
Differentially Expressed Genes Correlated with Fibrosis in a Rat Model of Chronic Partial Bladder Outlet Obstruction



Abstract
:1. Introduction
2. Materials and Methods
2.1. PBOO Animal Model
2.2. Bladder Tissue Preparation and RNA Preparation
2.3. Rat OneArray Plus Microarrays
2.4. Data Analysis
3. Results
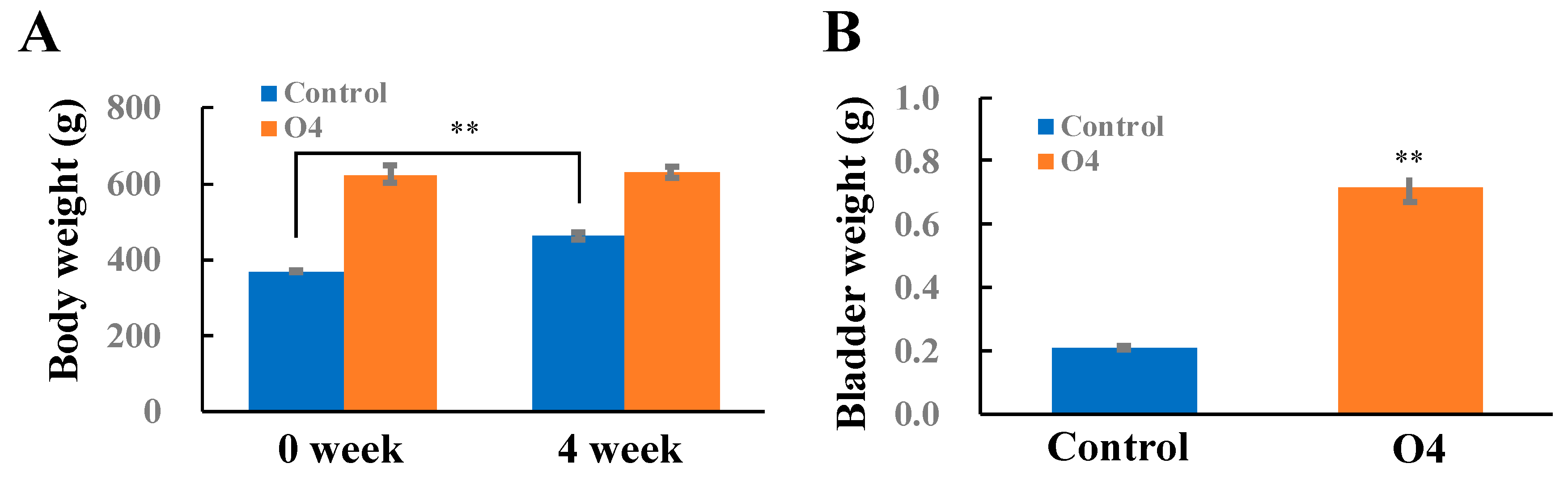
PBOO Increased Bladder Weight and Induced DEG Alterations Involved in Fibrosis, Downstream TGFB, and Extracellular Matrix
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Microarray Quality Control | ||
|---|---|---|
| Check Items | Description | Specification |
| Background arerage intensity | Cy5:B635 Median | Cy5 < 100 |
| Intrinsic Hybridization Controls (IHCs) | IHC Probes designed for monitoring overall hybridization efficiency | IHC < 15,000 IHC CV < 15% |
| Extrinsic Target Quality Control (ETQC) with Spikiins | ETQC probes and labeled spike-ins designed for monitoring the specificity of hybridization at low, medium, and high concentrations | High, Median, Low |
| Noise | 48 negative probes used in Rosetta Resolver for monitoring non-specific hybridization | Number of negative intensity > 200 should be less than 10 |
| Sample integrity in processing | Intrinsic Target Quality Control (ITQC) 3, 6, 7 probes designed for monitoring the sample integrity | Number of ITQC rations within spec (S1/S2 * < 3.5) >= 2 |
| Technical reproducibility | Pearson’s correlation coefficient between technical replicates | R value >= 0.975 |
| Il6 and Stat3 Signal Related Genes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Ldha | Socs3 | Ccl12 | Osmr | Nfkb1 | Il11 | Ccl5 | Cxcl12 | Cdc25a |
| Fold | 3.9 | 2.9 | 2.6 | 2.4 | 2.0 | 2.0 | 0.28 | 0.30 | 0.39 |
References
- Rosen, R.; Altwein, J.; Boyle, P.; Kirby, R.S.; Lukacs, B.; Meuleman, E.; O’Leary, M.P.; Puppo, P.; Robertson, C.; Giuliano, F. Lower urinary tract symptoms and male sexual dysfunction: The multinational survey of the aging male (MSAM-7). Eur. Urol. 2003, 44, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Vallancien, G.; Emberton, M.; Harving, N.; van Moorselaar, R.J.; Alf-One Study, G. Sexual dysfunction in 1274 European men suffering from lower urinary tract symptoms. J. Urol. 2003, 169, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Landau, E.H.; Jayanthi, V.R.; Churchill, B.M.; Shapiro, E.; Gilmour, R.F.; Khoury, A.E.; Macarak, E.J.; McLorie, G.A.; Steckler, R.E.; Kogan, B.A. Loss of elasticity in dysfunctional bladders: Urodynamic and histochemical correlation. J. Urol. 1994, 152 Pt 2, 702–705. [Google Scholar] [CrossRef]
- Palmer, L.S.; Lee, C.; Decker, R.S.; Lang, S.; Kaplan, W.E.; Firlit, C.F.; Cheng, E.Y. The effect of angiotensin converting enzyme inhibition and angiotensin II receptor antagonism on obstructed rat bladder. J. Urol. 1997, 158 Pt 2, 1100–1104. [Google Scholar] [CrossRef]
- Anumanthan, G.; Tanaka, S.T.; Adams, C.M.; Thomas, J.C.; Wills, M.L.; Adams, M.C.; Hayward, S.W.; Matusik, R.J.; Bhowmick, N.A.; Brock, J.W., 3rd; et al. Bladder stromal loss of transforming growth factor receptor II decreases fibrosis after bladder obstruction. J. Urol. 2009, 182 (Suppl. 4), 1775–1780. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Takeuchi, C.; Oyunzul, L.; Ito, Y. Bladder angiotensin-II receptors: Characterization and alteration in bladder outlet obstruction. Eur. Urol. 2009, 55, 482–489. [Google Scholar] [CrossRef]
- Persson, K.; Pandita, R.K.; Waldeck, K.; Andersson, K.E. Angiotensin II and bladder obstruction in the rat: Influence on hypertrophic growth and contractility. Am. J. Physiol. 1996, 271 Pt 2, R1186–R1192. [Google Scholar] [CrossRef]
- Myers, J.B.; Dall’era, J.E.; Koul, S.; Kumar, B.; Khandrika, L.; Flynn, B.J.; Koul, H.K. Biochemical alterations in partial bladder outlet obstruction in mice: Up-regulation of the mitogen activated protein kinase pathway. J. Urol. 2009, 181, 1926–1931. [Google Scholar] [CrossRef]
- Tanaka, S.T.; Martinez-Ferrer, M.; Makari, J.H.; Wills, M.L.; Thomas, J.C.; Adams, M.C.; Brock, J.W., 3rd; Pope, J.C.t.; Bhowmick, N.A. Recruitment of bone marrow derived cells to the bladder after bladder outlet obstruction. J. Urol. 2009, 182 (Suppl. 4), 1769–1774. [Google Scholar] [CrossRef]
- Yang, T.X.; Luo, D.Y.; Lin, Y.F.; Liu, Q.; Cai, X.; Ai, J.Z.; Li, H.; Shen, H.; Wang, K.J. The Antimuscarinic Agent Tolterodine Regulates Bladder Extracellular Matrix in Partial Bladder Outlet Obstruction in Rats. Cell Physiol. Biochem. 2018, 46, 36–45. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, P.; He, F.; Yang, X.; Wu, R.; Chen, W.; Li, L.; Yang, Z. Fibroblast Growth Factor 2 Promotes Bladder Hypertrophy Caused by Partial Bladder Outlet Obstruction. Front. Cell Dev. Biol. 2021, 9, 630228. [Google Scholar] [CrossRef]
- Lin, W.Y.; Lin, Y.P.; Levin, R.M.; Chen, M.L. The relevance of immune responses to partial bladder outlet obstruction and reversal. Neurourol. Urodyn. 2017, 36, 1306–1312. [Google Scholar] [CrossRef]
- Frangogiannis, N.G.; Dewald, O.; Xia, Y.; Ren, G.; Haudek, S.; Leucker, T.; Kraemer, D.; Taffet, G.; Rollins, B.J.; Entman, M.L. Critical role of monocyte chemoattractant protein-1/CC chemokine ligand 2 in the pathogenesis of ischemic cardiomyopathy. Circulation 2007, 115, 584–592. [Google Scholar] [CrossRef]
- Cohn, J.N.; Ferrari, R.; Sharpe, N. Cardiac remodeling–concepts and clinical implications: A consensus paper from an international forum on cardiac remodeling. Behalf of an International Forum on Cardiac Remodeling. J. Am. Coll Cardiol. 2000, 35, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Berk, B.C.; Fujiwara, K.; Lehoux, S. ECM remodeling in hypertensive heart disease. J. Clin. Investig. 2007, 117, 568–575. [Google Scholar] [CrossRef]
- Swaney, J.S.; Roth, D.M.; Olson, E.R.; Naugle, J.E.; Meszaros, J.G.; Insel, P.A. Inhibition of cardiac myofibroblast formation and collagen synthesis by activation and overexpression of adenylyl cyclase. Proc. Natl. Acad. Sci. USA 2005, 102, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Shimokawa, H.; Takeshita, A. Rho-kinase is an important therapeutic target in cardiovascular medicine. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1767–1775. [Google Scholar] [CrossRef]
- Wang, Y.X.; da Cunha, V.; Martin-McNulty, B.; Vincelette, J.; Li, W.; Choy, D.F.; Halks-Miller, M.; Mahmoudi, M.; Schroeder, M.; Johns, A.; et al. Inhibition of Rho-kinase by fasudil attenuated angiotensin II-induced cardiac hypertrophy in apolipoprotein E deficient mice. Eur. J. Pharmacol. 2005, 512, 215–222. [Google Scholar] [CrossRef]
- Sorescu, D. Smad3 mediates angiotensin II- and TGF-beta1-induced vascular fibrosis: Smad3 thickens the plot. Circ. Res. 2006, 98, 988–989. [Google Scholar] [CrossRef] [Green Version]
- Guven, A.; Onal, B.; Kalorin, C.; Whitbeck, C.; Chichester, P.; Kogan, B.; Levin, R.; Mannikarottu, A. Long term partial bladder outlet obstruction induced contractile dysfunction in male rabbits: A role for Rho-kinase. Neurourol. Urodyn. 2007, 26, 1043–1049. [Google Scholar] [CrossRef]
- Taylor, S.H.; Yeung, C.Y.; Kalson, N.S.; Lu, Y.; Zigrino, P.; Starborg, T.; Warwood, S.; Holmes, D.F.; Canty-Laird, E.G.; Mauch, C.; et al. Matrix metalloproteinase 14 is required for fibrous tissue expansion. Elife 2015, 4, e09345. [Google Scholar] [CrossRef]
- Xiao, T.; Zeng, O.; Luo, J.; Wu, Z.; Li, F.; Yang, J. Effects of hydrogen sulfide on myocardial fibrosis in diabetic rats: Changes in matrix metalloproteinases parameters. Biomed. Mater. Eng. 2015, 26 (Suppl. 1), S2033–S2039. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L.; Zeng, O.; Liu, J.M.; Yang, J. H2S improves renal fibrosis in STZ-induced diabetic rats by ameliorating TGF-beta1 expression. Ren. Fail. 2017, 39, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Spinale, F.G. Bioactive peptide signaling within the myocardial interstitium and the matrix metalloproteinases. Circ. Res. 2002, 91, 1082–1084. [Google Scholar] [CrossRef] [Green Version]
- Siwik, D.A.; Pagano, P.J.; Colucci, W.S. Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. Am. J. Physiol. Cell Physiol. 2001, 280, C53–C60. [Google Scholar] [CrossRef]
- Lin, W.Y.; Guven, A.; Juan, Y.S.; Neuman, P.; Whitbeck, C.; Chichester, P.; Kogan, B.; Levin, R.M.; Mannikarottu, A. Free radical damage as a biomarker of bladder dysfunction after partial outlet obstruction and reversal. BJU Int. 2008, 101, 621–626. [Google Scholar] [CrossRef]
- Takimoto, E.; Kass, D.A. Role of oxidative stress in cardiac hypertrophy and remodeling. Hypertension 2007, 49, 241–248. [Google Scholar] [CrossRef]
- Al-Saikan, B.; Ding, J.; Tredget, E.; Metcalfe, P. Benefits of mesenchymal stem cells after partial bladder outlet obstruction. Can Urol. Assoc. J. 2016, 10, E1–E6. [Google Scholar] [CrossRef] [Green Version]
- Wiafe, B.; Kadam, R.; Metcalfe, P.D. Intraperitoneal administration of mesenchymal stem cells is effective at mitigating detrusor deterioration after pBOO. Am. J. Physiol. Renal. Physiol. 2020, 318, F549–F556. [Google Scholar] [CrossRef]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, N.; Donmez, M.I.; Malykhina, A.P.; Carrasco, A., Jr.; Wilcox, D.T. Preventative effects of a HIF inhibitor, 17-DMAG, on partial bladder outlet obstruction-induced bladder dysfunction. Am. J. Physiol. Renal. Physiol. 2017, 313, F1149–F1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Chen, Y.; Zhu, H.; Wang, B.; Qu, P.; Chen, R.; Sun, X. Sodium Tanshinone IIA Sulfonate Ameliorates Bladder Fibrosis in a Rat Model of Partial Bladder Outlet Obstruction by Inhibiting the TGF-beta/Smad Pathway Activation. PLoS ONE 2015, 10, e0129655. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Fibrosis-Related Genes | |||||||||||
| Gene | Grem1 | Thbs1 | Lox | Serpine1 | Timp1 | Itga1 | Tgfb3 | Ldha | Ilk | Timp3 | Timp3 |
| Fold | 82.6 | 25.8 | 6.3 | 6.2 | 6.2 | 4.5 | 4.5 | 3.9 | 2.8 | 2.8 | 2.8 |
| Gene | Ccl12 | Mmp14 | Ltbp1 | Stat6 | Itgb | Nfkb | Thbs2 | Stat1 | Bmp7 | ||
| Fold | 2.6 | 2.4 | 2.4 | 2.2 | 2.2 | 2.0 | 0.24 | 0.47 | 0.48 | ||
| TgfbSignaling-Related Genes | |||||||||||
| Gene | Thbs1 | Serpine1 | Bhlhe40 | Gadd45b | Ldha | Hmox1 | Ptgs2 | Fn1 | Ptk2 | Cryab | Ctnnb1 |
| Fold | 25.8 | 6.2 | 5.0 | 4.5 | 3.9 | 3.3 | 3.2 | 2.7 | 2.7 | 2.6 | 2.2 |
| Gene | Fos | Sox4 | Srebf2 | Rybp | Bcl2l1 | Notch1 | Id2 | Ptk2b | Gtf2i | Msx2 | Herpud1 |
| Fold | 2.1 | 2.0 | 0.33 | 0.35 | 0.36 | 0.37 | 0.39 | 0.40 | 0.41 | 0.44 | 0.45 |
| Gene | Acvrl1 | Crebbp | Cdkn1b | ||||||||
| Fold | 0.47 | 0.49 | 0.49 | ||||||||
| Extracellular Matrix-Related Genes | |||||||||||
| Gene | Thbs1 | Col8a1 | Itga5 | Tnc | Timp1 | Col4a1 | Col4a2 | Ldha | Emilin1 | Col5a1 | Ncam1 |
| Fold | 25.8 | 15.2 | 9.3 | 6.6 | 6.2 | 6.1 | 5.7 | 3.9 | 3.5 | 3.3 | 3.2 |
| Gene | Lamc1 | Cd44 | Vcan | Timp3 | Fn1 | Adamts1 | Mmp14 | Ctnnb1 | Spock1 | Itgb1 | Lama1 |
| Fold | 3.0 | 2.8 | 2.8 | 2.8 | 2.7 | 2.6 | 2.4 | 2.2 | 2.2 | 2.2 | 2.0 |
| Gene | Thbs2 | Tgfbi | Vcam1 | Cdh1 | Mmp15 | Mmp11 | Mmp16 | Pecam1 | Cdh3 | ||
| Fold | 0.24 | 0.26 | 0.28 | 0.30 | 0.35 | 0.38 | 0.42 | 0.45 | 0.45 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, Y.-S.; Chang, H.H.; Ko, S.-Y.; Lin, Y.-P.; Lin, W.-Y. Differentially Expressed Genes Correlated with Fibrosis in a Rat Model of Chronic Partial Bladder Outlet Obstruction. Processes 2021, 9, 2219. https://doi.org/10.3390/pr9122219
Hsueh Y-S, Chang HH, Ko S-Y, Lin Y-P, Lin W-Y. Differentially Expressed Genes Correlated with Fibrosis in a Rat Model of Chronic Partial Bladder Outlet Obstruction. Processes. 2021; 9(12):2219. https://doi.org/10.3390/pr9122219
Chicago/Turabian StyleHsueh, Yuan-Shuo, Hui Hua Chang, Shun-Yao Ko, Yi-Pai Lin, and Wei-Yu Lin. 2021. "Differentially Expressed Genes Correlated with Fibrosis in a Rat Model of Chronic Partial Bladder Outlet Obstruction" Processes 9, no. 12: 2219. https://doi.org/10.3390/pr9122219
APA StyleHsueh, Y.-S., Chang, H. H., Ko, S.-Y., Lin, Y.-P., & Lin, W.-Y. (2021). Differentially Expressed Genes Correlated with Fibrosis in a Rat Model of Chronic Partial Bladder Outlet Obstruction. Processes, 9(12), 2219. https://doi.org/10.3390/pr9122219