
Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Scavenging Activity
[{(AControl − ABlankl) − (ATreatment − ABlank2)}/AControl − ABlank1] × 100
2.3. ABTS+ Scavenging Activity
[{(AControl − ABlankl) − (ATreatment − ABlank2)}/AControl − ABlank1] × 100
2.4. Cell Culture and UVB Treatment
2.5. Cell Viability
2.6. Intracellular ROS Generation
2.7. Protein Extraction
2.8. Western Blot
2.9. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.10. Total Phenolic Content (TPC)
2.11. High-Performance Liquid Chromatography (HPLC) Analysis
2.12. Statistical Analysis
3. Results
3.1. Heat Treatment Improved Antioxidant Activities of Licorice Extracts
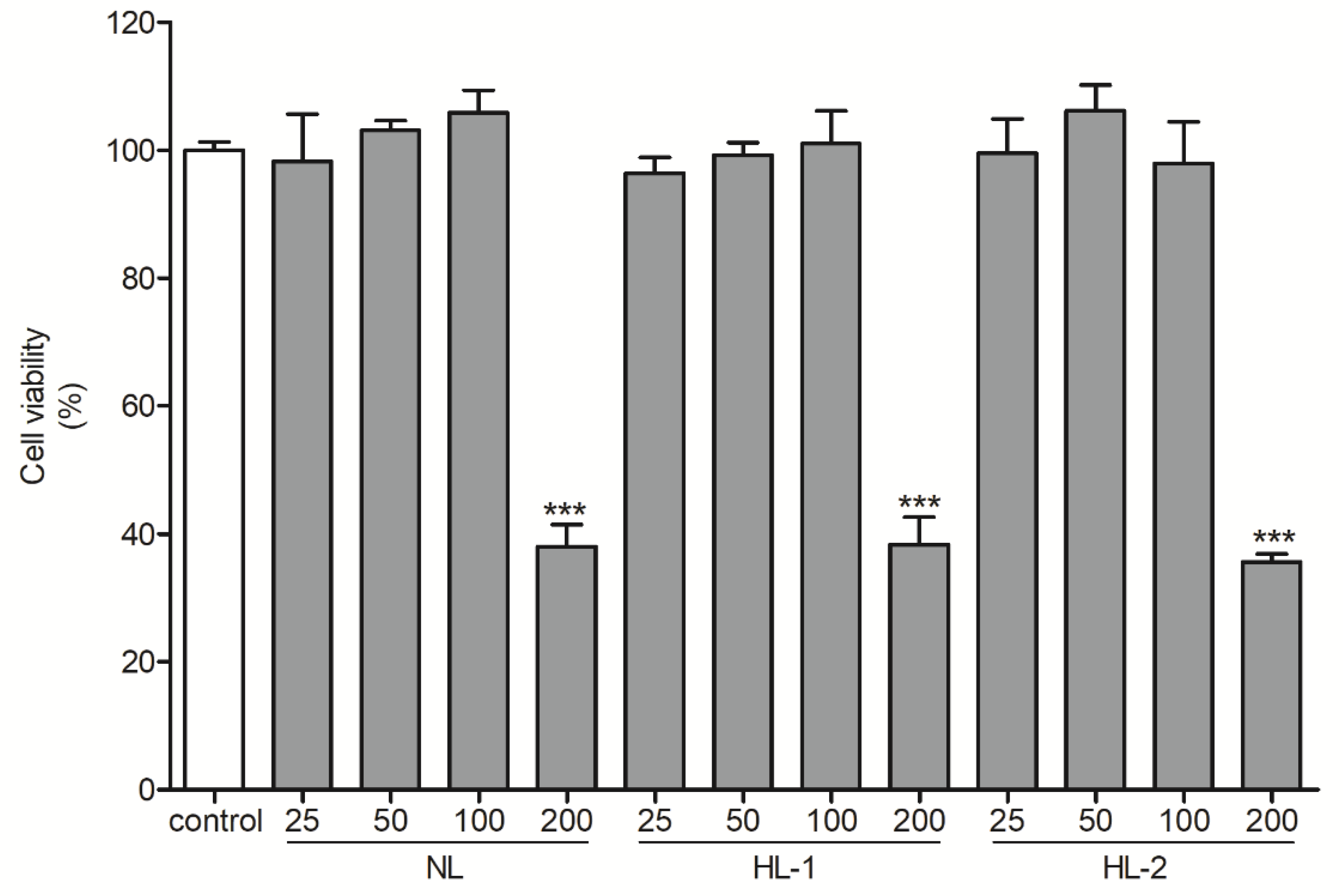
3.2. Heat Treatment Improved the UV Photoprotective Effect of Licorice Extract by Enhancing Antioxidant Properties
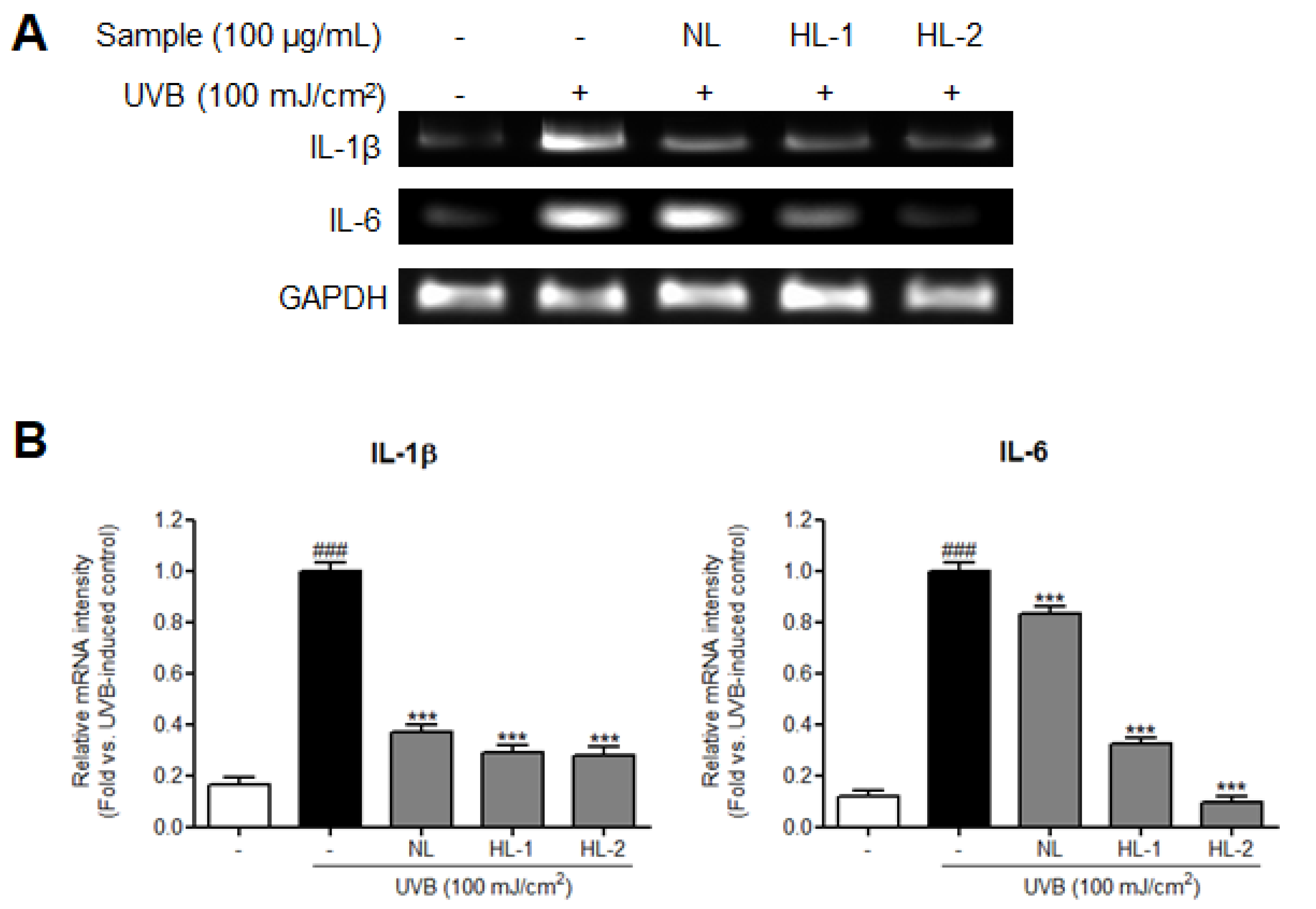
3.3. Heat Treatment Improved the UV Photoprotective Effect of Licorice Extract by Enhancing Anti-Inflammatory Properties
3.4. Heat Treatment Increased the Contents of Bioactive Compounds
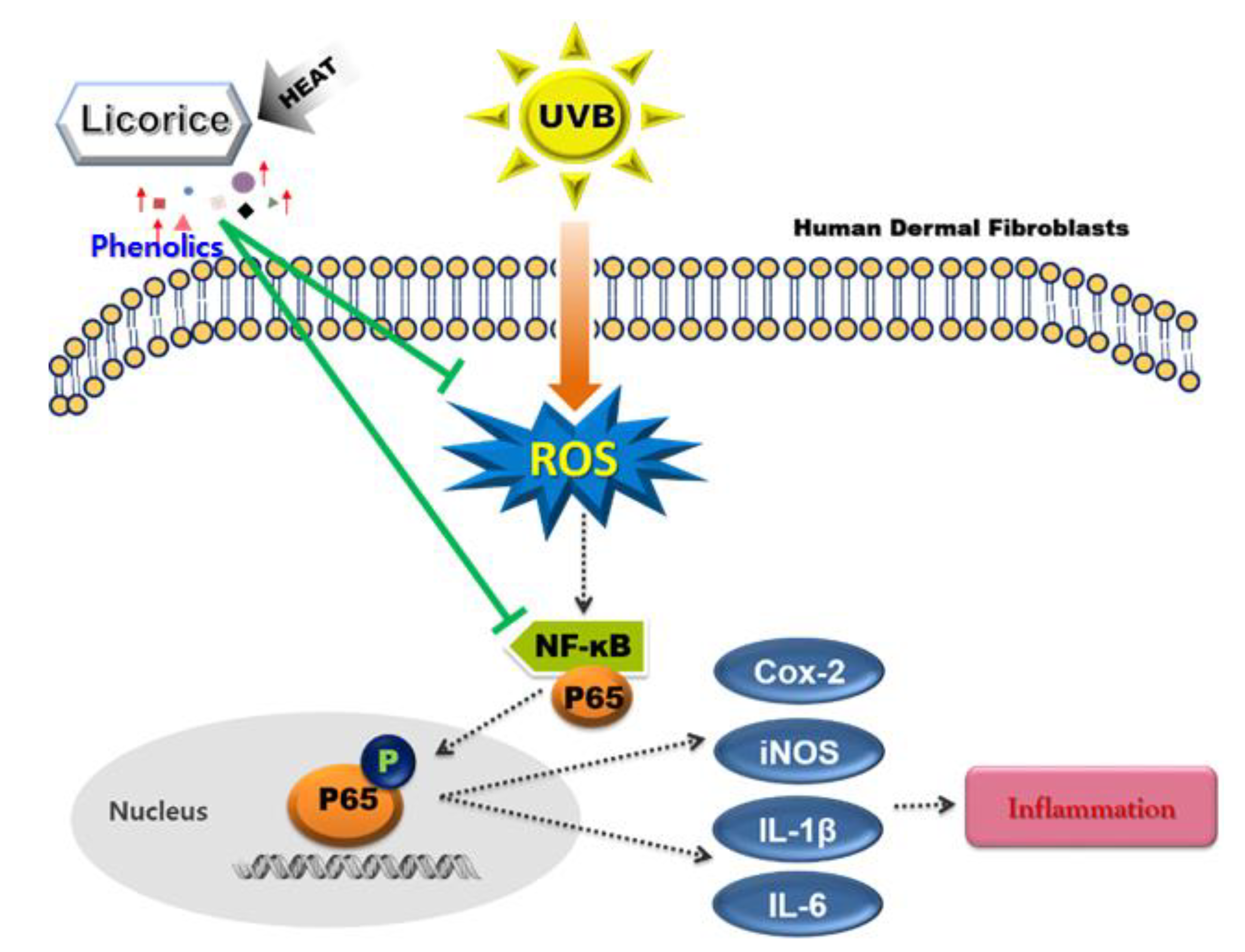
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Smart, R.; Wang, H.; Dambergs, B.; Sparrow, A.; Qian, M.C. Effect of grape bunch sunlight exposure and UV radiation on phenolics and volatile composition of Vitis vinifera L. cv. Pinot noir wine. Food Chem. 2015, 173, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.W.; Williamson, C.E.; Lucas, R.M.; Robinson, S.A.; Madronich, S.; Paul, N.D.; Bornman, J.F.; Bais, A.F.; Sulzberger, B.; Wilson, S.R. Ozone depletion, ultraviolet radiation, climate change and prospects for a sustainable future. Nat. Sustain. 2019, 2, 569–579. [Google Scholar] [CrossRef]
- Jung, Y.R.; Kim, D.H.; Kim, S.R.; An, H.J.; Lee, E.K.; Tanaka, T.; Kim, N.D.; Yokozawa, T.; Park, J.N.; Chung, H.Y. Anti-wrinkle effect of magnesium lithospermate B from Salvia miltiorrhiza BUNGE: Inhibition of MMPs via NF-kB signaling. PLoS ONE 2014, 9, e102689. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.M.; Ratti, B.A.; dos Santos Rando, F.; Fernandez, M.A.; Ueda-Nakamura, T.; Lautenschlager, S.d.O.S.; Nakamura, C.V. Metformin effect on driving cell survival pathway through inhibition of UVB-induced ROS formation in human keratinocytes. Mech. Ageing Dev. 2020, 192, 111387. [Google Scholar] [CrossRef]
- Gao, Y.; Lv, X.; Yang, H.; Peng, L.; Ci, X. Isoliquiritigenin exerts antioxidative and anti-inflammatory effects via activating the KEAP-1/Nrf2 pathway and inhibiting the NF-κB and NLRP3 pathways in carrageenan-induced pleurisy. Food Funct. 2020, 11, 2522–2534. [Google Scholar] [CrossRef]
- Kim, H.D.; Park, J.Y.; Noh, H.J.; Lee, S.E.; Lee, J.H.; Seo, K.H. Anti-inflammatory activity of Ganoderma lucidum by inhibition of NF-κB p65 phosphorylation. Korean J. Agric. Sci. 2019, 46, 653–660. [Google Scholar]
- Liu, Y.Y.; Yang, Y.N.; Feng, Z.M.; Jiang, J.S.; Zhang, P.C. Eight new triterpenoid saponins with antioxidant activity from the roots of Glycyrrhiza uralensis Fisch. Fitoterapia 2019, 133, 186–192. [Google Scholar] [CrossRef]
- Varsha, S.; Agrawal, R.; Sonam, P. Phytochemical screening and determination of anti-bacterial and anti-oxidant potential of Glycyrrhiza glabra root extracts. J. Environ. Res. Dev. 2013, 7, 4A. [Google Scholar]
- Conidi, C.; Fucà, L.; Drioli, E.; Cassano, A. A membrane-based process for the recovery of glycyrrhizin and phenolic compounds from licorice wastewaters. Molecules 2019, 24, 2279. [Google Scholar] [CrossRef]
- Martins, N.; Barros, L.; Duenas, M.; Santos Buelga, C.; Ferreira, I.C. Characterization of phenolic compounds and antioxidant properties of Glycyrrhiza glabra L. rhizomes and roots. RSC Adv. 2015, 5, 26991–26997. [Google Scholar] [CrossRef]
- Ji, S.; Li, Z.; Song, W.; Wang, Y.; Liang, W.; Li, K.; Tang, S.; Wang, Q.; Qiao, X.; Zhou, D. Bioactive constituents of Glycyrrhiza uralensis (licorice): Discovery of the effective components of a traditional herbal medicine. J. Nat. Prod. 2016, 79, 281–292. [Google Scholar] [CrossRef]
- Xuan, S.H.; Park, Y.M.; Ha, J.H.; Jeong, Y.J.; Park, S.N. The effect of dehydroglyasperin C on UVB-mediated MMPs expression in human HaCaT cells. Pharmacol. Rep. 2017. [Google Scholar] [CrossRef]
- Hamad, G.M.; Abd Elaziz, A.I.; Hassan, S.A.; Shalaby, M.A.; azim Mohdaly, A.A.A. Chemical Composition, Antioxidant, Antimicrobial and Anticancer Activities of Licorice (Glycyrrhiza glabra L.) Root and Its Application in Functional Yoghurt. Food Nutr. Res. 2020, 8, 707–715. [Google Scholar] [CrossRef]
- Vlaisavljević, S.; Šibul, F.; Sinka, I.; Zupko, I.; Ocsovszki, I.; Jovanović Šanta, S. Chemical composition, antioxidant and anticancer activity of licorice from Fruska Gora locality. IND 2018, 112, 217–224. [Google Scholar] [CrossRef]
- Yu, D.; Liu, X.; Zhang, G.; Ming, Z.; Wang, T. Isoliquiritigenin inhibits cigarette smoke-induced COPD by attenuating inflammation and oxidative stress via the regulation of the Nrf2 and NF-κB signaling pathways. Front. Pharmacol. 2018, 9, 1001. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, C.; Wang, Y.; Zhang, J.; Wang, F.; Li, L.; Meng, X.; Li, G.; Li, Y.; Wang, L. Isoliquiritigenin attenuates LPS-induced AKI by suppression of inflammation involving NF-κB pathway. Am. J. Transl. Res. 2018, 10, 4141. [Google Scholar]
- Hou, C.; Li, W.; Li, Z.; Gao, J.; Chen, Z.; Zhao, X.; Yang, Y.; Zhang, X.; Song, Y. Synthetic isoliquiritigenin inhibits human tongue squamous carcinoma cells through its antioxidant mechanism. Oxid. Med. Cell. Longev. 2017, 2017, 1379430. [Google Scholar] [CrossRef]
- Sung, M.W.; Li, P.C. Chemical analysis of raw, dry-roasted, and honey-roasted licorice by capillary electrophoresis. ELP 2004, 25, 3434–3440. [Google Scholar] [CrossRef]
- Ota, M.; Xu, F.; Li, Y.L.; Shang, M.Y.; Makino, T.; Cai, S.Q. Comparison of chemical constituents among licorice, roasted licorice, and roasted licorice with honey. J. Nat. Med. 2018, 72, 80–95. [Google Scholar] [CrossRef]
- Lee, S.K.; Park, K.K.; Park, J.H.Y.; Lim, S.S.; Chung, W.Y. The inhibitory effect of roasted licorice extract on human metastatic breast cancer cell-induced bone destruction. Phytother. Res. 2013, 27, 1776–1783. [Google Scholar] [CrossRef]
- Maghsoudlou, Y.; Ghajari, M.A.; Tavasoli, S. Effects of heat treatment on the phenolic compounds and antioxidant capacity of quince fruit and its tisane’s sensory properties. J. Food Sci. Technol. 2019, 56, 2365–2372. [Google Scholar] [CrossRef]
- Hwang, I.G.; Kim, H.Y.; Park, B.R.; Han, H.M.; Yoo, S.M. Effect of heat treatment on the antioxidant properties of yacon (Smallanthus sonchifolius). Korean J. Food Nutr. 2013, 26, 857–864. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Yang, K.-M.; Chiang, P.-Y. Roselle anthocyanins: Antioxidant properties and stability to heat and pH. Molecules 2018, 23, 1357. [Google Scholar] [CrossRef]
- Madihah, K.K.; Zaibunnisa, A.; Norashikin, S.; Rozita, O.; Misnawi, J. Optimization of roasting conditions for high-quality Arabica coffee. Int. Food Res. J. 2013, 20, 1623. [Google Scholar]
- Vodnar, D.C.; Călinoiu, L.F.; Dulf, F.V.; Ştefănescu, B.E.; Crişan, G.; Socaciu, C. Identification of the bioactive compounds and antioxidant, antimutagenic and antimicrobial activities of thermally processed agro-industrial waste. Food Chem. 2017, 231, 131–140. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Thermal processing for the release of phenolic compounds from wheat and oat bran. Biomolecules 2020, 10, 21. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, J.Y.; Seo, H.T.; Seong, H.A.; Ji, Y.J.; Lee, S.E.; Seo, K.H.; Kim, H.D. Samnamul (Shoots of Aruncus dioicus) Inhibit Adipogenesis by Downregulating Adipocyte-Specific Transcription Factors in 3T3-L1 Adipocytes. Processes 2020, 8, 1576. [Google Scholar] [CrossRef]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429. [Google Scholar]
- Fazio, A.; Plastina, P.; Meijerink, J.; Witkamp, R.F.; Gabriele, B. Comparative analyses of seeds of wild fruits of Rubus and Sambucus species from Southern Italy: Fatty acid composition of the oil, total phenolic content, antioxidant and anti-inflammatory properties of the methanolic extracts. Food Chem. 2013, 140, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.H.; Xu, S.P. Juglanin administration protects skin against UVB-induced injury by reducing Nrf2-dependent ROS generation. Int. J. Mol. Med. 2020, 46, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.H.; Kim, S.Y. Resveratrol-enriched rice attenuates UVB-ROS-induced skin aging via downregulation of inflammatory cascades. Oxi. Med. Cell. Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef] [PubMed]
- Kremer, J.L.; Melo, G.P.; Marinello, P.C.; Bordini, H.P.; Rossaneis, A.C.; Sábio, L.R.; Cecchini, R.; Cecchini, A.L.; Verri, W.A., Jr.; Luiz, R.C. Citral prevents UVB-induced skin carcinogenesis in hairless mice. J. Photochem. Photobiol. B 2019, 198, 111565. [Google Scholar] [CrossRef]
- Tang, S.C.; Liao, P.Y.; Hung, S.J.; Ge, J.S.; Chen, S.M.; Lai, J.C.; Hsiao, Y.P.; Yang, J.H. Topical application of glycolic acid suppresses the UVB induced IL-6, IL-8, MCP-1 and COX-2 inflammation by modulating NF-κB signaling pathway in keratinocytes and mice skin. J. Dermatol. Sci. 2017, 86, 238–248. [Google Scholar] [CrossRef]
- Aydogan-Coskun, B.; Coklar, H.; Akbuluta, M. Effect of heat treatment for liquefaction and pasteurization on antioxidant activity and phenolic compounds of Astragalus and sunflower-cornflower honeys. Food Sci. 2020, 40, 629–634. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, J.Y.; Park, J.Y.; Jang, G.Y.; Kim, H.D.; Lee, Y.S.; Kim, D.H. Differences in anti-inflammatory effect of immature and mature of Rubus coreanus fruits on LPS-induced RAW 264.7 macrophages via NF-κB signal pathways. BMC Complement. Altern. Med. 2019, 19, 89. [Google Scholar] [CrossRef]
- Kumar, N.; Gupta, S.; Chand Yadav, T.; Pruthi, V.; Kumar Varadwaj, P.; Goel, N. Extrapolation of phenolic compounds as multi-target agents against cancer and inflammation. DYN 2019, 37, 2355–2369. [Google Scholar] [CrossRef]
- Hudson da Silva Souza, I.; Nogueira, J.P.; Rajan, M.; Santos Leite Neta, M.T.; Narain, N. Babassu Oil (Orbynia phalerata), an Artisanal Product: Process Optimization of Seed Roasting on Yield, Phenolic Compounds, and Antioxidant Capacity. Eur. J. Lipid Sci. Technol. 2021, 123, 2000163. [Google Scholar] [CrossRef]
- Starowicz, M.; Ostaszyk, A.; Zielinski, H. The relationship between the browning index, total phenolics, color, and antioxidant activity of polish-originated honey samples. Foods 2021, 10, 967. [Google Scholar] [CrossRef]
- Xu, G.H.; Ye, X.Q.; Chen, J.C.; Liu, D.H. Effect of heat treatment on the phenolic compounds and antioxidant capacity of citrus peel extract. J. Agric. Food Chem. 2007, 55, 330–335. [Google Scholar] [CrossRef]
- Ji, B.; Guo, W.; Ma, H.; Xu, B.; Mu, W.; Zhang, Z.; Amat, A.; Cao, L. Isoliquiritigenin suppresses IL-1β induced apoptosis and inflammation in chondrocyte-like ATDC5 cells by inhibiting NF-κB and exerts chondroprotective effects on a mouse model of anterior cruciate ligament transection. Int. J. Mol. Med. 2017, 40, 1709–1718. [Google Scholar] [CrossRef]
- Ye, H.; Yang, X.; Chen, X.; Shen, L.; Le, R. Isoliquiritigenin protects against angiotensin II-induced fibrogenesis by inhibiting NF-κB/PPARγ inflammatory pathway in human Tenon’s capsule fibroblasts. Exp. Eye. Res. 2020, 199, 108146. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| IL-1β | F 1 | GTATCACTCATTGTGGCTGTG |
| R 2 | ATTTTGTCGTTGCTTGGTTCTC | |
| IL-6 | F | ATTACACATGTTCTCTGGGAAG |
| R | TTTTACCTCTTGGTTGAAGATATG | |
| GAPDH | F | AAAAGGGTCATCATCTCCGC |
| R | CTTCTTGATGTCATCATACTTGG | |
| Samples | DPPH (IC50, μg/mL) | ABTS + (IC50, μg/mL) |
|---|---|---|
| NL | 770.3 ± 42.3 a | 63.2 ± 0.8 a |
| HL-1 | 455.0 ± 36.5 b | 60.5 ± 1.0 b |
| HL-2 | 299.2 ± 10.8 c | 43.7 ± 0.8 c |
| Samples | Total Phenolic Content (mg GA/g Extract) | Isoliquiritigenin (mg/g Extract) |
|---|---|---|
| NL | 11.7 ± 0.7 b | 2.40 ± 0.24 c |
| HL-1 | 12.1 ± 0.6 b | 2.82 ± 0.14 b |
| HL-2 | 14.4 ± 0.6 a | 3.28 ± 0.18 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-Y.; Ji, Y.-J.; Seo, K.H.; Lee, J.Y.; Kim, G.-S.; Kang, M.H.; Lee, J.H.; Jang, G.Y.; Kim, H.D. Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts. Processes 2021, 9, 1040. https://doi.org/10.3390/pr9061040
Park J-Y, Ji Y-J, Seo KH, Lee JY, Kim G-S, Kang MH, Lee JH, Jang GY, Kim HD. Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts. Processes. 2021; 9(6):1040. https://doi.org/10.3390/pr9061040
Chicago/Turabian StylePark, Jeong-Yong, Yun-Jeong Ji, Kyung Hye Seo, Ji Yeon Lee, Geum-Soog Kim, Min Hye Kang, Jeong Hoon Lee, Gwi Yeong Jang, and Hyung Don Kim. 2021. "Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts" Processes 9, no. 6: 1040. https://doi.org/10.3390/pr9061040
APA StylePark, J.-Y., Ji, Y.-J., Seo, K. H., Lee, J. Y., Kim, G.-S., Kang, M. H., Lee, J. H., Jang, G. Y., & Kim, H. D. (2021). Heat Treatment Improves UV Photoprotective Effects of Licorice in Human Dermal Fibroblasts. Processes, 9(6), 1040. https://doi.org/10.3390/pr9061040