
Driving Sustainable Energy Co-Production: Gas Transfer and Pressure Dynamics Regulating Hydrogen and Carboxylic Acid Generation in Anaerobic Systems


 and
and
Abstract
1. Introduction
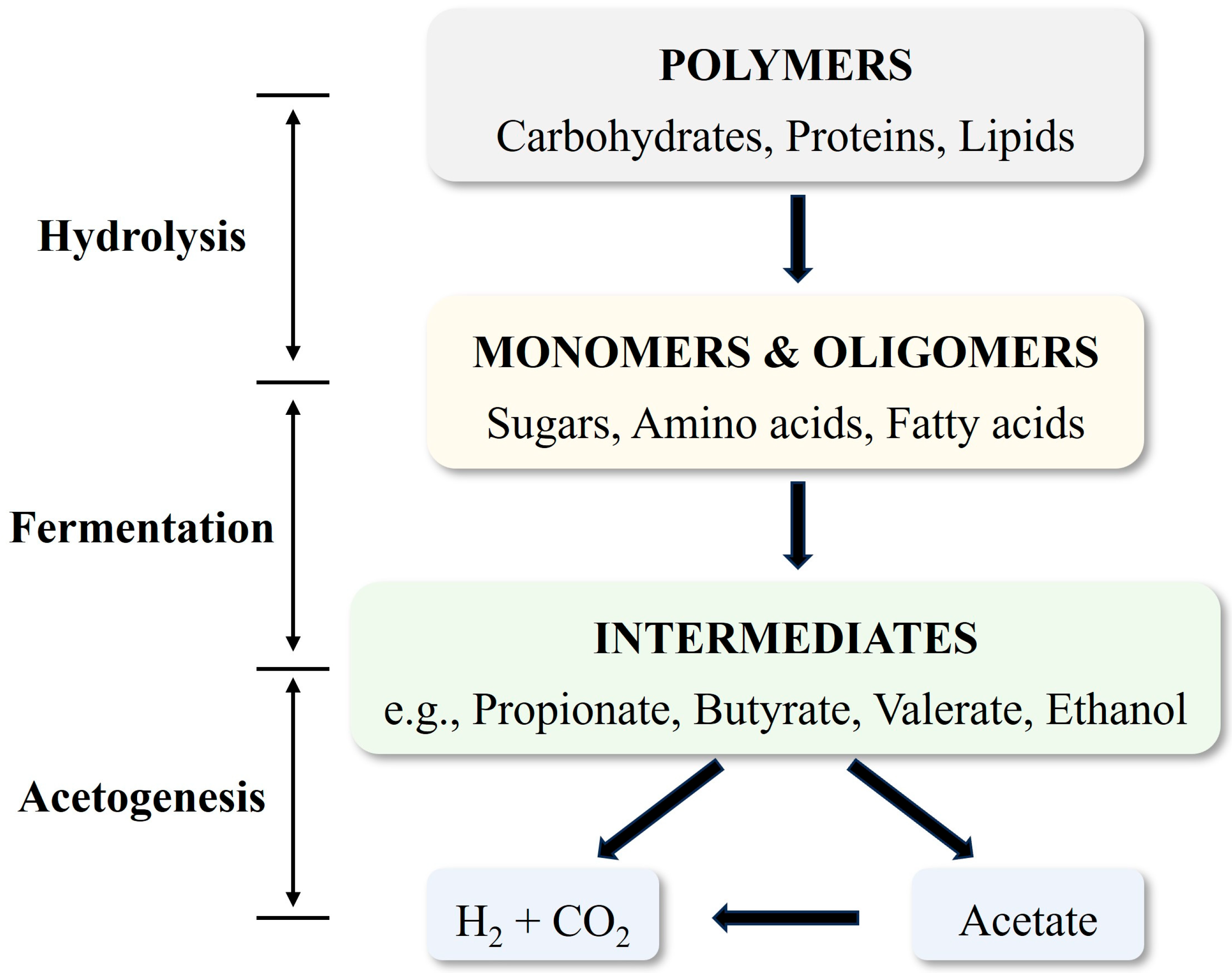
2. Biological Metabolism of Anaerobic Co-Production of Carboxylic Acids and Hydrogen
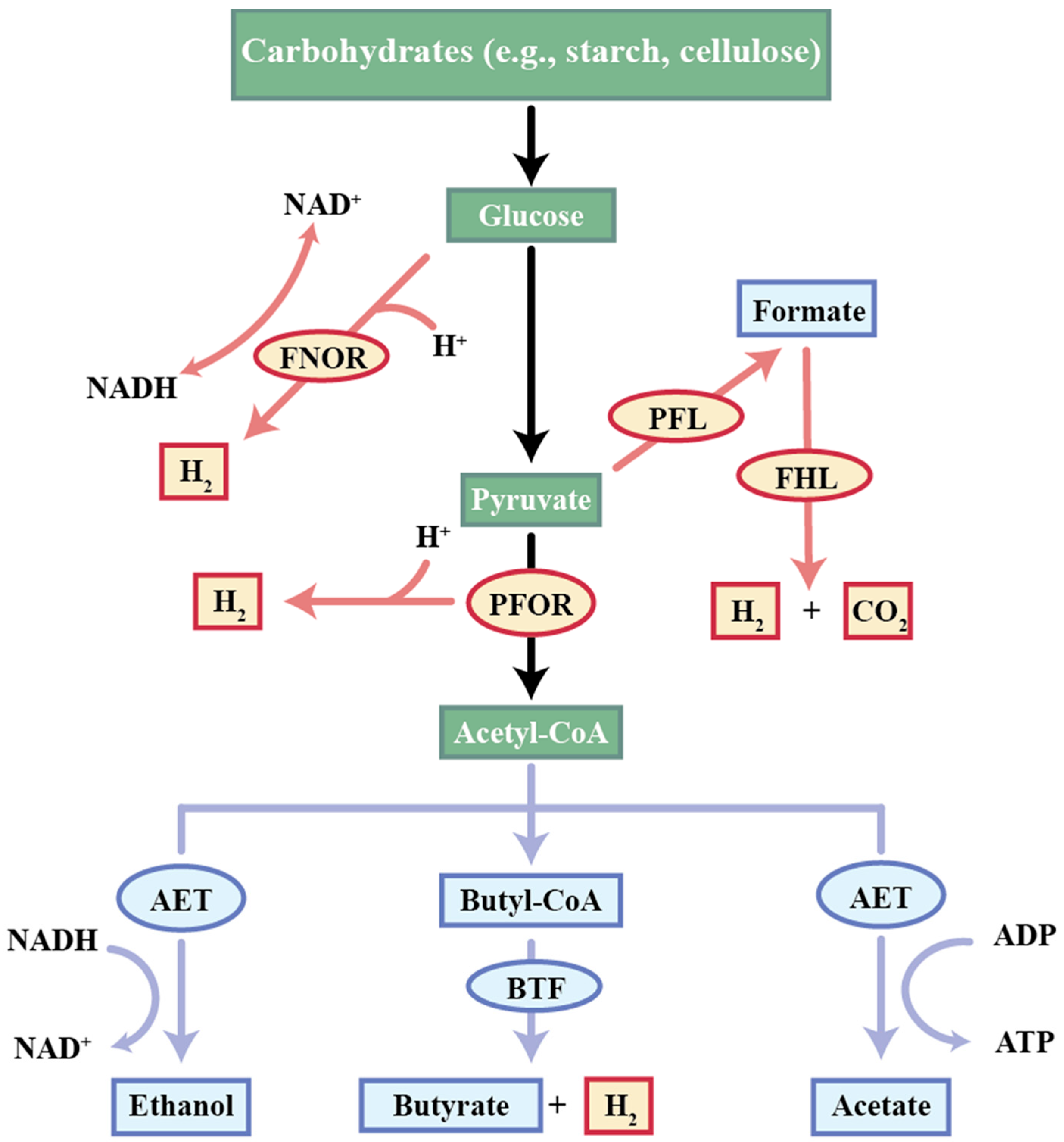
- Butyrate Fermentation
- Propionate Fermentation
- Acetate-Ethanol Fermentation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Operating Conditions | Key Microorganism | Main Metabolites | Co-Production Yield | Ref. |
|---|---|---|---|---|---|
| Waste activated sludge | With 90 mg/g VSS urea at a constant pH of 9.5 | Acinetobacter, Tissierella, and Petrimonas | H2, acetic and butyric acid | 24.57 mL/g VSS of H2 and 72.80 mg COD/g VSS of SCFAs. | [43] |
| Food waste | Control pH at 7 | Clostridium_sensu_stricto_1 | H2, acetic and butyric acid | 21.49 L/L of H2 and the production of butyrate increased to 42.13 g/L. | [44] |
| Food waste | Add 3 g/L lactic acid | Clostridium_sensu_stricto_12 | H2 and butyric acid | 57.02 ± 2.10 mL/g VSS of H2 | [45] |
| Melon and watermelon | A high HRT of 27 d, an organic loading rate of 3 g of VS per day | Ruminococcus | H2, iso-butyric, and caproic acids | 395.5 mL/g VS of H2, the content of iso-butyric and hexanoic acid can reach 76% of the SCFAs | [46] |
| Food waste | Add 8% oyster shells (w/w) | Lactobacillales, Gallicola, and Bacteroides | H2, acetic and butyric acid | 88.2 mL/g VS of H2, the highest concentrations of butyric and acetic acid can reach 8048.4 mg/L and 5604.6 mg/L, respectively. | [47] |
| Food waste | Add different levels of tar and BES | Clostridium_sensu_stricto and Clostridium_IV | H2, butyric and caproic acids | 65.0 mL/g VS of H2 (with a tar addition of 5 g/L), and the cumulative butyric acid production can reach 726.8 mg COD/g VS (with a tar addition of 30 g/L). | [48] |
3. Gas–Liquid Mass Transfer in Co-Production of Carboxylic Acids and Hydrogen
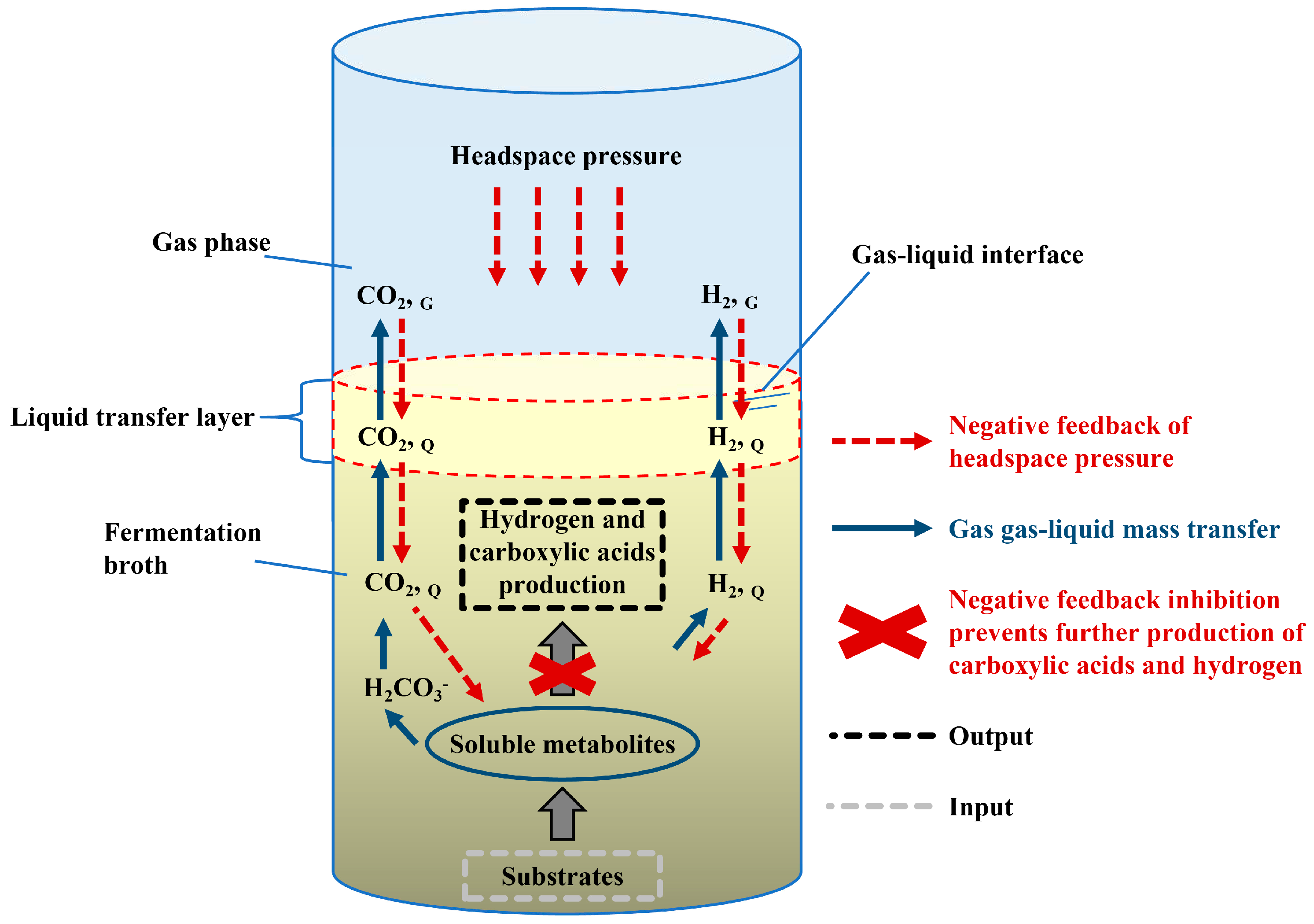
3.1. System Pressure in Co-Production of Carboxylic Acids and Hydrogen
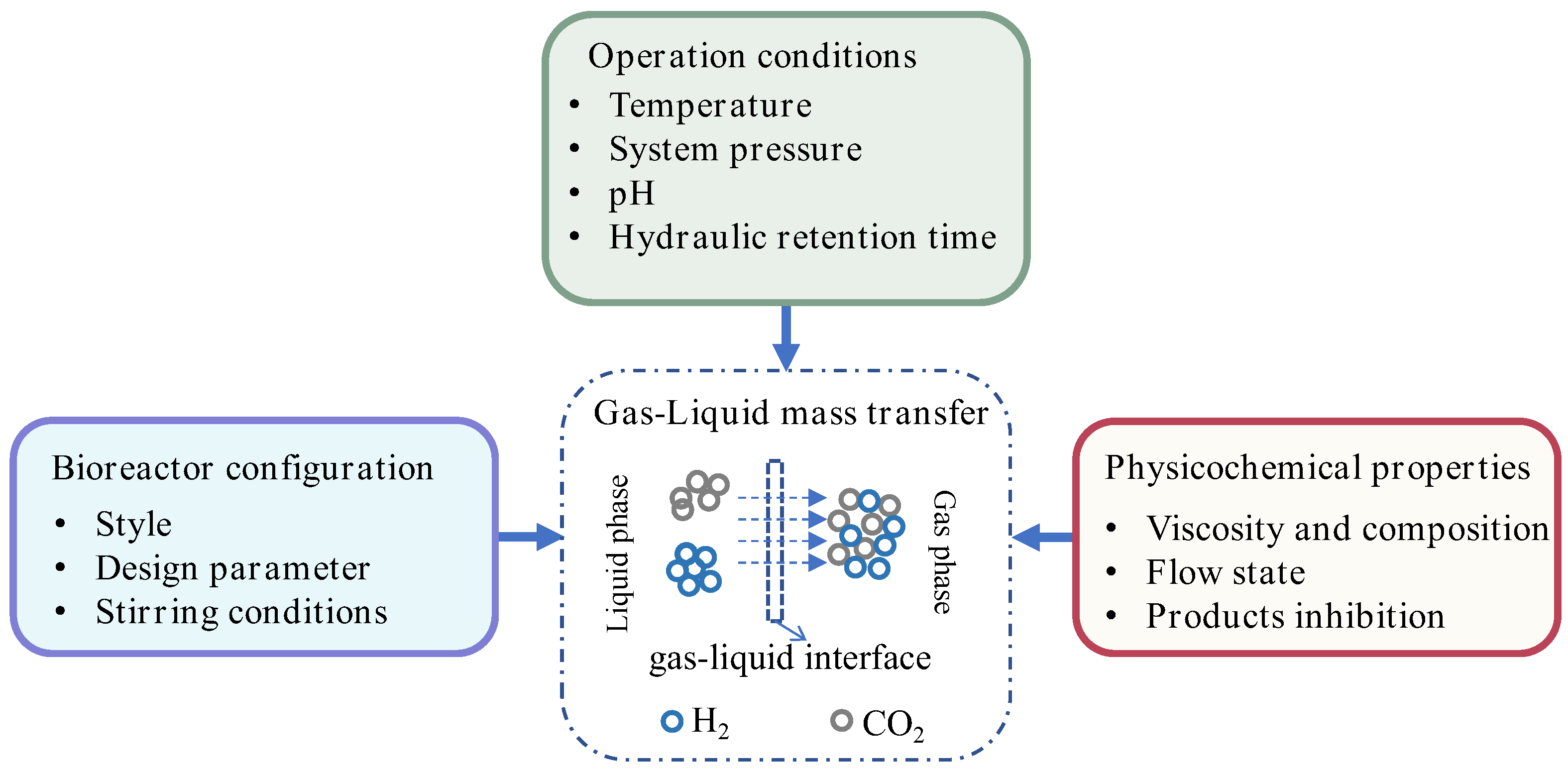
3.2. Mass Transfer Characteristic in Anaerobic Fermentation
3.3. Key Determinants of Interfacial Transport
4. Regulation of Hydrogen and Carboxylic Acid Co-Production
4.1. Regulation of Hydrogen and Carboxylic Acid Concentration
4.2. Bioreactor Configuration Optimization
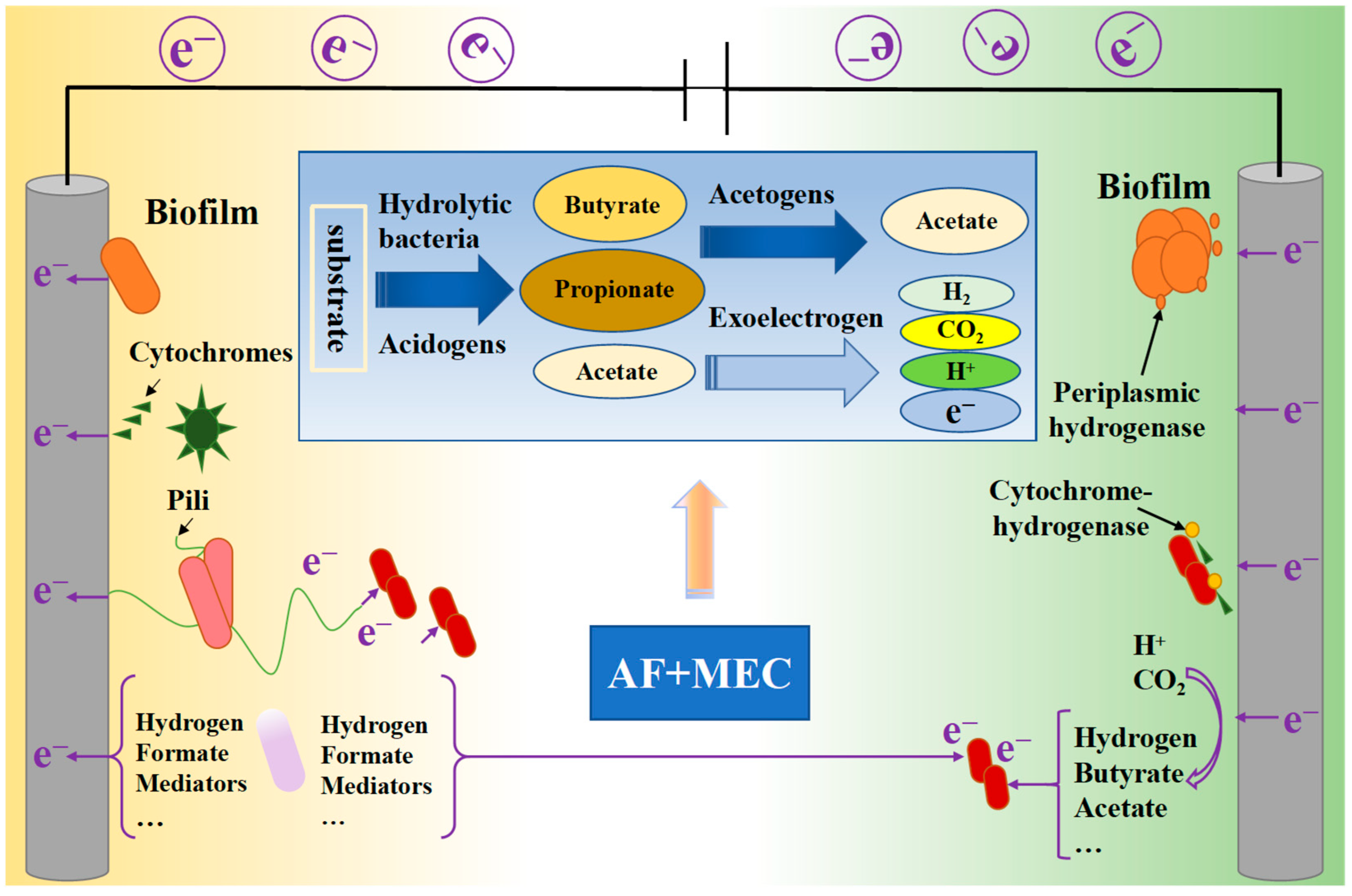
4.3. Coupling with Microbial Electrolysis Cell
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Anaerobic fermentation |
| NAD+ | Adenine dinucleotide ion |
| NADH | Nicotinamide adenine dinucleotide |
| Acetyl-CoA | Acetyl coenzyme A |
| PFL | Pyruvate formate lyase |
| PFOR | Pyruvate-ferredoxin oxidoreductase |
| FNOR | Ferredoxin-dependent NADP+ oxidoreductase |
| FHL | Formate hydrogen lyase |
| AET | Acetate–ethanol-type fermentation |
| BTF | Butyrate fermentation |
| PPF | Propionate fermentation |
| PTB | Phosphotransbutyrylase |
| BK | Butyrate kinase |
| APB | Acid-producing bacteria |
| HRT | Hydraulic retention time |
| SRT | Solid retention time |
| OLR | Organic loading rate |
| CSTR | Continuously stirred tank reactor |
| MBR | Membrane bioreactor |
| UASB | Upflow anaerobic sludge blanket |
| AFBR | Anaerobic fluidized bed reactor |
| PBR | Packed-bed bioreactor |
| MEC | Microbial electrolysis cell |
| COD | Chemical oxygen demand |
| Henry’s constant for gas i (m3·Pa·mol−1) | |
| Partial pressure of gas i in the gas phase (Pa) | |
| Dissolved concentration of gas i in the liquid phase (mol·m−3) | |
| NA | Mass transfer flux (mol·s−1·m2) |
| k | Mass transfer coefficient (m·s−1) |
| CAi − CA | Concentration difference between the two phases (mol·m3) |
| C0 | Initial concentration of substances (mol·m−3); |
| Cav | Concentration of substances at the gas–liquid interface (mol·m−3) |
| Ce | Equilibrium concentration of substances in the gas–liquid phase (mol·m−3) |
| Ef | Mass transfer efficiency |
| t | Mass transfer time (s) |
| KL | Liquid-side mass transfer coefficient (m·s−1) |
| a | Interface area (m2·m−3) |
| Q | Molar gas transfer rate (mol·s−1) |
| KLa | Volumetric mass transfer coefficient (s−1) |
| Partial pressure in the gas phase (Pa) | |
| Equilibrium partial pressure (Pa) | |
| H | Henry’s constant (Pa·m3·mol−1) |
References
- Posso, F.; Galeano, M.; Baranda, C.; Franco, D.; Rincón, A.; Zambrano, J.; Cavaliero, C.; Lópes, D. Towards the hydrogen economy in paraguay: Green hydrogen production potential and end-uses. Int. J. Hydrogen Energy 2022, 47, 30027–30049. [Google Scholar] [CrossRef]
- Nemestóthy, N.; Bélafi-Bakó, K.; Bakonyi, P. Enhancement of dark fermentative H2 production by gas separation membranes: A review. Bioresour. Technol. 2020, 302, 122828. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.; Gupta, P.; Saini, N.; Tiwari, A.K. Assessing the impact of renewable energy and non-renewable energy use on carbon emissions: Evidence from select developing and developed countries. Environ. Dev. Sustain. 2025, 27, 3059–3080. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Jin, Z.; Li, Z.; Liu, Q.; Zhang, K. Oil and gas pathway to net-zero: Review and outlook. Energy Strateg. Rev. 2023, 45, 101048. [Google Scholar] [CrossRef]
- Zhou, X.; Xie, F.; Li, H.; Zheng, C.; Zhao, X. Understanding inter-term fossil energy consumption pathways in China based on sustainable development goals. Geosci. Front. 2024, 15, 101687. [Google Scholar] [CrossRef]
- Fayyazbakhsh, A.; Bell, M.L.; Zhu, X.; Mei, X.; Koutný, M.; Hajinajaf, N.; Zhang, Y. Engine emissions with air pollutants and greenhouse gases and their control technologies. J. Clean. Prod. 2022, 376, 134260. [Google Scholar] [CrossRef]
- Liu, G.; Li, M.; Zhou, B.; Chen, Y.; Liao, S. General indicator for techno-economic assessment of renewable energy resources. Energy Convers. Manag. 2018, 156, 416–426. [Google Scholar] [CrossRef]
- Bhatt, A.H.; Ren, Z.J.; Tao, L. Value proposition of untapped wet wastes: Carboxylic acid production through anaerobic digestion. iScience 2020, 23, 101221. [Google Scholar] [CrossRef] [PubMed]
- Olokede, O.; Wu, H.; Holtzapple, M. Valorizing prickly pear cladodes via methane-arrested anaerobic digestion for carboxylic acid production. Biotechnol. Prog. 2022, 38, e3289. [Google Scholar] [CrossRef] [PubMed]
- Arimi, M.M.; Knodel, J.; Kiprop, A.; Namango, S.S.; Zhang, Y.; Geißen, S. Strategies for improvement of biohydrogen production from organic-rich wastewater: A review. Biomass Bioenerg. 2015, 75, 101–118. [Google Scholar] [CrossRef]
- Qian, Q.; Zhang, J.; Cui, M.; Han, B. Synthesis of acetic acid via methanol hydrocarboxylation with CO2 and H2. Nat. Commun. 2016, 7, 11481. [Google Scholar] [CrossRef] [PubMed]
- Kelbert, M.; Machado, T.O.; Araújo, P.H.H.; Sayer, C.; de Oliveira, D.; Maziero, P.; Simons, K.E.; Carciofi, B.A.M. Perspectives on biotechnological production of butyric acid from lignocellulosic biomass. Renew. Sustain. Energy Rev. 2024, 202, 114717. [Google Scholar] [CrossRef]
- Wang, Y.; Li, G.; Liu, Z.; Cui, P.; Zhu, Z.; Yang, S. Techno-economic analysis of biomass-to-hydrogen process in comparison with coal-to-hydrogen process. Energy 2019, 185, 1063–1075. [Google Scholar] [CrossRef]
- Boretti, A.; Banik, B.K. Advances in Hydrogen Production from Natural Gas Reforming. Adv. Energy Sustain. Res. 2021, 2, 2100097. [Google Scholar] [CrossRef]
- Shi, X.; Wu, L.; Wei, W.; Ni, B. Insights into the microbiomes for medium-chain carboxylic acids production from biowastes through chain elongation. Crit. Rev. Environ. Sci. Technol. 2022, 52, 3787–3812. [Google Scholar] [CrossRef]
- Das, D.; Veziroǧlu, T.N. Hydrogen production by biological processes: A survey of literature. Int. J. Hydrogen Energy 2001, 26, 13–28. [Google Scholar] [CrossRef]
- Chen, Y.; Yin, Y.; Wang, J. Influence of butyrate on fermentative hydrogen production and microbial community analysis. Int. J. Hydrogen Energy 2021, 46, 26825–26833. [Google Scholar] [CrossRef]
- Zhou, M.; Yan, B.; Wong, J.W.C.; Zhang, Y. Enhanced volatile fatty acids production from anaerobic fermentation of food waste: A mini-review focusing on acidogenic metabolic pathways. Bioresour. Technol. 2018, 248, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Zeng, H.; Gao, Y.; Mo, T.; Li, Y. Bio-hydrogen production by dark anaerobic fermentation of organic wastewater. Front. Chem. 2022, 10, 978907. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, C.; Zhao, W.; Mazarji, M.; Ren, L.; Liu, C.; Pan, J.; Yan, B. Anaerobic propionic acid production via succinate pathway at extremely low pH. Chem. Eng. J. 2024, 486, 150190. [Google Scholar] [CrossRef]
- Horiuchi, J.I.; Shimizu, T.; Tada, K.; Kanno, T.; Kobayashi, M. Selective production of organic acids in anaerobic acid reactor by pH control. Bioresour. Technol. 2002, 82, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, W.; Fang, S.; Li, Z.; Li, Z.; Wang, F.; Cheng, X.; Cao, J.; Feng, L.; Luo, J.; et al. Zinc pyrithione induced volatile fatty acids promotion derived from sludge anaerobic digestion: Interrelating the affected steps with microbial metabolic regulation and adaptive responses. Water Res. 2023, 234, 119816. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, S.; Lingam, Y.; Venkata Mohan, S. Understanding acidogenesis towards green hydrogen and volatile fatty acid production—Critical analysis and circular economy perspective. Chem. Eng. J. 2023, 464, 141550. [Google Scholar] [CrossRef]
- Ceron-Chafla, P.; Kleerebezem, R.; Rabaey, K.; van Lier, J.B.; Lindeboom, R.E.F. Direct and indirect effects of increased CO2 partial pressure on the bioenergetics of syntrophic propionate and butyrate conversion. Environ. Sci. Technol. 2020, 54, 12583–12592. [Google Scholar] [CrossRef] [PubMed]
- Abdelgadir, A.; Chen, X.; Liu, J.; Xie, X.; Zhang, J.; Zhang, K.; Wang, H.; Liu, N.; Tang, C. Characteristics, process parameters, and inner components of anaerobic bioreactors. BioMed Res. Int. 2014, 2014, 841573. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Nevin, K.P. Electrobiocommodities: Powering microbial production of fuels and commodity chemicals from carbon dioxide with electricity. Curr. Opin. Biotechnol. 2013, 24, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Strazzera, G.; Battista, F.; Garcia, N.H.; Frison, N.; Bolzonella, D. Volatile fatty acids production from food wastes for biorefinery platforms: A review. J. Environ. Manag. 2018, 226, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, J. Various additives for improving dark fermentative hydrogen production: A review. Renew. Sustain. Energy Rev. 2018, 95, 130–146. [Google Scholar] [CrossRef]
- Li, C.; Fang, H.H.P. Fermentative hydrogen production from wastewater and solid wastes by mixed cultures. Crit. Rev. Environ. Sci. Technol. 2007, 37, 1–39. [Google Scholar] [CrossRef]
- Voldsund, M.; Jordal, K.; Anantharaman, R. Hydrogen production with CO2 capture. Int. J. Hydrogen Energy 2016, 41, 4969–4992. [Google Scholar] [CrossRef]
- Lee, H.; Salerno, M.B.; Rittmann, B.E. Thermodynamic Evaluation on H2 Production in Glucose Fermentation. Environ. Sci. Technol. 2008, 42, 2401–2407. [Google Scholar] [CrossRef] [PubMed]
- Taheri, E.; Amin, M.; Pourzamani, H.; Fatehizadeh, A.; Ghasemian, M.; Bina, B. Comparison of acetate-butyrate and acetate-ethanol metabolic pathway in biohydrogen production. J. Med. Signals Sens. 2018, 8, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Saady, N.M.C. Homoacetogenesis during hydrogen production by mixed cultures dark fermentation: Unresolved challenge. Int. J. Hydrogen Energy 2013, 38, 13172–13191. [Google Scholar] [CrossRef]
- Rahimieh, A.; Nosrati, M. A review on biochemistry, microbiology and thermodynamic aspects of propionate: The key intermediate in the anaerobic digestion and wastewater treatment. Desalin Water Treat. 2024, 317, 100191. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, M.; Chen, Y.; Pan, Y. Achieving ethanol-type fermentation for hydrogen production in a granular sludge system by aeration. Bioresour. Technol. 2017, 224, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.; Sabaratnam, V.; Shirai, Y.; Hassan, M.A. Biohydrogen production from biomass and industrial wastes by dark fermentation. Int. J. Hydrogen Energy 2009, 34, 3277–3287. [Google Scholar] [CrossRef]
- Ciranna, A.; Ferrari, R.; Santala, V.; Karp, M. Inhibitory effects of substrate and soluble end products on biohydrogen production of the alkalithermophile Caloramator celer: Kinetic, metabolic and transcription analyses. Int. J. Hydrogen Energy 2014, 39, 6391–6401. [Google Scholar] [CrossRef]
- Khanal, S.K.; Chen, W.; Li, L.; Sung, S. Biological hydrogen production: Effects of pH and intermediate products. Int. J. Hydrogen Energy 2004, 29, 1123–1131. [Google Scholar] [CrossRef]
- Grupe, H.; Gottschalk, G. Physiological Events in Clostridium acetobutylicum during the Shift from Acidogenesis to Solventogenesis in Continuous Culture and Presentation of a Model for Shift Induction. Appl. Environ. Microb. 1992, 58, 3896–3902. [Google Scholar] [CrossRef] [PubMed]
- Beckers, L.; Masset, J.; Hamilton, C.; Delvigne, F.; Toye, D.; Crine, M.; Thonart, P.; Hiligsmann, S. Investigation of the links between mass transfer conditions, dissolved hydrogen concentration and biohydrogen production by the pure strain Clostridium butyricum CWBI1009. Biochem. Eng. J. 2015, 98, 18–28. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Kumar, G.; Bakonyi, P.; Kim, S.; Kobayashi, T.; Xu, K.Q.; Lakner, G.; Tóth, G.; Nemestóthy, N.; Bélafi-Bakó, K. A critical review on issues and overcoming strategies for the enhancement of dark fermentative hydrogen production in continuous systems. Int. J. Hydrogen Energy 2016, 41, 3820–3836. [Google Scholar] [CrossRef]
- Noblecourt, A.; Christophe, G.; Larroche, C.; Santa-Catalina, G.; Trably, E.; Fontanille, P. High hydrogen production rate in a submerged membrane anaerobic bioreactor. Int. J. Hydrogen Energy 2017, 42, 24656–24666. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, J.; Yuan, X.; Wang, D.; Luo, H.; Yang, R.; Wang, H. Urea promotes alkaline anaerobic fermentation of waste activated sludge for hydrogen production. Bioresour. Technol. 2025, 418, 131900. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Wang, D.; Wang, S.; Su, H.; Wang, Y. Regulatory mechanism of antioxidant enzymes on microbial metabolism and NADH in anaerobic fermentation of food waste for hydrogen production. J. Clean. Prod. 2024, 474, 143607. [Google Scholar] [CrossRef]
- Wang, X.; Ming, X.; Han, X.; Liu, Y.; Chen, M.; Zhang, T.; Li, X.; Zhang, D. Effect of lactic acid on short-chain fatty acids and hydrogen production during anaerobic fermentation of acidified food waste. Fuel 2025, 386, 134275. [Google Scholar] [CrossRef]
- Greses, S.; Tomás-Pejó, E.; González-Fernández, C. Short-chain fatty acids and hydrogen production in one single anaerobic fermentation stage using carbohydrate-rich food waste. J. Clean. Prod. 2021, 284, 124727. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, L.; Yuan, H.; Li, X.; Chang, Y.; Zuo, X. Oyster shells improve anaerobic dark fermentation performances of food waste: Hydrogen production, acidification performances, and microbial community characteristics. Bioresour. Technol. 2021, 335, 125268. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ji, X.; Zhu, K.; Wang, W.; Bao, Z.; Zhang, L. Biorefinery-oriented pyrolysis and anaerobic fermentation synergies: Leveraging biomass tar for enhanced chemical production from food waste. J. Clean. Prod. 2024, 477, 143830. [Google Scholar] [CrossRef]
- Sonnleitner, A.; Peintner, C.; Wukovits, W.; Friedl, A.; Schnitzhofer, W. Process investigations of extreme thermophilic fermentations for hydrogen production: Effect of bubble induction and reduced pressure. Bioresour. Technol. 2012, 118, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.B.; Pitt, L.; Love, M. Biohydrogen production: Prospects and limitations to practical application. Int. J. Hydrogen Energy 2004, 29, 173–185. [Google Scholar] [CrossRef]
- Valdez-Vazquez, I.; Ríos-Leal, E.; Carmona-Martínez, A.; Muñoz-Páez, K.M.; Poggi-Varaldo, H.M. Improvement of Biohydrogen Production from Solid Wastes by Intermittent Venting and Gas Flushing of Batch Reactors Headspace. Environ. Sci. Technol. 2006, 40, 3409–3415. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Liu, Y.; Li, Z. The synergistic hydrogen production of bicellular fermentation systems and fluid dynamics simulation in reactor under stirring. Bioresour. Technol. Rep. 2023, 22, 101473. [Google Scholar] [CrossRef]
- Oh, S.; Zuo, Y.; Zhang, H.; Guiltinan, M.J.; Logan, B.E.; Regan, J.M. Hydrogen production by Clostridium acetobutylicum ATCC 824 and megaplasmid-deficient mutant M5 evaluated using a large headspace volume technique. Int. J. Hydrogen Energy 2009, 34, 9347–9353. [Google Scholar] [CrossRef]
- Laurent, B.; Serge, H.; Julien, M.; Christopher, H.; Philippe, T. Effects of hydrogen partial pressure on fermentative biohydrogen production by a chemotropic Clostridium bacterium in a new horizontal rotating cylinder reactor. Energy Procedia 2012, 29, 34–41. [Google Scholar] [CrossRef]
- Castro-Carranza, A.; Vega-Hernández, P.; Nolasco, J.C.; Ladstätter-Weißenmayer, A.; Eickhoff, M.; Gutowski, J. Detection of Hydrogen Dissolved in Liquid Media: A Review and Outlook. Phys. Status Solidi A 2022, 219, 2100669. [Google Scholar] [CrossRef]
- Bundhoo, M.A.Z.; Mohee, R. Inhibition of dark fermentative bio-hydrogen production: A review. Int. J. Hydrogen Energy 2016, 41, 6713–6733. [Google Scholar] [CrossRef]
- Bakonyi, P.; Buitrón, G.; Valdez-Vazquez, I.; Nemestóthy, N.; Bélafi-Bakó, K. A novel gas separation integrated membrane bioreactor to evaluate the impact of self-generated biogas recycling on continuous hydrogen fermentation. Appl. Energy 2017, 190, 813–823. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimisation. Int. J. Hydrogen Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, X.; Hu, J.; Zhang, T.; Zhu, S.; Zhang, Q. Photo-bioreactor structure and light-heat-mass transfer properties in photo-fermentative bio-hydrogen production system: A mini review. Int. J. Hydrogen Energy 2017, 42, 12143–12152. [Google Scholar] [CrossRef]
- Sieborg, M.U.; Engelbrecht, N.; Singh, A.; Schnürer, A.; Ottosen, L.D.M.; Kofoed, M.V.W.; Sveriges, L. Unraveling the effects of temperature on mass transfer and microbiology in thermophilic and extreme thermophilic trickle bed biomethanation reactors. Chem. Eng. J. 2025, 509, 161179. [Google Scholar] [CrossRef]
- Ullrich, T.; Lindner, J.; Bär, K.; Mörs, F.; Graf, F.; Lemmer, A. Influence of operating pressure on the biological hydrogen methanation in trickle-bed reactors. Bioresour. Technol. 2018, 247, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ginkel, S.V.; Sung, S.; Lay, J. Biohydrogen Production as a Function of pH and Substrate Concentration. Environ. Sci. Technol. 2001, 35, 4726–4730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Y.; Chen, M.; Zeng, R.J. Hydrogen supersaturation in thermophilic mixed culture fermentation. Int. J. Hydrogen Energy 2012, 37, 17809–17816. [Google Scholar] [CrossRef]
- Maluta, F.; Paglianti, A.; Montante, G. Modelling of biohydrogen production in stirred fermenters by Computational Fluid Dynamics. Process Saf. Environ. Prot. 2019, 125, 342–357. [Google Scholar] [CrossRef]
- Gilbert, J.J.; Ray, S.; Das, D. Hydrogen production using Rhodobacter sphaeroides (O.U. 001) in a flat panel rocking photobioreactor. Int. J. Hydrogen Energy 2011, 36, 3434–3441. [Google Scholar] [CrossRef]
- Rodríguez-Reyes, J.J.; García-Depraect, O.; Cantera, S.; Mena-Navarro, V.; León-Becerril, E. Assessment of the recovery of hydrogen production activity in dark fermentation reactors after a long period of shutdown. Int. J. Hydrogen Energy 2025, 144, 1134–1146. [Google Scholar] [CrossRef]
- Clark, I.C.; Zhang, R.H.; Upadhyaya, S.K. The effect of low pressure and mixing on biological hydrogen production via anaerobic fermentation. Int. J. Hydrogen Energy 2012, 37, 11504–11513. [Google Scholar] [CrossRef]
- Song, Z.; Dai, Y.; Fan, Q.; Li, X.; Fan, Y.; Hou, H. Effects of pretreatment method of natural bacteria source on microbial community and bio-hydrogen production by dark fermentation. Int. J. Hydrogen Energy 2012, 37, 5631–5636. [Google Scholar] [CrossRef]
- Dreschke, G.; Papirio, S.; Lens, P.N.L.; Esposito, G. Influence of liquid-phase hydrogen on dark fermentation by Thermotoga neapolitana. Renew. Energy 2019, 140, 354–360. [Google Scholar] [CrossRef]
- Ananthi, V.; Bora, A.; Ramesh, U.; Yuvakkumar, R.; Raja, K.; Ponnuchamy, K.; Muthusamy, G.; Arun, A. A review on the technologies for sustainable biohydrogen production. Process Saf. Environ. 2024, 186, 944–956. [Google Scholar] [CrossRef]
- Sekoai, P.T.; Yoro, K.O.; Daramola, M.O. Effect of nitrogen gas sparging on dark fermentative biohydrogen production using suspended and immobilized cells of anaerobic mixed bacteria from potato waste. Biofuels 2018, 9, 595–604. [Google Scholar] [CrossRef]
- Winkler, J.; Neuner, T.; Hupfauf, S.; Arthofer, A.; Ebner, C.; Rauch, W.; Bockreis, A. Impact of impeller design on anaerobic digestion: Assessment of mixing dynamics, methane yield, microbial communities and digestate dewaterability. Bioresour. Technol. 2024, 406, 131095. [Google Scholar] [CrossRef] [PubMed]
- Clagnan, E.; Adani, F. Influence of feedstock source on the development of polyhydroxyalkanoates-producing mixed microbial cultures in continuously stirred tank reactors. New Biotechnol. 2023, 76, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Arslan, D.; Steinbusch, K.J.J.; Diels, L.; De Wever, H.; Hamelers, H.V.M.; Buisman, C.J.N. Selective carboxylate production by controlling hydrogen, carbon dioxide and substrate concentrations in mixed culture fermentation. Bioresour. Technol. 2013, 136, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Wang, Q.; Li, H.; Zhang, Y.; Deng, Z.; Liu, J.; Du, X. Effect of fermentation type regulation using alkaline addition on two-phase anaerobic digestion of food waste at different organic load rates. Renew. Energy 2020, 154, 385–393. [Google Scholar] [CrossRef]
- Dahiya, S.; Mohan, S.V. Selective control of volatile fatty acids production from food waste by regulating biosystem buffering: A comprehensive study. Chem. Eng. J. 2019, 357, 787–801. [Google Scholar] [CrossRef]
- Zhu, X.; Leininger, A.; Jassby, D.; Tsesmetzis, N.; Ren, Z.J. Will membranes break barriers on volatile fatty acid recovery from anaerobic digestion? ACS EST Eng. 2021, 1, 141–153. [Google Scholar] [CrossRef]
- Liu, G.; Lu, H.; Wu, K.; Guan, G.; Liang, B. Co-production of high-purity hydrogen and value-added molecules from cellulose under alkaline environment: Mechanism and application. Renew. Energy 2025, 252, 123472. [Google Scholar] [CrossRef]
- Song, S.; Ginige, M.P.; Yu Cheng, K.; Peacock, C.S.; Kaksonen, A.H. Ultrasonication-induced metabolic pathway shifts and reduced electron carrier washout with biomass enhanced hydrogen yield in a continuous stirred tank reactor. Chem. Eng. J. 2024, 493, 152594. [Google Scholar] [CrossRef]
- Oshiki, M.; Yamaguchi, G.; Takahashi, K.; Okabe, S.; Kawano, S.; Nakagawa, J.; Fukushima, T. Thermophilic dark fermentation for hydrogen and volatile fatty acids production from breadcrumbs. Chem. Eng. J. 2024, 501, 157633. [Google Scholar] [CrossRef]
- Naccache, G.; Paraschivoiu, M. Parametric study of the dual vertical axis wind turbine using CFD. J. Wind. Eng. Ind. Aerod. 2018, 172, 244–255. [Google Scholar] [CrossRef]
- Mokhtarani, B.; Zanganeh, J.; Moghtaderi, B. A Review on Biohydrogen Production Through Dark Fermentation, Process Parameters and Simulation. Energies 2025, 18, 1092. [Google Scholar] [CrossRef]
- Meersseman Arango, H.; Luis, P.; Leyssens, T.; Debecker, D.P. Enzyme-membrane reactors: Recent trends and applications for the production of fine chemicals and pharmaceutical building blocks. Comptes Rendus Chim. 2025, 28, 151–170. [Google Scholar] [CrossRef]
- Renaudie, M.; Clion, V.; Dumas, C.; Vuilleumier, S.; Ernst, B. Intensification and optimization of continuous hydrogen production by dark fermentation in a new design liquid/gas hollow fiber membrane bioreactor. Chem. Eng. J. 2021, 416, 129068. [Google Scholar] [CrossRef]
- Bérubé, P.R.; Lei, E. The effect of hydrodynamic conditions and system configurations on the permeate flux in a submerged hollow fiber membrane system. J. Membrane Sci. 2006, 271, 29–37. [Google Scholar] [CrossRef]
- Zhang, K.; Cui, Z.; Field, R.W. Effect of bubble size and frequency on mass transfer in flat sheet MBR. J. Membrane Sci. 2009, 332, 30–37. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, X.; Guan, J.; He, Z. Volatile Fatty Acid Production through Arresting Methanogenesis by Electro-Synthesized Hydrogen Peroxide in Anaerobic Digestion and Subsequent Recovery by Electrodialysis. ACS ES T Eng. 2024, 4, 2964–2973. [Google Scholar] [CrossRef]
- Show, K.Y.; Lee, D.J.; Tay, J.H.; Lin, C.Y.; Chang, J.S. Biohydrogen production: Current perspectives and the way forward. Int. J. Hydrogen Energy 2012, 37, 15616–15631. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, W.; Zhang, Y.; Yan, R.; Xie, Y.; Zhang, D.; Jia, F.; Yang, L.; Zaib, S.; Li, R.; et al. Effectiveness and mechanism of using sodium alginate-magnesium silicate carrier in UASB reactor to resist shock loading of coking wastewater. J. Clean. Prod. 2025, 506, 145519. [Google Scholar] [CrossRef]
- Afridi, Z.U.R.; Wu, J.; Li, Z.H.; Akand, R.; Cao, Z.P.; Poncin, S.; Li, H.Z. Novel insight of spatial mass transfer conditions of upflow anaerobic reactor. J. Clean. Prod. 2018, 204, 390–398. [Google Scholar] [CrossRef]
- Łukajtis, R.; Hołowacz, I.; Kucharska, K.; Glinka, M.; Rybarczyk, P.; Przyjazny, A.; Kamiński, M. Corrigendum to “Hydrogen production from biomass using dark fermentation” [Renew Sustain Energy Rev 91 (2018) 665–94]. Renew. Sustain. Energy Rev. 2018, 95, 354. [Google Scholar] [CrossRef]
- Tugtas, A.E.; Yesil, H.; Calli, B. Enhanced anaerobic digestion model no.1 for high solids fermentation: Integrating homoacetogenesis and chain elongation. Bioresour. Technol. 2025, 417, 131843. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Fu, Y.; Song, Y.; Wu, Q.; Cao, W.; Liu, Z.; Wei, X. Study on the process and performance of anaerobic circulating fluidized bed: Effects of fluidization velocity and particle circulation rate on hydrogen production. Int. J. Hydrogen Energy 2024, 91, 343–353. [Google Scholar] [CrossRef]
- Encina, P.A.G.; Hidalgo, M.D. Influence of substrate feed patterns on biofilm development in anaerobic fluidized bed reactors (AFBR). Process Biochem. 2005, 40, 2509–2516. [Google Scholar] [CrossRef]
- Han, W.; Hu, Y.; Li, S.; Nie, Q.; Zhao, H.; Tang, J. Effect of organic loading rate on dark fermentative hydrogen production in the continuous stirred tank reactor and continuous mixed immobilized sludge reactor from waste pastry hydrolysate. Waste Manag. 2016, 58, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Mendoza, L.J.; García-Depraect, O.; Muñoz, R. Unlocking the high-rate continuous performance of fermentative hydrogen bioproduction from fruit and vegetable residues by modulating hydraulic retention time. Bioresour. Technol. 2023, 373, 128716. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Lavagnolo, M.C.; Spagni, A. Biological hydrogen production via dark fermentation by using a side-stream dynamic membrane bioreactor: Effect of substrate concentration. Chem. Eng. J. 2018, 349, 719–727. [Google Scholar] [CrossRef]
- Sivagurunathan, P.; Anburajan, P.; Kumar, G.; Kim, S. Effect of hydraulic retention time (HRT) on biohydrogen production from galactose in an up-flow anaerobic sludge blanket reactor. Int. J. Hydrogen Energy 2016, 41, 21670–21677. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Azahar, A.M.; Tan, J.P.; Jahim, J.M.; Abdul, P.M.; Mastar, M.S.; Anuar, N.; Mohammed Yunus, M.F.; Asis, A.J.; Wu, S. Operation performance of up-flow anaerobic sludge blanket (UASB) bioreactor for biohydrogen production by self-granulated sludge using pre-treated palm oil mill effluent (POME) as carbon source. Renew. Energy 2019, 134, 1262–1272. [Google Scholar] [CrossRef]
- Rydén, M.; Arjmand, M. Continuous hydrogen production via the steam–iron reaction by chemical looping in a circulating fluidized-bed reactor. Int. J. Hydrogen Energy 2012, 37, 4843–4854. [Google Scholar] [CrossRef]
- Park, S.; Rhee, C.; Jadhav, D.A.; Jang, J.; Hwang, M.; Chae, K. Enhanced hydrogen production in microbial electrolysis cells through a magnetically induced electroactive anode biofilm. Chem. Eng. J. 2025, 505, 159071. [Google Scholar] [CrossRef]
- Guerrero-Sodric, O.; Baeza, J.A.; Guisasola, A. Enhancing bioelectrochemical hydrogen production from industrial wastewater using Ni-foam cathodes in a microbial electrolysis cell pilot plant. Water Res. 2024, 256, 121616. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Luo, S.; Yuan, R.; Hou, R.; Zhou, B.; Chen, H. Cathode materials and novel strategies for improving bioenergy production in microbial electrolysis cell: A review. J. Environ. Chem. Eng. 2025, 13, 115718. [Google Scholar] [CrossRef]
- Lee, H.; Xin, W.; Katakojwala, R.; Venkata Mohan, S.; Tabish, N.M.D. Microbial electrolysis cells for the production of biohydrogen in dark fermentation—A review. Bioresour. Technol. 2022, 363, 127934. [Google Scholar] [CrossRef] [PubMed]
- Chookaew, T.; Prasertsan, P.; Ren, Z.J. Two-stage conversion of crude glycerol to energy using dark fermentation linked with microbial fuel cell or microbial electrolysis cell. New Biotechnol. 2014, 31, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Chaitanya, N.K.; Rajpurohit, A.; Nair, P.S.; Chatterjee, P. Enhanced carbon capture and medium chain fatty acid production using microbial electrosynthesis: Role of electrode surface area. Bioresour. Technol. 2025, 435, 132916. [Google Scholar] [CrossRef] [PubMed]
- Bian, B.; Bajracharya, S.; Xu, J.; Pant, D.; Saikaly, P.E. Microbial electrosynthesis from CO2: Challenges, opportunities and perspectives in the context of circular bioeconomy. Bioresour. Technol. 2020, 302, 122863. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Kim, B.H.; Li, D.; Feng, Y.; Daud, W.R.W.; Scott, K.; Yu, E.H. Effects of applied potential and reactants to hydrogen-producing biocathode in a aicrobial electrolysis Ccell. Front. Chem. 2018, 6, 318. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Leininger, A.; May, H.D.; Ren, Z.J. H2 mediated mixed culture microbial electrosynthesis for high titer acetate production from CO2. Environ. Sci. Ecotechnol. 2024, 19, 100324. [Google Scholar] [CrossRef] [PubMed]
- Thulluru, L.P.; Dhanda, A.; Doki, M.M.; Ghangrekar, M.M.; Chowdhury, S. Sludge-derived hydrochar as a potential electrocatalyst for improved CO2 reduction in microbial electrosynthesis. RSC Sustain. 2025, 3, 471–485. [Google Scholar] [CrossRef]
- Yao, H.; Romans-Casas, M.; Vassilev, I.; Rinta-Kanto, J.M.; Puig, S.; Rissanen, A.J.; Kokko, M. Selective butyrate production from CO2 and methanol in microbial electrosynthesis—Influence of pH. Bioelectrochemistry 2025, 165, 109000. [Google Scholar] [CrossRef] [PubMed]
- Varghese, V.K.; Poddar, B.J.; Shah, M.P.; Purohit, H.J.; Khardenavis, A.A. A comprehensive review on current status and future perspectives of microbial volatile fatty acids production as platform chemicals. Sci. Total Environ. 2022, 815, 152500. [Google Scholar] [CrossRef] [PubMed]
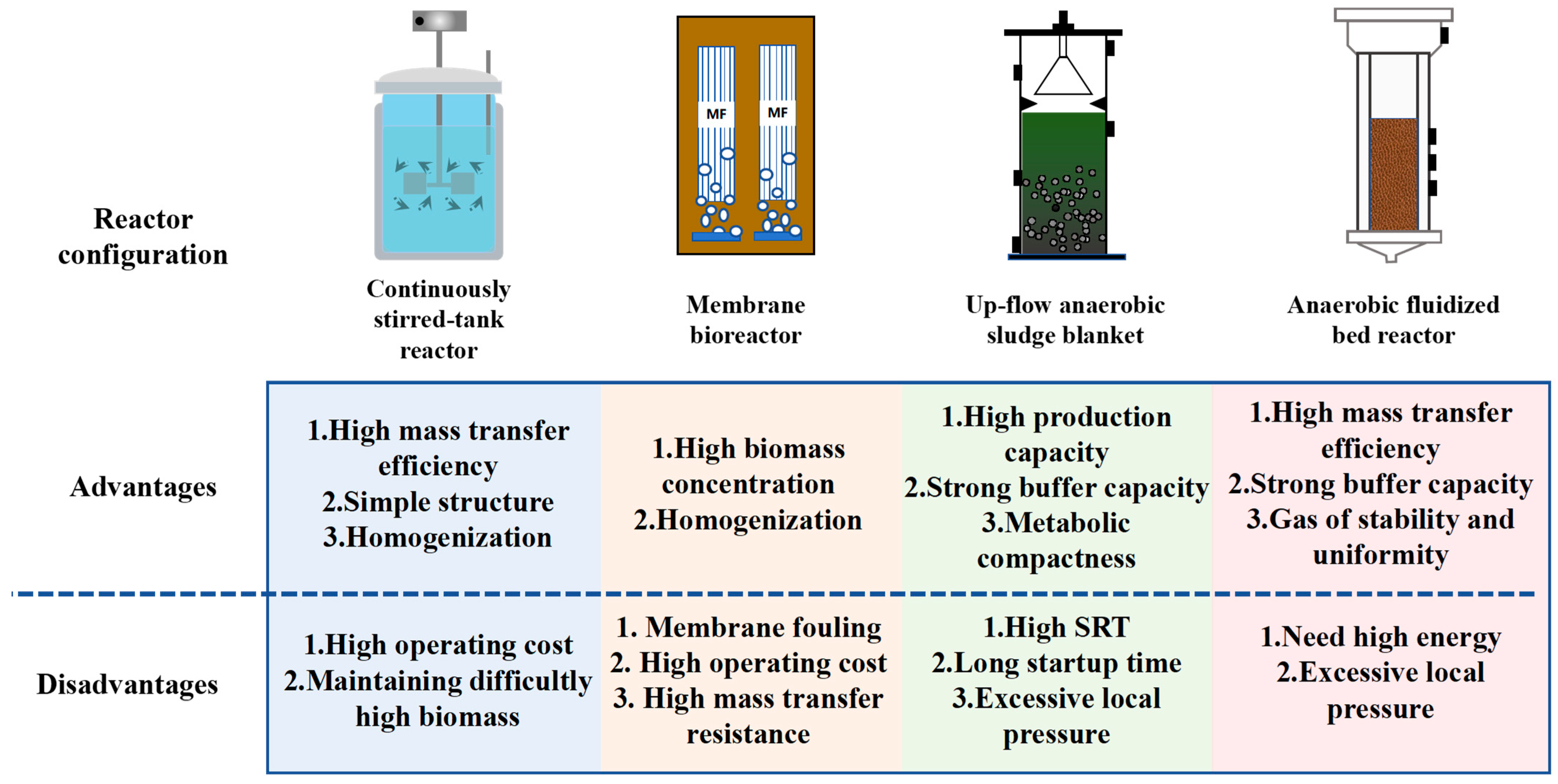
| Regulation Techniques | Advantages | Disadvantages | Ref. |
|---|---|---|---|
| Gas stripping | 1. Accelerating system mass transfer 2. Relieving the inhibition of hydrogen partial pressure | 1. Dilution of the generated biogas 2. The difficult of separating N2 and H2 3. CO2 stripping induces system acidification through carbonic acid dissociation | [49,57] |
| Vigorous stir | 1. Accelerating system mass transfer 2. Increasing the contact area between microorganisms and substrates | 1. Increasing operating costs 2. Breaking down the biological flocs | [69] |
| Reactor Type | Substrate | H2 Production Rate | Yield of Carboxylic acid | Ref. |
|---|---|---|---|---|
| CSTR | Waste pastry hydrolysate | The maximum hydrogen production rate (HPR) of CSTR (277.76 mL/(h·L) was achieved at OLR of 24 kgCOD/(m3·d) | Acetic acid (HAc) serves as the primary product, reaching a concentration of up to 13.85 mmol/L. | [95] |
| CSTR | Fruit-vegetable waste | When HRT is 6 h, the highest HPR can reach 11.8 NL·L-d−1. | Lactic acid is the main product, and its concentration can reach up to 18.4 ± 1.2 g/L. | [96] |
| CSTR | Breadcrumbs | The highest HPR was 7.0 L-H2·L−1·d−1 | 181 mM of HAc | [80] |
| MBR | Medium | The average HPR was 106.5 ± 10.6 mL-H2·L-medium−1·h−1 | HAc and butyric acid (HBc), the main products, can be produced at rates of 0.8–1.6 mmol/L/h and 1.1–2.2 mmol/L/h, respectively. | [84] |
| MBR | Synthetic feed containing sucrose | The highest HPR was 0.95 L-H2 L·−1·d−1 | HAc and HBc are the main products, with the highest concentrations reaching 79 mM and 96 mM, respectively. | [97] |
| UASB | Galactose | When the hydraulic retention time is 2 h, the maximum HPR can reach 56.8 L·L−1·d−1. | HAc and HBc, as the main soluble metabolites, can reach concentrations of up to 1700–2100 mg/L and 3500–3950 mg/L, respectively. | [98] |
| UASB | Palm oil mill effluent | The maximum HPR can achieved 11.75 L·L−1·d−1 | HAc is the main soluble product, with a maximum concentration of 550 mM. | [99] |
| AFBR | vinasse and cheese whey | 2.54 ± 0.39 L H2·d−1·L−1 | HAc and HBc are the main soluble products, with maximum percentages reaching 29.5% and 23.3%, respectively. | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; He, M.; Hou, Y.; Shuaibu, B.A.; Dong, W.; Liu, C.; Yan, B. Driving Sustainable Energy Co-Production: Gas Transfer and Pressure Dynamics Regulating Hydrogen and Carboxylic Acid Generation in Anaerobic Systems. Processes 2025, 13, 2343. https://doi.org/10.3390/pr13082343
Xiao X, He M, Hou Y, Shuaibu BA, Dong W, Liu C, Yan B. Driving Sustainable Energy Co-Production: Gas Transfer and Pressure Dynamics Regulating Hydrogen and Carboxylic Acid Generation in Anaerobic Systems. Processes. 2025; 13(8):2343. https://doi.org/10.3390/pr13082343
Chicago/Turabian StyleXiao, Xiao, Meng He, Yanning Hou, Bilal Abdullahi Shuaibu, Wenjian Dong, Chao Liu, and Binghua Yan. 2025. "Driving Sustainable Energy Co-Production: Gas Transfer and Pressure Dynamics Regulating Hydrogen and Carboxylic Acid Generation in Anaerobic Systems" Processes 13, no. 8: 2343. https://doi.org/10.3390/pr13082343
APA StyleXiao, X., He, M., Hou, Y., Shuaibu, B. A., Dong, W., Liu, C., & Yan, B. (2025). Driving Sustainable Energy Co-Production: Gas Transfer and Pressure Dynamics Regulating Hydrogen and Carboxylic Acid Generation in Anaerobic Systems. Processes, 13(8), 2343. https://doi.org/10.3390/pr13082343