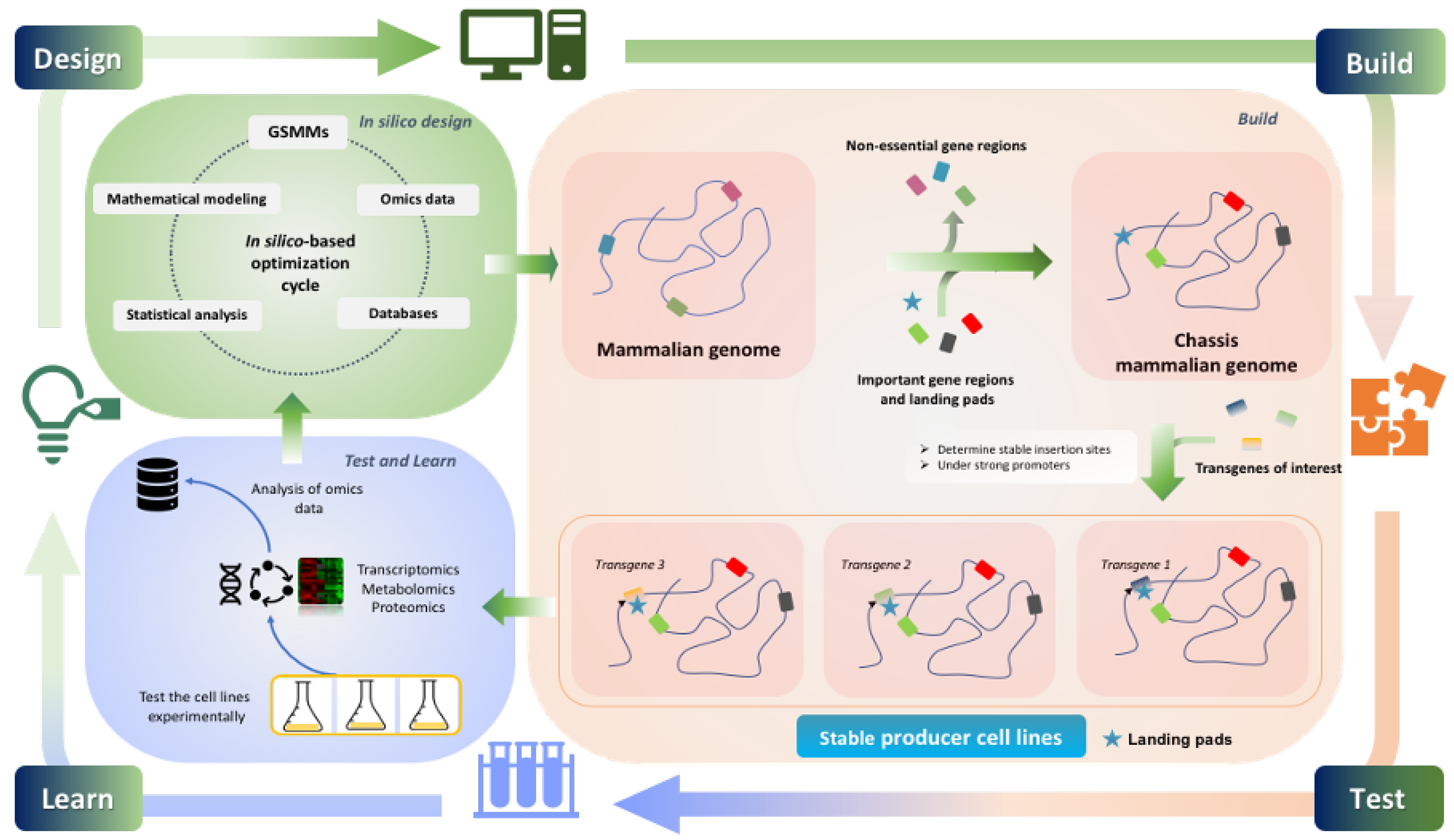
Key Challenges in Designing CHO Chassis Platforms

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Microbial Cell Factories: Current Engineering Paradigms for the Bio-based Production of Chemical Commodities
2.1. Engineering Paradigms
2.2. Model-based Engineering Approaches
2.3. Chassis Strains
2.4. Dynamic Control of Cell Metabolism
2.5. Requirements for Such Strategies to Work
3. Mammalian Expression Systems: CHO Cells
3.1. Importance of Mammalian Cells
3.2. Omics Landscape of CHO
3.3. Comparison of CHO to Bacteria and Yeast

{kind=link}
| Characteristic | E. coli | S. cerevisiae | CHO | References |
|---|---|---|---|---|
| Genome size (Mbp) | 4.6 | 12.1 | 2450 | [104,122,123] |
| Cell size (m) | <1 | 3–5 | 12–24 | [124,125,126] |
| Cell volume (m3) | 0.3–3 | 30–100 | 900–7200 | [125,126,127] |
| N-Linked Glycosylation | No | High Mannose | Complex | [128] |
| Cell culture medium complexity | Low | Low | High | [128] |
| Cost of cell culture medium | Low | Low | High | [128] |
| Doubling time (h) | Rapid (0.5–4) | Rapid (1.5–6) | Slow (18–48) | [75,129,130,131] |
| Number of protein coding genes | ∼4300 | ∼6600 | ∼24,000 | [115,132] |
| Gene length (bp) | ∼1000 | ∼1000 | ∼1300/18,000 † | [115,127] |
| Promoter length (bp) | ∼100 | ∼1000 | ∼104–105 * | [127,133] |
| Proteins per cell | ∼106 | ∼108 | ∼1010 * | [127,131] |
4. Engineering CHO for Improved Performance
4.1. Increasing Cell Specific Productivity and Process Performance
4.2. Maintaining the Balance between Growth and Productivity
4.3. Making Metabolism more Efficient
4.4. Ensuring Product Quality
4.5. Maintaining Stability
5. What Is Missing Towards the Construction of a CHO Chassis Cell?
Author Contributions
Funding
Conflicts of Interest
References
- Walsh, G. Biopharmaceutical benchmarks 2018. Nat. Biotechnol. 2018, 36, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Chan, G.; Hu, Y.; Hu, H.; Ouyang, D. A comprehensive map of FDA-approved pharmaceutical products. Pharmaceutics 2018, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Pohlscheidt, M.; Kiss, R.; Gottschalk, U. An Introduction to Recent Trends in the Biotechnology Industry: Development and Manufacturing of Recombinant Antibodies and Proteins. In New Bioprocessing Strategies: Development and Manufacturing of Recombinant Antibodies and Proteins; Springer: Cham, Switzerland, 2018; pp. 1–8. [Google Scholar]
- Market Research Future Bio process Technology Market to Expand in Size by Highest Revenue up to 2022. Available online: https://www.medgadget.com/2018/08/bio-process-technology-market-to-expand-in-size-by-highest-revenue-up-to-2022-market-research-future.html (accessed on 9 January 2020).
- Bandaranayake, A.D.; Almo, S.C. Recent advances in mammalian protein production. FEBS Lett. 2014, 588, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Harcum, S.W.; Lee, K.H. CHO Cells Can Make More Protein. Cell Syst. 2016, 3, 412–413. [Google Scholar] [CrossRef] [PubMed]
- Goeddel, D.V.; Kleid, D.G.; Bolivar, F.; Heyneker, H.L.; Yansura, D.G.; Crea, R.; Hirose, T.; Kraszewski, A.; Itakura, K.; Riggs, A.D. Expression in Escherichia coli of chemically synthesized genes for human insulin. Proc. Natl. Acad. Sci. USA 1979, 76, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Li, H.; Sourjik, V. Assembly and stability of flagellar motor in Escherichia coli. Mol. Microbiol. 2011, 80, 886–899. [Google Scholar] [CrossRef]
- Sivashanmugam, A.; Murray, V.; Cui, C.; Zhang, Y.; Wang, J.; Li, Q. Practical protocols for production of very high yields of recombinant proteins using Escherichia coli. Protein Sci. 2009, 18, 936–948. [Google Scholar] [CrossRef]
- Keasling, J.D. Manufacturing molecules through metabolic engineering. Science 2010, 330, 1355–1358. [Google Scholar] [CrossRef]
- Nielsen, J. Production of biopharmaceutical proteins by yeast: advances through metabolic engineering. Bioengineered 2013, 4, 207–211. [Google Scholar] [CrossRef]
- Paddon, C.J.; Keasling, J.D. Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development. Nat. Rev. Microbiol. 2014, 12, 355. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis—Key to sustainable industrial chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Cummings, R.; Esko, J.; Stanley, P.; Hart, G.; Aebi, M.; Darvill, A.; Kinoshita, T.; Packer, N.; Prestegard, J.; et al. Glycan-Recognizing Probes as Tools–Essentials of Glycobiology. In Essentials of Glycobiology [Internet], 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Liu, L. Antibody glycosylation and its impact on the pharmacokinetics and pharmacodynamics of monoclonal antibodies and Fc-fusion proteins. J. Pharm. Sci. 2015, 104, 1866–1884. [Google Scholar] [CrossRef] [PubMed]
- Aslankoohi, E.; Herrera-Malaver, B.; Rezaei, M.N.; Steensels, J.; Courtin, C.M.; Verstrepen, K.J. Non-conventional yeast strains increase the aroma complexity of bread. PLoS ONE 2016, 11, e0165126. [Google Scholar] [CrossRef] [PubMed]
- Saini, M.; Lin, L.J.; Chiang, C.J.; Chao, Y.P. Synthetic consortium of Escherichia coli for n-butanol production by fermentation of the glucose–xylose mixture. J. Agric. Food Chem. 2017, 65, 10040–10047. [Google Scholar] [CrossRef]
- Lewis, N.E.; Liu, X.; Li, Y.; Nagarajan, H.; Yerganian, G.; O’brien, E.; Bordbar, A.; Roth, A.M.; Rosenbloom, J.; Bian, C.; et al. Genomic landscapes of Chinese hamster ovary cell lines as revealed by the Cricetulus griseus draft genome. Nat. Biotechnol. 2013, 31, 759. [Google Scholar] [CrossRef]
- Hefzi, H.; Ang, K.S.; Hanscho, M.; Bordbar, A.; Ruckerbauer, D.; Lakshmanan, M.; Orellana, C.A.; Baycin-Hizal, D.; Huang, Y.; Ley, D.; et al. A Consensus Genome-scale Reconstruction of Chinese Hamster Ovary Cell Metabolism. Cell Syst. 2016, 3, 434–443.e8. [Google Scholar] [CrossRef]
- Lai, T.; Yang, Y.; Ng, S.K. Advances in mammalian cell line development technologies for recombinant protein production. Pharmaceuticals 2013, 6, 579–603. [Google Scholar] [CrossRef]
- Long, M.R.; Ong, W.K.; Reed, J.L. Computational methods in metabolic engineering for strain design. Curr. Opin. Biotechnol. 2015, 34, 135–141. [Google Scholar] [CrossRef]
- Ögmundarson, Ó.; Herrgård, M.J.; Forster, J.; Hauschild, M.Z.; Fantke, P. Addressing environmental sustainability of biochemicals. Nat. Sustain. 2020, 1–8. [Google Scholar] [CrossRef]
- Burgard, A.P.; Pharkya, P.; Maranas, C.D. Optknock: a bilevel programming framework for identifying gene knockout strategies for microbial strain optimization. Biotechnol. Bioeng. 2003, 84, 647–657. [Google Scholar] [CrossRef]
- Klamt, S.; Mahadevan, R. On the feasibility of growth-coupled product synthesis in microbial strains. Metab. Eng. 2015, 30, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Fong, S.S.; Burgard, A.P.; Herring, C.D.; Knight, E.M.; Blattner, F.R.; Maranas, C.D.; Palsson, B.O. In silico design and adaptive evolution of Escherichia coli for production of lactic acid. Biotechnol. Bioeng. 2005, 91, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Trinh, C.T.; Srienc, F. Metabolic engineering of Escherichia coli for efficient conversion of glycerol to ethanol. Appl. Environ. Microbiol. 2009, 75, 6696–6705. [Google Scholar] [CrossRef] [PubMed]
- Otero, J.M.; Cimini, D.; Patil, K.R.; Poulsen, S.G.; Olsson, L.; Nielsen, J. Industrial systems biology of Saccharomyces cerevisiae enables novel succinic acid cell factory. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Von Kamp, A.; Klamt, S. Growth-coupled overproduction is feasible for almost all metabolites in five major production organisms. Nat. Commun. 2017, 8, 15956. [Google Scholar] [CrossRef]
- Dinh, H.V.; King, Z.A.; Palsson, B.O.; Feist, A.M. Identification of growth-coupled production strains considering protein costs and kinetic variability. Metab. Eng. Commun. 2018, 7, e00080. [Google Scholar] [CrossRef]
- Portnoy, V.A.; Bezdan, D.; Zengler, K. Adaptive laboratory evolution–harnessing the power of biology for metabolic engineering. Curr. Opin. Biotechnol. 2011, 22, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Van Mastrigt, O.; Abee, T.; Lillevang, S.K.; Smid, E.J. Quantitative physiology and aroma formation of a dairy Lactococcus lactis at near-zero growth rates. Food Microbiol. 2018, 73, 216–226. [Google Scholar] [CrossRef]
- Boecker, S.; Zahoor, A.; Schramm, T.; Link, H.; Klamt, S. Broadening the Scope of Enforced ATP Wasting as a Tool for Metabolic Engineering in Escherichia coli. Biotechnol. J. 2019, 1800438. [Google Scholar] [CrossRef]
- Hädicke, O.; Bettenbrock, K.; Klamt, S. Enforced ATP futile cycling increases specific productivity and yield of anaerobic lactate production in Escherichia coli. Biotechnol. Bioeng. 2015, 112, 2195–2199. [Google Scholar] [CrossRef]
- Semkiv, M.V.; Dmytruk, K.V.; Abbas, C.A.; Sibirny, A.A. Activation of futile cycles as an approach to increase ethanol yield during glucose fermentation in Saccharomyces cerevisiae. Bioengineered 2016, 7, 106–111. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, J.; Kandasamy, V.; Würtz, A.; Jensen, P.R.; Solem, C. Stimulation of acetoin production in metabolically engineered Lactococcus lactis by increasing ATP demand. Appl. Microbiol. Biotechnol. 2016, 100, 9509–9517. [Google Scholar] [CrossRef] [PubMed]
- Holm, A.K.; Blank, L.M.; Oldiges, M.; Schmid, A.; Solem, C.; Jensen, P.R.; Vemuri, G.N. Metabolic and transcriptional response to cofactor perturbations in Escherichia coli. J. Biol. Chem. 2010, 285, 17498–17506. [Google Scholar] [CrossRef] [PubMed]
- Feist, A.M.; Palsson, B.Ø. The growing scope of applications of genome-scale metabolic reconstructions using Escherichia coli. Nat. Biotechnol. 2008, 26, 659. [Google Scholar] [CrossRef]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current status and applications of genome-scale metabolic models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef]
- Lewis, N.E.; Nagarajan, H.; Palsson, B.O. Constraining the metabolic genotype–phenotype relationship using a phylogeny of in silico methods. Nat. Rev. Microbiol. 2012, 10, 291–305. [Google Scholar] [CrossRef]
- Orth, J.D.; Thiele, I.; Palsson, B.Ø. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245. [Google Scholar] [CrossRef]
- Zanghellini, J.; Ruckerbauer, D.E.; Hanscho, M.; Jungreuthmayer, C. Elementary flux modes in a nutshell: properties, calculation and applications. Biotechnol. J. 2013, 8, 1009–1016. [Google Scholar] [CrossRef]
- Gagneur, J.; Klamt, S. Computation of elementary modes: a unifying framework and the new binary approach. BMC Bioinform. 2004, 5, 175. [Google Scholar] [CrossRef]
- Maia, P.; Rocha, M.; Rocha, I. In Silico Constraint-Based Strain Optimization Methods: The Quest for Optimal Cell Factories. Microbiol. Mol. Biol. Rev. 2016, 80, 45–67. [Google Scholar] [CrossRef]
- Alter, T.B.; Ebert, B.E. Determination of growth-coupling strategies and their underlying principles. BMC Bioinform. 2019, 20, 447. [Google Scholar] [CrossRef] [PubMed]
- Tepper, N.; Shlomi, T. Predicting metabolic engineering knockout strategies for chemical production: accounting for competing pathways. Bioinformatics 2009, 26, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Pharkya, P.; Burgard, A.P.; Maranas, C.D. OptStrain: a computational framework for redesign of microbial production systems. Genome Res. 2004, 14, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Pharkya, P.; Maranas, C.D. An optimization framework for identifying reaction activation/inhibition or elimination candidates for overproduction in microbial systems. Metab. Eng. 2006, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.R.; Rocha, I.; Förster, J.; Nielsen, J. Evolutionary programming as a platform for in silico metabolic engineering. BMC Bioinform. 2005, 6, 308. [Google Scholar]
- Lun, D.S.; Rockwell, G.; Guido, N.J.; Baym, M.; Kelner, J.A.; Berger, B.; Galagan, J.E.; Church, G.M. Large-scale identification of genetic design strategies using local search. Mol. Syst. Biol. 2009, 5, 296. [Google Scholar] [CrossRef]
- Rocha, I.; Maia, P.; Evangelista, P.; Vilaça, P.; Soares, S.; Pinto, J.P.; Nielsen, J.; Patil, K.R.; Ferreira, E.C.; Rocha, M. OptFlux: An open-source software platform for in silico metabolic engineering. BMC Syst. Biol. 2010, 4, 45. [Google Scholar] [CrossRef]
- Hamdi, A.; Santos, S.T.; Rocha, I. Towards metabolic optimization of CHO cells: in silico improvement of culture medium. In Proceedings of the 8th IFAC Conference on Foundations of Systems Biology in Engineering, Valencia, Spain, 15–18 October 2019. [Google Scholar]
- Kim, J.; Reed, J.L. OptORF: Optimal metabolic and regulatory perturbations for metabolic engineering of microbial strains. BMC Syst. Biol. 2010, 4, 53. [Google Scholar] [CrossRef]
- Kim, J.; Reed, J.L.; Maravelias, C.T. Large-scale bi-level strain design approaches and mixed-integer programming solution techniques. PLoS ONE 2011, 6, e24162. [Google Scholar] [CrossRef]
- Xu, Z.; Zheng, P.; Sun, J.; Ma, Y. ReacKnock: identifying reaction deletion strategies for microbial strain optimization based on genome-scale metabolic network. PLoS ONE 2013, 8, e72150. [Google Scholar] [CrossRef]
- Ren, S.; Zeng, B.; Qian, X. Adaptive bi-level programming for optimal gene knockouts for targeted overproduction under phenotypic constraints. BMC Bioinform. 2013, 14, S17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ji, B.; Mardinoglu, A.; Nielsen, J.; Hua, Q. Logical transformation of genome-scale metabolic models for gene level applications and analysis. Bioinformatics 2015, 31, 2324–2331. [Google Scholar] [CrossRef][Green Version]
- Jian, X.; Zhou, S.; Zhang, C.; Hua, Q. In silico identification of gene amplification targets based on analysis of production and growth coupling. Biosystems 2016, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Zhang, C.; Zhou, S.; Wei, L.; Hua, Q. IdealKnock: a framework for efficiently identifying knockout strategies leading to targeted overproduction. Comput. Biol. Chem. 2016, 61, 229–237. [Google Scholar] [CrossRef]
- Hartmann, A.; Vila-Santa, A.; Kallscheuer, N.; Vogt, M.; Julien-Laferrière, A.; Sagot, M.F.; Marienhagen, J.; Vinga, S. OptPipe-a pipeline for optimizing metabolic engineering targets. BMC Syst. Biol. 2017, 11, 143. [Google Scholar] [CrossRef]
- Jensen, K.; Broeken, V.; Hansen, A.S.L.; Sonnenschein, N.; Herrgård, M.J. OptCouple: Joint simulation of gene knockouts, insertions and medium modifications for prediction of growth-coupled strain designs. Metab. Eng. Commun. 2019, 8, e00087. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Sun, R.; Yao, J.; Li, J.; Liu, Q.; Price, N.D.; Liu, C.; Wang, Z. OptRAM: In-silico strain design via integrative regulatory-metabolic network modeling. PLoS Comput. Biol. 2019, 15, e1006835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z. egKnock: Identifying direct gene knockout strategies for microbial strain optimization based on metabolic network with gene-protein-reaction relationships. bioRxiv 2019, 514653. [Google Scholar]
- Campodonico, M.A.; Andrews, B.A.; Asenjo, J.A.; Palsson, B.O.; Feist, A.M. Generation of an atlas for commodity chemical production in Escherichia coli and a novel pathway prediction algorithm, GEM-Path. Metab. Eng. 2014, 25, 140–158. [Google Scholar] [CrossRef]
- Shepelin, D.; Hansen, A.S.L.; Lennen, R.; Luo, H.; Herrgård, M.J. Selecting the best: evolutionary engineering of chemical production in microbes. Genes 2018, 9, 249. [Google Scholar] [CrossRef]
- Harder, B.J.; Bettenbrock, K.; Klamt, S. Model-based metabolic engineering enables high yield itaconic acid production by Escherichia coli. Metab. Eng. 2016, 38, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Klamt, S. Characterizing and ranking computed metabolic engineering strategies. Bioinformatics 2019, 35, 3063–3072. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Trinh, C.T. Harnessing natural modularity of cellular metabolism to design a modular chassis cell for a diverse class of products by using goal attainment optimization. bioRxiv 2019, 748350. [Google Scholar] [CrossRef]
- Matsumoto, T.; Tanaka, T.; Kondo, A. Engineering metabolic pathways in Escherichia coli for constructing a “microbial chassis” for biochemical production. Bioresour. Technol. 2017, 245, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Jouhten, P.; Boruta, T.; Andrejev, S.; Pereira, F.; Rocha, I.; Patil, K.R. Yeast metabolic chassis designs for diverse biotechnological products. Sci. Rep. 2016, 6, 29694. [Google Scholar] [CrossRef]
- Trinh, C.T.; Mendoza, B. Modular cell design for rapid, efficient strain engineering toward industrialization of biology. Curr. Opin. Chem. Eng. 2016, 14, 18–25. [Google Scholar] [CrossRef]
- Pereira, F.; Lopes, H.; Maia, P.; Meyer, B.; Konstantinidis, D.; Kafkia, E.; Kötter, P.; Rocha, I.; Patil, K. Yeast chassis design for production of dicarboxylic acids. New Biotechnol. 2018, 44, S8. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lin, J.; Jiang, Y.; Yang, S. Improved n-butanol production from Clostridium cellulovorans by integrated metabolic and evolutionary engineering. Appl. Environ. Microbiol. 2019, 85, e02560-18. [Google Scholar] [CrossRef]
- Xavier, J.C.; Patil, K.R.; Rocha, I. Systems biology perspectives on minimal and simpler cells. Microbiol. Mol. Biol. Rev. 2014, 78, 487–509. [Google Scholar] [CrossRef]
- Pósfai, G.; Plunkett, G.; Fehér, T.; Frisch, D.; Keil, G.M.; Umenhoffer, K.; Kolisnychenko, V.; Stahl, B.; Sharma, S.S.; De Arruda, M.; et al. Emergent properties of reduced-genome Escherichia coli. Science 2006, 312, 1044–1046. [Google Scholar] [CrossRef]
- Couto, J.M.; McGarrity, A.; Russell, J.; Sloan, W.T. The effect of metabolic stress on genome stability of a synthetic biology chassis Escherichia coli K12 strain. Microb. Cell Factories 2018, 17, 8. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Kildegaard, K.R.; Li, M.; Borodina, I.; Nielsen, J. Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis. Metab. Eng. 2015, 31, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Chen, Y.; Siewers, V.; Nielsen, J. Improving production of malonyl coenzyme A-derived metabolites by abolishing Snf1-dependent regulation of Acc1. MBio 2014, 5, e01130-14. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Si, T.; Nair, N.U.; Zhao, H. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab. Eng. 2014, 24, 139–149. [Google Scholar] [CrossRef]
- Kozak, B.U.; van Rossum, H.M.; Luttik, M.A.; Akeroyd, M.; Benjamin, K.R.; Wu, L.; de Vries, S.; Daran, J.M.; Pronk, J.T.; van Maris, A.J. Engineering acetyl coenzyme A supply: functional expression of a bacterial pyruvate dehydrogenase complex in the cytosol of Saccharomyces cerevisiae. MBio 2014, 5, e01696-14. [Google Scholar] [CrossRef]
- Soma, Y.; Hanai, T. Self-induced metabolic state switching by a tunable cell density sensor for microbial isopropanol production. Metab. Eng. 2015, 30, 7–15. [Google Scholar] [CrossRef]
- Soma, Y.; Fujiwara, Y.; Nakagawa, T.; Tsuruno, K.; Hanai, T. Reconstruction of a metabolic regulatory network in Escherichia coli for purposeful switching from cell growth mode to production mode in direct GABA fermentation from glucose. Metab. Eng. 2017, 43, 54–63. [Google Scholar] [CrossRef]
- Brockman, I.M.; Prather, K.L. Dynamic metabolic engineering: new strategies for developing responsive cell factories. Biotechnol. J. 2015, 10, 1360–1369. [Google Scholar] [CrossRef]
- Soma, Y.; Tsuruno, K.; Wada, M.; Yokota, A.; Hanai, T. Metabolic flux redirection from a central metabolic pathway toward a synthetic pathway using a metabolic toggle switch. Metab. Eng. 2014, 23, 175–184. [Google Scholar] [CrossRef]
- Tsuruno, K.; Honjo, H.; Hanai, T. Enhancement of 3-hydroxypropionic acid production from glycerol by using a metabolic toggle switch. Microb. Cell Factories 2015, 14, 155. [Google Scholar] [CrossRef]
- Moser, F.; Borujeni, A.E.; Ghodasara, A.N.; Cameron, E.; Park, Y.; Voigt, C.A. Dynamic control of endogenous metabolism with combinatorial logic circuits. Mol. Syst. Biol. 2018, 14, e8605. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Breitling, R.; Bovenberg, R.; Takano, E. Exploiting plug-and-play synthetic biology for drug discovery and production in microorganisms. Nat. Rev. Microbiol. 2011, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A. Systems biology: its practice and challenges. Cell 2005, 121, 511–513. [Google Scholar] [CrossRef] [PubMed]
- McCombie, W.R.; McPherson, J.D.; Mardis, E.R. Next-generation sequencing technologies. Cold Spring Harb. Perspect. Med. 2019, 9, a036798. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230. [Google Scholar] [CrossRef]
- Khalil, A.S.; Collins, J.J. Synthetic biology: Applications come of age. Nat. Rev. Genet. 2010, 11, 367. [Google Scholar] [CrossRef]
- Walsh, G.; Jefferis, R. Post-translational modifications in the context of therapeutic proteins. Nat. Biotechnol. 2006, 24, 1241. [Google Scholar] [CrossRef]
- Meehl, M.A.; Stadheim, T.A. Biopharmaceutical discovery and production in yeast. Curr. Opin. Biotechnol. 2014, 30, 120–127. [Google Scholar] [CrossRef]
- Anurag, A.J.R.A.W.; Rathore, S. Defining critical quality attributes for monoclonal antibody therapeutic products. BioPharm Int. 2014, 27, 34–43. [Google Scholar]
- Batra, J.; Rathore, A.S. Glycosylation of monoclonal antibody products: Current status and future prospects. Biotechnol. Prog. 2016, 32, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Higgins, E. Carbohydrate analysis throughout the development of a protein therapeutic. Glycoconj. J. 2010, 27, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Gifford, J.; Albee, A.; Deeds, Z.; Delong, B.; Kao, K.; Ross, S.; Caple, M. An Efficient Approach to Cell Culture Medium Optimization—A statistical method to medium mixing. In Animal Cell Technology Meets Genomics; Springer: Dordrecht, The Netherlands, 2005; pp. 549–553. [Google Scholar]
- Puente-Massaguer, E.; Badiella, L.; Gutiérrez-Granados, S.; Cervera, L.; Gòdia, F. A statistical approach to improve compound screening in cell culture media. Eng. Life Sci. 2019, 19, 315–327. [Google Scholar] [CrossRef]
- Puck, T.T.; Cieciura, S.J.; Robinson, A. Genetics of somatic mammalian cells: III. Long-term cultivation of euploid cells from human and animal subjects. J. Exp. Med. 1958, 108, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Urlaub, G.; Käs, E.; Carothers, A.M.; Chasin, L.A. Deletion of the diploid dihydrofolate reductase locus from cultured mammalian cells. Cell 1983, 33, 405–412. [Google Scholar] [CrossRef]
- Fan, L.; Kadura, I.; Krebs, L.E.; Hatfield, C.C.; Shaw, M.M.; Frye, C.C. Improving the efficiency of CHO cell line generation using glutamine synthetase gene knockout cells. Biotechnol. Bioeng. 2012, 109, 1007–1015. [Google Scholar] [CrossRef]
- Bort, J.A.H.; Stern, B.; Borth, N. CHO-K1 host cells adapted to growth in glutamine-free medium by FACS-assisted evolution. Biotechnol. J. 2010, 5, 1090–1097. [Google Scholar] [CrossRef]
- Xu, X.; Nagarajan, H.; Lewis, N.E.; Pan, S.; Cai, Z.; Liu, X.; Chen, W.; Xie, M.; Wang, W.; Hammond, S.; et al. The genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nat. Biotechnol. 2011, 29, 735. [Google Scholar] [CrossRef]
- Cao, Y.; Kimura, S.; Itoi, T.; Honda, K.; Ohtake, H.; Omasa, T. Construction of BAC-based physical map and analysis of chromosome rearrangement in Chinese hamster ovary cell lines. Biotechnol. Bioeng. 2012, 109, 1357–1367. [Google Scholar] [CrossRef]
- Derouazi, M.; Martinet, D.; Schmutz, N.B.; Flaction, R.; Wicht, M.; Bertschinger, M.; Hacker, D.; Beckmann, J.; Wurm, F. Genetic characterization of CHO production host DG44 and derivative recombinant cell lines. Biochem. Biophys. Res. Commun. 2006, 340, 1069–1077. [Google Scholar] [CrossRef]
- Lee, J.S.; Kildegaard, H.F.; Lewis, N.E.; Lee, G.M. Mitigating Clonal Variation in Recombinant Mammalian Cell Lines. Trends Biotechnol. 2019, 37, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, H.; Gerstl, M.P.; Ruckerbauer, D.; Hanscho, M.; Himmelbauer, H.; Clarke, C.; Barron, N.; Zanghellini, J.; Borth, N. Genetic and epigenetic variation across genes involved in energy metabolism and mitochondria of Chinese Hamster Ovary cell lines. Biotechnol. J. 2019, 14, 1800681. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, J.; Hernández, I.; Fischer, C.; Hanscho, M.; Auer, N.; Hackl, M.; Jadhav, V.; Baumann, M.; Krempl, P.M.; Schmidl, C.; et al. Comprehensive genome and epigenome characterization of CHO cells in response to evolutionary pressures and over time. Biotechnol. Bioeng. 2016, 113, 2241–2253. [Google Scholar] [CrossRef]
- Kaas, C.S.; Kristensen, C.; Betenbaugh, M.J.; Andersen, M.R. Sequencing the CHO DXB11 genome reveals regional variations in genomic stability and haploidy. BMC Genom. 2015, 16, 160. [Google Scholar] [CrossRef] [PubMed]
- Vcelar, S.; Melcher, M.; Auer, N.; Hrdina, A.; Puklowski, A.; Leisch, F.; Jadhav, V.; Wenger, T.; Baumann, M.; Borth, N. Changes in chromosome counts and patterns in CHO cell lines upon generation of recombinant cell lines and subcloning. Biotechnol. J. 2018, 13, 1700495. [Google Scholar] [CrossRef] [PubMed]
- Vcelar, S.; Jadhav, V.; Melcher, M.; Auer, N.; Hrdina, A.; Sagmeister, R.; Heffner, K.; Puklowski, A.; Betenbaugh, M.; Wenger, T.; et al. Karyotype variation of CHO host cell lines over time in culture characterized by chromosome counting and chromosome painting. Biotechnol. Bioeng. 2018, 115, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Pilbrough, W.; Munro, T.P.; Gray, P. Intraclonal protein expression heterogeneity in recombinant CHO cells. PLoS ONE 2009, 4, e8432. [Google Scholar] [CrossRef]
- Brinkrolf, K.; Rupp, O.; Laux, H.; Kollin, F.; Ernst, W.; Linke, B.; Kofler, R.; Romand, S.; Hesse, F.; Budach, W.E.; et al. Chinese hamster genome sequenced from sorted chromosomes. Nat. Biotechnol. 2013, 31, 694. [Google Scholar] [CrossRef]
- Rupp, O.; MacDonald, M.L.; Li, S.; Dhiman, H.; Polson, S.; Griep, S.; Heffner, K.; Hernandez, I.; Brinkrolf, K.; Jadhav, V.; et al. A reference genome of the Chinese hamster based on a hybrid assembly strategy. Biotechnol. Bioeng. 2018, 115, 2087–2100. [Google Scholar] [CrossRef]
- Zhang, J. Protein-length distributions for the three domains of life. Trends Genet. 2000, 16, 107–109. [Google Scholar] [CrossRef]
- Wang, M.; Kurland, C.G.; Caetano-Anollés, G. Reductive evolution of proteomes and protein structures. Proc. Natl. Acad. Sci. USA 2011, 108, 11954–11958. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G.; Canbäck, B.; Berg, O.G. The origins of modern proteomes. Biochimie 2007, 89, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Struhl, K.; Moqtaderi, Z. The TAFs in the HAT. Cell 1998, 94, 1–4. [Google Scholar] [CrossRef][Green Version]
- Jadhav, V.; Hackl, M.; Druz, A.; Shridhar, S.; Chung, C.Y.; Heffner, K.M.; Kreil, D.P.; Betenbaugh, M.; Shiloach, J.; Barron, N.; et al. CHO microRNA engineering is growing up: Recent successes and future challenges. Biotechnol. Adv. 2013, 31, 1501–1513. [Google Scholar] [CrossRef]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef]
- Cherry, J.M.; Ball, C.; Weng, S.; Juvik, G.; Schmidt, R.; Adler, C.; Dunn, B.; Dwight, S.; Riles, L.; Mortimer, R.K.; et al. Genetic and physical maps of Saccharomyces cerevisiae. Nature 1997, 387, 67. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, N. On the way to commercializing plant cell culture platform for biopharmaceuticals: present status and prospect. Pharm. Bioprocess. 2014, 2, 499. [Google Scholar] [CrossRef]
- Martínez, V.S.; Buchsteiner, M.; Gray, P.; Nielsen, L.K.; Quek, L.E. Dynamic metabolic flux analysis using B-splines to study the effects of temperature shift on CHO cell metabolism. Metab. Eng. Commun. 2015, 2, 46–57. [Google Scholar] [CrossRef]
- Pan, X.; Dalm, C.; Wijffels, R.H.; Martens, D.E. Metabolic characterization of a CHO cell size increase phase in fed-batch cultures. Appl. Microbiol. Biotechnol. 2017, 101, 8101–8113. [Google Scholar] [CrossRef]
- Milo, R.; Phillips, R. Cell Biology by the Numbers; Garland Science: New York, NY, USA; Abingdon, UK, 2015. [Google Scholar]
- Fernandez, J.M.; Hoeffler, J.P. Gene Expression Systems: Using Nature for the Art of Expression; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Bitter, G.A.; Egan, K.M.; Burnette, W.N.; Samal, B.; Fieschko, J.C.; Peterson, D.L.; Downing, M.R.; Wypych, J.; Langley, K.E. Hepatitis B vaccine produced in yeast. J. Med. Virol. 1988, 25, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Salari, R.; Salari, R. Investigation of the best Saccharomyces cerevisiae growth condition. Electron. Phys. 2017, 9, 3592. [Google Scholar] [CrossRef] [PubMed]
- Széliová, D.; E Ruckerbauer, D.; N Galleguillos, S.; B Petersen, L.; Natter, K.; Hanscho, M.; Troyer, C.; Causon, T.; Schoeny, H.; B Christensen, H.; et al. What CHO is made of: Variations in the biomass composition of Chinese hamster ovary cell lines. Metab. Eng. 2020. Accepted manuscript. [Google Scholar]
- Rocha, M.; Ferreira, P.G. Bioinformatics Algorithms: Design and Implementation in Python; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Nguyen, L.N.; Novak, N.; Baumann, M.; Koehn, J.; Borth, N. Bioinformatic Identification of Chinese Hamster Ovary (CHO) Cold-Shock Genes and Biological Evidence of their Cold-Inducible Promoters. Biotechnol. J. 2020, 15, 1900359. [Google Scholar] [CrossRef] [PubMed]
- Monger, C.; Kelly, P.S.; Gallagher, C.; Clynes, M.; Barron, N.; Clarke, C. Towards next generation CHO cell biology: Bioinformatics methods for RNA-Seq-based expression profiling. Biotechnol. J. 2015, 10, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, H.F.; Baycin-Hizal, D.; Lewis, N.E.; Betenbaugh, M.J. The emerging CHO systems biology era: Harnessing the ‘omics revolution for biotechnology. Curr. Opin. Biotechnol. 2013, 24, 1102–1107. [Google Scholar] [CrossRef]
- Borth, N.; Mattanovich, D.; Kunert, R.; Katinger, H. Effect of increased expression of protein disulfide isomerase and heavy chain binding protein on antibody secretion in a recombinant CHO cell line. Biotechnol. Prog. 2005, 21, 106–111. [Google Scholar] [CrossRef]
- Mohan, C.; Park, S.H.; Chung, J.Y.; Lee, G.M. Effect of doxycycline-regulated protein disulfide isomerase expression on the specific productivity of recombinant CHO cells: thrombopoietin and antibody. Biotechnol. Bioeng. 2007, 98, 611–615. [Google Scholar] [CrossRef]
- Tigges, M.; Fussenegger, M. Xbp1-based engineering of secretory capacity enhances the productivity of Chinese hamster ovary cells. Metab. Eng. 2006, 8, 264–272. [Google Scholar] [CrossRef]
- Harreither, E.; Hackl, M.; Pichler, J.; Shridhar, S.; Auer, N.; Łabaj, P.P.; Scheideler, M.; Karbiener, M.; Grillari, J.; Kreil, D.P.; et al. Microarray profiling of preselected CHO host cell subclones identifies gene expression patterns associated with in-creased production capacity. Biotechnol. J. 2015, 10, 1625–1638. [Google Scholar] [CrossRef]
- Vishwanathan, N.; Yongky, A.; Johnson, K.C.; Fu, H.Y.; Jacob, N.M.; Le, H.; Yusufi, F.N.; Lee, D.Y.; Hu, W.S. Global insights into the Chinese hamster and CHO cell transcriptomes. Biotechnol. Bioeng. 2015, 112, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Fu, H.Y.; Raju, R.; Vishwanathan, N.; Hu, W.S. Unveiling gene trait relationship by cross-platform meta-analysis on Chinese hamster ovary cell transcriptome. Biotechnol. Bioeng. 2017, 114, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Diendorfer, A.B.; Hackl, M.; Klanert, G.; Jadhav, V.; Reithofer, M.; Stiefel, F.; Hesse, F.; Grillari, J.; Borth, N. Annotation of additional evolutionary conserved microRNAs in CHO cells from updated genomic data. Biotechnol. Bioeng. 2015, 112, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Hackl, M.; Jadhav, V.; Jakobi, T.; Rupp, O.; Brinkrolf, K.; Goesmann, A.; Pühler, A.; Noll, T.; Borth, N.; Grillari, J. Computational identification of microRNA gene loci and precursor microRNA sequences in CHO cell lines. J. Biotechnol. 2012, 158, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Hackl, M.; Jakobi, T.; Blom, J.; Doppmeier, D.; Brinkrolf, K.; Szczepanowski, R.; Bernhart, S.H.; zu Siederdissen, C.H.; Bort, J.A.H.; Wieser, M.; et al. Next-generation sequencing of the Chinese hamster ovary microRNA transcriptome: Identification, annotation and profiling of microRNAs as targets for cellular engineering. J. Biotechnol. 2011, 153, 62–75. [Google Scholar] [CrossRef]
- Loh, W.P.; Loo, B.; Zhou, L.; Zhang, P.; Lee, D.Y.; Yang, Y.; Lam, K.P. Overexpression of microRNAs enhances recombinant protein production in Chinese hamster ovary cells. Biotechnol. J. 2014, 9, 1140–1151. [Google Scholar] [CrossRef]
- Druz, A.; Son, Y.J.; Betenbaugh, M.; Shiloach, J. Stable inhibition of mmu-miR-466h-5p improves apoptosis resistance and protein production in CHO cells. Metab. Eng. 2013, 16, 87–94. [Google Scholar] [CrossRef]
- Jadhav, V.; Hackl, M.; Klanert, G.; Bort, J.A.H.; Kunert, R.; Grillari, J.; Borth, N. Stable overexpression of miR-17 enhances recombinant protein production of CHO cells. J. Biotechnol. 2014, 175, 38–44. [Google Scholar] [CrossRef]
- Kelly, P.S.; Breen, L.; Gallagher, C.; Kelly, S.; Henry, M.; Lao, N.T.; Meleady, P.; O’Gorman, D.; Clynes, M.; Barron, N. Re-programming CHO cell metabolism using miR-23 tips the balance towards a highly productive phenotype. Biotechnol. J. 2015, 10, 1029–1040. [Google Scholar] [CrossRef]
- Fischer, S.; Marquart, K.F.; Pieper, L.A.; Fieder, J.; Gamer, M.; Gorr, I.; Schulz, P.; Bradl, H. miRNA engineering of CHO cells facilitates production of difficult-to-express proteins and increases success in cell line development. Biotechnol. Bioeng. 2017, 114, 1495–1510. [Google Scholar] [CrossRef]
- Baek, E.; Noh, S.M.; Lee, G.M. Anti-apoptosis engineering for improved protein production from CHO cells. In Heterologous Protein Production in CHO Cells; Springer: New York, NY, USA, 2017; pp. 71–85. [Google Scholar]
- Lee, S.; Lee, G.M. Bcl-2 overexpression in CHO cells improves polyethylenimine-mediated gene transfection. Process. Biochem. 2013, 48, 1436–1440. [Google Scholar] [CrossRef]
- Kroll, P.; Eilers, K.; Fricke, J.; Herwig, C. Impact of cell lysis on the description of cell growth and death in cell culture. Eng. Life Sci. 2017, 17, 440–447. [Google Scholar] [CrossRef]
- Klein, T.; Heinzel, N.; Kroll, P.; Brunner, M.; Herwig, C.; Neutsch, L. Quantification of cell lysis during CHO bioprocesses: Impact on cell count, growth kinetics and productivity. J. Biotechnol. 2015, 207, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, N.; Senga, Y.; Honda, S. Anxa2-and Ctsd-knockout CHO cell lines to diminish the risk of contamination with host cell proteins. Biotechnol. Prog. 2019, 35, e2820. [Google Scholar] [CrossRef] [PubMed]
- Kol, S.; Ley, D.; Wulff, T.; Decker, M.; Arnsdorf, J.; Schoffelen, S.; Hansen, A.H.; Jensen, T.L.; Gutierrez, J.M.; Chiang, A.W.; et al. Multiplex secretome engineering enhances recombinant protein production and purity. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Chiu, J.; Valente, K.N.; Levy, N.E.; Min, L.; Lenhoff, A.M.; Lee, K.H. Knockout of a difficult-to-remove CHO host cell protein, lipoprotein lipase, for improved polysorbate stability in monoclonal antibody formulations. Biotechnol. Bioeng. 2017, 114, 1006–1015. [Google Scholar] [CrossRef]
- Fischer, S.; Handrick, R.; Otte, K. The art of CHO cell engineering: a comprehensive retrospect and future perspectives. Biotechnol. Adv. 2015, 33, 1878–1896. [Google Scholar] [CrossRef]
- Simpson, N.; Singh, R.; Emery, A.; Al-Rubeai, M. Bcl-2 over-expression reduces growth rate and prolongs G1 phase in continuous chemostat cultures of hybridoma cells. Biotechnol. Bioeng. 1999, 64, 174–186. [Google Scholar] [CrossRef]
- Kaufmann, H.; Mazur, X.; Fussenegger, M.; Bailey, J.E. Influence of low temperature on productivity, proteome and protein phosphorylation of CHO cells. Biotechnol. Bioeng. 1999, 63, 573–582. [Google Scholar] [CrossRef]
- Xu, J.; Tang, P.; Yongky, A.; Drew, B.; Borys, M.C.; Liu, S.; Li, Z.J. Systematic Development of Temperature Shift Strategies for Chinese Hamster Ovary Cells Based on Short Duration Cultures and Kinetic Modeling. mAbs 2019, 11, 191–204. [Google Scholar] [CrossRef]
- Bedoya-López, A.; Estrada, K.; Sanchez-Flores, A.; Ramirez, O.T.; Altamirano, C.; Segovia, L.; Miranda-Rios, J.; Trujillo-Roldan, M.A.; Valdez-Cruz, N.A. Effect of temperature downshift on the transcriptomic responses of Chinese hamster ovary cells using recombinant human tissue plasminogen activator production culture. PLoS ONE 2016, 11, e0151529. [Google Scholar] [CrossRef]
- Jiang, Z.; Sharfstein, S.T. Sodium butyrate stimulates monoclonal antibody over-expression in CHO cells by improving gene accessibility. Biotechnol. Bioeng. 2008, 100, 189–194. [Google Scholar] [CrossRef]
- Bi, J.X.; Shuttleworth, J.; Al-Rubeai, M. Uncoupling of cell growth and proliferation results in enhancement of productivity in p21CIP1-arrested CHO cells. Biotechnol. Bioeng. 2004, 85, 741–749. [Google Scholar] [CrossRef]
- Fussenegger, M.; Schlatter, S.; Dätwyler, D.; Mazur, X.; Bailey, J.E. Controlled proliferation by multigene metabolic engineering enhances the productivity of Chinese hamster ovary cells. Nat. Biotechnol. 1998, 16, 468. [Google Scholar] [CrossRef]
- Klamt, S.; Mahadevan, R.; Hädicke, O. When do two-stage processes outperform one-stage processes? Biotechnol. J. 2018, 13, 1700539. [Google Scholar] [CrossRef]
- Kallehauge, T.B.; Li, S.; Pedersen, L.E.; Ha, T.K.; Ley, D.; Andersen, M.R.; Kildegaard, H.F.; Lee, G.M.; Lewis, N.E. Ribosome profiling-guided depletion of an mRNA increases cell growth rate and protein secretion. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- O’Brien, E.J.; Monk, J.M.; Palsson, B.O. Using genome-scale models to predict biological capabilities. Cell 2015, 161, 971–987. [Google Scholar] [CrossRef]
- Galleguillos, S.N.; Ruckerbauer, D.; Gerstl, M.P.; Borth, N.; Hanscho, M.; Zanghellini, J. What can mathematical modelling say about CHO metabolism and protein glycosylation? Comput. Struct. Biotechnol. J. 2017, 15, 212–221. [Google Scholar] [CrossRef]
- Calmels, C.; McCann, A.; Malphettes, L.; Andersen, M.R. Application of a curated genome-scale metabolic model of CHO DG44 to an industrial fed-batch process. Metab. Eng. 2019, 51, 9–19. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Feizi, A.; Li, S.; Kallehauge, T.B.; Hefzi, H.; Grav, L.M.; Ley, D.; Hizal, D.B.; Betenbaugh, M.J.; Voldborg, B.; et al. Genome-scale reconstructions of the mammalian secretory pathway predict metabolic costs and limitations of protein secretion. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, G.M. Down-regulation of lactate dehydrogenase-A by siRNAs for reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin. Appl. Microbiol. Biotechnol. 2007, 74, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Kildegaard, H.F.; Andersen, M.R. Impact of CHO metabolism on cell growth and protein production: an overview of toxic and inhibiting metabolites and nutrients. Biotechnol. J. 2018, 13, 1700499. [Google Scholar] [CrossRef] [PubMed]
- Mulukutla, B.C.; Kale, J.; Kalomeris, T.; Jacobs, M.; Hiller, G.W. Identification and control of novel growth inhibitors in fed-batch cultures of Chinese hamster ovary cells. Biotechnol. Bioeng. 2017, 114, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Mulukutla, B.C.; Mitchell, J.; Geoffroy, P.; Harrington, C.; Krishnan, M.; Kalomeris, T.; Morris, C.; Zhang, L.; Pegman, P.; Hiller, G.W. Metabolic engineering of Chinese hamster ovary cells towards reduced biosynthesis and accumulation of novel growth inhibitors in fed-batch cultures. Metab. Eng. 2019, 54, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Ley, D.; Pereira, S.; Pedersen, L.E.; Arnsdorf, J.; Hefzi, H.; Davy, A.M.; Ha, T.K.; Wulff, T.; Kildegaard, H.F.; Andersen, M.R. Reprogramming AA catabolism in CHO cells with CRISPR/Cas9 genome editing improves cell growth and reduces byproduct secretion. Metab. Eng. 2019, 56, 120–129. [Google Scholar] [CrossRef]
- O’Brien, C.; Allman, A.; Daoutidis, P.; Hu, W.S. Kinetic model optimization and its application to mitigating the Warburg effect through multiple enzyme alterations. Metab. Eng. 2019, 56, 154–164. [Google Scholar] [CrossRef]
- Geoghegan, D.; Arnall, C.; Hatton, D.; Noble-Longster, J.; Sellick, C.; Senussi, T.; James, D.C. Control of amino acid transport into Chinese hamster ovary cells. Biotechnol. Bioeng. 2018, 115, 2908–2929. [Google Scholar] [CrossRef]
- Butler, M.; Spearman, M. The choice of mammalian cell host and possibilities for glycosylation engineering. Curr. Opin. Biotechnol. 2014, 30, 107–112. [Google Scholar] [CrossRef]
- Spearman, M.; Butler, M. Glycosylation in cell culture. In Animal Cell Culture; Springer: Cham, Switzerland, 2015; pp. 237–258. [Google Scholar]
- Ehret, J.; Zimmermann, M.; Eichhorn, T.; Zimmer, A. Impact of cell culture media additives on IgG glycosylation produced in Chinese hamster ovary cells. Biotechnol. Bioeng. 2019, 116, 816–830. [Google Scholar]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Malphettes, L.; Freyvert, Y.; Chang, J.; Liu, P.Q.; Chan, E.; Miller, J.C.; Zhou, Z.; Nguyen, T.; Tsai, C.; Snowden, A.W.; et al. Highly efficient deletion of FUT8 in CHO cell lines using zinc-finger nucleases yields cells that produce completely nonfucosylated antibodies. Biotechnol. Bioeng. 2010, 106, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Mascarenhas, J.; Sealover, N.R.; George, H.J.; Brooks, J.; Kayser, K.J.; Gau, B.; Yasa, I.; Azadi, P.; Archer-Hartmann, S. Chinese hamster ovary (CHO) host cell engineering to increase sialylation of recombinant therapeutic proteins by modulating sialyltransferase expression. Biotechnol. Prog. 2015, 31, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Coats, M.T.; Bydlinski, N.; Maresch, D.; Diendorfer, A.; Klanert, G.; Borth, N. mRNA Transfection into CHO-Cells Reveals Production Bottlenecks. Biotechnol. J. 2019, e1900198. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Ye, Z.; Wang, S.; Schulz, M.A.; Van Coillie, J.; Sun, L.; Chen, Y.H.; Narimatsu, Y.; Hansen, L.; Kristensen, C.; et al. The glycosylation design space for recombinant lysosomal replacement enzymes produced in CHO cells. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stach, C.S.; McCann, M.G.; O’Brien, C.M.; Le, T.S.; Somia, N.; Chen, X.; Lee, K.; Fu, H.Y.; Daoutidis, P.; Zhao, L.; et al. Model-driven engineering of N-linked glycosylation in Chinese Hamster Ovary cells. ACS Synth. Biol. 2019, 8, 2524–2535. [Google Scholar] [CrossRef]
- Kotidis, P.; Jedrzejewski, P.; Sou, S.N.; Sellick, C.; Polizzi, K.; del Val, I.J.; Kontoravdi, C. Model-based optimization of antibody galactosylation in CHO cell culture. Biotechnol. Bioeng. 2019, 116, 1612–1626. [Google Scholar] [CrossRef]
- Dahodwala, H.; Lee, K.H. The fickle CHO: A review of the causes, implications, and potential alleviation of the CHO cell line instability problem. Curr. Opin. Biotechnol. 2019, 60, 128–137. [Google Scholar] [CrossRef]
- Willbanks, A.; Leary, M.; Greenshields, M.; Tyminski, C.; Heerboth, S.; Lapinska, K.; Haskins, K.; Sarkar, S. The evolution of epigenetics: From prokaryotes to humans and its biological consequences. Genet. Epigenet. 2016, 8, GEG–S31863. [Google Scholar] [CrossRef]
- Hernandez, I.; Dhiman, H.; Klanert, G.; Jadhav, V.; Auer, N.; Hanscho, M.; Baumann, M.; Esteve-Codina, A.; Dabad, M.; Gómez, J.; et al. Epigenetic regulation of gene expression in Chinese Hamster Ovary cells in response to the changing environment of a batch culture. Biotechnol. Bioeng. 2019, 116, 677–692. [Google Scholar] [CrossRef]
- Marx, N.; Grünwald-Gruber, C.; Bydlinski, N.; Dhiman, H.; Ngoc Nguyen, L.; Klanert, G.; Borth, N. CRISPR-Based Targeted Epigenetic Editing Enables Gene Expression Modulation of the Silenced Beta-Galactoside Alpha-2, 6-Sialyltransferase 1 in CHO Cells. Biotechnol. J. 2018, 13, 1700217. [Google Scholar] [CrossRef] [PubMed]
- Karottki, K.J.l.C.; Hefzi, H.; Xiong, K.; Shamie, I.; Hansen, A.H.; Li, S.; Pedersen, L.E.; Li, S.; Lee, J.S.; Lee, G.M.; et al. Awakening dormant glycosyltransferases in CHO cells with CRISPRa. Biotechnol. Bioeng. 2019, 117, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Weinguny, M.; Eisenhut, P.; Klanert, G.; Virgolini, N.; Johnson, A.; Ivansson, D.; Lövgren, A.; Borth, N. Random epigenetic modulation of CHO cells by repeated knock-down of DNA-methyltransferases increases population diversity and enables sorting of cells with higher production capacities. Biotechnol. Bioeng. 2020. Submitted manuscript. [Google Scholar]
- Kostyrko, K.; Neuenschwander, S.; Junier, T.; Regamey, A.; Iseli, C.; Schmid-Siegert, E.; Bosshard, S.; Majocchi, S.; Le Fourn, V.; Girod, P.A.; et al. MAR-mediated transgene integration into permissive chromatin and increased expression by recombination pathway engineering. Biotechnol. Bioeng. 2017, 114, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Betts, Z.; Dickson, A.J. Assessment of UCOE on recombinant EPO production and expression stability in amplified Chinese hamster ovary cells. Mol. Biotechnol. 2015, 57, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chusainow, J.; Yap, M.G. DNA methylation contributes to loss in productivity of monoclonal antibody-producing CHO cell lines. J. Biotechnol. 2010, 147, 180–185. [Google Scholar] [CrossRef]
- Moritz, B.; Woltering, L.; Becker, P.B.; Göpfert, U. High levels of histone H 3 acetylation at the CMV promoter are predictive of stable expression in Chinese hamster ovary cells. Biotechnol. Prog. 2016, 32, 776–786. [Google Scholar] [CrossRef]
- Soo, B.P.; Tay, J.; Ng, S.; Ho, S.C.; Yang, Y.; Chao, S.H. Correlation between expression of recombinant proteins and abundance of H3K4Me3 on the enhancer of human cytomegalovirus major immediate-early promoter. Mol. Biotechnol. 2017, 59, 315–322. [Google Scholar] [CrossRef]
- Spencer, S.; Gugliotta, A.; Koenitzer, J.; Hauser, H.; Wirth, D. Stability of single copy transgene expression in CHOK1 cells is affected by histone modifications but not by DNA methylation. J. Biotechnol. 2015, 195, 15–29. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Schimmel, J.; Kool, H.; Kanaar, R.; Tijsterman, M. Inactivation of Pol θ and C-NHEJ eliminates off-target integration of exogenous DNA. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Saito, S.; Maeda, R.; Adachi, N. Dual loss of human POLQ and LIG4 abolishes random integration. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wang, J.; Luo, M.; Luo, H.; Zhao, M.; Han, L.; Zhang, M.; Yang, H.; Xie, Y.; Jiang, H.; et al. Rapid development of stable transgene CHO cell lines by CRISPR/Cas9-mediated site-specific integration into C12orf35. Appl. Microbiol. Biotechnol. 2018, 102, 6105–6117. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; O’Callaghan, P.M.; Droms, K.A.; James, D.C. A mechanistic understanding of production instability in CHO cell lines expressing recombinant monoclonal antibodies. Biotechnol. Bioeng. 2011, 108, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.A.; O’Brien, S.A.; Zhao, L.; Fu, H.Y.; Vishwanathan, N.; Hu, W.S. Recurring genomic structural variation leads to clonal instability and loss of productivity. Biotechnol. Bioeng. 2019, 116, 41–53. [Google Scholar] [CrossRef]
- O’Brien, S.A.; Lee, K.; Fu, H.Y.; Lee, Z.; Le, T.S.; Stach, C.S.; McCann, M.G.; Zhang, A.Q.; Smanski, M.J.; Somia, N.V.; et al. Single Copy Transgene Integration in a Transcriptionally Active Site for Recombinant Protein Synthesis. Biotechnol. J. 2018, 13, 1800226. [Google Scholar] [CrossRef]
- Baumann, M.; Gludovacz, E.; Sealover, N.; Bahr, S.; George, H.; Lin, N.; Kayser, K.; Borth, N. Preselection of recombinant gene integration sites enabling high transcription rates in CHO cells using alternate start codons and recombinase mediated cassette exchange. Biotechnol. Bioeng. 2017, 114, 2616–2627. [Google Scholar] [CrossRef]
- Gaidukov, L.; Wroblewska, L.; Teague, B.; Nelson, T.; Zhang, X.; Liu, Y.; Jagtap, K.; Mamo, S.; Tseng, W.A.; Lowe, A.; et al. A multi-landing pad DNA integration platform for mammalian cell engineering. Nucleic Acids Res. 2018, 46, 4072–4086. [Google Scholar] [CrossRef]
- Pristovšek, N.; Nallapareddy, S.; Grav, L.M.; Hefzi, H.; Lewis, N.E.; Rugbjerg, P.; Hansen, H.G.; Lee, G.M.; Andersen, M.R.; Kildegaard, H.F. Systematic Evaluation of Site-Specific Recombinant Gene Expression for Programmable Mammalian Cell Engineering. ACS Synth. Biol. 2019, 8, 758–774. [Google Scholar] [CrossRef]
- Crawford, Y.; Zhou, M.; Hu, Z.; Joly, J.; Snedecor, B.; Shen, A.; Gao, A. Fast identification of reliable hosts for targeted cell line development from a limited-genome screening using combined φC31 integrase and CRE-Lox technologies. Biotechnol. Prog. 2013, 29, 1307–1315. [Google Scholar] [CrossRef]
- Irion, S.; Luche, H.; Gadue, P.; Fehling, H.J.; Kennedy, M.; Keller, G. Identification and targeting of the ROSA26 locus in human embryonic stem cells. Nat. Biotechnol. 2007, 25, 1477. [Google Scholar] [CrossRef]
- Zambrowicz, B.P.; Imamoto, A.; Fiering, S.; Herzenberg, L.A.; Kerr, W.G.; Soriano, P. Disruption of overlapping transcripts in the ROSA βgeo 26 gene trap strain leads to widespread expression of β-galactosidase in mouse embryos and hematopoietic cells. Proc. Natl. Acad. Sci. USA 1997, 94, 3789–3794. [Google Scholar] [CrossRef] [PubMed]
- Zboray, K.; Sommeregger, W.; Bogner, E.; Gili, A.; Sterovsky, T.; Fauland, K.; Grabner, B.; Stiedl, P.; Moll, H.P.; Bauer, A.; et al. Heterologous protein production using euchromatin-containing expression vectors in mammalian cells. Nucleic Acids Res. 2015, 43, e102. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Zheng, Q.; Jiang, R.; Chen-Tsai, R.Y.; Kong, L.J. A system for site-specific integration of transgenes in mammalian cells. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Brown, A.J.; Gibson, S.J.; Hatton, D.; Arnall, C.L.; James, D.C. Whole synthetic pathway engineering of recombinant protein production. Biotechnol. Bioeng. 2019, 116, 375–387. [Google Scholar] [CrossRef] [PubMed]
- LncBook a Curated Knowledgebase of Human Long Non-coding RNAs. Available online: http://bigd.big.ac.cn/lncbook/index (accessed on 1 May 2020).
- A Comprehensive Compendium of Human Long Non-coding RNAs. Available online: https://lncipedia.org/ (accessed on 1 May 2020).
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.K.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef]
- Davies, S.L.; Lovelady, C.S.; Grainger, R.K.; Racher, A.J.; Young, R.J.; James, D.C. Functional heterogeneity and heritability in CHO cell populations. Biotechnol. Bioeng. 2013, 110, 260–274. [Google Scholar] [CrossRef]
- Fernandez-Martell, A.; Johari, Y.B.; James, D.C. Metabolic phenotyping of CHO cells varying in cellular biomass accumulation and maintenance during fed-batch culture. Biotechnol. Bioeng. 2018, 115, 645–660. [Google Scholar] [CrossRef]
- Pichler, J.; Galosy, S.; Mott, J.; Borth, N. Selection of CHO host cell subclones with increased specific antibody production rates by repeated cycles of transient transfection and cell sorting. Biotechnol. Bioeng. 2011, 108, 386–394. [Google Scholar] [CrossRef]
- Ausländer, S.; Ausländer, D.; Müller, M.; Wieland, M.; Fussenegger, M. Programmable single-cell mammalian biocomputers. Nature 2012, 487, 123. [Google Scholar] [CrossRef]
- Guinn, M.; Bleris, L. Biological 2-input decoder circuit in human cells. ACS Synth. Biol. 2014, 3, 627–633. [Google Scholar] [CrossRef]
- Weinberg, B.H.; Pham, N.H.; Caraballo, L.D.; Lozanoski, T.; Engel, A.; Bhatia, S.; Wong, W.W. Large-scale design of robust genetic circuits with multiple inputs and outputs for mammalian cells. Nat. Biotechnol. 2017, 35, 453. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.N.; Baumann, M.; Dhiman, H.; Marx, N.; Schmieder, V.; Hussein, M.; Eisenhut, P.; Hernandez, I.; Koehn, J.; Borth, N. Novel promoters derived from Chinese Hamster Ovary cells via in silico and in vitro analysis. Biotechnol. J. 2019, 14, 1900125. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Gibson, S.J.; Hatton, D.; James, D.C. In silico design of context-responsive mammalian promoters with user-defined functionality. Nucleic Acids Res. 2017, 45, 10906–10919. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Sweeney, B.; Mainwaring, D.O.; James, D.C. Synthetic promoters for CHO cell engineering. Biotechnol. Bioeng. 2014, 111, 1638–1647. [Google Scholar] [CrossRef]
- Zucchelli, S.; Patrucco, L.; Persichetti, F.; Gustincich, S.; Cotella, D. Engineering translation in mammalian cell factories to increase protein yield: the unexpected use of long non-coding SINEUP RNAs. Comput. Struct. Biotechnol. J. 2016, 14, 404–410. [Google Scholar] [CrossRef]
- Martella, A.; Pollard, S.M.; Dai, J.; Cai, Y. Mammalian synthetic biology: time for big MACs. ACS Synth. Biol. 2016, 5, 1040–1049. [Google Scholar] [CrossRef]
- Pikaart, M.J.; Recillas-Targa, F.; Felsenfeld, G. Loss of transcriptional activity of a transgene is accompanied by DNA methylation and histone deacetylation and is prevented by insulators. Genes Dev. 1998, 12, 2852–2862. [Google Scholar] [CrossRef]
- Dang, Q.; Auten, J.; Plavec, I. Human beta interferon scaffold attachment region inhibits de novo methylation and confers long-term, copy number-dependent expression to a retroviral vector. J. Virol. 2000, 74, 2671–2678. [Google Scholar] [CrossRef][Green Version]
- Hejnar, J.; Hájková, P.; Plachỳ, J.; Elleder, D.; Stepanets, V.; Svoboda, J. CpG island protects Rous sarcoma virus-derived vectors integrated into nonpermissive cells from DNA methylation and transcriptional suppression. Proc. Natl. Acad. Sci. USA 2001, 98, 565–569. [Google Scholar] [CrossRef]
- Choe, D.; Cho, S.; Kim, S.C.; Cho, B.K. Minimal genome: worthwhile or worthless efforts toward being smaller? Biotechnol. J. 2016, 11, 199–211. [Google Scholar] [CrossRef]
- Cartwright, J.F.; Arnall, C.L.; Patel, Y.D.; Barber, N.O.; Lovelady, C.S.; Rosignoli, G.; Harris, C.L.; Dunn, S.; Field, R.P.; Dean, G.; et al. A platform for context-specific genetic engineering of recombinant protein production by CHO cells. J. Biotechnol. 2020, 312, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Mias, G.I.; Li-Pook-Than, J.; Jiang, L.; Lam, H.Y.; Chen, R.; Miriami, E.; Karczewski, K.J.; Hariharan, M.; Dewey, F.E.; et al. Personal omics profiling reveals dynamic molecular and medical phenotypes. Cell 2012, 148, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems biology and multi-omics integration: Viewpoints from the metabolomics research community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.A.; Palsson, B.O. Context-specific metabolic networks are consistent with experiments. PLoS Comput. Biol. 2008, 4, e1000082. [Google Scholar] [CrossRef]
- Colijn, C.; Brandes, A.; Zucker, J.; Lun, D.S.; Weiner, B.; Farhat, M.R.; Cheng, T.Y.; Moody, D.B.; Murray, M.; Galagan, J.E. Interpreting expression data with metabolic flux models: predicting Mycobacterium tuberculosis mycolic acid production. PLoS Comput. Biol. 2009, 5, e1000489. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Price, N.D. Probabilistic integrative modeling of genome-scale metabolic and regulatory networks in Escherichia coli and Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 107, 17845–17850. [Google Scholar] [CrossRef]
- Zur, H.; Ruppin, E.; Shlomi, T. iMAT: An integrative metabolic analysis tool. Bioinformatics 2010, 26, 3140–3142. [Google Scholar] [CrossRef]
- Schmidt, B.J.; Ebrahim, A.; Metz, T.O.; Adkins, J.N.; Palsson, B.Ø.; Hyduke, D.R. GIM3E: Condition-specific models of cellular metabolism developed from metabolomics and expression data. Bioinformatics 2013, 29, 2900–2908. [Google Scholar] [CrossRef]
- Song, H.S.; Reifman, J.; Wallqvist, A. Prediction of metabolic flux distribution from gene expression data based on the flux minimization principle. PLoS ONE 2014, 9, e112524. [Google Scholar] [CrossRef]
- Motamedian, E.; Mohammadi, M.; Shojaosadati, S.A.; Heydari, M. TRFBA: An algorithm to integrate genome-scale metabolic and transcriptional regulatory networks with incorporation of expression data. Bioinformatics 2016, 33, 1057–1063. [Google Scholar] [CrossRef]
- Banos, D.T.; Trébulle, P.; Elati, M. Integrating transcriptional activity in genome-scale models of metabolism. BMC Syst. Biol. 2017, 11, 134. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Reed, J.L. Integrating proteomic or transcriptomic data into metabolic models using linear bound flux balance analysis. Bioinformatics 2018, 34, 3882–3888. [Google Scholar] [CrossRef] [PubMed]

| Tool | Year | Predictions | Application (Strains) | References |
|---|---|---|---|---|
| OptKnock | 2003 | Possible knockouts that can improve production yield. Maximize the target compound production using Bi-Level linear programming | E. coli, G. sulfurreducens, S. cerevisiae | [24,45,46] |
| OptStrain | 2004 | Reaction addition/deletion Possible knockouts. Possible insertions | E. coli, C. acetobutylicum, M. extorquens | [47] |
| OptReg | 2005 | Up and downregulation of reactions for strain design | E. coli | [48] |
| OptGene | 2008 | Optimization of non-linear objective functions Faster predictions compared to OptKnock | S. cerevisiae | [49] |
| RobustKnock | 2009 | Triple level optimization pipeline The framework targets the maximization of minimal target fluxes of industrial targets based on FBA and FVA Production of chemicals of interest is an obligatory by-product of growth rate maximization | E. coli | [45,46] |
| GDLS | 2009 | Employs MILP search approach Finding direct gene-deletion targets Low-complexity search of the space of genetic manipulations ignoring Gene-Protein-Reaction relationships | E. coli | [50] |
| Optflux | 2010 | Bi-level Optimization (OptGene/OptKnock) Multiobjective Optimization | E. coli, yeast, mammalian cells | [51,52] |
| OptORF | 2010 | Depict metabolic engineering strategies based on gene deletion/overexpression | E. coli | [53] |
| SimOptstrain | 2011 | Concurrent gene insertions and knockouts Non-native reaction addition. Bi-level strain design | E. coli | [54] |
| BiMoMa | 2011 | Knockout predictions based on mixed-integer programming solution techniques GPR association | E. coli | [54] |
| ReacKnock | 2013 | Predictions using Bi-Level FBA Improved computation time compared to OptKnock | E. coli | [55] |
| MOMAKnock | 2013 | Relies on MOMA assumption to restrict constraints to steady-state fluxes Identification of robust knockout strategies | E. coli core model | [56] |
| OptGeneKnock | 2015 | Incorporates logic transformation of model (LTM) with a bilevel mixed integer linear programming (MILP)-based knockout method Designing fast genetic intervention strategies | E. coli | [57] |
| APCG | 2016 | Analysis of production and growth coupling Identification of gene targets for improving production of the desired metabolite | E. coli | [58] |
| IdealKnock | 2016 | Predict knockout strategies for overproduction of compounds of interest | Y. lipolytica | [59] |
| SelFI | 2017 | Identification of selection pathways for directed enzyme evolution | ||
| OptPipe | 2017 | Knockout prediction procedures and rank the suggested mutants according to the expected growth rate, production and a new adaptability measure | C. glutamicum | [60] |
| gcOPT | 2018 | Prediction of possible strong growth-coupling combinations and intervention strategies | E. coli | [45] |
| OptCouple | 2019 | Determination of knockouts, insertions and also medium supplements | E. coli | [61] |
| OptRAM | 2019 | Overexpression, knockdown or knockout of both metabolic genes and transcription factors | Saccharomyces cerevisiae | [62] |
| egKnock | 2019 | Find gene deletion targets for maximization of minimum target flux of industrial objective in flux variability analysis This tool takes gene-protein-reaction relationships into consideration | E. coli | [63] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdi, A.; Széliová, D.; Ruckerbauer, D.E.; Rocha, I.; Borth, N.; Zanghellini, J. Key Challenges in Designing CHO Chassis Platforms. Processes 2020, 8, 643. https://doi.org/10.3390/pr8060643
Hamdi A, Széliová D, Ruckerbauer DE, Rocha I, Borth N, Zanghellini J. Key Challenges in Designing CHO Chassis Platforms. Processes. 2020; 8(6):643. https://doi.org/10.3390/pr8060643
Chicago/Turabian StyleHamdi, Anis, Diana Széliová, David E. Ruckerbauer, Isabel Rocha, Nicole Borth, and Jürgen Zanghellini. 2020. "Key Challenges in Designing CHO Chassis Platforms" Processes 8, no. 6: 643. https://doi.org/10.3390/pr8060643
APA StyleHamdi, A., Széliová, D., Ruckerbauer, D. E., Rocha, I., Borth, N., & Zanghellini, J. (2020). Key Challenges in Designing CHO Chassis Platforms. Processes, 8(6), 643. https://doi.org/10.3390/pr8060643