
Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries


Abstract
1. Introduction
2. Materials and Methods
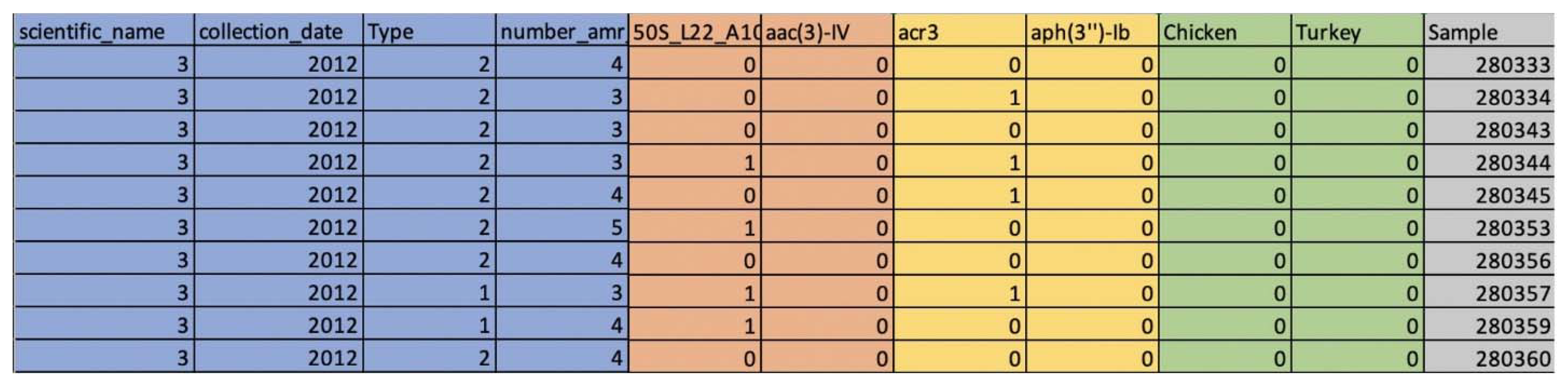
2.1. The Stress Response Gene Data from the NCBI Pathogen Detection Isolates Browser
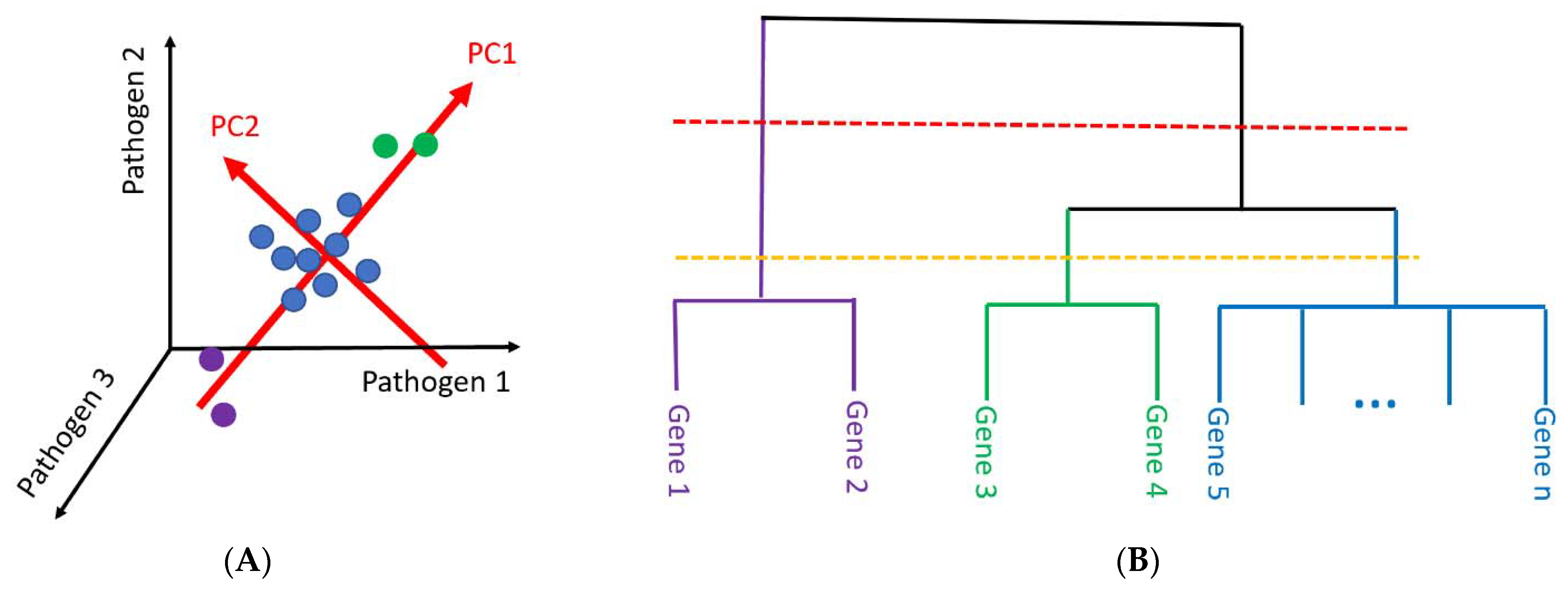
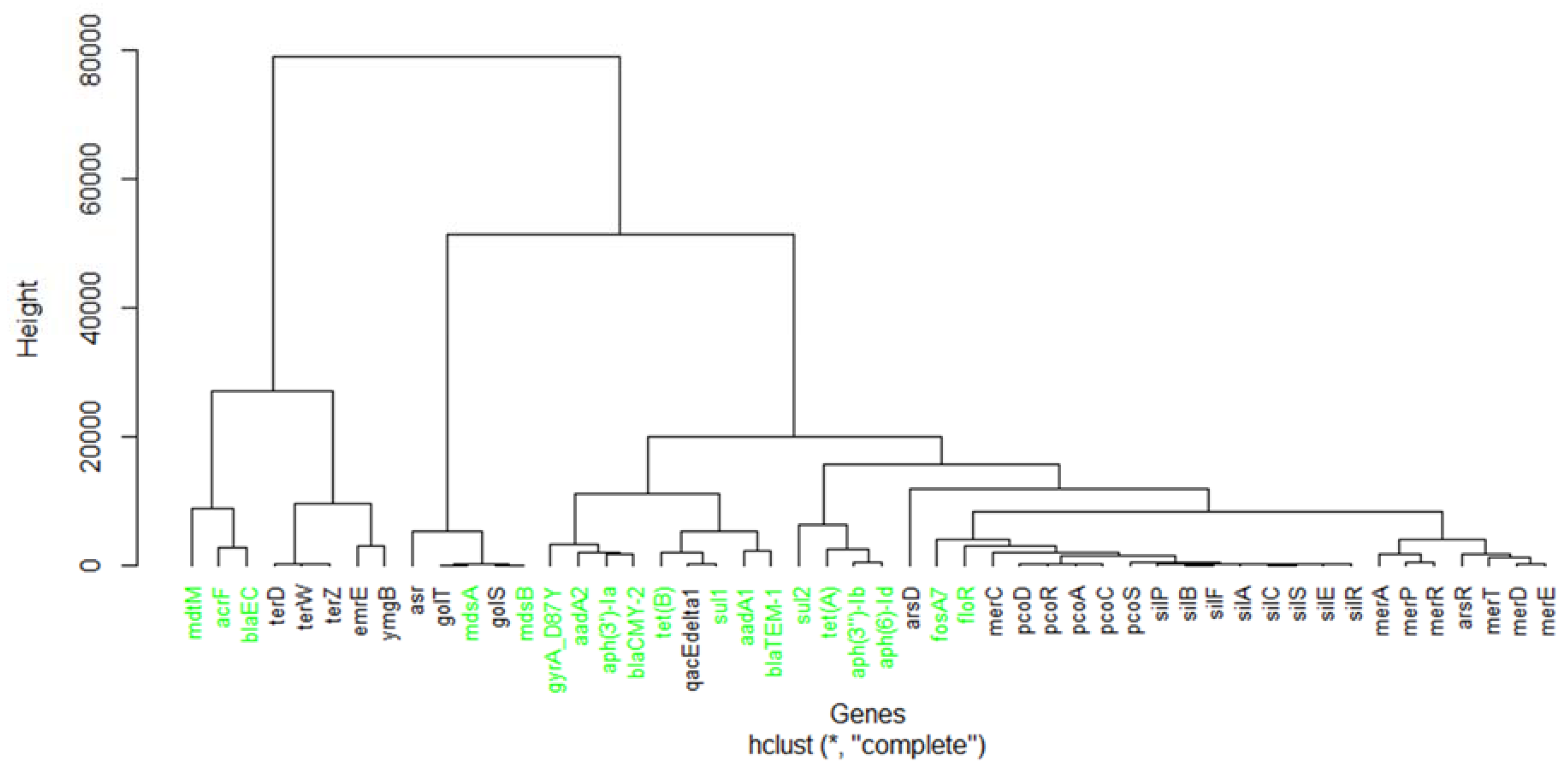
2.2. Principal Component Analysis and Hierarchical Clustering
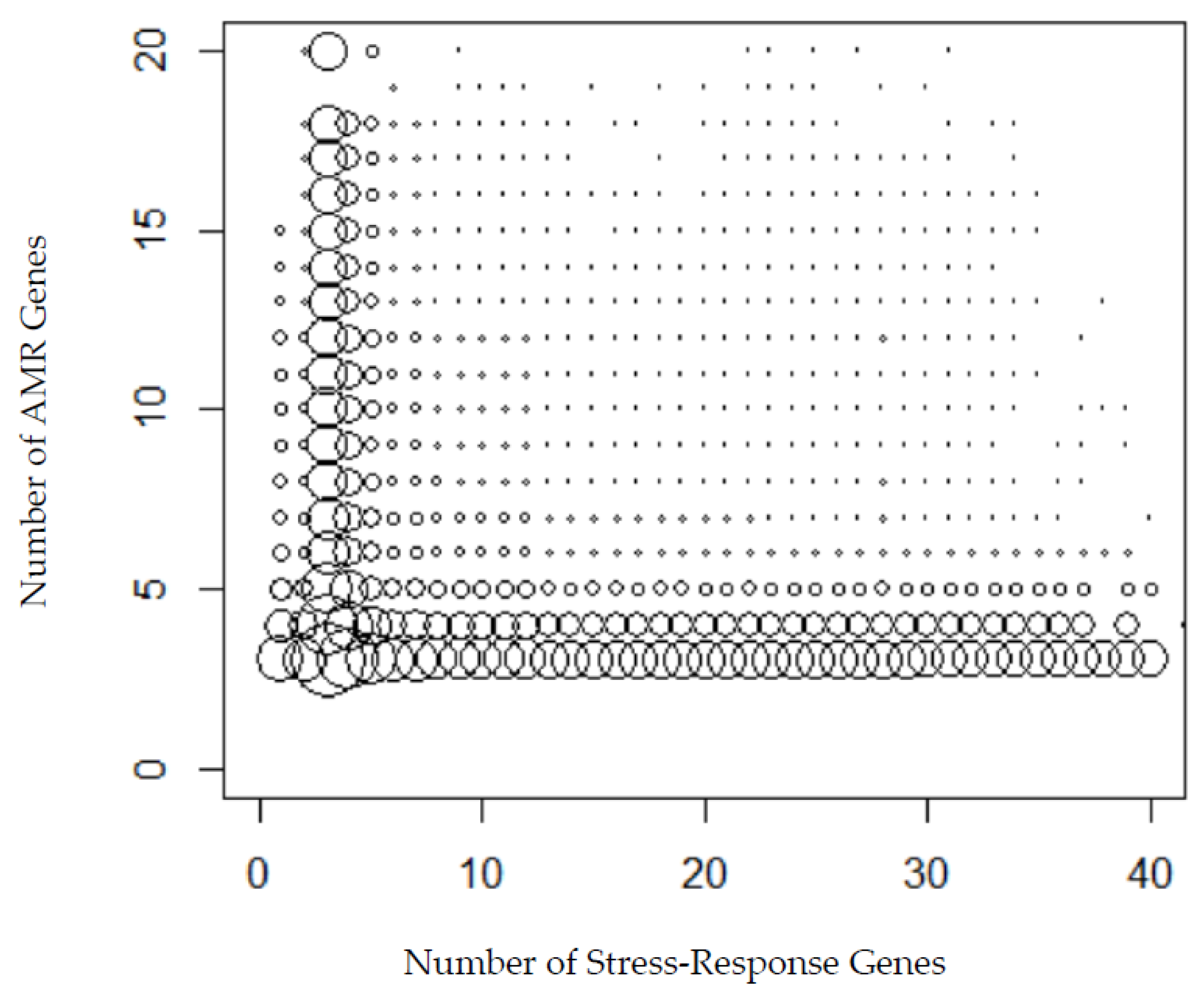
2.3. Investigating the Occurrence Correlation between the Stress Response Genes and AMR Genes
3. Results
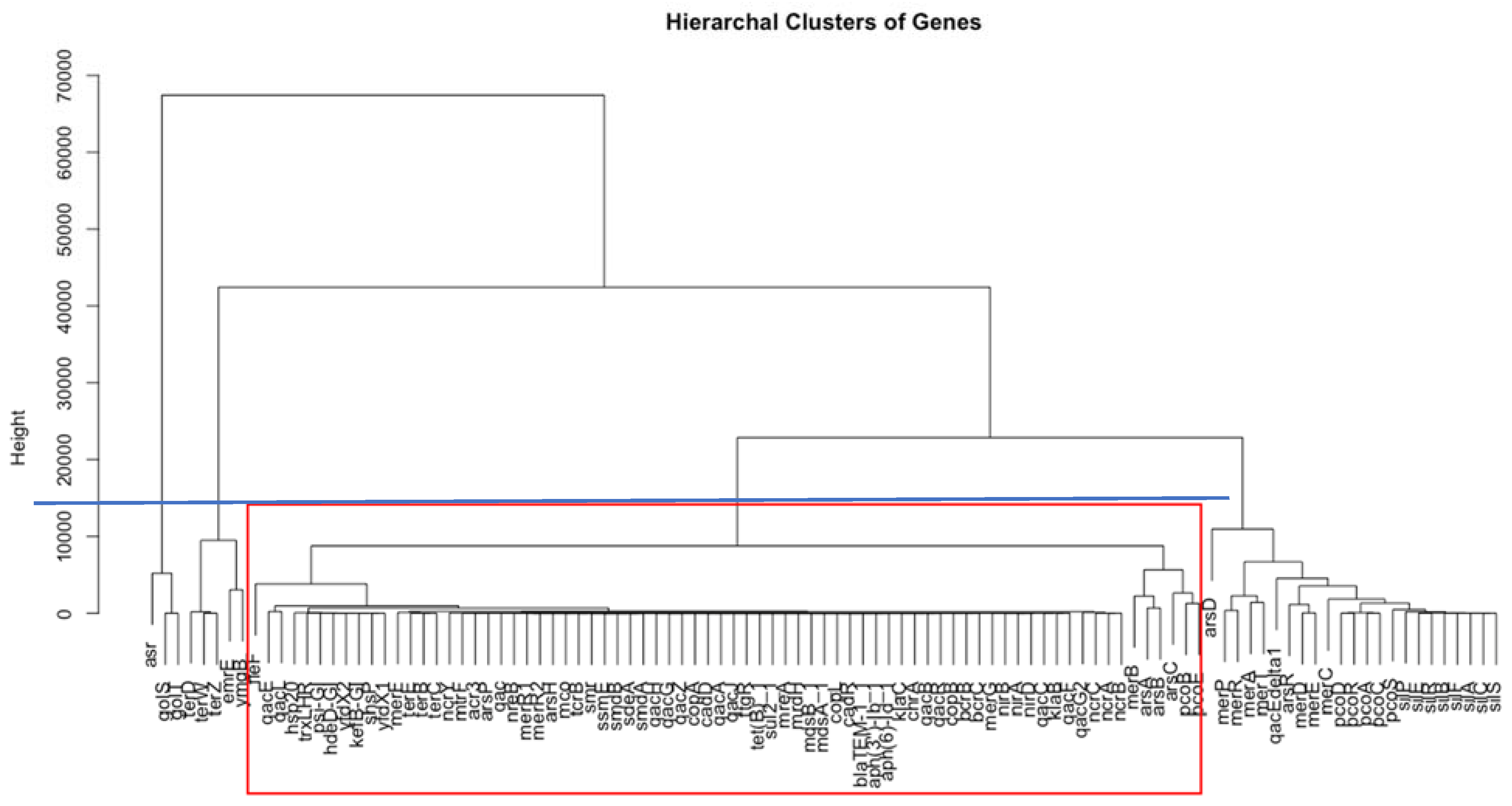
3.1. Identification of Important Stress Response Genes for the Five Countries
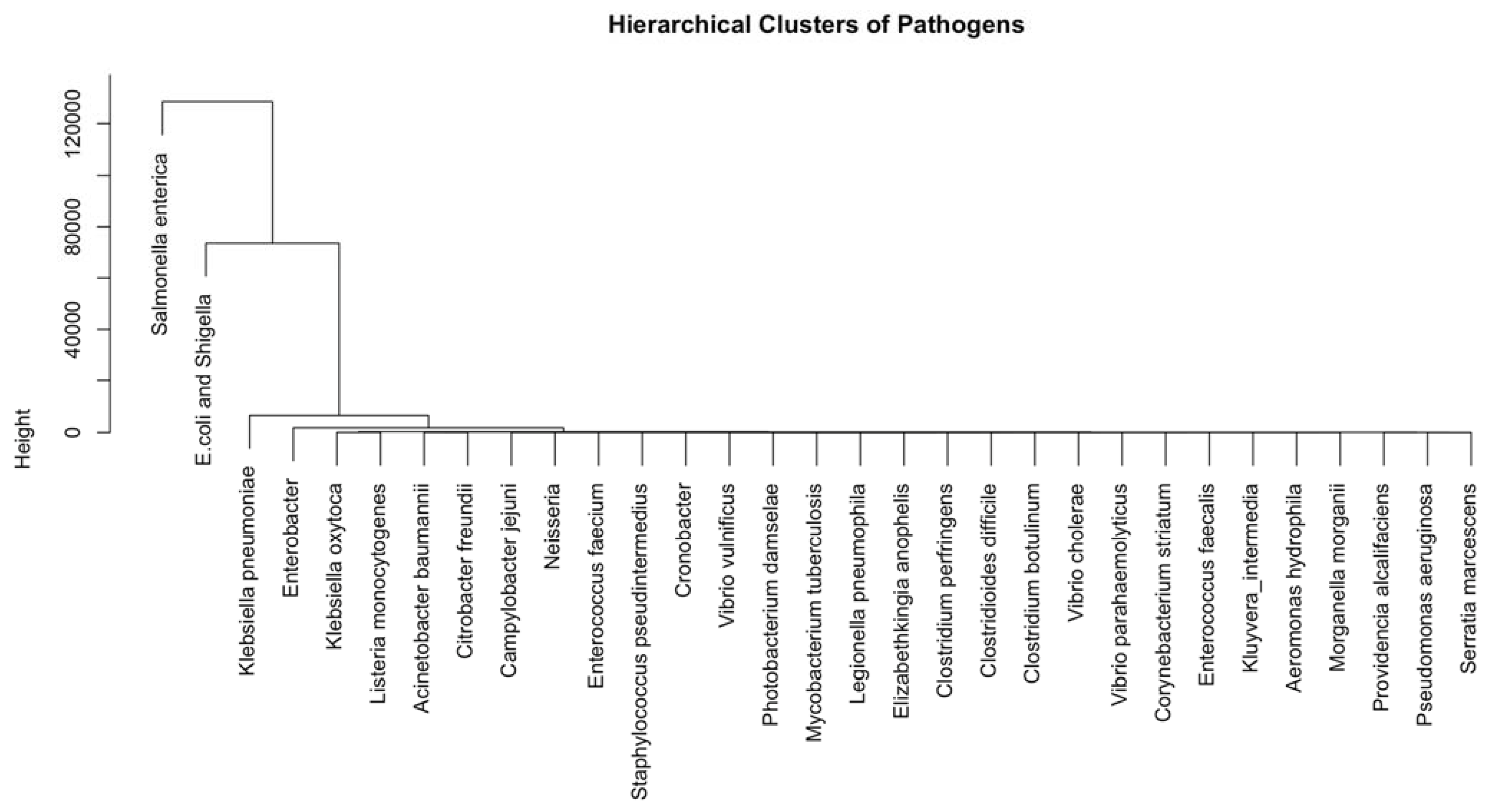
3.2. Identification of Important Stress Response Pathogens for the Five Countries
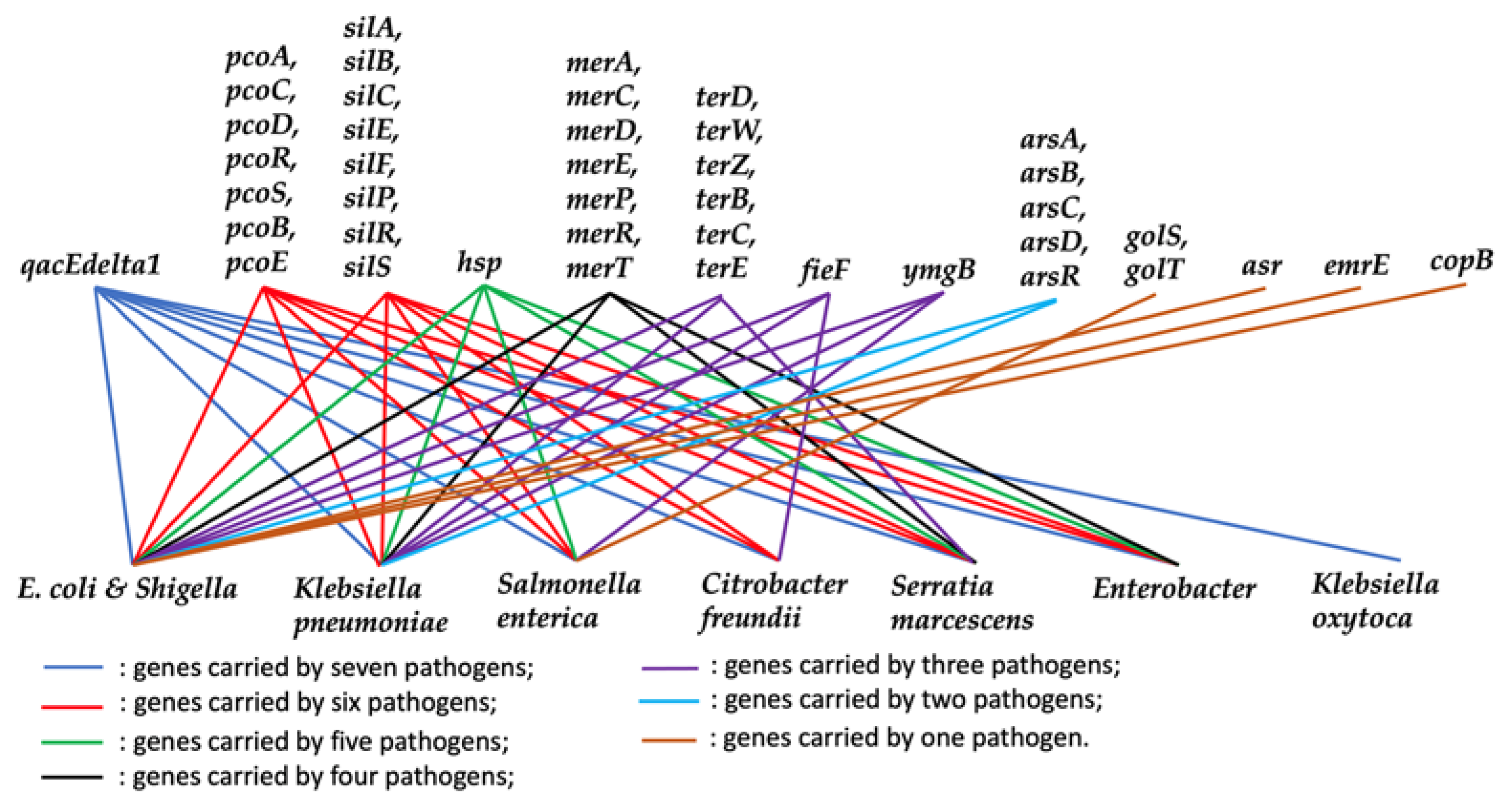
3.3. The Relationship between Stress Response Genes and AMR Genes for Individual Countries
3.4. Comparing the Numbers of Stress Response Genes and AMR Genes in Individual Samples for Each Country
4. Discussion
4.1. Significance of Important and Shared Stress Response Genes
4.2. Relationship between Stress Response Genes and AMR Genes
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Scallan, E.; Hoekstra, R.M.; Widdowson, M.A.; Hall, A.J.; Griffin, P.M. Foodborne Illness Acquired in the United States Response. Emerg. Infect. Dis. 2011, 17, 1339–1340. [Google Scholar] [CrossRef]
- Boor, K.J. Bacterial stress responses: What doesn’t kill them can make them stronger. PLoS Biol. 2006, 4, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Smith, D.W.; Li, Y. Natural freezing as a wastewater treatment method: E. coli inactivation capacity. Water Res. 2006, 40, 2321–2326. [Google Scholar] [CrossRef] [PubMed]
- Weiser, R.S.; Hargiss, C.O. Studies on the Death of Bacteria at Low Temperatures. The Comparative Effects of Crystallization, Vitromelting, and Devitrification on the Mortality of Escherichia-Coli. J. Bacteriol. 1946, 52, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Bang, W.; Drake, M.A. Stress response of Escherichia coli. Compr. Rev. Food Sci. Food Saf. 2006, 5, 52–64. [Google Scholar] [CrossRef]
- Duhring, S.; Germerodt, S.; Skerka, C.; Zipfel, P.F.; Dandekar, T.; Schuster, S. Host-pathogen interactions between the human innate immune system and Candida albicans—Understanding and modeling defense and evasion strategies. Front. Microbiol. 2015, 6, 625. [Google Scholar] [CrossRef]
- National Library of Medicine (US). The NCBI Pathogen Detection Project; NCBI: Bethesda, MD, USA, 2016. [Google Scholar]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- O’Brien, T.F. Emergence, spread, and environmental effect of antimicrobial resistance: How use of an antimicrobial anywhere can increase resistance to any antimicrobial anywhere else. Clin. Infect. Dis. 2002, 34, S78–S84. [Google Scholar] [CrossRef]
- Harbottle, H.; Thakur, S.; Zhao, S.; White, D.G. Genetics of antimicrobial resistance. Anim. Biotechnol. 2006, 17, 111–124. [Google Scholar] [CrossRef]
- Brazas, M.D.; Hancock, R.E.W. Using microarray gene signatures to elucidate mechanisms of antibiotic action and resistance. Drug Discov. Today 2005, 10, 1245–1252. [Google Scholar] [CrossRef]
- Lee, S.; Hinz, A.; Bauerle, E.; Angermeyer, A.; Juhaszova, K.; Kaneko, Y.; Singh, P.K.; Manoil, C. Targeting a bacterial stress response to enhance antibiotic action. Proc. Natl. Acad. Sci. USA 2009, 106, 14570–14575. [Google Scholar] [CrossRef] [PubMed]
- Ebinesh, A. Bacterial Stress Response and Cross Resistance to Antibiotics in the Light of Natural Selection. J. Infect. Dis. Immune Ther. 2017, 1, 1. [Google Scholar]
- Poole, K. Stress responses as determinants of antimicrobial resistance in Pseudomonas aeruginosa: Multidrug efflux and more. Can. J. Microbiol. 2014, 60, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zheng, J.; Deng, T.; Peng, J.; Daniel, D.; Jia, Q.; Huang, Z.Y. An Analysis of Antimicrobial Resistance of Clinical Pathogens from Historical Samples for Six Countries. Processes 2019, 7, 964. [Google Scholar] [CrossRef]
- Yang, K.; Wang, A.; Fu, M.; Wang, A.; Chen, K.; Jia, Q.; Huang, Z.Y. Investigation of Incidents and Trends of Antimicrobial Resistance in Foodborne Pathogens in Eight Countries from Historical Sample Data. Int. J. Environ. Res. Public Health 2020, 17, 472. [Google Scholar] [CrossRef]
- Hua, M.G.; Huang, W.L.; Chen, A.; Rehmet, M.; Jin, C.; Huang, Z.Y. Comparison of Antimicrobial Resistance Detected in Environmental and Clinical Isolates from Historical Data for the US. Biomed Res. Int. 2020, 2020. [Google Scholar] [CrossRef]
- Kourti, T. Application of latent variable methods to process control and multivariate statistical process control in industry. Int. J. Adapt. Control. 2005, 19, 213–246. [Google Scholar] [CrossRef]
- Qin, S.J. Statistical process monitoring: Basics and beyond. J. Chemom. 2003, 17, 480–502. [Google Scholar] [CrossRef]
- Bar-Joseph, Z.; Demaine, E.D.; Gifford, D.K.; Srebro, N.; Hamel, A.M.; Jaakkola, T.S. K-ary clustering with optimal leaf ordering for gene expression data. Bioinformatics 2003, 19, 1070–1078. [Google Scholar] [CrossRef]
- Wang, X.Z.; Smith, K.; Hyndman, R. Characteristic-based clustering for time series data. Data Min. Knowl. Discov. 2006, 13, 335–364. [Google Scholar] [CrossRef]
- Boyd, E.S.; Barkay, T. The mercury resistance operon: From an origin in a geothermal environment to an efficient detoxification machine. Front. Microbiol. 2012, 3, 349. [Google Scholar] [CrossRef]
- Butcher, B.G.; Deane, S.M.; Rawlings, D.E. The chromosomal arsenic resistance genes of Thiobacillus ferrooxidans have an unusual arrangement and confer increased arsenic and antimony resistance to Escherichia coli. Appl. Environ. Microbol. 2000, 66, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Uyaguari-Diaz, M.I.; Croxen, M.A.; Luo, Z.Y.; Cronin, K.I.; Chan, M.; Baticados, W.N.; Nesbitt, M.J.; Li, S.R.; Miller, K.M.; Dooley, D.; et al. Human Activity Determines the Presence of Integron-Associated and Antibiotic Resistance Genes in Southwestern British Columbia. Front. Microbiol. 2018, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Suziedeliene, E.; Suziedelis, K.; Garbenciute, V.; Normark, S. The acid-inducible asr gene in Escherichia coli: Transcriptional control by the phoBR operon. J. Bacteriol. 1999, 181, 2084–2093. [Google Scholar] [CrossRef]
- Munoz-Villagran, C.M.; Mendez, K.N.; Cornejo, F.; Figueroa, M.; Undabarrena, A.; Morales, E.H.; Arenas-Salinas, M.; Arenas, F.A.; Castro-Nallar, E.; Vasquez, C.C. Comparative genomic analysis of a new tellurite-resistant Psychrobacter strain isolated from the Antarctic Peninsula. PeerJ 2018, 6, e4402. [Google Scholar] [CrossRef] [PubMed]
- Bondarczuk, K.; Piotrowska-Seget, Z. Molecular basis of active copper resistance mechanisms in Gram-negative bacteria. Cell Biol. Toxicol. 2013, 29, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Shafeeq, S.; Yesilkaya, H.; Kloosterman, T.G.; Narayanan, G.; Wandel, M.; Andrew, P.W.; Kuipers, O.P.; Morrissey, J.A. The cop operon is required for copper homeostasis and contributes to virulence in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 1255–1270. [Google Scholar] [CrossRef]
- Wu, J.; Rosen, B.P. The Arsr Protein Is a Trans-Acting Regulatory Protein. Mol. Microbiol. 1991, 5, 1331–1336. [Google Scholar] [CrossRef]
- Firrincieli, A.; Presentato, A.; Favoino, G.; Marabottini, R.; Allevato, E.; Stazi, S.R.; Mugnozza, G.S.; Harfouche, A.; Petruccioli, M.; Turner, R.J.; et al. Identification of Resistance Genes and Response to Arsenic in Rhodococcus aetherivorans BCP1. Front. Microbiol. 2019, 10, 888. [Google Scholar] [CrossRef]
- Seputiene, V.; Motiejunas, D.; Suziedelis, K.; Tomenius, H.; Normark, S.; Melefors, O.; Suziedeliene, E. Molecular characterization of the acid-inducible asr gene of Escherichia coli and its role in acid stress response. J. Bacteriol. 2003, 185, 2475–2484. [Google Scholar] [CrossRef]
- Parnanen, K.M.M.; Narciso-da-Rocha, C.; Kneis, D.; Berendonk, T.U.; Cacace, D.; Do, T.T.; Elpers, C.; Fatta-Kassinos, D.; Henriques, I.; Jaeger, T.; et al. Antibiotic resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 2019, 5, eaau9124. [Google Scholar] [CrossRef]
- Surleac, M.; Barbu, I.C.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu, I.; Talapan, D.; Nita, M.; et al. Whole genome sequencing snapshot of multi-drug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15, e0228079. [Google Scholar] [CrossRef] [PubMed]
- Pontel, L.B.; Audero, M.E.; Espariz, M.; Checa, S.K.; Soncini, F.C. GolS controls the response to gold by the hierarchical induction of Salmonella-specific genes that include a CBA efflux-coding operon. Mol. Microbiol. 2007, 66, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Page, R.; Garcia-Contreras, R.; Palermino, J.M.; Zhang, X.S.; Doshi, O.; Wood, T.K.; Peti, W. Structure and function of the Escherichia coli protein YmgB: A protein critical for biofilm formation and acid-resistance. J. Mol. Biol. 2007, 373, 11–26. [Google Scholar] [CrossRef]
- Nishino, K.; Yamaguchi, A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol. 2001, 183, 5803–5812. [Google Scholar] [CrossRef] [PubMed]
- Ranjith, K.; Arunasri, K.; Reddy, G.S.; Adicherla, H.; Sharma, S.; Shivaji, S. Global gene expression in Escherichia coli, isolated from the diseased ocular surface of the human eye with a potential to form biofilm. Gut Pathog. 2017, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Huang, J.; Shang, Z.; Xu, R.; Lou, Y.; Chen, X. Co-existence of resistance genes and their association with the genetic marker of integrons among multi-resistant Escherichia coli isolates. Acta Microbiol. Sin. 2011, 51, 458–467. [Google Scholar]
- Okade, H.; Nakagawa, S.; Sakagami, T.; Hisada, H.; Nomura, N.; Mitsuyama, J.; Yamagishi, Y.; Mikamo, H. Characterization of plasmid-mediated quinolone resistance determinants in Klebsiella pneumoniae and Escherichia coli from Tokai, Japan. J. Infect. Chemother. 2014, 20, 778–783. [Google Scholar] [CrossRef]
- Chiu, C.H.; Chen, H.L.; Kao, L.S.; Yang, C.Y.; Chu, C.; Doublet, B.; Praud, K.; Cloeckaert, A. Variant Salmonella genomic island 1 antibiotic resistance gene clusters in Salmonella enterica serovar Derby isolates from humans in Taiwan. J. Antimicrob. Chemother. 2007, 59, 325–326. [Google Scholar] [CrossRef]
- Moura, A.; Araujo, S.; Alves, M.S.; Henriques, I.; Pereira, A.; Correia, A.C.M. The contribution of Escherichia coli from human and animal sources to the integron gene pool in coastal waters. Front. Microbiol. 2014, 5, 419. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | US | UK | China | South Africa | Australia |
|---|---|---|---|---|---|
| arsA | |||||
| arsB | |||||
| arsC | |||||
| arsD | |||||
| arsR | |||||
| asr | |||||
| copB | |||||
| emrE | |||||
| fieF | |||||
| golS | |||||
| golT | |||||
| hsp | |||||
| merA | |||||
| merC | |||||
| merD | |||||
| merE | |||||
| merP | |||||
| merR | |||||
| merT | |||||
| pcoA | |||||
| pcoB | |||||
| pcoC | |||||
| pcoD | |||||
| pcoE | |||||
| pcoR | |||||
| pcoS | |||||
| qacEdelta1 | |||||
| silA | |||||
| silB | |||||
| silC | |||||
| silE | |||||
| silF | |||||
| silP | |||||
| silR | |||||
| silS | |||||
| terB | |||||
| terC | |||||
| terD | |||||
| terE | |||||
| terW | |||||
| terZ | |||||
| ymgB |
| South Africa | China | Australia | UK | US | |
|---|---|---|---|---|---|
| E. coli and Shigella | |||||
| Salmonella enterica | |||||
| Klebsiella pneumoniae | |||||
| Enterobacter | |||||
| Enterococcus faecium | |||||
| Klebsiella oxytoca | |||||
| Citrobacter freundii | |||||
| Cronobacter | |||||
| Serratia marcescens |
| Countries | Stress Response Genes | AMR Genes |
|---|---|---|
| The US | golT, golS | mdsA, mdsB |
| qacEdelta1 | sul1 | |
| The UK | golT, golS | mdsA, mdsB |
| emrE | acrF | |
| ygmB | emrD | |
| fief | sat2 | |
| arsR | tet(A) | |
| China | ymgB | mdtM |
| emrE | gyrA_s83L | |
| pcoA | oqxA | |
| qacEdelta1 | sul1 | |
| South Africa | golS, golT | mdsA, mdsB |
| hsp20, arsA, arsB, arsR | aac(3)-lla | |
| arsC, pcoB, pcoC, pcoD, pcoR, silS | catB3 | |
| Australia | ymgB | mdtM |
| copB | aac()-l, msr(C), eat(A) | |
| terD, terW, terZ | cyaA_S352T | |
| merC, merP, merT | blaCTX-M-15, gyrA_D87N | |
| qacEdelta1 | sul1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, R.; Zhang, L.; Duan, C.; Gao, M.; Feng, R.; Jia, Q.; Huang, Z. Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries. Processes 2021, 9, 927. https://doi.org/10.3390/pr9060927
Pei R, Zhang L, Duan C, Gao M, Feng R, Jia Q, Huang Z. Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries. Processes. 2021; 9(6):927. https://doi.org/10.3390/pr9060927
Chicago/Turabian StylePei, Rachael, Liz Zhang, Catherine Duan, Michael Gao, Rachel Feng, Qian Jia, and Zuyi (Jacky) Huang. 2021. "Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries" Processes 9, no. 6: 927. https://doi.org/10.3390/pr9060927
APA StylePei, R., Zhang, L., Duan, C., Gao, M., Feng, R., Jia, Q., & Huang, Z. (2021). Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries. Processes, 9(6), 927. https://doi.org/10.3390/pr9060927