, J.K.; Versalovic, J. Healthy Human Gastrointestinal Microbiome: Composition and Function After a Decade of Exploration. Dig. Dis. Sci. 2020, 65, 695–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, M.; Cao, H. Fast Quantification of Short Chain Fatty Acids and Ketone Bodies by Liquid Chromatography-Tandem Mass Spectrometry after Facile Derivatization Coupled with Liquid-Liquid Extraction. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1083, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Ziętek, M.; Celewicz, Z.; Szczuko, M. Short-Chain Fatty Acids, Maternal Microbiota and Metabolism in Pregnancy. Nutrients 2021, 13, 1244. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of Short-Chain Fatty Acid Production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic Health: Fermentation and Short Chain Fatty Acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Jaochico, A.; Sangaraju, D.; Shahidi-Latham, S.K. A Rapid Derivatization Based LC-MS/MS Method for Quantitation of Short Chain Fatty Acids in Human Plasma and Urine. Bioanalysis 2019, 11, 741–753. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouteau, E.; Meirim, I.; Métairon, S.; Fay, L.B. Acetate, Propionate and Butyrate in Plasma: Determination of the Concentration and Isotopic Enrichment by Gas Chromatography/mass Spectrometry with Positive Chemical Ionization. J. Mass Spectrom. 2001, 36, 798–805. [Google Scholar] [CrossRef]
- Zalar, B.; Haslberger, A.; Peterlin, B. The Role of Microbiota in Depression—A Brief Review. Psychiatr. Danub. 2018, 30, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.C.; Verdin, E. Ketone Bodies as Signaling Metabolites. Trends Endocrinol. Metab. 2014, 25, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Harada, E.; Nakagawa, H.; Morikawa, Y.; Shono, M.; Kugimiya, F.; Yoshimura, M.; Yasue, H. The Diabetic Heart Utilizes Ketone Bodies as an Energy Source. Metabolism 2017, 77, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Soto-Mota, A.; Norwitz, N.G.; Clarke, K. Why a D-β-Hydroxybutyrate Monoester? Biochem. Soc. Trans. 2020, 48, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierziak, J.; Burgberger, M.; Wojtasik, W. 3-Hydroxybutyrate as a Metabolite and a Signal Molecule Regulating Processes of Living Organisms. Biomolecules 2021, 11, 402. [Google Scholar] [CrossRef]
- Møller, N. Ketone Body, 3-Hydroxybutyrate: Minor Metabolite—Major Medical Manifestations. J. Clin. Endocrinol. Metab. 2020, 105. [Google Scholar] [CrossRef]
- Spanou, L.; Dalakleidi, K.; Zarkogianni, K.; Papadimitriou, A.; Nikita, K.; Vasileiou, V.; Alevizaki, M.; Anastasiou, E. Ketonemia and Ketonuria in Gestational Diabetes Mellitus. Hormones 2015, 14, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Dudzik, D.; Zorawski, M.; Skotnicki, M.; Zarzycki, W.; García, A.; Angulo, S.; Paz Lorenzo, M.; Barbas, C.; Pilar Ramos, M. GC–MS Based Gestational Diabetes Mellitus Longitudinal Study: Identification of 2-and 3-Hydroxybutyrate as Potential Prognostic Biomarkers. J. Pharm. Biomed. Anal. 2017, 144, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Shafaei, A.; Vamathevan, V.; Pandohee, J.; Lawler, N.G.; Broadhurst, D.; Boyce, M.C. Sensitive and Quantitative Determination of Short-Chain Fatty Acids in Human Serum Using Liquid Chromatography Mass Spectrometry. Anal. Bioanal. Chem. 2021. [Google Scholar] [CrossRef]
- Gu, H.; Jasbi, P.; Patterson, J.; Jin, Y. Enhanced Detection of Short-Chain Fatty Acids Using Gas Chromatography Mass Spectrometry. Curr. Protoc. 2021, 1, e177. [Google Scholar] [CrossRef]
- Douny, C.; Dufourny, S.; Brose, F.; Verachtert, P.; Rondia, P.; Lebrun, S.; Marzorati, M.; Everaert, N.; Delcenserie, V.; Scippo, M.-L. Development of an Analytical Method to Detect Short-Chain Fatty Acids by SPME-GC–MS in Samples Coming from an in Vitro Gastrointestinal Model. J. Chromatogr. B 2019, 1124, 188–196. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Zhu, M.-J. A Sensitive GC/MS Detection Method for Analyzing Microbial Metabolites Short Chain Fatty Acids in Fecal and Serum Samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Song, H.E.; Lee, H.Y.; Kim, S.J.; Back, S.H.; Yoo, H.J. A Facile Profiling Method of Short Chain Fatty Acids Using Liquid Chromatography-Mass Spectrometry. Metabolites 2019, 9, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monleón, D.; Morales, J.M.; Barrasa, A.; López, J.A.; Vázquez, C.; Celda, B. Metabolite Profiling of Fecal Water Extracts from Human Colorectal Cancer. NMR Biomed. 2009, 22, 342–348. [Google Scholar] [CrossRef]
- Ndagijimana, M.; Laghi, L.; Vitali, B.; Placucci, G.; Brigidi, P.; Guerzoni, M.E. Effect of a Synbiotic Food Consumption on Human Gut Metabolic Profiles Evaluated by (1)H Nuclear Magnetic Resonance Spectroscopy. Int. J. Food Microbiol. 2009, 134, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, G.; Noor, S.O.; Ridgway, K.; Scovell, L.; Jamieson, C.; Johnson, I.T.; Colquhoun, I.J.; Kemsley, E.K.; Narbad, A. Metabolomics of Fecal Extracts Detects Altered Metabolic Activity of Gut Microbiota in Ulcerative Colitis and Irritable Bowel Syndrome. J. Proteome Res. 2011, 10, 4208–4218. [Google Scholar] [CrossRef]
- Jacobs, D.M.; Deltimple, N.; van Velzen, E.; van Dorsten, F.A.; Bingham, M.; Vaughan, E.E.; van Duynhoven, J. (1)H NMR Metabolite Profiling of Feces as a Tool to Assess the Impact of Nutrition on the Human Microbiome. NMR Biomed. 2008, 21, 615–626. [Google Scholar] [CrossRef]
- Garcia, A.; Olmo, B.; Lopez-Gonzalvez, A.; Cornejo, L.; Rupérez, F.J.; Barbas, C. Capillary Electrophoresis for Short Chain Organic Acids in Faeces Reference Values in a Mediterranean Elderly Population. J. Pharm. Biomed. Anal. 2008, 46, 356–361. [Google Scholar] [CrossRef]
- Jiang, R.; Jiao, Y.; Zhang, P.; Liu, Y.; Wang, X.; Huang, Y.; Zhang, Z.; Xu, F. Twin Derivatization Strategy for High-Coverage Quantification of Free Fatty Acids by Liquid Chromatography–Tandem Mass Spectrometry. Anal. Chem. 2017, 89, 12223–12230. [Google Scholar] [CrossRef]
- Weathington, B.L.; Cunningham, C.J.L.; Pittenger, D.J. Understanding Business Research: Weathington/Understanding Business Research; John Wiley & Sons: Nashville, TN, USA, 2012; ISBN 9781118134269. [Google Scholar]
- Pinto, J.; Almeida, L.M.; Martins, A.S.; Duarte, D.; Barros, A.S.; Galhano, E.; Pita, C.; Almeida, M.D.C.; Carreira, I.M.; Gil, A.M. Prediction of Gestational Diabetes through NMR Metabolomics of Maternal Blood. J. Proteome Res. 2015, 14, 2696–2706. [Google Scholar] [CrossRef] [PubMed]
- Andersson-Hall, U.; Gustavsson, C.; Pedersen, A.; Malmodin, D.; Joelsson, L.; Holmäng, A. Higher Concentrations of BCAAs and 3-HIB Are Associated with Insulin Resistance in the Transition from Gestational Diabetes to Type 2 Diabetes. J. Diabetes Res. 2018, 2018, 4207067. [Google Scholar] [CrossRef]
- Cibickova, L.; Schovanek, J.; Karasek, D. Changes in Serum Lipid Levels during Pregnancy in Women with Gestational Diabetes. A Narrative Review. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2021, 165, 8–12. [Google Scholar] [CrossRef]
- Antuna-Puente, B.; Faraj, M.; Karelis, A.D.; Garrel, D.; Prud’homme, D.; Rabasa-Lhoret, R.; Bastard, J.-P. HOMA or QUICKI: Is It Useful to Test the Reproducibility of Formulas? Diabetes Metab. 2008, 34, 294–296. [Google Scholar] [CrossRef]
- Funakoshi, S.; Fujimoto, S.; Hamasaki, A.; Fujiwara, H.; Fujita, Y.; Ikeda, K.; Takahara, S.; Nagashima, K.; Hosokawa, M.; Seino, Y.; et al. Utility of Indices Using C-Peptide Levels for Indication of Insulin Therapy to Achieve Good Glycemic Control in Japanese Patients with Type 2 Diabetes. J. Diabetes Investig. 2011, 2, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, P.; Shao, P.; Liu, H.; Li, W.; Wang, L.; Wang, J.; Zhang, S.; Leng, J.; Li, N.; Tian, H.; et al. C-Peptide Levels and the Risk of Diabetes and Pre-Diabetes among Chinese Women with Gestational Diabetes. J. Diabetes Complicat. 2017, 31, 1658–1662. [Google Scholar] [CrossRef]
- Somm, E.; Jornayvaz, F.R. Fibroblast Growth Factor 15/19: From Basic Functions to Therapeutic Perspectives. Endocr. Rev. 2018, 39, 960–989. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.C.; Coskun, T.; Rovira, A.R.I.; Schneider, M.A.; Raches, D.W.; Micanovic, R.; Bina, H.A.; Dunbar, J.D.; Kharitonenkov, A. Fundamentals of FGF19 & FGF21 Action in Vitro and in Vivo. PLoS ONE 2012, 7, e38438. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhu, W.; Li, J.; An, C.; Wang, Z. Serum Concentrations of Fibroblast Growth Factors 19 and 21 in Women with Gestational Diabetes Mellitus: Association with Insulin Resistance, Adiponectin, and Polycystic Ovary Syndrome History. PLoS ONE 2013, 8, e81190. [Google Scholar] [CrossRef]
- Rios-Covian, D.; González, S.; Nogacka, A.M.; Arboleya, S.; Salazar, N.; Gueimonde, M.; de Los Reyes-Gavilán, C.G. An Overview on Fecal Branched Short-Chain Fatty Acids Along Human Life and as Related With Body Mass Index: Associated Dietary and Anthropometric Factors. Front. Microbiol. 2020, 11, 973. [Google Scholar] [CrossRef] [PubMed]
- Farup, P.G.; Valeur, J. Changes in Faecal Short-Chain Fatty Acids after Weight-Loss Interventions in Subjects with Morbid Obesity. Nutrients 2020, 12, 802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanadchangsaeng, N.; Kitagawa, A.; Yamamoto, T.; Abe, H.; Tsuge, T. Identification, Biosynthesis, and Characterization of Polyhydroxyalkanoate Copolymer Consisting of 3-Hydroxybutyrate and 3-Hydroxy-4-Methylvalerate. Biomacromolecules 2009, 10, 2866–2874. [Google Scholar] [CrossRef]
- Lau, N.-S.; Chee, J.-Y.; Tsuge, T.; Sudesh, K. Biosynthesis and Mobilization of a Novel Polyhydroxyalkanoate Containing 3-Hydroxy-4-Methylvalerate Monomer Produced by Burkholderia Sp. USM (JCM15050). Bioresour. Technol. 2010, 101, 7916–7923. [Google Scholar] [CrossRef] [PubMed]
- Barbut, F.; Mastrantonio, P.; Delmée, M.; Brazier, J.; Kuijper, E.; Poxton, I. European Study Group on Clostridium difficile (ESGCD) Prospective Study of Clostridium Difficile Infections in Europe with Phenotypic and Genotypic Characterisation of the Isolates. Clin. Microbiol. Infect. 2007, 13, 1048–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, M.D.; Stabler, R.A.; Shetty, N.; Wren, B.W. The Continually Evolving Clostridium Difficile Species. Future Microbiol. 2012, 7, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Kotanko, P.; Carter, M.; Levin, N.W. Intestinal Bacterial Microflora—A Potential Source of Chronic Inflammation in Patients with Chronic Kidney Disease. Nephrol. Dial. Transplant. 2006, 21, 2057–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, K.; Konop, M.; Bielinska, K.; Hutsch, T.; Dziekiewicz, M.; Banaszkiewicz, A.; Ufnal, M. Inflammatory Bowel Disease Is Associated with Increased Gut-to-Blood Penetration of Short-Chain Fatty Acids: A New, Non-Invasive Marker of a Functional Intestinal Lesion. Exp. Physiol. 2019, 104, 1226–1236. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanovová, E.; Piskláková, B.; Friedecká, J.; Krystyník, O.; Friedecký, D.; Karásek, D. Plasma Short-Chain Fatty Acids and Their Derivatives in Women with Gestational Diabetes Mellitus. Separations 2021, 8, 188. https://doi.org/10.3390/separations8100188
Ivanovová E, Piskláková B, Friedecká J, Krystyník O, Friedecký D, Karásek D. Plasma Short-Chain Fatty Acids and Their Derivatives in Women with Gestational Diabetes Mellitus. Separations. 2021; 8(10):188. https://doi.org/10.3390/separations8100188
Chicago/Turabian StyleIvanovová, Eliška, Barbora Piskláková, Jaroslava Friedecká, Ondřej Krystyník, David Friedecký, and David Karásek. 2021. "Plasma Short-Chain Fatty Acids and Their Derivatives in Women with Gestational Diabetes Mellitus" Separations 8, no. 10: 188. https://doi.org/10.3390/separations8100188
APA StyleIvanovová, E., Piskláková, B., Friedecká, J., Krystyník, O., Friedecký, D., & Karásek, D. (2021). Plasma Short-Chain Fatty Acids and Their Derivatives in Women with Gestational Diabetes Mellitus. Separations, 8(10), 188. https://doi.org/10.3390/separations8100188