Effects of Bromelain and Trypsin Hydrolysis on the Phytochemical Content, Antioxidant Activity, and Antibacterial Activity of Roasted Butterfly Pea Seeds



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
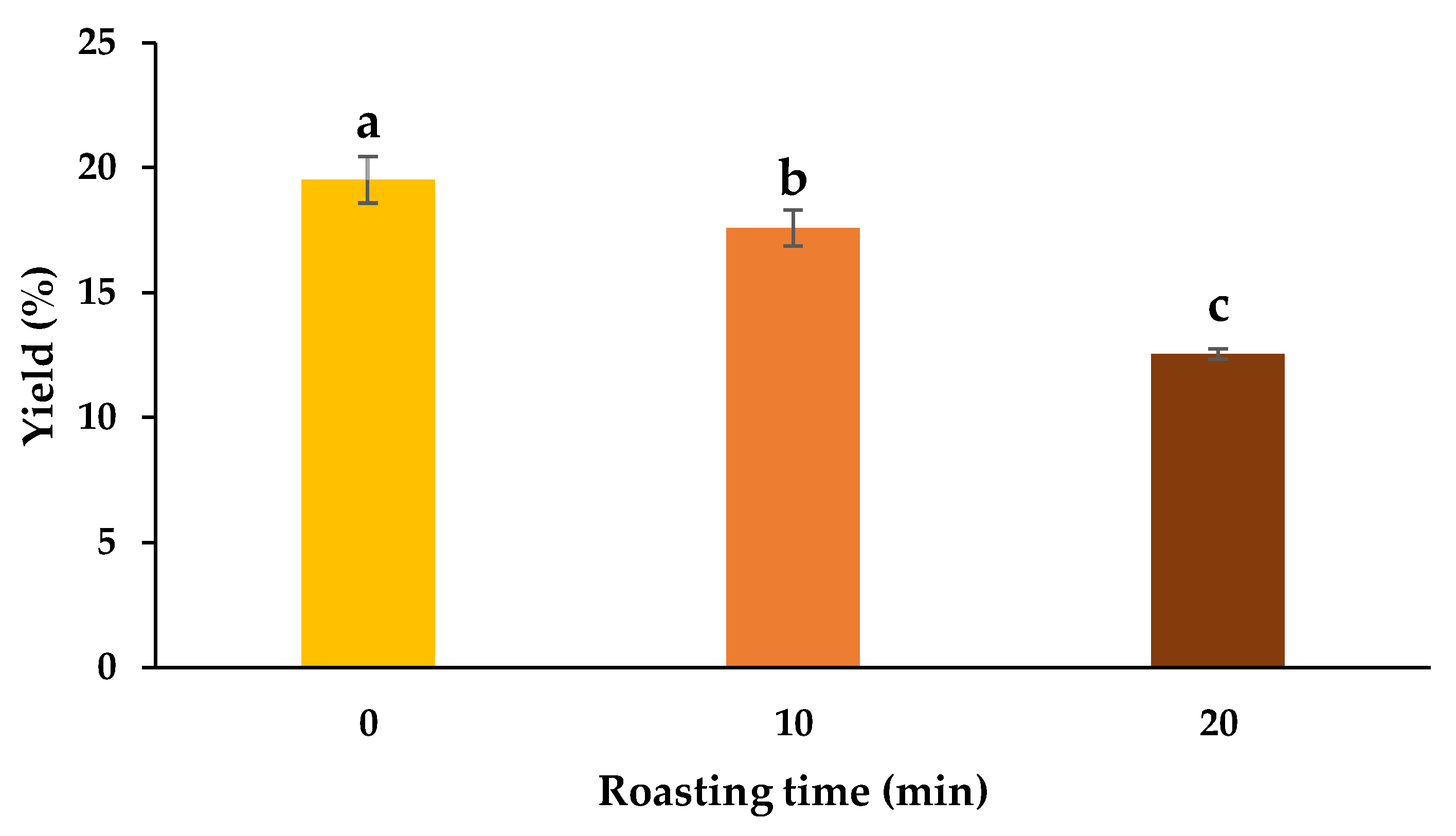
2.3. Extraction and Yield Determination
2.4. Enzymatic Hydrolysis
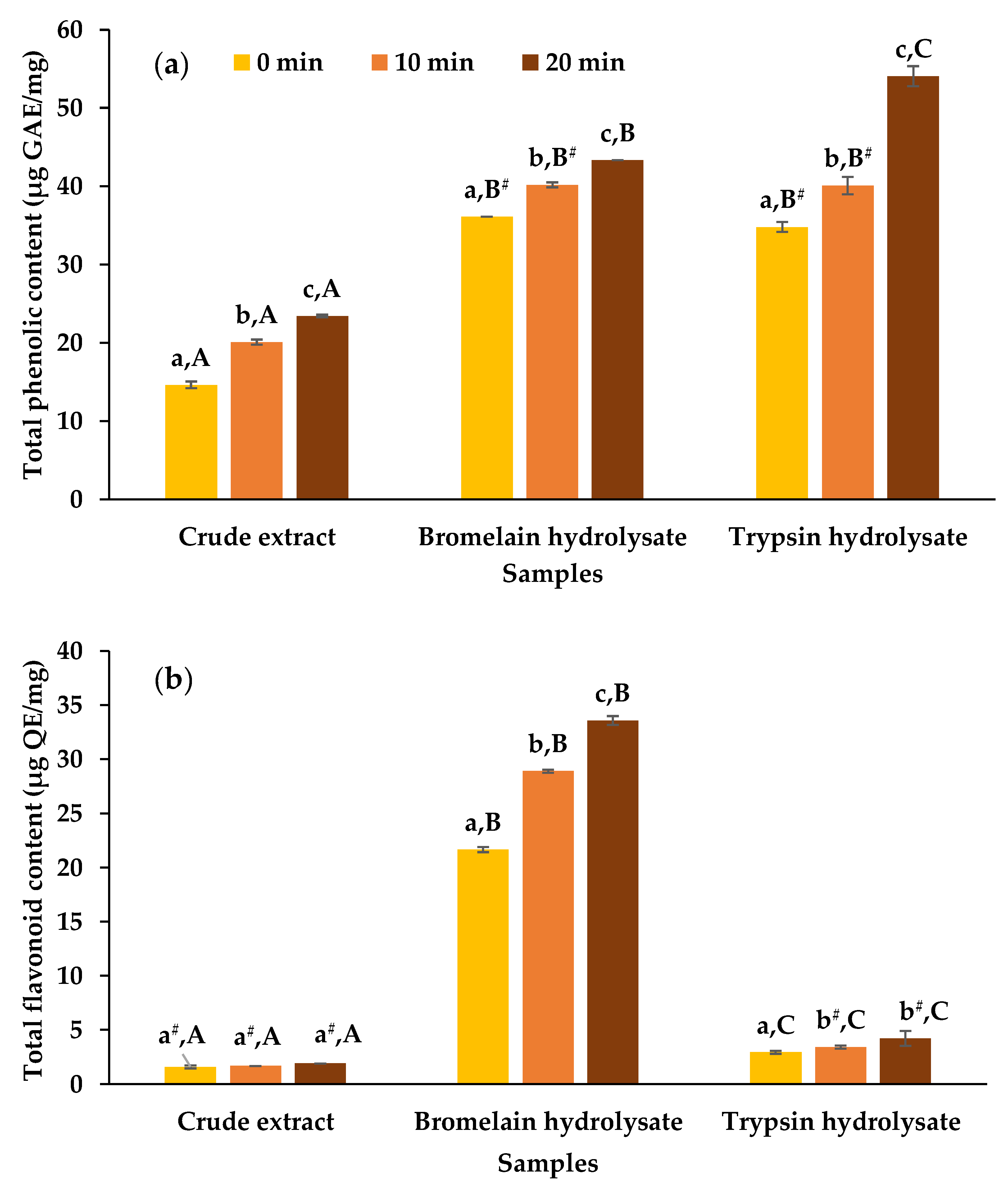
2.5. Determination of Total Phenolic Content
2.6. Determination of Total Flavonoid Content
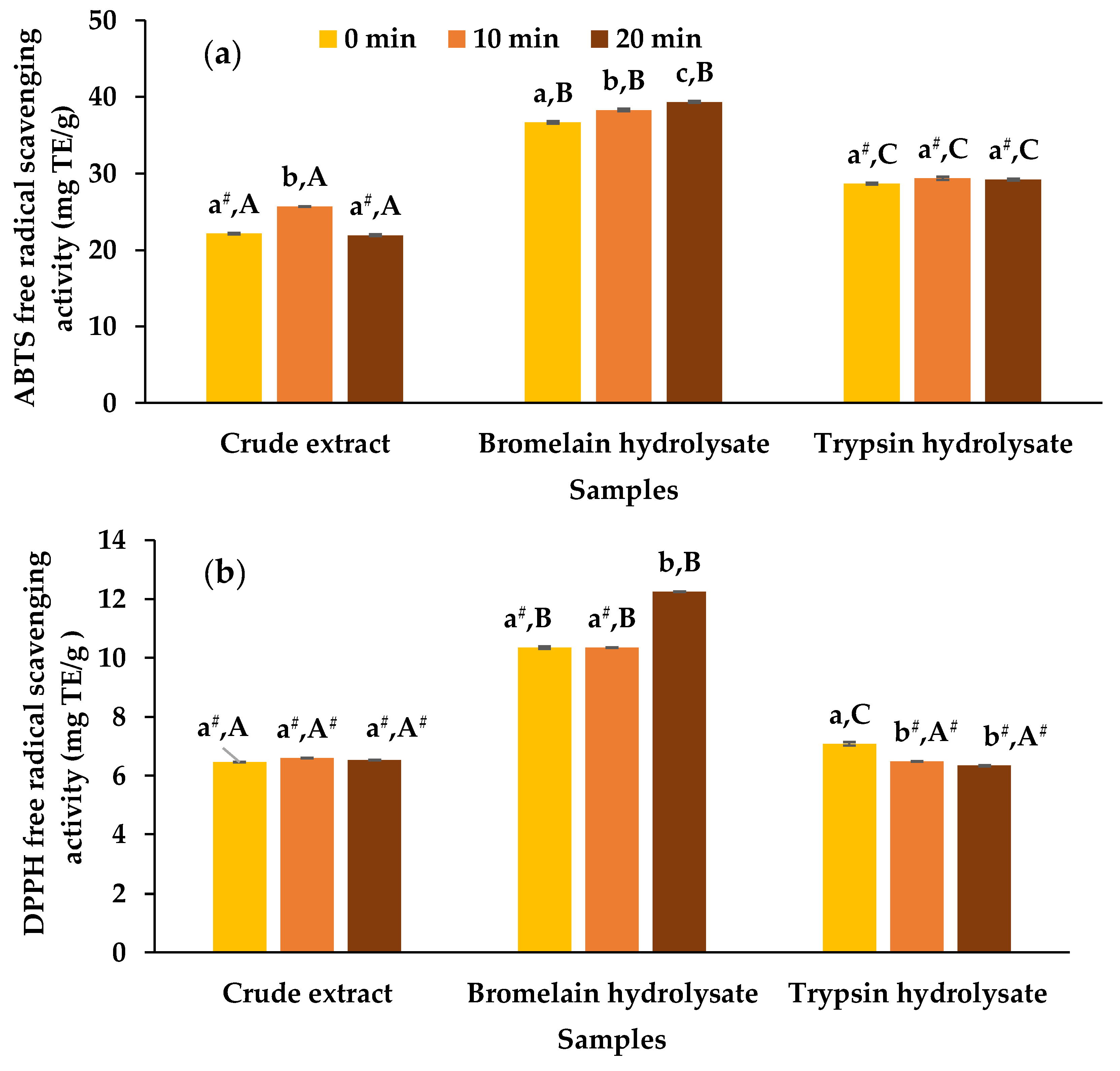
2.7. Determination of Antioxidant Activity
2.7.1. 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) diammonium salt (ABTS) Free Radical Scavenging Assay
2.7.2. 2,2-diphenyl-1-picrylhydrazyl (DPPH) Free Radical Scavenging Assay
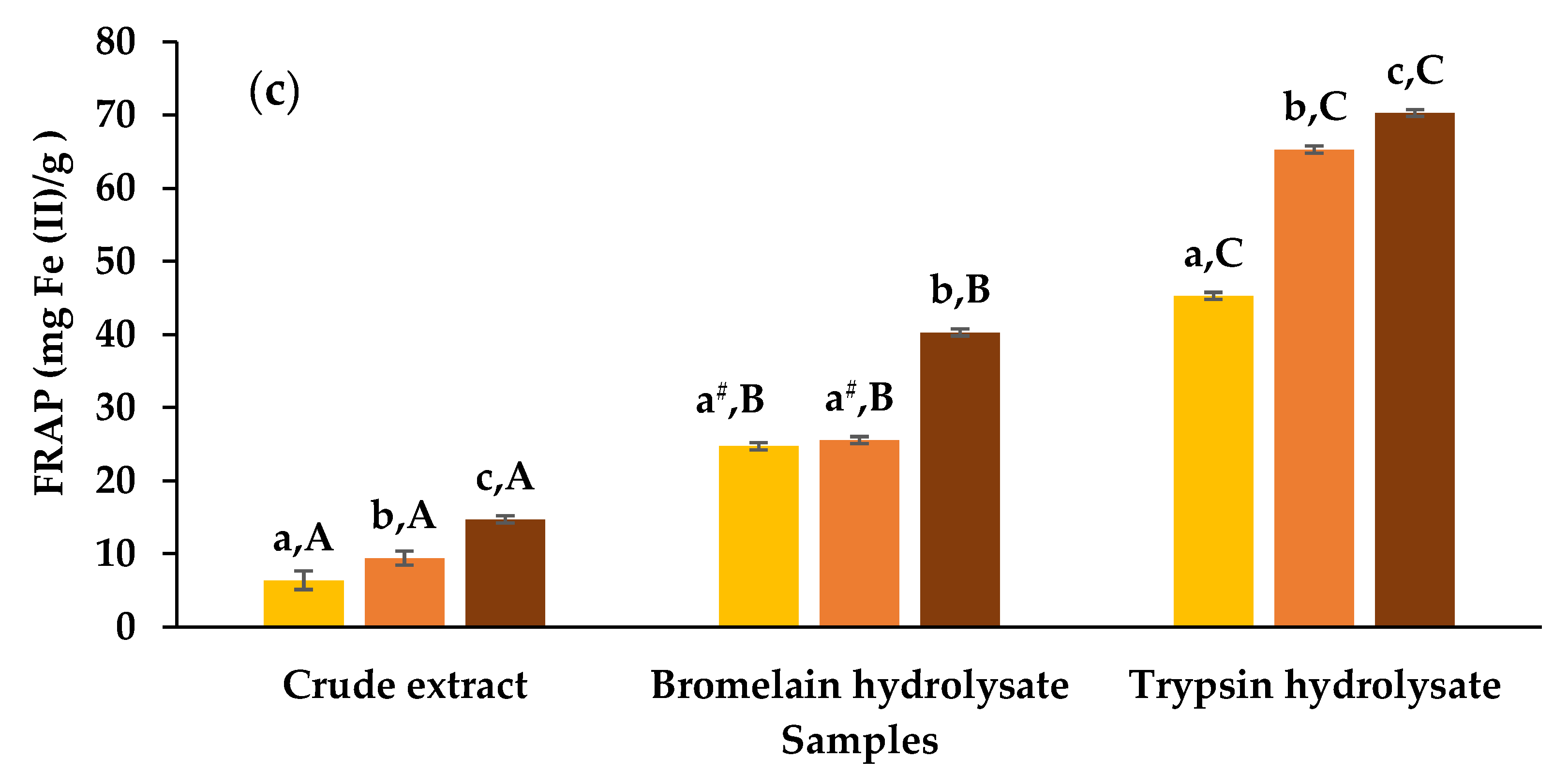
2.7.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.8. Well-Diffusion Antibacterial Assay
2.9. Statistical Analysis
3. Results and Discussion
3.1. Yield, TPC, and TFC
3.2. Antioxidant Activities of Crude Extracts and Hydrolysates
3.3. Antibacterial Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Najiya, A.; Alugoju, P.; Krishnaswamy, V.; Latha, P. Phytochemical analysis, antioxidant and antibacterial activity of Clitoria ternatea. IJBTT 2017, 7, 10–21. [Google Scholar]
- Neda, G.D.; Rabeta, M.S.; Ong, M.T. Chemical composition and anti-proliferative properties of flowers of Clitoria ternatea. Int. Food Res. J. 2013, 20, 1229–1234. [Google Scholar]
- Zingare, M.; Zingare, P.; Dubey, A.; Ansari, M. Clitoria ternatea (aparajita): A review of the antioxidant, antidiabetic and hepatoprotective potentials. Int. J. Pharm. Biol. Sci. 2013, 3, 203–213. [Google Scholar]
- Makasana, J.; Dholakiya, B.; Gajbhiye, N.; Raju, S. Extractive determination of bioactive flavonoids from butterfly pea (Clitoria ternatea Linn.). Res. Chem. Intermediat. 2016, 43, 783–799. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef] [PubMed]
- El Anany, A. Nutritional composition, antinutritional factors, bioactive compounds and antioxidant activity of guava seeds (Psidium Myrtaceae) as affected by roasting processes. J. Food Sci. Technol. 2013, 52, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, C.; Balachandran, I. Total phenolics and total flavonoids in selected Indian medicinal plants. Indian J. Pharm. Sci. 2012, 74, 258–260. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, S. Antioxidant activity and phenolic compounds in selected herbs. J. Agri. Food Chem. 2001, 49, 5165–5170. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 1–19. [Google Scholar] [CrossRef]
- Pham-Huy, L.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Cowan, M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.T.; Tan, Y.N.; Ee, K.Y.; Wong, F.C. Seeds, fermented plant foods, and agricultural by-products as sources of plant-derived antibacterial peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, S162–S177. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Tian, W.; Li, H.; Cao, J.; Jiang, W. Improving antioxidant activities of whey protein hydrolysates obtained by thermal preheat treatment of pepsin, trypsin, alcalase and flavourzyme. Int. J. Food Sci. Technol. 2012, 47, 2045–2051. [Google Scholar] [CrossRef]
- Liu, L.; Wen, W.; Zhang, R.; Wei, Z.; Deng, Y.; Xiao, J.; Zhang, M. Complex enzyme hydrolysis releases antioxidative phenolics from rice bran. Food Chem. 2017, 214, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Malaguti, M.; Dinelli, G.; Leoncini, E.; Bregola, V.; Bosi, S.; Cicero, A.; Hrelia, S. Bioactive peptides in cereals and legumes: Agronomical, biochemical and clinical aspects. Int. J. Mol. Sci. 2014, 15, 21120–21135. [Google Scholar] [CrossRef] [PubMed]
- Salas, C.; Badillo-Corona, J.; Ramírez-Sotelo, G.; Oliver-Salvador, C. Biologically active and antimicrobial peptides from plants. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef]
- Aluko, R.E. Antihypertensive peptides from food protein. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef]
- Thidarat, S.; Udomsak, M.; Jindawan, W.; Namphung, D.; Suneerat, Y.; Sawan, T.; Pisamai, T. Effect of roasting on phytochemical properties of Thai soybeans. Int. Food Res. J. 2016, 23, 606–612. [Google Scholar]
- Ribeiro, B.; Barreto, D.; Coelho, M. Enzyme-enhanced extraction of phenolic compounds and proteins from flaxseed meal. ISRN Biotechnol. 2013, 2013. [Google Scholar] [CrossRef]
- Kavitha, S.; Parimalavalli, R. Effect of processing methods on proximate composition of cereal and legume flours. J. Human Nutr. Food Sci. 2014, 2, 1051. [Google Scholar]
- Olanipekun, O.; Omenna, E.; Olapade, O.; Suleiman, P.; Omodara, O. Effect of boiling and roasting on the nutrient composition of kidney beans seed flour. Sky J. Food Sci. 2015, 4, 24–29. [Google Scholar]
- Vaz Patto, M.; Amarowicz, R.; Aryee, A.; Boye, J.; Chung, H.; Martín-Cabrejas, M.; Domoney, C. Achievements and challenges in improving the nutritional quality of food legumes. Crit. Rev. Plant. Sci. 2014, 34, 105–143. [Google Scholar] [CrossRef]
- Kaur, T.; Kaur, A.; Grewal, R. Kinetics studies with fruit bromelain (Ananas comosus) in the presence of cysteine and divalent ions. J. Food Sci. Technol. 2014, 52, 5954–5960. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Hang, S.; Zhou, Y.; Liu, Q.; Yang, H. Bromelain kinetics and mechanism on myofibril from golden pomfret (Trachinotus blochii). J. Food Sci. 2018, 83, 2148–2158. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.; Ong, S.; Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Al-Saeedi, A.; Hossain, M. Total phenols, total flavonoids contents and free radical scavenging activity of seeds crude extracts of pigeon pea traditionally used in Oman for the treatment of several chronic diseases. Asian. Pac. J. Trop. Dis. 2015, 5, 316–321. [Google Scholar] [CrossRef]
- Baba, S.; Malik, S. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Ahmed, D.; Khan, M.; Saeed, R. Comparative analysis of phenolics, flavonoids, and antioxidant and antibacterial potential of methanolic, hexanic and aqueous extracts from Adiantum caudatum leaves. Antioxidants 2015, 4, 394–409. [Google Scholar] [CrossRef]
- Ng, W.J.; Lim, M.S. Anti-staphylococcal activity of Melaleuca honey. Southeast. Asian J. Trop. Med. Public Health. 2015, 6, 472–479. [Google Scholar]
- Shahidi, F.; Yeo, J.D. Insoluble-bound phenolic in food. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Aryee, A.; Boye, J. Improving the digestibility of lentil flours and protein isolate and characterization of their enzymatically prepared hydrolysates. Int. J. Food Prop. 2016, 19, 2649–2665. [Google Scholar] [CrossRef]
- Macedo, M.; Xavier-Filho, J. Purification and partial characterisation of trypsin inhibitors from seeds of Clitoria ternatea. J. Sci. Food Agri. 1992, 58, 55–58. [Google Scholar] [CrossRef]
- Je, J.; Lee, K.; Lee, M.; Ahn, C. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food Res. Int. 2009, 42, 1266–1272. [Google Scholar] [CrossRef]
- Selamassakul, O.; Laohakunjit, N.; Kerdchoechuen, O.; Ratanakhanokchai, K. A novel multi-biofunctional protein from brown rice hydrolysed by endo/endo-exoproteases. Food Funct. 2016, 7, 2635–2644. [Google Scholar] [CrossRef]
- Liu, B.; Chiang, P. Production of hydrolysate with antioxidative activity and functional properties by enzymatic hydrolysis of defatted sesame (Sesamum indicum L.). IJASE. 2008, 6, 73–83. [Google Scholar]
- Xu, B.; Yuan, S.; Chang, S. Comparative analyses of phenolic composition, antioxidant capacity, and color of cool season legumes and other selected food legumes. J. Food Sci. 2007, 72, S167–S177. [Google Scholar] [CrossRef]
- Bordbar, S.; Ebrahimpour, A.; Abdul Hamid, A.; Abdul Manap, M.; Anwar, F.; Saari, N. The improvement of the endogenous antioxidant property of stone fish (Actinopyga lecanora) tissue using enzymatic proteolysis. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Karamać, M.; Kosińska-Cagnazzo, A.; Kulczyk, A. Use of different proteases to obtain flaxseed protein hydrolysates with antioxidant activity. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Osman, A.; El-Araby, G.; Taha, H. Potential use as a bio-preservative from lupin protein hydrolysate generated by Alcalase in food system. J. Appl. Bio. Biotech. 2016, 4, 76–81. [Google Scholar]
- Hwang, C.; Chen, Y.; Luo, C.; Chiang, W. Antioxidant and antibacterial activities of peptide fractions from flaxseed protein hydrolysed by protease from Bacillus altitudinis HK02. Int. J. Food Sci. Technol. 2016, 51, 681–689. [Google Scholar] [CrossRef]
- Trang, H.; Pasuwan, P. Screening antimicrobial activity against pathogens from protein hydrolysate of rice bran and Nile Tilapia by-products. Int. Food Res. J. 2018, 25, 2157–2163. [Google Scholar]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta. 2009, 1794, 808–816. [Google Scholar] [CrossRef]
- Makinen, P.L.; Makinen, K.K. The Enterococcus faecalis extracellular metalloendopeptidase (EC 3.4.24.30; coccolysin) inactivates human endothelin at bonds involving hydrophobic amino acid residues. Biochem. Biophys. Res. Commun. 1994, 200, 981–985. [Google Scholar]
- Schmidtchen, A.; Frick, I.; Andersson, E.; Tapper, H.; Bjorck, L. Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol. Microbiol. 2002, 46, 157–168. [Google Scholar] [CrossRef]
- Nesuta, O.; Budesinsky, M.; Hadravova, R.; Monincova, L.; Humpolickova, J.; Cerovsky, V. How proteases from Enterococcus faecalis contribute to its resistance to short α-helical antimicrobial peptides. Pathog. Dis. 2017, 75. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Roasting Time (min) | Amount (mg) | Zone of Inhibition (mm)# | |||
|---|---|---|---|---|---|---|
| E. ca | K. pb | E. fc | S. ad | |||
| Ampicillin disc | na | 0.01 | - | - | 26 | 31 |
| Crude extract | 0 | 0.375–3 | - | - | - | - |
| 10 | 0.375–3 | - | - | - | - | |
| 20 | 0.375–3 | - | - | - | - | |
| Bromelain hydrolysate | 0 | 0.375 | - | - | - | 10 |
| 0.75 | - | - | - | 8 | ||
| 1.5 | - | - | - | 10 | ||
| 3 | - | - | - | 7 | ||
| 10 | 0.375 | - | - | - | 11 | |
| 0.75 | - | - | - | 8 | ||
| 1.5 | - | - | - | 8 | ||
| 3 | - | - | - | 8 | ||
| 20 | 0.375 | - | - | - | 8 | |
| 0.75 | - | - | - | - | ||
| 1.5 | - | - | - | - | ||
| 3 | - | - | - | - | ||
| Trypsin hydrolysate | 0 | 0.375–3 | - | - | - | - |
| 10 | 0.375–3 | - | - | - | - | |
| 20 | 0.375–3 | - | - | - | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ee, K.-Y.; Khoo, L.-Y.; Ng, W.-J.; Wong, F.-C.; Chai, T.-T. Effects of Bromelain and Trypsin Hydrolysis on the Phytochemical Content, Antioxidant Activity, and Antibacterial Activity of Roasted Butterfly Pea Seeds. Processes 2019, 7, 534. https://doi.org/10.3390/pr7080534
Ee K-Y, Khoo L-Y, Ng W-J, Wong F-C, Chai T-T. Effects of Bromelain and Trypsin Hydrolysis on the Phytochemical Content, Antioxidant Activity, and Antibacterial Activity of Roasted Butterfly Pea Seeds. Processes. 2019; 7(8):534. https://doi.org/10.3390/pr7080534
Chicago/Turabian StyleEe, Kah-Yaw, Li-Ying Khoo, Wen-Jie Ng, Fai-Chu Wong, and Tsun-Thai Chai. 2019. "Effects of Bromelain and Trypsin Hydrolysis on the Phytochemical Content, Antioxidant Activity, and Antibacterial Activity of Roasted Butterfly Pea Seeds" Processes 7, no. 8: 534. https://doi.org/10.3390/pr7080534
APA StyleEe, K.-Y., Khoo, L.-Y., Ng, W.-J., Wong, F.-C., & Chai, T.-T. (2019). Effects of Bromelain and Trypsin Hydrolysis on the Phytochemical Content, Antioxidant Activity, and Antibacterial Activity of Roasted Butterfly Pea Seeds. Processes, 7(8), 534. https://doi.org/10.3390/pr7080534