Modification of the Bacterial Cell Wall—Is the Bioavailability Important in Creosote Biodegradation?




Abstract
1. Introduction
- Aromatic hydrocarbons, including polycyclic aromatic hydrocarbons (PAHs), alkylated PAHs (up to 90%) and benzene-toluene-ethylbenzene-xylene (BTEX) pollutants group,
- Phenolic compounds, such as cresols, phenols and xylenols,
- Heterocycles containing nitrogen, sulfur or oxygen (e.g., pyridines, quinolones, benzotiophenes, dibenzofurans and their derivatives),
2. Materials and Methods
2.1. Chemicals
2.2. Bacteria Strains Isolation
2.3. Creosote Biodegradation Test
2.4. Cell Wall Properties
2.5. Glutathione S-Transferase Activity
2.6. Statistical Analysis
3. Results and discussion
3.1. Bacterial strains
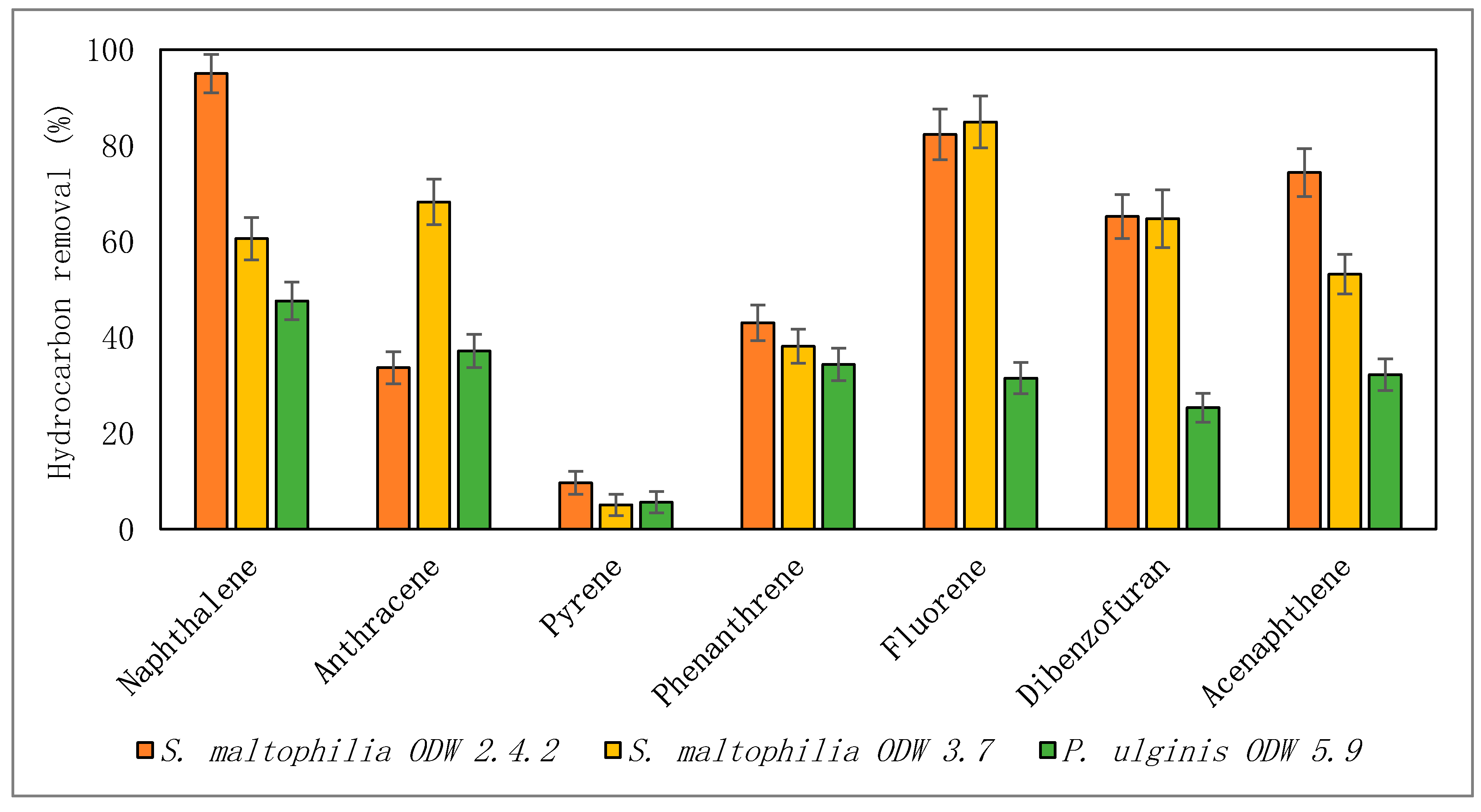
3.2. Creosote Biodegradation
3.3. Enzymatic Activity in the Presence of Creosote
3.4. Cell Wall Properties
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hicknell, B.N.; Mumford, K.G.; Kueper, B.H. Laboratory study of creosote removal from sand at elevated temperatures. J. Contam. Hydrol. 2018, 219, 40–49. [Google Scholar] [CrossRef]
- Madrid, F.; Rubio-Bellido, M.; Villaverde, J.; Peña, A.; Morillo, E. Natural and assisted dissipation of polycyclic aromatic hydrocarbons in a long-term co-contaminated soil with creosote and potentially toxic elements. Sci. Total Environ. 2019, 660, 705–714. [Google Scholar] [CrossRef]
- Webb, D.A. Creosote, its biodegradation and environmental effects. Amer. Wood-Preserv. Assoc. 1980, 76, 65–69. [Google Scholar]
- Melber, C.; Kielhorn, J.; Mangelsdorf, I. Concise International Chamical Assessment Document 62. Coal Car Creosote; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Ye, Y.-F.; Ma, F.-Y.; Wu, M.; Wei, X.-Y.; Liu, J.-M. Increase of acenaphthene content in creosote oil by hydrodynamic cavitation. Chem. Eng. Process. 2016, 104, 66–74. [Google Scholar] [CrossRef]
- Blum, P.; Sagner, A.; Tiehm, A.; Martus, P.; Wendel, T.; Grathwohl, P. Importance of heterocylic aromatic compounds in monitored natural attenuation for coal tar contaminated aquifers: A review. J. Contam. Hydrol. 2011, 126, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.G.; Chapman, P.J.; Pritchard, P.H. Creosote-contaminated sites. Their potential for bioremediation. Environ. Sci. Technol. 1989, 23, 1197–1201. [Google Scholar] [CrossRef]
- Vähäoja, P.; Piltonen, P.; Hyvönen, A.; Niinimäki, J.; Jalonen, J.; Kuokkanen, T. Biodegradability Studies of Certain Wood Preservatives in Groundwater as Determined by the Respirometric Bod Oxitop Method. Water Air Soil Pollut. 2005, 165, 313–324. [Google Scholar] [CrossRef]
- Carlsten, C.; Hunt, S.C.; Kaufman, J.D. Squamous Cell Carcinoma of the Skin and Coal Tar Creosote Exposure in a Railroad Worker. Environ. Health Perspect. 2005, 113, 96–97. [Google Scholar] [CrossRef][Green Version]
- Clapp, R.W.; Jacobs, M.M.; Loechler, E.L. Environmental and Occupational Causes of Cancer: New Evidence 2005-2007. Rev. Environ. Health 2008, 23, 1–38. [Google Scholar] [CrossRef]
- Hu, J.; Mao, Y.; White, K. Renal cell carcinoma and occupational exposure to chemicals in Canada. Occup. Med. 2002, 52, 157–164. [Google Scholar] [CrossRef]
- Krzyśko-Łupicka, T.; Cybulska, K.; Kołosowski, P.; Telesiński, A.; Sudoł, A. Influence of environmental pollution with creosote oil or its vapors on biomass and selected physiological groups of microorganisms. E3S Web Conf. 2017, 22, 00092. [Google Scholar] [CrossRef]
- Kaczorek, E.; Pacholak, A.; Zdarta, A.; Smułek, W. The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase. Colloids Interfaces 2018, 2, 35. [Google Scholar] [CrossRef]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wan, J.; Liu, Y.; Yu, J.; Yi, H.; Ye, S.; Deng, R. Sorption, transport and biodegradation—An insight into bioavailability of persistent organic pollutants in soil. Sci. Total Environ. 2018, 610, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, Y.; Zhong, H.; Xiao, R.; Zeng, G.; Liu, Z.; Cheng, M.; Lai, C.; Zhang, C.; Liu, G.; et al. Mechanisms for rhamnolipids-mediated biodegradation of hydrophobic organic compounds. Sci. Total Environ. 2018, 634, 1–11. [Google Scholar] [CrossRef]
- Xing, S.-F.; Sun, X.-F.; Taylor, A.A.; Walker, S.L.; Wang, Y.-F.; Wang, S.-G. D-Amino acids inhibit initial bacterial Adhesion: Thermodynamic evidence. Biotechnol. Bioeng. 2015, 112, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Hearn, E.M.; Patel, D.R.; van den Berg, B. Outer-membrane transport of aromatic hydrocarbons as a first step in biodegradation. Proc. Natl. Acad. Sci. USA 2008, 105, 8601–8606. [Google Scholar] [CrossRef]
- Hua, F.; Wang, H.Q. Uptake and trans-membrane transport of petroleum hydrocarbons by microorganisms. Biotechnol. Biotechnol. Equip. 2014, 28, 165–175. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59(2), 201–222. [Google Scholar] [CrossRef]
- Kaczorek, E.; Urbanowicz, M.; Olszanowski, A. The influence of surfactants on cell surface properties of Aeromonas hydrophila during diesel oil biodegradation. Colloids Surf. B 2010, 81, 363–368. [Google Scholar] [CrossRef]
- Kaczorek, E.; Sałek, K.; Guzik, U.; Jesionowski, T.; Cybulski, Z. Biodegradation of alkyl derivatives of aromatic hydrocarbons and cell surface properties of a strain of Pseudomonas stutzeri. Chemosphere 2013, 90, 471–478. [Google Scholar] [CrossRef]
- Hassanshahian, M. Isolation and characterization of biosurfactant producing bacteria from Persian Gulf (Bushehr provenance). Mar. Pollut. Bull. 2014, 86, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Ambalam, P.; Kondepudi, K.K.; Nilsson, I.; Wadström, T.; Ljungh, Å. Bile stimulates cell surface hydrophobicity, Congo red binding and biofilm formation of Lactobacillus strains. FEMS Microbiol. Lett. 2012, 333, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Pacholak, A.; Simlat, J.; Zgoła-Grześkowiak, A.; Kaczorek, E. Biodegradation of clotrimazole and modification of cell properties after metabolic stress and upon addition of saponins. Ecotoxicol. Environ. Saf. 2018, 161, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Sakthivel, R.; Nisha, S.A.; Suganthy, N.; Pandian, S.K. Eugenol alters the integrity of cell membrane and acts against the nosocomial pathogen Proteus mirabilis. Arch. Pharmacal Res. 2013, 36, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef]
- Daane, L.L.; Harjono, I.; Barns, S.M.; Launen, L.A.; Palleron, N.J.; Häggblom, M.M. PAH-degradation by Paenibacillus spp. and description of Paenibacillus naphthalenovorans sp. nov., a naphthalene-degrading bacterium from the rhizosphere of salt marsh plants. Int. J. Syst. Evol. Microbiol. 2002, 52, 131–139. [Google Scholar] [CrossRef]
- Mesbaiah, F.Z.; Eddouaouda, K.; Badis, A.; Chebbi, A.; Hentati, D.; Sayadi, S.; Chamkha, M. Preliminary characterization of biosurfactant produced by a PAH-degrading Paenibacillus sp. under thermophilic conditions. Environ. Sci. Pollut. Res. 2016, 23, 14221–14230. [Google Scholar] [CrossRef]
- Tripathi, V.; Gaur, V.K.; Dhiman, N.; Gautam, K.; Manickam, N. Characterization and properties of the biosurfactant produced by PAH-degrading bacteria isolated from contaminated oily sludge environment. Environ. Sci. Pollut. Res. 2019. [Epub ahead of print]. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, K.; Pandey, A.K.; Sharma, A.; Singh, S.B.; Kumar, K.; Arora, A.; Nain, L. Pyrene Degradation by Biosurfactant Producing Bacterium Stenotrophomonas maltophilia. Agric. Res. 2015, 4, 42–47. [Google Scholar] [CrossRef]
- Arulazhagan, P.; Al-Shekri, K.; Huda, Q.; Godon, J.J.; Basahi, J.M.; Jeyakumar, D. Biodegradation of polycyclic aromatic hydrocarbons by an acidophilic Stenotrophomonas maltophilia strain AJH1 isolated from a mineral mining site in Saudi Arabia. Extremophiles 2017, 21, 163–174. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Inoue, C. Polycyclic aromatic hydrocarbons (PAHs) biodegradation potential and diversity of microbial consortia enriched from tsunami sediments in Miyagi, Japan. J. Hazard. Mater. 2015, 283, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, C.; Alessandri, R.; Aunola, T.; Cortina, J.L.; Gamisans, X.; Tuhkanen, T. Oxidation by Fenton as reagent combined with biological treatment applied to a creosote-contaminated soil. J. Hazard. Mater. 2009, 166, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Jurys, A.; Gailiũte, I.; Aikaitė-Stanaitienė, J.; Grigiškis, S.; Maruška, A.; Stankevičius, M.; Levišauskas, D. Review of creosote pollution toxicity and possibilities of bioremediation. Environ. Technol. Resour. 2013, 1, 34–38. [Google Scholar] [CrossRef][Green Version]
- Grant, R.J.; Muckian, L.M.; Clipson, N.J.W.; Doyle, E.M. Microbial community changes during the bioremediation of creosote-contaminated soil. Lett. Appl. Microbiol. 2007, 44, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Widada, J.; Nojiri, H.; Kasuga, K.; Yoshida, T.; Habe, H.; Omori, T. Molecular detection and diversity of polycyclic aromatic hydrocarbon-degrading bacteria isolated from geographically diverse sites. Appl. Microbiol. Biotechnol. 2002, 58, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Shor, L.M.; Liang, W.; Rockne, K.J.; Young, L.Y.; Taghon, G.L.; Kosson, D.S. Intraaggregate mass transport-limited bioavailability of polycyclic aromatic hydrocarbons to Mycobacterium strain PC01. Environ. Sci. Technol. 2003, 37, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Kanaly, R.A.; Harayama, S. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J. Bacteriol. 2000, 182(8), 2059–2067. [Google Scholar] [CrossRef]
- Muangchinda, C.; Yamazoe, A.; Polrit., D.; Thoetkiattikul, H.; Mhuantong, W.; Champreda, V.; Pinyakong, O. Biodegradation of high concentrations of mixed polycyclic aromatic hydrocarbons by indigenous bacteria from a river sediment: a microcosm study and bacterial community analysis. Environ. Sci. Pollut. Res. 2017, 24, 4591–4602. [Google Scholar] [CrossRef]
- Subashchandrabosea, S.R.; Venkateswarlub, K.; Naidua, R.; Meghara, M. Biodegradation of high-molecular weight PAHs by Rhodococcus wratislaviensis strain 9: Overexpression of amidohydrolase induced by pyrene and BaP. Sci. Total Environ. 2019, 651, 813–821. [Google Scholar] [CrossRef]
- Monostori, P.; Wittmann, G.; Karg, E.; Túri, S. Determination of glutathione and glutathione disulfide in biological samples: An in-depth review. J. Chrom. B. 2009, 877, 3331–3346. [Google Scholar] [CrossRef]
- Pour, L.M.; Farahnak, A.; Rad, M.M.; Golmohamadi, T.; Eshraghian, M. Activity Assay of Glutathione S-Transferase (GSTs) Enzyme as a Diagnostic Biomarker for Liver Hydatid Cyst in Vitro. Iran J. Public Health. 2014, 43, 994. [Google Scholar]
- Allocati, N.; Federici, L.; Masulli, M.; di Ilio, C. Glutathione transferases in bacteria: Bacterial. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Zablotowicz, R.M.; Hoagland, R.E.; Locke, M.A.; Hickey, W.J. Glutathione-S-Transferase Activity and Metabolism of Glutathione Conjugates by Rhizosphere Bacteria. Appl. Environ. Microbiol. 1995, 61, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, I.; Mukherji, S. Diverse effect of surfactants on pyrene biodegradation by a Pseudomonas strain utilizing pyrene by cell surface hydrophobicity induction. Int. Biodeter. Biodegrad. 2016, 108, 67–75. [Google Scholar] [CrossRef]
- Obuekwe, C.O.; Al-Jadi, Z.K.; Al-Saleh, E.S. Hydrocarbon degradation in relation to cell-surface hydrophobicity among bacterial hydrocarbon degraders from petroleum-contaminated Kuwait desert environment. Int. Biodeter. Biodegrad. 2009, 63, 273–279. [Google Scholar] [CrossRef]
- Bezza, F.A.; Chirwa, E.M.N. Pyrene biodegradation enhancement potential of lipopeptide biosurfactant produced by Paenibacillus dendritiformis CN5 strain. J. Hazard. Mater. 2017, 321, 218–227. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, J.; Li, J.; Cao, B.; Chen, Y.; Liu, D.; Wang, X.; Zhang, Y. Exogenous Zn2+ enhance the biodegradation of atrazine by regulating the chlorohydrolase gene trzN transcription and membrane permeability of the degrader Arthrobacter sp. DNS10. Chemosphere 2020, 238, 124594. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B.; Korshunova, I.O.; Rubtsova, E.V. Assessment of bacterial resistance to organic solvents using a combined confocal laser scanning and atomic force microscopy (CLSM/AFM). J. Microbiol. Methods 2014, 107, 23–29. [Google Scholar] [CrossRef]
- Dam, S.; Pagès, J.-M.; Masi, M. Stress responses, outer membrane permeability control and antimicrobial resistance in Enterobacteriaceae. Microbiology 2018, 164, 260–267. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Sample | Strain | GenBank (NCBI) Number | Hemolysis Test |
|---|---|---|---|
| O10 | Stenotrophomonas maltophilia ODW 2.4.2 | MK503432.1 | alpha |
| O6 | Stenotrophomonas maltophilia ODW 3.7 | MK503436.1 | alpha |
| O8 | Paenibacillus ulginis ODW 5.9 | MK503429.1 | beta |
| Strain | Carbon Source | CR (%) | ONPG (μM min−1) | CV (%) |
|---|---|---|---|---|
| Stenotrophomonas maltophilia ODW 2.4.2 | succinate | 7.0 ± 0.8 | 0.28 ± 0.01 | 37 ± 3 |
| creosote | 1.8 ± 0.6 | 0.15 ± 0.01 | 24 ± 2 | |
| Stenotrophomonas maltophilia ODW 3.7 | succinate | 4.2 ± 0.1 | 0.51 ± 0.06 | 40 ± 3 |
| creosote | 5.1 ± 0.5 | 0.44 ± 0.09 | 59 ± 4 | |
| Paenibacillus ulginis ODW 5.9 | succinate | 3.4 ± 0.8 | 1.24 ± 0.21 | 16 ± 2 |
| creosote | 13.6 ± 0.9 | 0.13 ± 0.06 | 36 ± 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smułek, W.; Pacholak, A.; Kaczorek, E. Modification of the Bacterial Cell Wall—Is the Bioavailability Important in Creosote Biodegradation? Processes 2020, 8, 147. https://doi.org/10.3390/pr8020147
Smułek W, Pacholak A, Kaczorek E. Modification of the Bacterial Cell Wall—Is the Bioavailability Important in Creosote Biodegradation? Processes. 2020; 8(2):147. https://doi.org/10.3390/pr8020147
Chicago/Turabian StyleSmułek, Wojciech, Amanda Pacholak, and Ewa Kaczorek. 2020. "Modification of the Bacterial Cell Wall—Is the Bioavailability Important in Creosote Biodegradation?" Processes 8, no. 2: 147. https://doi.org/10.3390/pr8020147
APA StyleSmułek, W., Pacholak, A., & Kaczorek, E. (2020). Modification of the Bacterial Cell Wall—Is the Bioavailability Important in Creosote Biodegradation? Processes, 8(2), 147. https://doi.org/10.3390/pr8020147