
The Temporal Variation of Secondary Metabolites in the Mycobiont Culture and Thallus of Parmelina carporrhizans and Parmelina quercina Analyzed using High-Performance Liquid Chromatography

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Lichen Sampling
2.2. Mycobiont Isolation and Aposymbiotic Culture
2.3. DNA Extraction and Molecular Identification
2.4. High-Performance Liquid Chromatography
3. Results
3.1. Mycobiont Growth and Qualitative Observations
3.2. Identification of secondary metabolites by High-Performance Liquid Chromatography
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmadjian, V. The Lichen Symbiosis; John Wiley & Sons: New York, NY, USA, 1993. [Google Scholar]
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete Yeasts in the Cortex of Ascomycete Macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef]
- Meeßen, J.; Ott, S. Recognition Mechanisms during the Pre-Contact State of Lichens: I. Mycobiont-Photobiont Interactions of the Mycobiont of Fulgensia bracteata. Symbiosis 2013, 59, 121–130. [Google Scholar] [CrossRef]
- Timsina, B.A.; Sorensen, J.L.; Weihrauch, D.; Piercey-Normore, M.D. Effect of Aposymbiotic Conditions on Colony Growth and Secondary Metabolite Production in the Lichen-Forming Fungus Ramalina dilacerata. Fungal Biol. 2013, 117, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Casselman, K.D. Lichen Dyes: The New Source Book, 2nd ed.; Dover Publications: Mineola, NY, USA, 2003. [Google Scholar]
- Wang, L.-S.; Narui, T.; Harada, H.; Culberson, C.F.; Culberson, W.L. Ethnic Uses of Lichens in Yunnan, China. Bryologist 2001, 104, 345–349. [Google Scholar] [CrossRef]
- Ferreira, E.S.B.; Hulme, A.N.; McNab, H.; Quye, A. The Natural Constituents of Historical Textile Dyes. Chem. Soc. Rev. 2004, 33, 329–336. [Google Scholar] [CrossRef]
- Devkota, S.; Chaudhary, R.P.; Werth, S.; Scheidegger, C. Indigenous Knowledge and Use of Lichens by the Lichenophilic Communities of the Nepal Himalaya. J. Ethnobiol. Ethnomed. 2017, 13, 15. [Google Scholar] [CrossRef]
- Stocker-Wörgötter, E. Metabolic Diversity of Lichen-Forming Ascomycetous Fungi: Culturing, Polyketide and Shikimatemetabolite Production, and PKS Genes. Nat. Prod. Rep. 2008, 25, 188–200. [Google Scholar] [CrossRef]
- White, P.A.S.; Oliveira, R.C.M.; Oliveira, A.P.; Serafini, M.R.; Araújo, A.A.S.; Gelain, D.P.; Moreira, J.C.F.; Almeida, J.R.G.S.; Quintans, J.S.S.; Quintans-Junior, L.J.; et al. Antioxidant Activity and Mechanisms of Action of Natural Compounds Isolated from Lichens: A Systematic Review. Molecules 2014, 19, 14496–14527. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Darokar, M.P.; Saikia, D.; Pal, A.; Fatima, A.; Khanuja, S.P.S. Antimycobacterial Activity of Lichens. Pharm. Biol. 2007, 45, 200–204. [Google Scholar] [CrossRef]
- Cheng, B.; Cao, S.; Vasquez, V.; Annamalai, T.; Tamayo-Castillo, G.; Clardy, J.; Tse-Dinh, Y.-C. Identification of Anziaic Acid, a Lichen Depside from Hypotrachyna sp., as a New Topoisomerase Poison Inhibitor. PLoS ONE 2013, 8, e60770. [Google Scholar] [CrossRef]
- Kosanić, M.; Manojlović, N.; Janković, S.; Stanojković, T.; Ranković, B. Evernia prunastri and Pseudoevernia furfuraceae Lichens and Their Major Metabolites as Antioxidant, Antimicrobial and Anticancer Agents. Food Chem. Toxicol. 2013, 53, 112–118. [Google Scholar] [CrossRef]
- Honda, N.; Freitas, D.; Micheletti, A.; Carvalho, N.; Spielmann, A.; Canêz, L. Parmotrema screminiae (Parmeliaceae), a Novel Lichen Species from Brazil with Potent Antimicrobial Activity. Orbital Electron. J. Chem. 2016, 8, 334–340. [Google Scholar] [CrossRef]
- Jeon, H.-S.; Lokos, L.; Han, K.S.; Ryu, J.-A.; Kim, J.-A.; Koh, Y.-J.; Hur, J.-S. Isolation of Lichen-Forming Fungi from Hungarian Lichens and Their Antifungal Activity Against Fungal Pathogens of Hot Pepper Anthracnose. Plant Pathol. J. 2009, 25, 38–46. [Google Scholar] [CrossRef]
- Kowalski, M.; Hausner, G.; Piercey-Normore, M.D. Bioactivity of Secondary Metabolites and Thallus Extracts from Lichen Fungi. Mycoscience 2011, 52, 413–418. [Google Scholar] [CrossRef]
- Rancan, F.; Rosan, S.; Boehm, K.; Fernández, E.; Hidalgo, M.E.; Quihot, W.; Rubio, C.; Boehm, F.; Piazena, H.; Oltmanns, U. Protection against UVB Irradiation by Natural Filters Extracted from Lichens. J. Photochem. Photobiol. B 2002, 68, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.-H.; Chollet-Krugler, M.; Gouault, N.; Tomasi, S. UV-Protectant Metabolites from Lichens and Their Symbiotic Partners. Nat. Prod. Rep. 2013, 30, 1490–1508. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C.; Mahadik, N.; Morey, M. Antioxidative and Cardiovascular-Protective Activities of Metabolite Usnic Acid and Psoromic Acid Produced by Lichen Species Usnea complanata under Submerged Fermentation. Pharm. Biol. 2012, 50, 968–979. [Google Scholar] [CrossRef]
- Fernández-Moriano, C.; Divakar, P.K.; Crespo, A.; Gómez-Serranillos, M.P. In Vitro Neuroprotective Potential of Lichen Metabolite Fumarprotocetraric Acid via Intracellular Redox Modulation. Toxicol. Appl. Pharmacol. 2017, 316, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Fazio, A.T.; Bertoni, M.D.; Adler, M.T.; Ruiz, L.B.; Rosso, M.L.; Muggia, L.; Hager, A.; Stocker-Wörgötter, E.; Maier, M.S. Culture Studies on the Mycobiont Isolated from Parmotrema reticulatum (Taylor) Choisy: Metabolite Production under Different Conditions. Mycol. Progress. 2009, 8, 359–365. [Google Scholar] [CrossRef]
- Thadhani, V.M.; Karunaratne, V. Potential of Lichen Compounds as Antidiabetic Agents with Antioxidative Properties: A Review. Oxid. Med. Cell. Longev. 2017, 2017, 2079697. [Google Scholar] [CrossRef]
- Rezanka, T.; Dembitsky, V.M. The Colleflaccinosides, Two Chiral Bianthraquinone Glycosides with Antitumor Activity from the Lichen Collema flaccidum Collected in Israel and Russia. Nat. Prod. Res. 2006, 20, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-S.; Kim, T.K.; Kim, J.E.; Hong, J.-M.; Nguyen, T.T.T.; Han, S.J.; Youn, U.J.; Yim, J.H.; Kim, I.-C. Anticancer Activity of Ramalin, a Secondary Metabolite from the Antarctic Lichen Ramalina terebrata, against Colorectal Cancer Cells. Molecules 2017, 22, 1361. [Google Scholar] [CrossRef] [PubMed]
- Carlos, I.Z.; Quilles, M.B.; Carli, C.B.A.; Maia, D.C.G.; Benzatti, F.P.; Lopes, T.I.B.; Gianini, A.S.; Brum, R.L.; Vilegas, W.; dos Santos, L.C.; et al. Lichen Metabolites Modulate Hydrogen Peroxide and Nitric Oxide in Mouse Macrophages. Z. Nat. C. J. Biosci. 2009, 64, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Upreti, D.K.; Divakar, P.K.; Nayaka, S. Commercial and Ethnic Use of Lichens in India. Econ. Bot. 2005, 59, 269–273. [Google Scholar] [CrossRef]
- Joulain, D.; Tabacchi, R. Lichen Extracts as Raw Materials in Perfumery. Part 1: Oakmoss. Flavour Fragr. J. 2009, 24, 49–61. [Google Scholar] [CrossRef]
- Shukla, P.; Upreti, D.K. Lichen Dyes: Current Scenario and Future Prospects. In Recent Advances in Lichenology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 209–230. [Google Scholar] [CrossRef]
- Shaheen, S.; Iqbal, Z.; Hussain, M. First Report of Dye Yielding Potential and Compounds of Lichens; a Cultural Heritage of Himalayan Communities, Pakistan. Pak. J. Bot. 2019, 51, 341–360. [Google Scholar] [CrossRef]
- Devkota, S.; Weerakoon, G. Everniastrum nepalense. In The IUCN Red List of Threatened Species 2017; IUCN: Gland, Switzerland, 2017. [Google Scholar] [CrossRef]
- Aptroot, A.; Perez-Ortega, S. Xanthoparmelia beccae. In IUCN Red List Assessment; IUCN: Gland, Switzerland, 2018. [Google Scholar] [CrossRef]
- Salgado, F.; Albornoz, L.; Cortéz, C.; Stashenko, E.; Urrea-Vallejo, K.; Nagles, E.; Galicia-Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary Metabolite Profiling of Species of the Genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2017, 23, 54. [Google Scholar] [CrossRef]
- Horák, J.; Materna, J.; Halda, J.P.; Mladenović, S.; Bogusch, P.; Pech, P. Biodiversity in Remnants of Natural Mountain Forests under Conservation-Oriented Management. Sci. Rep. 2019, 9, 89. [Google Scholar] [CrossRef]
- Pykälä, J. Habitat Loss and Deterioration Explain the Disappearance of Populations of Threatened Vascular Plants, Bryophytes and Lichens in a Hemiboreal Landscape. Glob. Ecol. Conserv. 2019, 18, e00610. [Google Scholar] [CrossRef]
- Díaz, E.M.; Zamora, J.C.; Ruibal, C.; Divakar, P.K.; González-Benítez, N.; Le Devehat, F.; Chollet, M.; Ferron, S.; Sauvager, A.; Boustie, J.; et al. Axenic Culture and Biosynthesis of Secondary Compounds in Lichen Symbiotic Fungi, the Parmeliaceae. Symbiosis 2020, 82, 79–93. [Google Scholar] [CrossRef]
- Zakeri, Z.; Junne, S.; Jäger, F.; Dostert, M.; Otte, V.; Neubauer, P. Lichen Cell Factories: Methods for the Isolation of Photobiont and Mycobiont Partners for Defined Pure and Co-Cultivation. Microb. Cell Factories 2022, 21, 80. [Google Scholar] [CrossRef]
- Culberson, C.F.; Culberson, W.L. Chemosyndromic Variation in Lichens. Syst. Bot. 1976, 1, 325–339. [Google Scholar] [CrossRef]
- Elix, J.A. Biochemistry and Secondary Metabolites. In Lichen Biology; Nash, T., III, Ed.; Cambridge University Press: Cambridge, UK, 1996; pp. 154–180. [Google Scholar]
- Brunauer, G.; Hager, A.; Grube, M.; Türk, R.; Stocker-Wörgötter, E. Alterations in Secondary Metabolism of Aposymbiotically Grown Mycobionts of Xanthoria elegans and Cultured Resynthesis Stages. Plant. Physiol. Biochem. 2007, 45, 146–151. [Google Scholar] [CrossRef]
- Stocker-Wörgötter, E.; Elix, J.A.; Grube, M. Secondary Chemistry of Lichen-Forming Fungi: Chemosyndromic Variation and DNA-Analyses of Cultures and Chemotypes in the Ramalina farinacea Complex. bryo 2004, 107, 152–162. [Google Scholar] [CrossRef]
- Hager, A.; Stocker-Wörgötter, E. Secondary Chemistry and DNA-Analyses of the Australian Lichen Heterodea muelleri (Hampe) Nyl. and Culture of the Symbionts. Symbiosis 2005, 39, 13–19. [Google Scholar]
- Bertrand, R.L.; Sorensen, J.L. Lost in Translation: Challenges with Heterologous Expression of Lichen Polyketide Synthases. ChemistrySelect 2019, 4, 6473–6483. [Google Scholar] [CrossRef]
- Pichler, G.; Candotto Carniel, F.; Muggia, L.; Holzinger, A.; Tretiach, M.; Kranner, I. Enhanced Culturing Techniques for the Mycobiont Isolated from the Lichen Xanthoria parietina. Mycol. Progress. 2021, 20, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.C.; Divakar, P.K.; Zhang, N.; González, N.; Struwe, L. Non-Developing Ascospores in Apothecia of Asexually Reproducing Lichen-Forming Fungi. Int. Microbiol. 2013, 16, 145–155. [Google Scholar] [CrossRef]
- Molina, M.; Divakar, P.; González-Benítez, N. Success in the Isolation and Axenic Culture of Anaptychia ciliaris (Physciaceae, Lecanoromycetes) Mycobiont. Mycoscience 2015, 56, 351–358. [Google Scholar] [CrossRef]
- Hale, M.E. A Monograph of the Lichen Genus Parmelia Acharius Sensu Stricto (Ascomycotina: Parmeliaceae); Smithsonian Institution Press: Washington, DC, USA, 1987; pp. 1–64. [Google Scholar]
- Alors, D.; Cendón-Flórez, Y.; Divakar, P.K.; Crespo, A.; Benítez, G.N.; Molina, M.C. Differences in the Sexual Aposymbiotic Phase of the Reproductive Cycles of Parmelina carporrhizans and P. quercina. Possible Implications for Their Reproductive Biology. Lichenologist 2019, 51, 175–186. [Google Scholar] [CrossRef]
- del Molina, M.C.; Crespo, A. Comparison of Development of Axenic Cultures of Five Species of Lichen-Forming Fungi. Mycol. Res. 2000, 104, 595–602. [Google Scholar] [CrossRef]
- Deason, T.R.; Bold, H.C. Psychological Studies: I. Exploratory Studies of Texas Soil Algae; The University of Texas Publication: Austin, TX, USA, 1960. [Google Scholar]
- Lilly, H.L.; Barnett, V.G. Physiology of the Fungi; McGraw-Hill: New York, NY, USA, 1951. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Sninsky, J.J., White, T.J., Eds.; Academic Press: Millbrae, CA, USA, 1989; Volume 38, pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Alors, D.; Dal Grande, F.; Schmitt, I.; Kraichak, E.; Lumbsch, H.T.; Crespo, A.; Divakar, P.K. Characterization of Fungus-Specific Microsatellite Markers in the Lichen-Forming Fungus Parmelina carporrhizans (Parmeliaceae). Appl. Plant. Sci. 2014, 2, 1400081. [Google Scholar] [CrossRef] [PubMed]
- Feige, G.B.; Lumbsch, T.; Huneck, S.; Elix, J. Identification of Lichen Substances by a Standardize High-Performance Liquid Chromatography Method. J. Chromatogr. A 1993, 646, 417–427. [Google Scholar] [CrossRef]
- Benatti, M.N.; Gernert, M.; Schmitt, I. Parmotrema hydrium, a New Species of Parmeliaceae in Southeastern Brazil. Acta Bot. Bras. 2013, 27, 810–814. [Google Scholar] [CrossRef]
- Clerc, P.; Truong, C. The Non-Sorediate and Non-Isidiate Parmelina Species (Lichenized Ascomycetes, Parmeliaceae) in Switzerland—Parmelina atricha (Nyl.) P. Clerc Reinstated in the European Lichen Flora. Sauteria 2008, 15, 175–194. [Google Scholar]
- Molina, M.C.; Crespo, A.; Vicente, C.; Elix, J.A. Differences in the Composition of Phenolics and Fatty Acids of Cultured Mycobiont and Thallus of Physconia distorta. Plant Physiol. Biochem. 2003, 41, 175–180. [Google Scholar] [CrossRef]
- Liu, X.-T.; Winkler, A.L.; Schwan, W.R.; Volk, T.J.; Rott, M.A.; Monte, A. Antibacterial Compounds from Mushrooms I: A Lanostane-Type Triterpene and Prenylphenol Derivatives from Jahnoporus hirtus and Albatrellus flettii and Their Activities against Bacillus cereus and Enterococcus faecalis. Planta Med. 2010, 76, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Nadal, B.; Thetiot-Laurent, S.; Pin, S.; Renault, J.-P.; Cressier, D.; Rima, G.; Le Roux, A.; Meunier, S.; Wagner, A.; Lion, C.; et al. Synthesis and Antioxidant Properties of Pulvinic Acids Analogues. Bioorganic Med. Chem. 2010, 18, 7931–7939. [Google Scholar] [CrossRef]
- Cordeiro, L.M.C.; Iacomini, M.; Stocker-Wörgötter, E. Culture Studies and Secondary Compounds of Six Ramalina Species. Mycol. Res. 2004, 108, 489–497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| P. carporrhizans | P. quercina | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Substances | RT | Thallus | Early | Late | Secreted | Thallus | Early | Late | Secreted |
| Lecanoric acid | 4.66 | Ma | - | - | - | Ma | - | - | - |
| Atranorin acid | 7.68 | mi | - | - | - | tra | - | - | - |
| Chloratranorin | 8.12 | mi | - | - | - | tra | - | - | - |
| Evernic acid | 6.11 | tra | - | - | - | - | - | - | - |
| Confluentic acid | 6.32 | - | Ma | - | - | - | - | - | - |
| Pulvinic acid | 4.15 | - | - | - | - | - | mi | - | - |
| Norlobaridone | 6.26 | - | - | Ma | - | - | - | - | - |
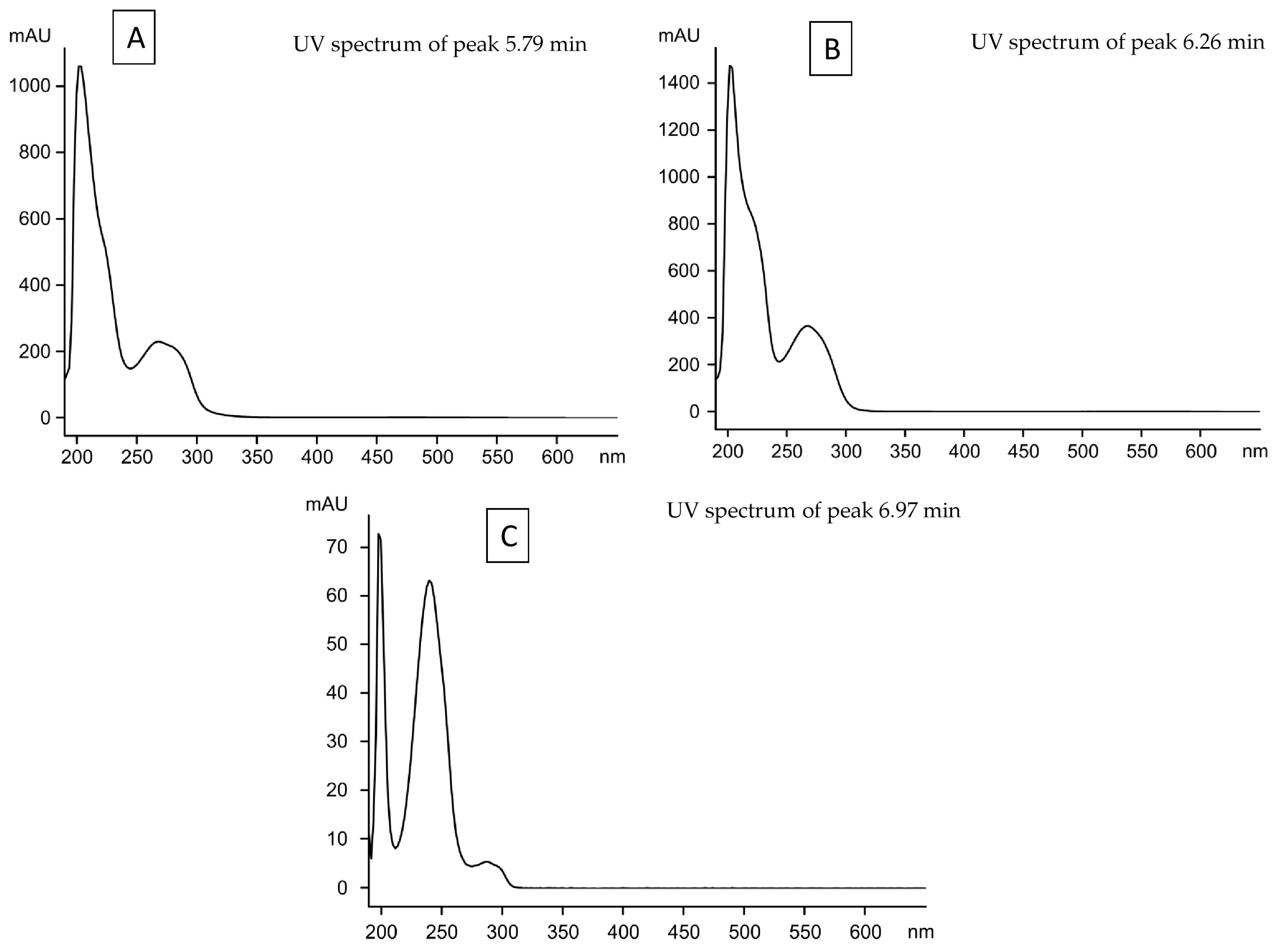
| Unknown 1 | 5.79 | - | - | - | - | - | - | Ma | - |
| Unknown 2 | 6.26 | - | - | - | - | - | - | Ma | - |
| Unknown 3 | 6.97 | - | - | - | Ma | - | Ma | - | Ma |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alors, D.; Divakar, P.K.; Calchera, A.; Schmitt, I.; Crespo, A.; Molina, M.C. The Temporal Variation of Secondary Metabolites in the Mycobiont Culture and Thallus of Parmelina carporrhizans and Parmelina quercina Analyzed using High-Performance Liquid Chromatography. Separations 2023, 10, 399. https://doi.org/10.3390/separations10070399
Alors D, Divakar PK, Calchera A, Schmitt I, Crespo A, Molina MC. The Temporal Variation of Secondary Metabolites in the Mycobiont Culture and Thallus of Parmelina carporrhizans and Parmelina quercina Analyzed using High-Performance Liquid Chromatography. Separations. 2023; 10(7):399. https://doi.org/10.3390/separations10070399
Chicago/Turabian StyleAlors, David, Pradeep Kumar Divakar, Anjuli Calchera, Imke Schmitt, Ana Crespo, and María Carmen Molina. 2023. "The Temporal Variation of Secondary Metabolites in the Mycobiont Culture and Thallus of Parmelina carporrhizans and Parmelina quercina Analyzed using High-Performance Liquid Chromatography" Separations 10, no. 7: 399. https://doi.org/10.3390/separations10070399
APA StyleAlors, D., Divakar, P. K., Calchera, A., Schmitt, I., Crespo, A., & Molina, M. C. (2023). The Temporal Variation of Secondary Metabolites in the Mycobiont Culture and Thallus of Parmelina carporrhizans and Parmelina quercina Analyzed using High-Performance Liquid Chromatography. Separations, 10(7), 399. https://doi.org/10.3390/separations10070399