Control of Specific Growth Rate in Fed-Batch Bioprocesses: Novel Controller Design for Improved Noise Management


Abstract
1. Introduction
2. Materials and Methods
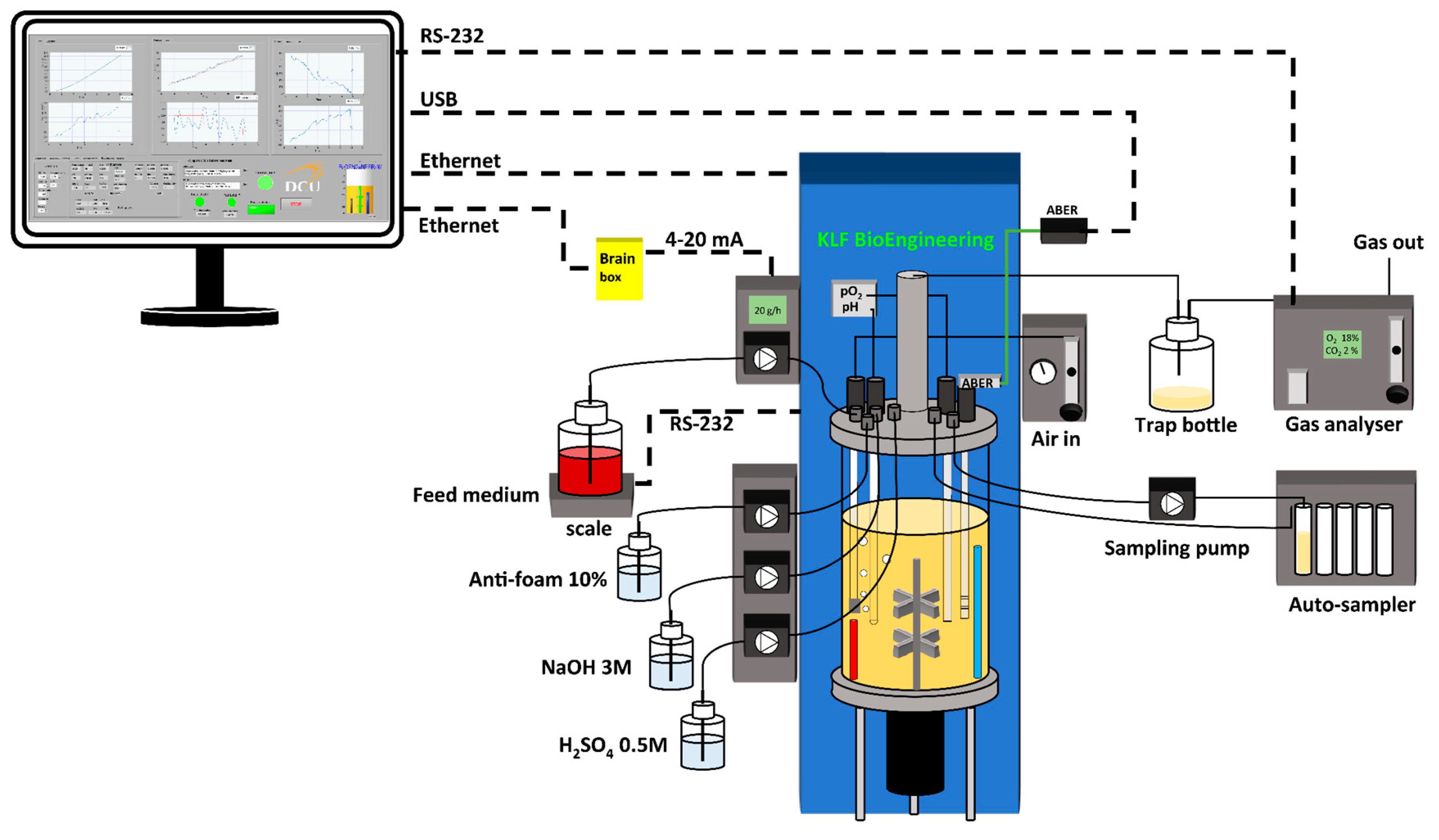
2.1. Cell Strain and Culture Conditions
2.2. Signal Filtering and Smoothing
2.3. Dielectric Spectroscopy Adaptive Calibration
2.4. Specific Growth Rate Estimation
2.5. Controller Design
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mears, L.; Stocks, S.M.; Sin, G.; Gernaey, K.V. A review of control strategies for manipulating the feed rate in fed-batch fermentation processes. J. Biotechnol. 2017, 245, 34–46. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.Y.; Park, S.; Middelberg, A.P.J. Control of fed-batch fermentations. Biotechnol. Adv. 1999, 17, 29–48. [Google Scholar] [CrossRef]
- Kim, B.S.; Lee, S.C.; Lee, S.Y.; Chang, Y.K.; Chang, H.N. High cell density fed-batch cultivation of Escherichia coli using exponential feeding combined with pH-stat. Bioprocess Biosyst. Eng. 2004, 26, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Johnston, W.; Cord-Ruwisch, R.; Cooney, M.J. Industrial control of recombinant E. coli fed-batch culture: New perspectives on traditional controlled variables. Bioprocess Biosyst. Eng. 2002, 25, 111–120. [Google Scholar]
- Habegger, L.; Rodrigues Crespo, K.; Dabros, M. Preventing Overflow Metabolism in Crabtree-Positive Microorganisms through On-Line Monitoring and Control of Fed-Batch Fermentations. Fermentation 2018, 4, 79. [Google Scholar] [CrossRef]
- Kesson, M.; Hagander, P.; Axelsson, J.P. Avoiding acetate accumulation in Escherichia coli cultures using feedback control of glucose feeding. Biotechnol. Bioeng. 2001, 73, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Graham, C.; Ciccarelli, R.B. Automated fed-batch fermentation with feed-back controls based on dissolved oxygen (DO) and pH for production of DNA vaccines. J. Ind. Microbiol. Biotechnol. 1997, 18, 43–48. [Google Scholar] [CrossRef]
- Soons, Z.I.T.A.; Voogt, J.A.; van Straten, G.; van Boxtel, A.J.B. Constant specific growth rate in fed-batch cultivation of Bordetella pertussis using adaptive control. J. Biotechnol. 2006, 125, 252–268. [Google Scholar] [CrossRef]
- Junker, B.H.; Wang, H.Y. Bioprocess monitoring and computer control: Key roots of the current PAT initiative. Biotechnol. Bioeng. 2006, 95, 226–261. [Google Scholar] [CrossRef]
- Jenzsch, M.; Simutis, R.; Eisbrenner, G.; Stückrath, I.; Lübbert, A. Estimation of biomass concentrations in fermentation processes for recombinant protein production. Bioprocess Biosyst. Eng. 2006, 29, 19–27. [Google Scholar] [CrossRef]
- Claes, J.E.; Van Impe, J.F. Combining yield coefficients and exit-gas analysis for monitoring of the baker’s yeast fed-batch fermentation. Bioprocess Eng. 2000, 22, 195–200. [Google Scholar] [CrossRef]
- Chen, L.Z.; Nguang, S.K.; Li, X.M.; Chen, X.D. Soft sensors for on-line biomass measurements. Bioprocess Biosyst. Eng. 2004, 26, 191–195. [Google Scholar]
- Luttmann, R.; Bracewell, D.G.; Cornelissen, G.; Gernaey, K.V.; Glassey, J.; Hass, V.C.; Kaiser, C.; Preusse, C.; Striedner, G.; Mandenius, C.F. Soft sensors in bioprocessing: A status report and recommendations. Biotechnol. J. 2012, 7, 1040–1048. [Google Scholar] [CrossRef]
- Kiviharju, K.; Salonen, K.; Moilanen, U.; Eerikäinen, T. Biomass measurement online: The performance of in situ measurements and software sensors. J. Ind. Microbiol. Biotechnol. 2008, 35, 657–665. [Google Scholar] [CrossRef]
- Arnoux, A.S.; Preziosi-Belloy, L.; Esteban, G.; Teissier, P.; Ghommidh, C. Lactic acid bacteria biomass monitoring in highly conductive media by permittivity measurements. Biotechnol. Lett. 2005, 27, 1551–1557. [Google Scholar] [CrossRef]
- Schuler, M.M.; Marison, I.W. Real-time monitoring and control of microbial bioprocesses with focus on the specific growth rate: Current state and perspectives. Appl. Microbiol. Biotechnol. 2012, 94, 1469–1482. [Google Scholar] [CrossRef]
- Kiviharju, K.; Salonen, K.; Moilanen, U.; Meskanen, E.; Leisola, M.; Eerikäinen, T. On-line biomass measurements in bioreactor cultivations: Comparison study of two on-line probes. J. Ind. Microbiol. Biotechnol. 2007, 34, 561–566. [Google Scholar] [CrossRef]
- Austin, G.D.; Watson, R.W.J.; D’Amore, T. Studies of on-line viable yeast biomass with a capacitance biomass monitor. Biotechnol. Bioeng. 1994, 43, 337–341. [Google Scholar] [CrossRef]
- Biechele, P.; Busse, C.; Solle, D.; Scheper, T.; Reardon, K. Sensor systems for bioprocess monitoring. Eng. Life Sci. 2015, 15, 469–488. [Google Scholar] [CrossRef]
- Olsson, L.; Nielsen, J. On-line and in situ monitoring of biomass in submerged cultivations. Trends Biotechnol. 1997, 15, 517–522. [Google Scholar] [CrossRef]
- Schuler, M.M.; Sivaprakasam, S.; Freeland, B.; Hama, A.; Hughes, K.M.; Marison, I.W. Investigation of the potential of biocalorimetry as a process analytical technology (PAT) tool for monitoring and control of Crabtree-negative yeast cultures. Appl. Microbiol. Biotechnol. 2012, 93, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Araki, K.; Shioya, S.; Suga, K.I. Optimal production of glutathione by controlling the specific growth rate of yeast in fed-batch culture. Biotechnol. Bioeng. 1991, 38, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Ehgartner, D.; Hartmann, T.; Heinzl, S.; Frank, M.; Veiter, L.; Kager, J.; Herwig, C.; Fricke, J. Controlling the specific growth rate via biomass trend regulation in filamentous fungi bioprocesses. Chem. Eng. Sci. 2017, 172, 32–41. [Google Scholar] [CrossRef]
- Dabros, M.; Schuler, M.M.; Marison, I.W. Simple control of specific growth rate in biotechnological fed-batch processes based on enhanced online measurements of biomass. Bioprocess Biosyst. Eng. 2010, 33, 1109–1118. [Google Scholar] [CrossRef]
- Gnoth, S.; Jenzsch, M.; Simutis, R.; Lübbert, A. Control of cultivation processes for recombinant protein production: A review. Bioprocess Biosyst. Eng. 2008, 31, 21–39. [Google Scholar] [CrossRef]
- Riesenberg, D.; Schulz, V.; Knorre, W.A.; Pohl, H.D.; Korz, D.; Sanders, E.A.; Roß, A.; Deckwer, W.D. High cell density cultivation of Escherichia coli at controlled specific growth rate. J. Biotechnol. 1991, 20, 17–27. [Google Scholar] [CrossRef]
- Marison, I.; Hennessy, S.; Foley, R.; Schuler, M.; Sivaprakasam, S.; Freeland, B. The choice of suitable online analytical techniques and data processing for monitoring of bioprocesses. Adv. Biochem. Eng. Biotechnol. 2013, 132, 249–280. [Google Scholar]
- Liu, C.; Gong, Z.; Shen, B.; Feng, E. Modelling and optimal control for a fed-batch fermentation process. Appl. Math. Model. 2013, 37, 695–706. [Google Scholar] [CrossRef]
- Galvanauskas, V.; Simutis, R.; Vaitkus, V. Adaptive control of biomass specific growth rate in fed-batch biotechnological processes. A comparative study. Processes 2019, 7, 810. [Google Scholar] [CrossRef]
- Mohan, N.; Sivaprakasam, S. Heat Compensation Calorimeter as a Process Analytical Tool to Monitor and Control Bioprocess Systems. Ind. Eng. Chem. Res. 2017, 56, 8416–8427. [Google Scholar] [CrossRef]
- Steinwandter, V.; Zahel, T.; Sagmeister, P.; Herwig, C. Propagation of measurement accuracy to biomass soft-sensor estimation and control quality. Anal. Bioanal. Chem. 2017, 409, 693–706. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sagmeister, P.; Wechselberger, P.; Jazini, M.; Meitz, A.; Langemann, T.; Herwig, C. Soft sensor assisted dynamic bioprocess control: Efficient tools for bioprocess development. Chem. Eng. Sci. 2013, 96, 190–198. [Google Scholar] [CrossRef]
- Paulsson, D.; Gustavsson, R.; Mandenius, C.F. A soft sensor for bioprocess control based on sequential filtering of metabolic heat signals. Sensors (Switzerland) 2014, 14, 17864–17882. [Google Scholar] [CrossRef] [PubMed]
- Metze, S.; Ruhl, S.; Greller, G.; Grimm, C.; Scholz, J. Monitoring online biomass with a capacitance sensor during scale-up of industrially relevant CHO cell culture fed-batch processes in single-use bioreactors. Bioprocess Biosyst. Eng. 2020, 43, 193–205. [Google Scholar] [CrossRef]
- Moore, B.; Sanford, R.; Zhang, A. Case study: The characterization and implementation of dielectric spectroscopy (biocapacitance) for process control in a commercial GMP CHO manufacturing process. Biotechnol. Prog. 2019, 35, e2782. [Google Scholar] [CrossRef]
- Justice, C.; Brix, A.; Freimark, D.; Kraume, M.; Pfromm, P.; Eichenmueller, B.; Czermak, P. Process control in cell culture technology using dielectric spectroscopy. Biotechnol. Adv. 2011, 29, 391–401. [Google Scholar] [CrossRef]
- Carvell, J.; Lee, M.; Bhat, A.R. Recent developments in scaling down and using single use probes for measuring the live cell concentration by dielectric spectroscopy. BMC Proc. 2015, 9, P46. [Google Scholar] [CrossRef]
- Voisard, D.; Pugeaud, P.; Kumar, A.R.; Jenny, K.; Jayaraman, K.; Marison, I.W.; Von Stockar, U. Development of a large-scale biocalorimeter to monitor and control bioprocesses. Biotechnol. Bioeng. 2002, 80, 125–138. [Google Scholar] [CrossRef]
- Cole, H.E.; Demont, A.; Marison, I.W. The application of dielectric spectroscopy and biocalorimetry for the monitoring of biomass in immobilized mammalian cell cultures. Processes 2015, 3, 384–405. [Google Scholar] [CrossRef]
- Urit, T.; Li, M.; Bley, T.; Löser, C. Growth of Kluyveromyces marxianus and formation of ethyl acetate depending on temperature. Appl. Microbiol. Biotechnol. 2013, 97, 10359–10371. [Google Scholar] [CrossRef]
- Löser, C.; Urit, T.; Keil, P.; Bley, T. Studies on the mechanism of synthesis of ethyl acetate in Kluyveromyces marxianus DSM 5422. Appl. Microbiol. Biotechnol. 2015, 99, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- GRANT, C.L.; PRAMER, D. Minor element composition of yeast extract. J. Bacteriol. 1962, 84, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Urit, T.; Stukert, A.; Bley, T.; Löser, C. Formation of ethyl acetate by Kluyveromyces marxianus on whey during aerobic batch cultivation at specific trace element limitation. Appl. Microbiol. Biotechnol. 2012, 96, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Dabros, M.; Dennewald, D.; Currie, D.J.; Lee, M.H.; Todd, R.W.; Marison, I.W.; Von Stockar, U. Cole-Cole, linear and multivariate modeling of capacitance data for on-line monitoring of biomass. Bioprocess Biosyst. Eng. 2009, 32, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.; Cooper, D. A novel pattern-based approach for diagnostic controller performance monitoring. Control Eng. Pract. 2010, 18, 279–288. [Google Scholar] [CrossRef]
- Meshram, P.M.; Kanojiya, R.G. Tuning of PID controller using Ziegler-Nichols method for speed control of DC motor. In Proceedings of the IEEE-international conference on advances in engineering, science and management (ICAESM-2012), Nagapattinam, India, 30–31 March 2012; pp. 117–122. [Google Scholar]
- Löser, C.; Urit, T.; Stukert, A.; Bley, T. Formation of ethyl acetate from whey by Kluyveromyces marxianus on a pilot scale. J. Biotechnol. 2013, 163, 17–23. [Google Scholar] [CrossRef]
- Urit, T.; Löser, C.; Wunderlich, M.; Bley, T. Formation of ethyl acetate by Kluyveromyces marxianus on whey: Studies of the ester stripping. Bioprocess Biosyst. Eng. 2011, 34, 547–559. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Batch Medium g/L | Feed Medium g/L |
|---|---|---|
| Lactose | 5 | 300 |
| (NH4)2SO4 | 5 | 50 |
| KH2PO4 | 3 | 25 |
| MgSO4·7H2O | 0.5 | 3 |
| Peptone | 5 | 25 |
| Yeast extract | 5 | 25 |
| Antifoam | 2 | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brignoli, Y.; Freeland, B.; Cunningham, D.; Dabros, M. Control of Specific Growth Rate in Fed-Batch Bioprocesses: Novel Controller Design for Improved Noise Management. Processes 2020, 8, 679. https://doi.org/10.3390/pr8060679
Brignoli Y, Freeland B, Cunningham D, Dabros M. Control of Specific Growth Rate in Fed-Batch Bioprocesses: Novel Controller Design for Improved Noise Management. Processes. 2020; 8(6):679. https://doi.org/10.3390/pr8060679
Chicago/Turabian StyleBrignoli, Yann, Brian Freeland, David Cunningham, and Michal Dabros. 2020. "Control of Specific Growth Rate in Fed-Batch Bioprocesses: Novel Controller Design for Improved Noise Management" Processes 8, no. 6: 679. https://doi.org/10.3390/pr8060679
APA StyleBrignoli, Y., Freeland, B., Cunningham, D., & Dabros, M. (2020). Control of Specific Growth Rate in Fed-Batch Bioprocesses: Novel Controller Design for Improved Noise Management. Processes, 8(6), 679. https://doi.org/10.3390/pr8060679