Effect of Sialic Acid on Mammalian Cell Culture and Protein Expression: A Potential Productivity Enhancer for Biopharmaceutical Cell Culture Processes



Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line, Media, and Seed Expansion
2.2. Fed-Batch Production Culture Conditions
2.3. Cell and Metabolite Analysis
2.4. Analysis of Antibody Concentration (Titer)
2.5. Physicochemical Analysis
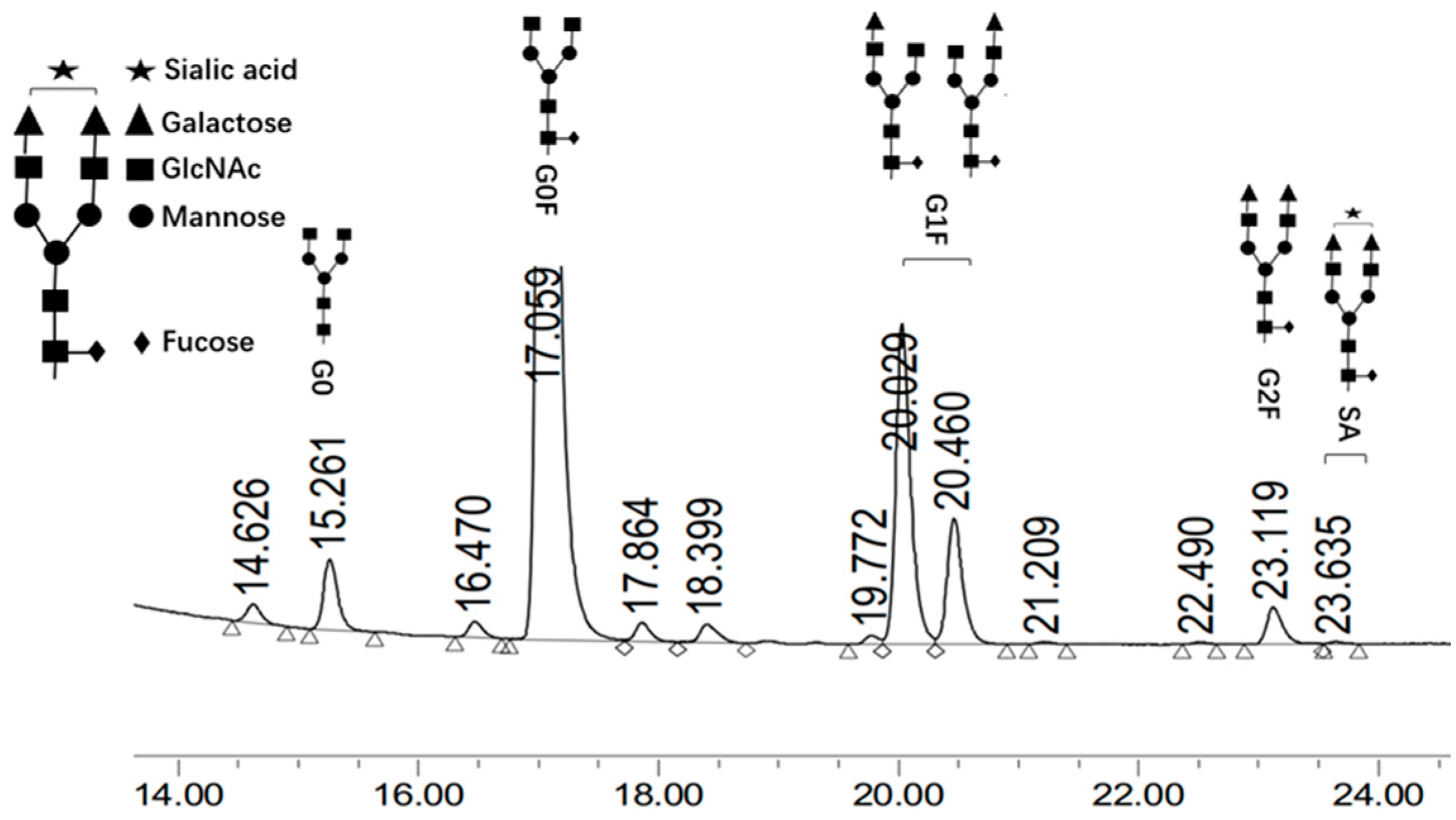
2.6. Analysis of N-Glycan SA
2.7. Analysis of ROS
2.8. Statistical Analysis
3. Results and Discussion
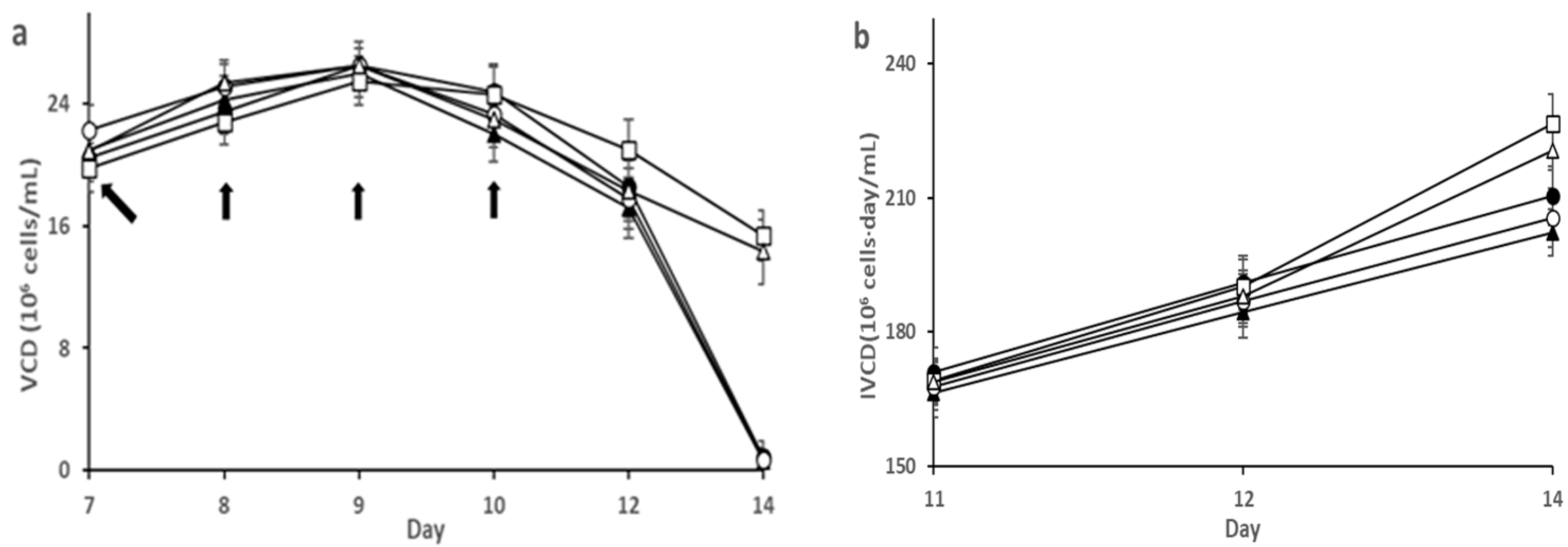
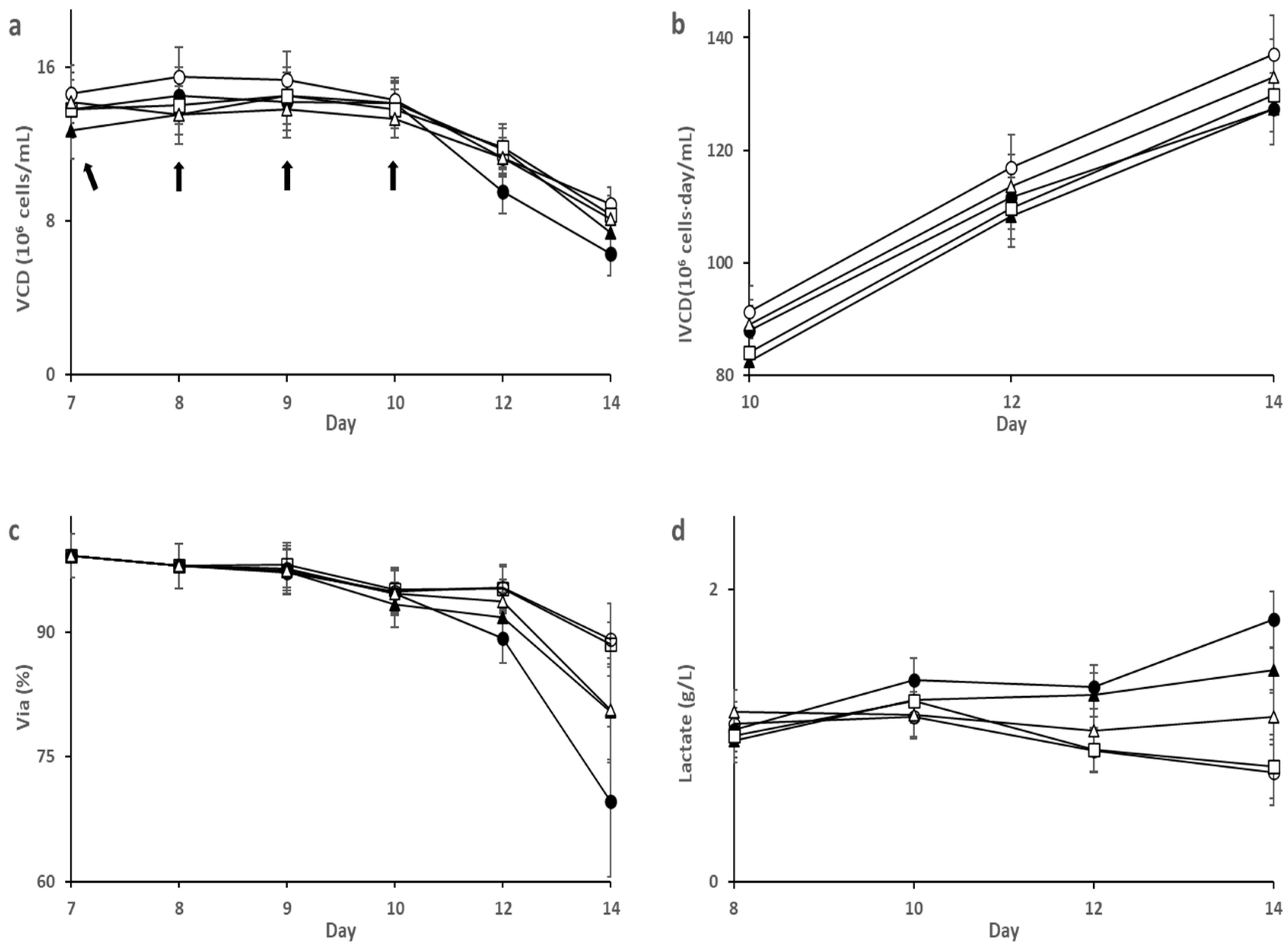
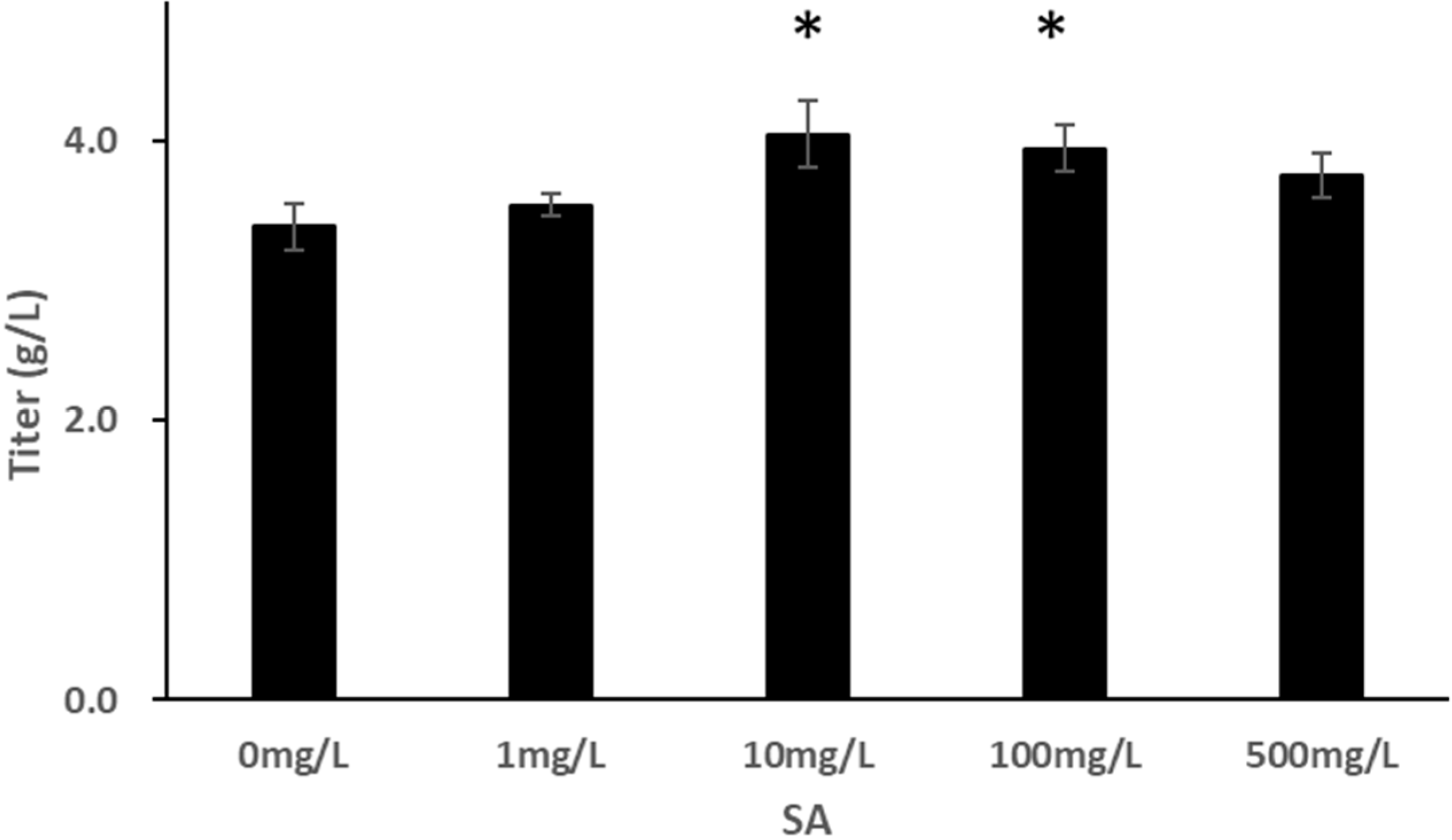
3.1. Effect of SA on HEK293 Cell Culture
3.2. Effect of SA on CHO-K1 Cell Culture
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Couzin-Frankel, J. Breakthrough of the year 2013. Cancer immunotherapy. Science 2013, 342, 1432–1433. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.; Meneses-Acosta, A. Recent advances in technology supporting biopharmaceutical production from mammalian cells. Appl. Microbiol. Biotechnol. 2012, 96, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Dumont, J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, Y.G.; Lee, G.M. CHO cells in biotechnology for production of recombinant proteins: Current state and further potential. Appl. Microbiol. Biotechnol. 2012, 93, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Cell culture, oxidative stress, and antioxidants: Avoiding pitfalls. Biomed. J. 2014, 37, 99–105. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Fritz, R.; Bol, J.; Hebling, U.; Angermuller, S.; Volkl, A.; Fahimi, H.D.; Mueller, S. Compartment-dependent management of H2O2 by peroxisomes. Free Radic. Biol. 2007, 42, 1119–1129. [Google Scholar] [CrossRef]
- Halliwell, B.; Cross, C.E. Oxygen-derived species: Their relation to human disease and environmental stress. Environ. Health Perspect. 1994, 102, 5–12. [Google Scholar]
- Schnellbaecher, A.; Binder, D.; Bellmaine, S.; Zimmer, A. Vitamins in cell culture media: Stability and stabilization strategies. Biotechnol. Bioeng. 2019, 116, 1537–1555. [Google Scholar] [CrossRef]
- Kurano, S.; Kurano, N.; Leist, C.; Fiechter, A. Utilization and stability of vitamins in serum-containing and serum-free media in CHO cell culture. Cytotechnology 1990, 4, 243–250. [Google Scholar] [CrossRef]
- Hata, R.; Senoo, H. L-ascorbic acid 2-phosphate stimulates collagen accumulation, cell proliferation, and formation of a three-dimensional tissue like substance by skin fibroblasts. J. Cell Physiol. 1989, 138, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Yun, Z.; Takagi, M.; Yoshida, T. Effect of antioxidants on the apoptosis of CHO cells and production of tissue plasminogen activator in suspension culture. J. Biosci. Bioeng. 2001, 91, 581–585. [Google Scholar] [CrossRef]
- Angata, T.; Varki, A. Chemical diversity in the sialic acids and related alpha-keto acids: An evolutionary perspective. Chem. Rev. 2002, 102, 439–469. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.C.; Kang, K.A.; Lim, C.M.; Park, J.H.; Jung, K.S.; Hyun, J.W. Water extract of edible bird’s nest attenuated the oxidative stress-induced matrix metalloproteinase-1 by regulating the mitogen-activated protein kinase and activator protein-1 pathway in human keratinocytes. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 347–354. [Google Scholar] [CrossRef]
- Wang, B.; Brand-Miller, J. The role and potential of sialic acid in human nutrition. Eur. J. Clin. Nutr. 2003, 57, 1351–1369. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Yang, B.; Zhu, Q.J. Theoretical studies on the electronic structure parameters and reactive activity of Neu5Gc and Neu5Ac under food processing solvent environment. Molecules 2019, 24, 313. [Google Scholar] [CrossRef]
- Iijima, R.; Takahashi, H.; Namme, R.; Ikegami, S.; Yamazaki, M. Novel biological function of sialic acid (N-acetylneuraminic acid) as a hydrogen peroxide scavenger. FEBS Lett. 2004, 561, 163–166. [Google Scholar] [CrossRef]
- Iijima, R.; Ichikawa, T.; Yamazaki, M. Sialic acid attenuates the cytotoxicity of the lipid hydroperoxides HpODE and HpETE. Carbohydr. Res. 2009, 334, 933–935. [Google Scholar] [CrossRef]
- Kang, D.G.; Kim, C.S.; Cha, H.J. Coexpression of CMP-sialic acid transporter reduces N-glycolylneuraminic acid levels of recombinant glycoproteins in Chinese hamster ovary cells. Biotechnol. Bioeng. 2019, 116, 2815–2822. [Google Scholar] [CrossRef]
- Liste-Calleja, L.; Lecina, M.; Lopez-Repullo, J.; Albiol, J.; Solà, C.; Cairó, J.J. Lactate and glucose concomitant consumption as a self-regulated pH detoxification mechanism in HEK293 cell cultures. Appl. Microbiol. Biotechnol. 2015, 99, 9951–9960. [Google Scholar] [CrossRef]
- Karengera, E.; Robotham, A.; Kelly, J. Concomitant reduction of lactate and ammonia accumulation in fed-batch cultures: Impact on glycoprotein production and quality. Biotechnol. Prog. 2018, 34, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Dhara, V.G.; Naik, H.M.; Majewska, N.I.; Betenbaugh, M.J. Recombinant antibody production in CHO and NS0 Cells: Differences and similarities. BioDrugs 2018, 32, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Val, I.J.; Muller, C. Amino acid and glucose metabolism in fed-batch CHO cell culture affects antibody production and glycosylation. Biotechnol. Bioeng. 2015, 112, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Yuk, I.H.; Russell, S.; Tang, Y. Effects of copper on CHO cells: Cellular requirements and product quality considerations. Biotechnol. Prog. 2015, 31, 226–238. [Google Scholar] [CrossRef]
- Chevallier, V.; Andersen, M.R.; Malphettes, L. Oxidative stress-alleviating strategies to improve recombinant protein production in CHO cells. Biotechnol. Bioeng. 2020, 117, 1172–1186. [Google Scholar] [CrossRef]
- Zagari, F.; Jordan, M.; Stettler, M. Lactate metabolism shift in CHO cell culture: The role of mitochondrial oxidative activity. New Biotechnol. 2013, 30, 238–245. [Google Scholar] [CrossRef]
- Kyriakopoulos, S.; Kontoravdi, C. A framework for the systematic design of fed-batch strategies in mammalian cell culture. Biotechnol. Bioeng. 2014, 111, 2466–2476. [Google Scholar] [CrossRef]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef]
- Pan, X.; Streefland, M.; Dalm, C. Selection of chemically defined media for CHO cell fed-batch culture processes. Cytotechnology 2017, 69, 39–56. [Google Scholar] [CrossRef]
- Kildegaard, H.F.; Fan, Y.; Sen, J.W. Glycoprofiling effects of media additives on IgG produced by CHO cells in fed-batch bioreactors. Biotechnol. Bioeng. 2016, 113, 359–366. [Google Scholar] [CrossRef]
- Xu, S.; Hoshan, L.; Chen, H. Improving lactate metabolism in an intensified CHO culture process: Productivity and product quality considerations. Bioprocess Biosyst. Eng. 2016, 39, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, C.; Illanes, A.; Becerra, S. Considerations on the lactate consumption by CHO cells in the presence of galactose. J. Biotechnol. 2006, 125, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Mascarenhas, J.; Sealover, N. Chinese hamster ovary (CHO) host cell engineering to increase sialylation of recombinant therapeutic proteins by modulating sialyltransferase expression. Biotechnol. Prog. 2015, 31, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Feary, M.; Racher, A.J.; Young, R.J.; Smales, C.M. Methionine Sulfoximine supplementation enhances productivity in GS-CHOK1SV cell lines through Glutathione biosynthesis. Biotechnol. Prog. 2017, 33, 17–25. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Basal Medium | Feed Medium | Time of Adding Sialic Acid (Day) |
|---|---|---|---|
| HEK293 | Dynamis | Cell Boost 7a/7b | 9, 10, 11, 12 |
| CHO cell line A | Dynamis | Cell Boost 7a/7b | 7, 8, 9, 10 |
| CHO cell line B | Dynamis | Cell Boost 7a/7b | 7, 8, 9, 10 |
| Cell Line | Condition | SEC Major Peak (%) | Charge Heterogeneity | N-Glycan Profile | |||||
|---|---|---|---|---|---|---|---|---|---|
| Acidic (%) | Main (%) | Basic (%) | G0F (%) | G1F (%) | G2F (%) | SA (%) | |||
| A | Control | 98.8 ± 0.3 | 23.4 ± 1.3 | 68.2 ± 2.4 | 8.4 ± 0.9 | 85.2 ± 3.1 | 10.2 ± 1.2 | 2.1 ± 0.2 | 0.5 ± 0.1 |
| SA:10 mg/L | 99.1± 0.2 | 20.9 ± 1.2 | 70.3 ± 2.5 | 8.8 ± 0.9 | 83.7 ± 3.0 | 11.3 ± 1.1 | 1.9 ± 0.2 | 0.5 ± 0.1 | |
| SA:100 mg/L | 98.5 ± 0.3 | 17.0 ± 1.0 | 73.9 ± 2.4 | 9.1 ± 1.1 | 86.3 ± 2.8 | 8.76 ± 1.1 | 2.4 ± 0.3 | 0.6 ± 0.1 | |
| B | Control | 99.2 ± 0.2 | 20.8 ± 1.1 | 72.4 ± 2.4 | 6.8 ± 0.7 | 78.2 ± 2.9 | 14.4 ± 1.3 | 3.2 ± 0.3 | 0.7 ± 0.1 |
| SA:10 mg/L | 98.8 ± 0.3 | 17.1 ± 1.1 | 75.7 ± 2.6 | 7.2 ± 0.7 | 79.3 ± 2.4 | 13.8 ± 1.3 | 3.0 ± 0.2 | 0.8 ± 0.1 | |
| SA:100 mg/L | 99.1 ± 0.3 | 16.8 ± 1.3 | 76.1 ± 2.6 | 7.1 ± 0.8 | 81.2 ± 2.5 | 12.2 ± 1.2 | 2.8 ± 0.3 | 0.8 ± 0.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Xiao, S.; Wu, J.; Yao, J. Effect of Sialic Acid on Mammalian Cell Culture and Protein Expression: A Potential Productivity Enhancer for Biopharmaceutical Cell Culture Processes. Processes 2020, 8, 1449. https://doi.org/10.3390/pr8111449
Chen X, Xiao S, Wu J, Yao J. Effect of Sialic Acid on Mammalian Cell Culture and Protein Expression: A Potential Productivity Enhancer for Biopharmaceutical Cell Culture Processes. Processes. 2020; 8(11):1449. https://doi.org/10.3390/pr8111449
Chicago/Turabian StyleChen, Xiangsong, Shang Xiao, Jinyong Wu, and Jianming Yao. 2020. "Effect of Sialic Acid on Mammalian Cell Culture and Protein Expression: A Potential Productivity Enhancer for Biopharmaceutical Cell Culture Processes" Processes 8, no. 11: 1449. https://doi.org/10.3390/pr8111449
APA StyleChen, X., Xiao, S., Wu, J., & Yao, J. (2020). Effect of Sialic Acid on Mammalian Cell Culture and Protein Expression: A Potential Productivity Enhancer for Biopharmaceutical Cell Culture Processes. Processes, 8(11), 1449. https://doi.org/10.3390/pr8111449