Abstract
Postbiotics, defined as non-viable microbial cells or their components that confer health benefits to the host, have emerged as promising functional ingredients for safe and shelf-stable products. This study aimed to develop postbiotic acerola juice through fermentation with Lacticaseibacillus casei NRRL B-442, followed by thermosonication (TS; 3300 W/L, 60–65 °C, 10 min) or thermal treatment (TH; 80 °C, 10 min). Both TS and TH effectively inactivated the probiotic strain while preserving key bioactive compounds. Total phenolic content ranged from 1.22 to 1.46 g/L, and ascorbic acid levels were retained between 2.36 ± 0.1 and 2.78 ± 0.1 g/L. Antioxidant activity remained stable, with ABTS values between 16,516 ± 246 and 16,689 ± 249 μmol Trolox/L, and FRAP values ranging from 2860 ± 130 to 2910 ± 160 μg Fe2+/mL. These parameters showed no significant degradation (p > 0.05) over 35 days of cold storage (4 °C). TS at 60 °C enhanced antibacterial activity in dialyzed samples, achieving 65% inhibition against Escherichia coli and 56% against Salmonella Typhimurium (7 log CFU/mL), outperforming TH at 80 °C (51% and 57%, respectively). The antibacterial effect remained stable during refrigerated storage, confirming thermosonication as a viable strategy for producing functional, microbiologically safe postbiotic beverages.
1. Introduction
The effectiveness of probiotic food products relies on the number of viable and active cells per gram or milliliter at the time of consumption [1]. On the other hand, products containing inanimate microorganisms are not subject to this limitation [2,3]. In this context, the term “postbiotic” has emerged, referring to a preparation of inanimate microorganisms and/or their components that confer a health benefit on the host [4]. Beyond inactivated cells, the term ‘postbiotics’ also encompasses soluble components, such as vitamins, organic acids, enzymes, short-chain fatty acids (SCFAs), peptides, cell surface proteins, among others [5,6]. Interest in postbiotics has grown due to their wide range of functional properties, including antimicrobial, antioxidant, and immunomodulatory activity [2]. Although probiotics might be consumed by most of the population, postbiotics are particularly promising for immunocompromised individuals as they attenuate the risk of infection associated with the consumption of live microorganisms [5,7]. Studies demonstrate that postbiotics derived from lactic acid bacteria possess inhibitory properties against various microorganisms, including Listeria monocytogenes, Staphylococcus aureus, Escherichia coli, Salmonella spp., Yersinia spp., Aeromonas spp., Bacillus spp., as well as viruses, yeast, and molds [8,9,10,11].
Postbiotics can arise from microbial cell structures or metabolites produced during microbial fermentation in nutrient-rich environments. As highlighted by Aguilar-Toalá et al. [3], a variety of processing techniques are used to obtain postbiotic preparations, especially those aimed at disrupting microbial cells to release intracellular components. These techniques include thermal and high-pressure treatments, enzymatic hydrolysis, co-cultivation, solvent extraction, chemical inactivation, and sonication. The inactivation method is decisive as it directly influences the composition, integrity, and functionality of the resulting postbiotic compounds, ultimately determining their health-promoting potential. Emerging technologies are being explored to optimize safety and functionality in postbiotic foods. Ultrasound emerges as an eco-friendly and fast-processing method that helps to preserve nutrients and bioactive compounds while extending shelf life [12,13]. However, ultrasound alone is insufficient for effective microbial inactivation. Therefore, combining ultrasound with a thermal treatment, a process known as thermosonication, has proven to be an effective approach for enhancing the efficiency of this technology in microbial inactivation, with reduced impact on the overall food quality [14,15]. This technique has shown promise for producing postbiotic formulations with enhanced antioxidant and antibacterial potential.
The metabolic activity of LAB strains and their ability to produce beneficial metabolites depend significantly on the food matrix, fermentation conditions, and storage parameters, resulting in distinct postbiotic profiles [16]. Lacto-fermentation of fruit juices has been explored as a sustainable strategy for producing natural bio-preservatives [17,18,19,20,21]. LAB can produce antimicrobial and antioxidant compounds, such as hydrogen peroxide, diacetyl, bacteriocins, peptides, and phenolics during juice fermentation [8].
Among fruit juices, acerola (Malpighia emarginata) stands out due to its exceptionally high vitamin C and phenolic compound content, making it an attractive candidate for functional beverage development. Despite these benefits, its low pH and limited sugar content pose challenges for probiotic fermentation.
Given this context, the present study aimed to: (1) develop and characterize a fermented acerola juice containing potential postbiotics from L. casei B-442 inactivated by thermosonication or thermal processing; (2) evaluate changes in bioactive compounds and antibacterial activity; and (3) assess the stability and functionality of the juices during refrigerated storage (4 °C) for 35 days.
2. Materials and Methods
2.1. Juice Preparation
A pasteurized acerola frozen pulp was diluted in potable water (100 g of acerola pulp to 200 mL of water). The acerola pulp was purchased from the local market without preservatives (Nossa Fruta Brasil®, Fortaleza, Brazil).
2.2. Lacticaseibacillus casei Inoculum Preparation and Enumeration
Lacticaseibacillus casei (former Lactobacillus casei) NRRL B-442 was obtained from the NRRL Culture Collection (United States Department of Agriculture, Peoria, Illinois, USA. The stock culture of L. casei NRRL B-442 was activated in MRS broth (30 °C, 16 h) to obtain approximately 9.0 log CFU/mL of initial cell concentration. The probiotic viability (log CFU/mL) was assessed using the micro drop plating technique [22]. A total of 10 μL of each serial dilution was spot-plated onto MRS agar plates, with each divided into four quadrants to accommodate four different dilutions. All plates were prepared in triplicate, and only quadrants containing 3–30 colonies were enumerated to ensure statistical reliability.
2.3. Acerola Juice Fermentation
Due to the acerola juice’s low pH and sugar content, 25 g/L of glucose was supplemented to the acerola juice, and the pH was adjusted to 5.5. The fermentation was carried out in sterile Schott flasks (500 mL) containing 400 mL of juice with 1% (v/v) of the activated L. casei culture (described in Section 2.1) at 30 °C for 8 h without agitation. After fermentation, citric acid was added to restore the pH to its initial value of 3.5.
2.4. Thermosonication (TS)
Fermented acerola juices were subjected to ultrasound processing using a probe-type ultrasound device (Unique model USD500, 18 kH, Ultronique, São Paulo, Brazil) equipped with a 13 mm titanium macroprobe immersed 1.5 cm below the liquid surface. The juices (100 mL) were processed in a 250 mL glass-jacketed reactor. Thermosonication (TS) was applied at a power intensity of 3300 W. L−1 for 10 min at temperatures of 50 ± 2 °C, 60 ± 2 °C, and 65 ± 2 °C (Table 1). An external circulating water bath was used for temperature control. The TS processing started after the juice reached the desired temperature. After treatment, the juice was immediately cooled in an ice bath, as described by Nascimento et al. [23].

Table 1.
Thermosonication and heat-processed potentially postbiotic acerola juice samples.
2.5. Thermal Processing (TH)
Thermal treatment was performed in a water bath (SOLAB, São Paulo, Brazil) at a controlled temperature of 80 ± 2 °C for 10 min. Time was set after the juice reached the desired temperature. Afterward, the juice was cooled down in an ice bath. The working volume was 100 mL of fermented acerola juice [24]. Table 1 presents the six products evaluated in the present study.
2.6. Storage Stability
After fermentation and subsequent TS and TH processing, the following parameters were assessed during storage at 4 °C for 35 days. Throughout the storage period, samples were periodically collected from each juice to monitor changes in L. casei viability, microbial counts, lactic acid production, ascorbic acid content, glucose consumption, antioxidant activity, and total phenolic compounds.
2.7. Determination of Sugars and Organic Acids
An Agilent HPLC system equipped with a UV-DAD detector at 210 nm and a RID detector was used to evaluate sugars and organic acids. Organic acids (lactic and ascorbic) were separated using an Aminex BIORAD HPX-87H column (300 × 7.8 mm, Bio-Rad Laboratories, Hercules, CA, USA) at 50 °C. Sulfuric acid (5 mM) at 0.4 mL/min was used as the mobile phase. Each run lasted 40 min. Glucose was separated using a Supelcogel™ Ca column (300 × 7.8 mm, Merck Supelco, Darmstadt, Germany) at 80 °C. Ultrapure water was the mobile phase at 0.5 mL/min. The sugars were detected by a refractive index detector (RI) at 35 °C. Each run lasted 30 min. HPLC methodologies were used according to Nascimento et al. [23].
2.8. Total Phenolic Compounds
Total phenolic content was determined using the Folin–Ciocalteu method as determined by Oliveira et al. [25]. A 10 μL aliquot of each sample was mixed with 200 μL of Folin–Ciocalteu reagent (diluted 1:10 in distilled water). After 3 min of incubation in the dark at 25 °C, 100 μL of a 4% Na2CO3 solution was added. The mixture was incubated, and absorbance was measured at 765 nm. Quantification was performed using a gallic acid (GA) calibration curve, and results were expressed as grams of gallic acid equivalents (GAE) per liter (g GAE/L).
2.9. Antioxidant Activity
In vitro antioxidant activity was assessed according to Fonteles et al. [26], using the 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) and ferric reducing antioxidant power (FRAP) assays, representing different antioxidant compounds’ actions. For the ABTS+ assay, the stock solutions were as follows: 7 mM ABTS+(solution A); and 140 mM potassium persulfate solution (solution B). The working solution was prepared by mixing 5000 μL solution A and 88 μL solution B and allowing them to react for 16 h at room temperature (28 °C) in the dark. The solution was diluted by mixing the ABTS+ solution with absolute ethanol to obtain an absorbance of 0.700 ± 0.02 at 734 nm. Diluted juices (30 μL) were allowed to react with 3000 μL ABTS+ solution for 6 min in the dark. Ethanol was used as a blank. Absorbances were measured at 734 nm. Trolox 2mM was the antioxidant standard; the results were expressed in the moles of Trolox/L.
For FRAP assay, stock solutions included 300 mM acetate buffer (3.1 g C2H3NaO2·3H2O and 16 mL C2H4O2), pH 3.6, 10 mM TPTZ (2,4,6-tripyridyl-s-triazine) solution in 40 mM HCl, and 20 mM FeCl3·6H2O solution. The working solution was freshly prepared by mixing µL acetate buffer, 250 µL TPTZ, and 250 µL FeCl3·6H2O; the solution was then warmed to 37 °C before the analysis. The sample (15 μL) was reacted with 285 μL FRAP solution for 30 min in the dark condition. Measurements of the colored product (ferrous tripyridyltriazine complex) were taken at 593 nm. A standard vitamin C 1 M curve was prepared, and the results were expressed in μg Fe2+/mL.
2.10. Microbiological Quality
Total yeast and mold enumeration was completed by spread plating 100 μL of sample dilution in potato-agar medium. All plates were prepared in triplicate, and only plates containing 30–300 colonies were enumerated to ensure statistical reliability. Average colony counts (±standard deviation) were calculated, and the results were expressed in log of colony-forming units per milliliter of juice (log CFU/mL). Yeasts and molds were specifically monitored in this study because they represent the main spoilage microorganisms in acidic fruit juices such as acerola, in which the low pH environment favors their growth [27].
2.11. Antibacterial Activity Assay
Fermented and processed acerola juice samples were centrifuged at 10,000 g for 20 min at 4 °C. The supernatants were collected and dialyzed for 24 h using 12 kDa molecular weight cutoff membranes in distilled water to remove lactic acid. After dialysis, the samples were adjusted to 25 g/L glucose and pH 6.0.
Escherichia coli ATCC 25922 and Salmonella Typhimurium ATCC 14028 were grown in nutrient broth (Merck Millipore, Darmstadt, Germany) at 30 °C and 125 rpm for 16 h (overnight). The bacterial suspensions were serially diluted and adjusted to concentrations of 4, 5, and 7 log CFU/mL according to their optical density (OD) at 600 nm. Plate counts on nutrient agar (microdrop inoculation method) confirmed the counts.
A microtiter plate assay was used to assess antibacterial activity (growth inhibition, %). In 96-well plates, 100 μL of each juice sample (centrifuged and/or dialyzed) was mixed with 100 μL of bacterial suspension (4, 5, and 7 log CFU/mL) and incubated at 30 °C for 24 h.
Absorbance at 600 nm was measured at 0 and 24 h using a microplate reader (Bio-Tek Instruments, Winooski, VT, USA). Antibacterial activity was calculated using the following equation:
where:
ΔABS sample = ABS 24 h sample − ABS 0 h sample
- -
- 24 h sample: 100 μL of probiotic or postbiotic juice + 100 μL of bacterial suspension (E. coli or S. typhimurium), incubated for 24 h at 30 °C;
- -
- 0 h sample: 100 μL of probiotic or postbiotic juice + 100 μL of bacterial suspension (E. coli or S. typhimurium), measured immediately (0 h) at 30 °C;
ΔABS negative control = ABS 24 negative control − ABS 0 h negative control;
- -
- 24 h negative control: 100 μL of nutrient broth + 100 μL of bacterial suspension (E. coli or S. typhimurium), incubated for 24 h at 30 °C;
- -
- 0 h negative control: 100 μL of nutrient broth + 100 μL of bacterial suspension (E. coli or S. typhimurium), measured immediately (0 h) at 30 °C.
A positive control composed of 100 μL of lactic acid solution (3 g/L) + 100 μL of bacterial suspension (E. coli or S. typhimurium) was also evaluated.
2.12. Statistics
The assays were carried out in replicates, and the analysis in triplicate (n = 6). The Tukey test at 95% confidence level was applied to the pertinent data. The data was handled using Statistica software (TIBCO, StatSoft 14.0.0.15).
3. Results and Discussion
3.1. Acerola Juice Fermentation by L. casei
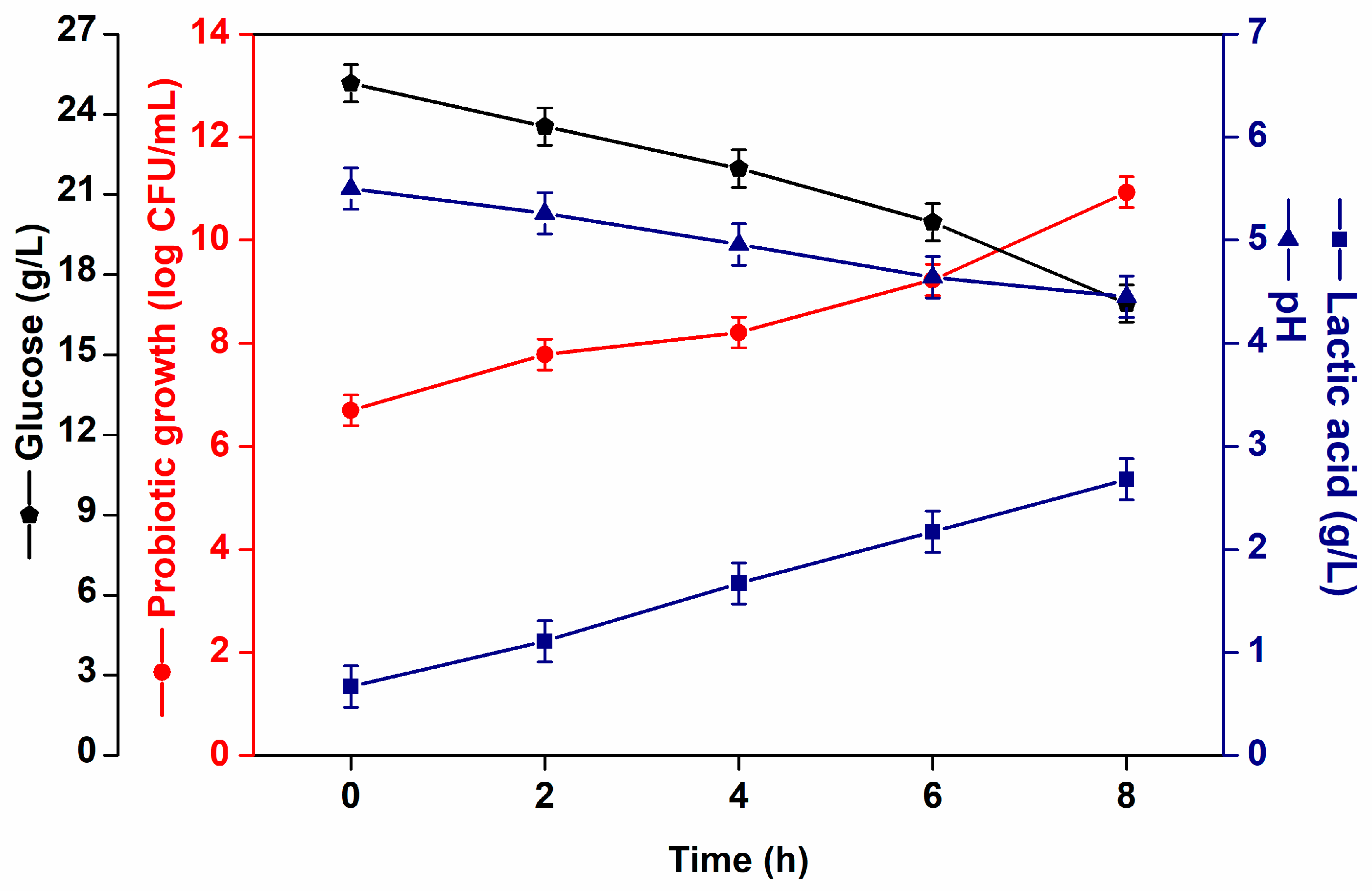
L. casei NRRL B-442 effectively fermented acerola juice, exhibiting a high growth rate by utilizing added glucose as a carbon source. The viable cell counts of L. casei in the fermented acerola juices are shown in Figure 1. After 8h of fermentation, the cell viability was 10.9 log CFU/mL, starting from an initial count of 6.70 log CFU/mL, and consuming approximately 32.4% of the initial glucose. These values are above the minimum threshold of 6–7 log CFU/mL typically required for probiotic functionality and health claims [28,29]. After 8 h of fermentation, L. casei produced 2.7 ± 0.1 g/L of lactic acid, resulting in a pH drop from 5.5 to 4.5.
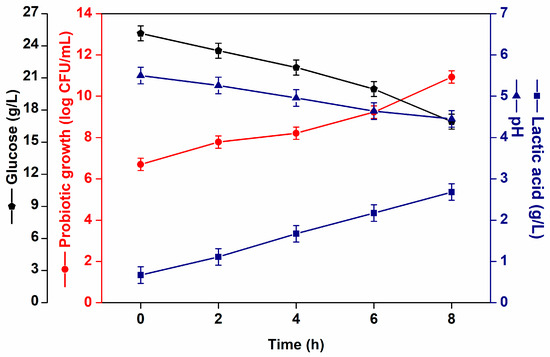
Figure 1.
L. casei NRRL B-442 growth (log CFU/mL) ( ) glucose consumption (g/L) (
) glucose consumption (g/L) ( ), pH (
), pH ( ) and lactic acid (g/L) (
) and lactic acid (g/L) ( ) formation during fermentation.
) formation during fermentation.
) glucose consumption (g/L) (), pH () and lactic acid (g/L) () formation during fermentation.
In a prior study conducted by our research group, acerola juice was effectively fermented by L. casei NRRL B-442, both with and without the addition of prebiotics, resulting in microbial growth and metabolite production [23]. However, the addition of an external carbon source favors microbial growth. In the present study, glucose was added to acerola juice to increase microbial counts.
The present results agree with those reported in similar studies. Santos et al. [21] evaluated the fermentation of cocoa juice by L. casei NRRL B-442 (pH 6.2, 33 °C, and 12 h) and reported a maximum probiotic count of approximately 9 log CFU/mL and lactic acid production of 4.3 g/L after 24h. Probiotic cashew apple juice [18] (pH 6.4, 30 °C and 16 h) showed probiotic count of 8.43 Log CFU/mL and lactic acid production of 2.9 ± 0.01 g/L; probiotic cantaloupe melon juice [19] (pH 6.1, 31 °C and 12 h) showed counts of 8.85 Log CFU/mL and lactic acid production of 3.9 ± 0.01 g/L.
Table 2 shows changes in ascorbic acid, antioxidant activity (ABTS and FRAP), and phenolic compounds concentrations in probiotic acerola juice after 8 h of fermentation. The fermentation process significantly enhanced the bioactive profile of the acerola juice (p < 0.05), leading to increases in total phenolic compounds (1.44 ± 0.05 g/L), ascorbic acid (2.4 ± 0.10 g/L), and antioxidant activity measured by ABTS (19,196 ± 272 μmol Trolox/L) and FRAP (3170 ± 150 μg Fe2+/mL) assays (Table 2). Fermentation-generated microbial metabolites may facilitate the release of cell wall-bound bioactive compounds, such as tannins, flavonoids, and phenolic acids, thereby increasing their bioavailability. Furthermore, the hydrolysis of complex phenolic structures into free forms also contributes to the elevated concentrations of phenolic compounds [30]. These findings highlight the potential of fermentation to enhance the functional properties of fruit juice matrices, aligning with the work of Xu et al. [31], who reported similar improvements in vitamin C, total phenolics, and antioxidant activity in citrus juice fermented with Lactobacillus strains.
Table 2.
Antioxidant activity (ABTS and FRAP), ascorbic acid, and phenolic compounds concentrations after 8 h of fermentation in acerola fermented juice *.
Tran et al. [32] found no statistically significant changes (p ≤ 0.05) in antioxidant activity or total phenolic content following 24 h of grapefruit juice fermentation with L. plantarum 01, L. fermentum D13, L. rhamnosus B01725, and B. bifidum B7.5. Similarly, Dzandu et al. [33] observed no notable improvement (p ≤ 0.05) in antioxidant capacity in tomato juices fermented for 72h with L. rhamnosus ATCC 53103 and L. casei ATCC 393. These findings demonstrate that glucose-supplemented acerola juice is a suitable fermentation matrix for L. casei NRRL B-442.
Probiotic Stability During Cold Storage
Ensuring probiotic viability throughout both production and storage is essential for maintaining health claims from probiotic consumption [21]. However, preserving the stability of live microorganisms presents a significant technological challenge, as many probiotic strains are sensitive to environmental stressors such as oxygen exposure and extreme temperatures.
After fermentation, probiotic acerola juice was stored at 4 °C for 35 days. Figure 2 shows the viable cell counts of L. casei B-442, lactic acid, pH, and glucose concentrations in probiotic acerola juice throughout storage at 4 °C. In probiotic juice, fluctuations in cell viability were observed during the storage period. An increase in viability of approximately 7 log cycles was observed after 21 days of storage. However, cell viability decreased by 8 log cycles between 21 and 35 days of storage. Concurrently, a decrease in glucose concentration of approximately 83% was observed throughout the storage period, confirming the microbial activity. Despite this reduction, which was caused by glucose depletion, the viable cell count remained consistently above 10.00 ± 0.5 log CFU/mL (Figure 2), which exceeds the minimum threshold typically required for probiotic functionality in fermented foods [1,23,29].
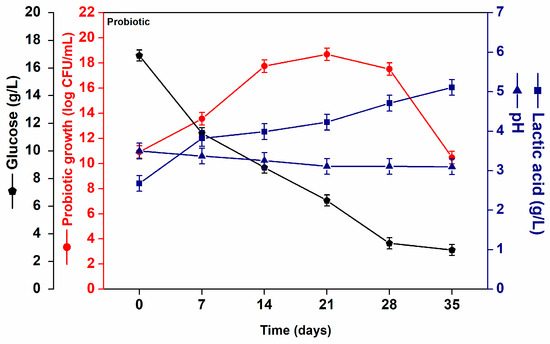
Figure 2.
L. casei NRRL B-442 growth (log CFU/mL) () glucose consumption (g/L) (), pH () and lactic acid (g/L) () formation in probiotic acerola juice throughout storage at 4 °C.
) glucose consumption (g/L) (), pH () and lactic acid (g/L) () formation in probiotic acerola juice throughout storage at 4 °C.
The present findings agree with Santos et al. [21], who reported that the viability of L. casei in sucrose-supplemented probiotic cocoa juice increased until the 21st day of storage at 4 °C. However, a decline in viability was observed after day 28, from 8.99 ± 0.02 to 7.52 ± 0.01 log CFU/mL by day 42. These results highlight the dynamic behavior of LAB viability in acidic, sugar-rich fruit matrices during refrigerated storage (Figure 2).
The viability of probiotic microorganisms in food products is influenced by various factors throughout production, processing, and storage, with lactic acid being a key determinant. Over the 35-day storage period, we observed a 91% increase in lactic acid concentration (from 2.68 ± 0.15 g/L to 5.11 ± 0.25 g/L; p < 0.05) in probiotic acerola juice, while pH values dropped from 3.5 to 3.1 (Figure 2). The increase in lactic acid was expected, given that L. casei maintains its metabolic activity even under refrigerated conditions. These findings are consistent with Maciel et al. [34], who reported increased lactic acid content in sapota-do-solimões functional juice, from 6.0 ± 0.01 g/L to 8.0 ± 0.01 g/L within 15 days of storage.
Table 3 shows changes in ascorbic acid, antioxidant activity (ABTS and FRAP), and phenolic compounds concentrations in probiotic acerola juice during storage at 4 °C. The ascorbic acid concentration remained stable during the 35-day storage period, with values ranging from 2.45 ± 0.15 g/L to 2.55 ± 0.15 g/L (Table 3). Antioxidant activity (FRAP) followed a similar trend, maintaining values between 3150 ± 150 and 3200 ± 150 μg Fe2+/mL (Table 3). The FRAP method assesses antioxidant potential based on electron donation capacity rather than hydrogen atom [35].
Table 3.
Antioxidant activity (ABTS and FRAP), ascorbic acid, and phenolic compounds concentrations at the beginning and end of cold storage in acerola fermented juice *.
These findings suggest a relationship between ascorbic acid and FRAP activity in fermented acerola juice. Supporting this observation, Adebayo-Tayo et al. [36] reported stability in vitamin C content in probiotic mango juice stored at 4 °C, ranging from 41.35 to 48.77 mg/100 mL, after 4 weeks.
In contrast, a significant reduction (p < 0.05) in the phenolic compounds was observed (Table 3), decreasing from 1.40 ± 0.1 to 1.17 ± 0.1, and in the antioxidant activity (ABTS), ranging from 19,290 ± 272 to 16,289 ± 261 μmol Trolox/L. The ABTS method measures the ability of lipophilic and hydrophilic antioxidants to quench a 2,2′-azinobis, 3-ethylbenzthiazoline-6-sulphonic acid radical cation, reflecting the juice’s overall antioxidant potential [37]. The observed trends suggest a relationship between total phenolic content and ABTS antioxidant capacity in fermented acerola juice.
3.2. Postbiotic Production by Thermosonication (TS) and Thermal Processing (TH)
The inactivation of the probiotic microorganism following fermentation is required to confer postbiotic status to the fermented juice. Although ultrasonic processing is generally considered less detrimental to bioactive compounds than conventional thermal treatments, its effectiveness and impact can vary significantly depending on the characteristics of the food matrix and the specific operational parameters applied [38]. Preliminary ultrasound processing at 25 °C resulted in only 10% inactivation of L. casei. Since this study aimed to obtain postbiotic preparations requiring complete microbial inactivation, this condition was considered insufficient, and it was excluded from the final experimental design.
Complete inactivation (≥10.9-log reduction) of L. casei NRRL B-442 was achieved in fermented acerola juice using thermosonication (TS: 3300 W/L, 10 min at 60–65 °C) and thermal treatment (TH: 80 °C, 10 min). In contrast, TS at 50 °C resulted in a maximum reduction of 63% (≤8-log reduction), highlighting the temperature-dependent synergistic effect between ultrasound and heat for effective probiotic inactivation (Table 4). Notably, TS at 60–65 °C demonstrated inactivation efficacy comparable to conventional thermal processing at 80 °C, indicating its potential as an efficient alternative for postbiotic juice production.
Table 4.
L. casei B-442 inactivation (%), yeasts and molds (CFU/mL), ascorbic acid, antioxidant activity (ABTS and FRAP), and phenolic compounds concentrations after 8 h of fermentation and different processing methods in acerola fermented juice *,**.
Alcántara-Zavala et al. [39] reported a 5.85 log CFU/mL reduction of Lactobacillus sp. in pulque, a traditional non-dairy fermented beverage, using thermosonication at 95% amplitude (500 W) for 9 min at 50 °C. Almada et al. [24], observed higher resistance to ultrasound alone (792 W; 20 kHz; room temperature), in which L. casei and L. acidophilus at an initial concentration of 7 log CFU/mL required a minimum of 40 min to achieve >5-log reduction (93%) in phosphate-buffered solution. These findings suggest that the thermal component plays an important role in the efficacy of ultrasound inactivation.
No growth of yeasts and molds was observed in juice samples processed by thermosonication and conventional heat treatment (Table 4). Thermosonication is an effective antifungal method used in liquid food systems [40]. The synergistic effect is particularly useful against heat-resistant fungal spores, which are more susceptible to ultrasonic-induced mechanical stress when exposed to sublethal temperatures [41].
Processing plays a critical role in the production of postbiotics, not only for the inactivation of microbial cells but also for optimizing the release and preservation of bioactive compounds [2]. Techniques such as heat treatment and thermosonication can enhance the availability of key metabolites, including peptides, organic acids, and cell wall components, which contribute to the health-promoting effects of postbiotics. Additionally, processing ensures product stability, safety, and consistency, essential for industrial-scale applications [42,43].
The primary mechanism responsible for microbial inactivation by ultrasound is known as acoustic cavitation. This phenomenon involves the formation, growth, and subsequent implosion of microbubbles within a liquid medium subjected to ultrasonic oscillations. The collapse of these bubbles generates localized high temperatures and pressures, producing intense mechanical forces. Additionally, acoustic streaming—the steady fluid flow resulting from acoustic energy dissipation during cavitation—also contributes to microbial inactivation. Together, these effects can cause rupture of the cell wall, disruption and thinning of the cell membrane, and damage to intracellular components, including DNA, ultimately leading to the loss of microbial viability [12,44].
A secondary mechanism contributing to microbial inactivation during thermosonication is the temperature of the process. Increasing the processing temperature elevates the vapor pressure within the cavitation bubbles, mitigating the intensity of the collapse while simultaneously increasing the number of bubbles formed. [13,15]. In most studies combining ultrasound with heat, processing temperatures are typically mild, ranging from 30 °C to 70 °C [14,45], illustrating the potential of thermosonication to reduce the intensity of conventional thermal treatments while maintaining microbial inactivation efficacy.
Thermosonication (TS: 50–65 °C) and conventional thermal processing (TH: 80 °C) effectively preserved the bioactive compounds in fermented acerola juice, with minimal significant statistical differences observed (p > 0.05). Total phenolic content remained between 1.22 and 1.46 g/L, ascorbic acid retention ranged from 2.36 ± 0.1 to 2.78 ± 0.1 g/L, and antioxidant activity—measured by ABTS and FRAP assays—was maintained at 16,516 ± 246 to 16,689 ± 249 μmol Trolox/L and 2860 ± 130 to 2910 ± 160 μg Fe2+/mL, respectively (Table 4). These findings demonstrate that both treatments preserved the functional compounds of the juice with minimal degradation under the tested conditions.
Both thermosonication (60–65 °C) and thermal processing (80 °C) achieved complete probiotic inactivation (≥10.9-log reduction), enabling the development of a stable, potentially postbiotic beverage while preventing significant degradation and ensuring their continued efficacy [37,46,47].
Postbiotic Stability During Cold Storage
To assess the impact of thermosonication and heat treatments on the microbial stability of postbiotic acerola juices, the viability of L. casei, yeast, and molds was monitored throughout 35 days of refrigerated storage.
No growth of L. casei, yeasts, or molds was observed during the storage period in fermented acerola juice samples treated with thermosonication and conventional thermal processing (Table 5), indicating effective microbial control and prevention of post-processing proliferation. The synergistic action of ultrasound-induced cavitation and mild heat likely caused irreversible structural damage to L. casei cells, thereby ensuring microbial stability and safety throughout the product’s shelf life [46].
Table 5.
L. casei B-442 inactivation (%), yeasts and molds (CFU/mL), glucose (g/L), and lactic acid (g/L) during cold storage in postbiotic acerola fermented juice *.
The microbial quality observed (non-contamination by yeasts and molds) may be attributed to the presence of antimicrobial peptides (AMPs) produced during fermentation and retained after processing [48,49]. These low-molecular-weight protein compounds, synthesized by bacteria such as L. casei, exhibit potent antibacterial activity against a variety of pathogenic and spoilage microorganisms. Their mode of action typically involves targeting the cell membranes of closely related bacterial strains, leading to cell disruption and death [50].
Glucose and lactic acid remained stable during storage as expected, given the low temperature and the absence of viable microorganisms (Table 5). Lactic acid is among the organic acids that are extensively employed for food preservation, capable of inducing either lethal or sublethal harm to microorganisms (antimicrobial) by modifying the permeability of the outer cell membrane [51]. The lactic acid and glucose content is constant in all postbiotic juice samples, suggesting that any minor variations observed are likely attributable to experimental variability rather than the processing conditions themselves.
The elevated levels of phenolic compounds, antioxidant activity, and ascorbic acid in acerola juice reveal its potential as a functional food ingredient, showing its notable nutraceutical and health-promoting properties [52]. Table 6 presents the variations in phenolic compounds, ascorbic acid, and antioxidant activity (ABTS and FRAP) in postbiotic acerola juice during refrigerated storage.
Table 6.
Phenolic compounds, ascorbic acid, and antioxidant activity (ABTS and FRAP) during cold storage in postbiotic acerola fermented juice *.
Both ultrasonically and thermally treated juices maintained stable total phenolic content over the 35-day storage period (Table 6), starting at 1.32 ± 0.05 g/L on day 0 and 1.48 ± 0.05 g/L on day 35. Similar effects of thermosonication treatments on total phenolic compounds were reported in blueberry wine [53] and blueberry juice [37]. Ultrasound and heat treatments can lead to chemical reactions, such as releasing phenolic compounds from their connections with various components of cell walls, such as polysaccharides or proteins, which are then released over time [54,55].
Ascorbic acid content remained stable during the 35-day storage period (Table 6). The final concentrations ranged from 2.62 ± 0.15 to 2.85 ± 0.15 g/L for TS60 °C, 2.63 ± 0.15 to 2.89 ± 0.15 g/L for TS65 °C, and from 2.36 ± 0.15 to 2.59 ± 0.15 g/L for TH80 °C. This observation is consistent with Aguilar et al. [56], who demonstrated that combining ultrasound with mild heat (55 °C) can ensure microbiological safety while preserving ascorbic acid content.
Consistent with the results for bioactive compounds, the antioxidant activity remained stable during storage in all postbiotic samples (Table 6), starting at 13,516 ± 1261 to 17,976 ± 150 μmol Trolox/L (ABTS) and 2850 ± 80 to 3200 ± 80 μg Fe2+/mL (FRAP). The antioxidant activity was preserved throughout cold storage, as shown by the stable levels maintained over this period. This stability can be attributed to the retention of phenolic compounds and ascorbic acid.
These findings demonstrate that postbiotic acerola juice produced via thermosonication (60–65 °C) or thermal processing (80 °C) achieves both microbial stability and bioactive preservation.
3.3. Antibacterial Activity
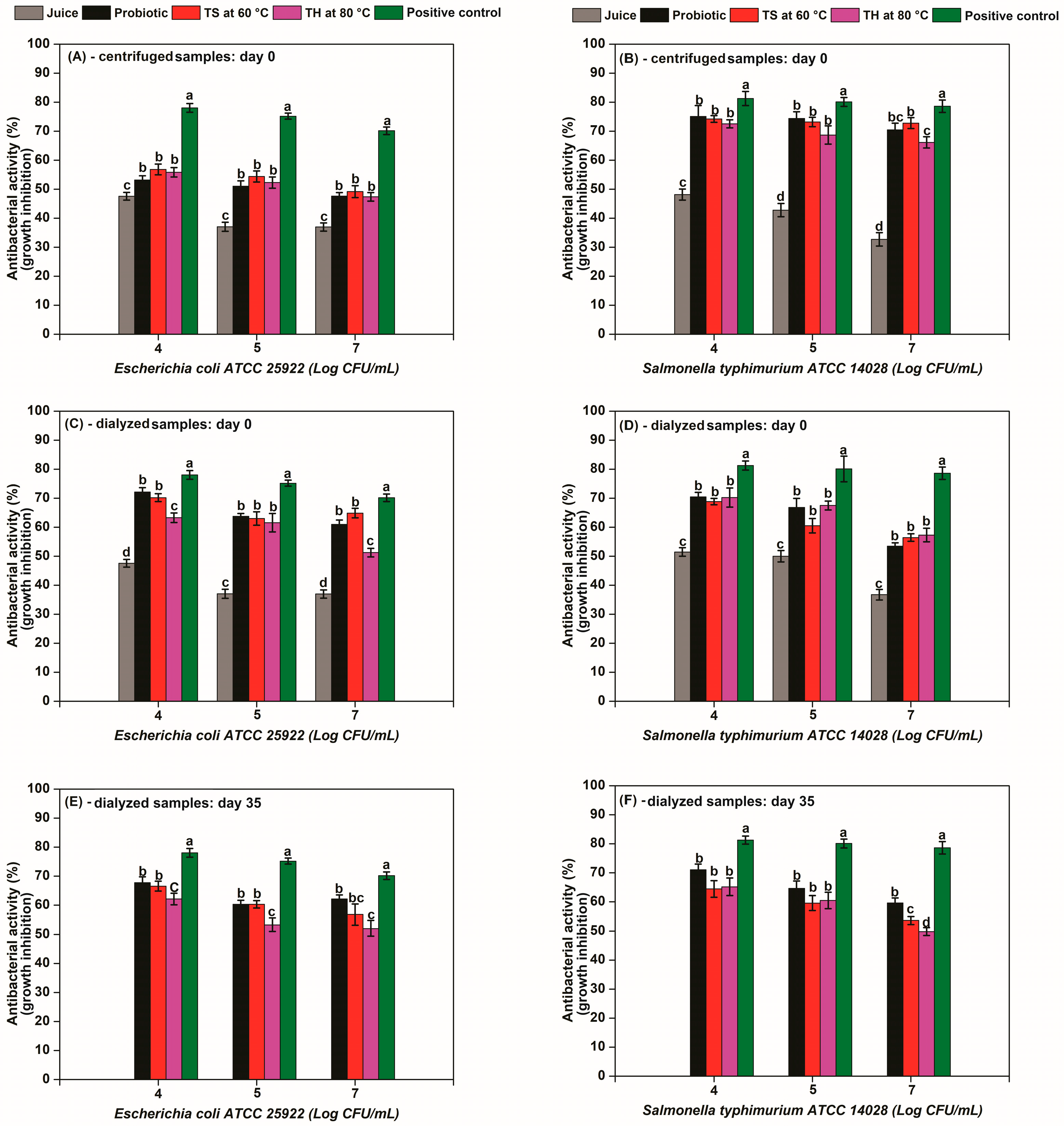
Lactic acid bacteria and their metabolites play a significant role in ensuring food safety and extending shelf-life through the inhibition of pathogenic and spoilage microorganisms [9,57,58]. Figure 3 shows the antibacterial activity (growth inhibition, %) of potentially postbiotic acerola juices subjected to thermosonication (3300 W L−1, 60 °C) and heat processing (80 °C) before and after storage at 4 °C. Since lactic acid has antimicrobial activity, a positive control sample containing 3 g/L of lactic was also evaluated. The antimicrobial activity of acerola juice, probiotic acerola juice, and postbiotic acerola juice and the positive control sample are presented in Figure 3.
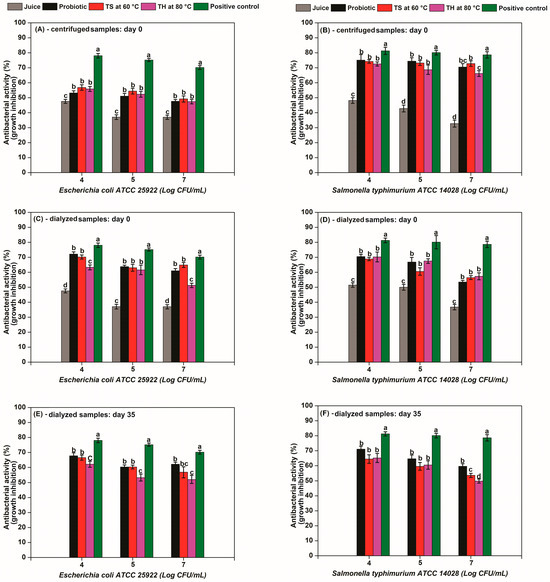
Figure 3.
Antibacterial activity (growth inhibition, %) of potentially postbiotic acerola juices subjected to thermosonication (3300 W L−1 at 60 °C) and heat processing (80 °C) before and after storage at 4 °C. Antimicrobial activity of centrifuged samples against (A) E. coli and (B) S. typhimurium at day 0; dialyzed samples against (C) E. coli and (D) S. Typhimurium at day 0; dialyzed samples against (E) E. coli and (F) S. Typhimurium after 35 days at 4 °C. Positive control is a solution of lactic acid 3 g/L.
Centrifuged jui ces samples (10,000 g for 20 min at 4 °C) showed strong growth inhibition against E. coli and S. typhimurium (antibacterial activity, %) (Figure 3A,B). Acerola juice inhibited E. coli growth by 24% and S. typhimurium growth by 32% (at 7 log CFU/mL). This reduction in growth is likely due to the bioactive compounds found in acerola juice.
Probiotic acerola juice enhanced growth inhibition, reaching 48% against E. coli and 70% against S. typhimurium (7 log CFU/mL) (Figure 3A,B). This indicates that the fermentation process produced metabolites with antibacterial properties. Antimicrobial agents can include bacteriocins and organic acids (lactic acid, acetic acid, formic acid, caproic acid), short-chain fatty acids (such as propionic acid, butyric acid, and isobutyric acid), carbon dioxide, hydrogen peroxide, reuterin, reuterocycline, diacetyl [10,59]. In this work, the antibacterial properties were likely attributed to antimicrobial peptides or lactic acid, since L. casei is a homofermentative bacteria.
The postbiotic juices retained antimicrobial activity at levels comparable to the probiotic juice, indicating that the processing method preserved the antibacterial compounds. TS60 exhibited 49% and 73% inhibition against E. coli and S. typhimurium (7 log CFU/mL), respectively, while TH80 showed 47% and 66% inhibition against the same pathogens at the same concentration (Figure 3A,B). The centrifuged samples demonstrated stronger antimicrobial activity against S. typhimurium compared to E. coli, suggesting greater efficacy of the inhibitory compounds against Salmonella species.
Fruit and fruit juices have traditionally been regarded as low-risk foods for microbial contamination due to their low pH levels. However, they are increasingly recognized as potential sources of foodborne illnesses caused by pathogens (e.g., Salmonella and Escherichia coli) [60].
These findings are consistent with previous research, demonstrating that fermentation enhances the antimicrobial activity of probiotic-containing products. Muhialdin et al. [8] reported that a 48 h fermentation significantly increased the antibacterial activity of fermented cantaloupe juice using L. plantarum FBS05 towards 106 CFU/mL of E. coli and S. typhimurium to 96.73 ± 0.00% and 96.83 ± 0.00%, respectively. Algboory et al. [9] reported that a 48 h fermentation enhanced the antibacterial activity of fermented camel milk using L. plantarum IS10 towards 106 CFU/mL of E. coli and S. aureus subsp. aureus to 75% and 79%, respectively.
To ensure that the antibacterial effect of the juices did not come only from the lactic acid produced during fermentation, dialysis was performed to remove lactic acid from centrifuged samples juices, as confirmed by HPLC analysis, thereby allowing for the evaluation of antibacterial effects arising from other bioactive constituents. As shown in Figure 3C,D, the dialyzed samples (24 h using 12 kDa molecular weight cutoff membranes in distilled water) effectively maintained the antibacterial activity against E. coli and S. typhimurium. The dialysis process consistently enhanced anti-E. coli activity across all samples (probiotic: +12%; TS60: +16%; TH80: +4%) (Figure 3C), while also reducing anti-Salmonella efficacy (probiotic: −20%; TS60: −17%; TH80: −9%) (Figure 3D) compared to centrifuged samples at 7 log CFU/mL. These findings indicate the presence of additional antimicrobial compounds beyond lactic acid—potentially including bioactive peptides that contribute to the inhibitory effect [9,10,61].
Moreover, the postbiotics produced by thermal processing at 80 °C were less effective against E. coli when compared to the probiotic and postbiotics produced by thermosonication at 60 °C (p < 0.05) (Figure 3C). Majkut et al. [62] reported that honey subjected to thermal treatment showed a significant reduction of antimicrobial properties (MBC ⩾ 50%) at 82 °C and 62 °C after 15 and 120 min exposure.
The antimicrobial activity of dialyzed juices is likely due to the presence of antimicrobial peptides, with bacteriocins being among the most well-known [63]. These compounds exert their effects by disrupting the integrity of target bacterial cell membranes, forming pores that lead to cell leakage, loss of membrane potential, and ultimately cell death. In some cases, bacteriocins may also interfere with essential enzymatic systems or inhibit protein synthesis [64].
The antibacterial efficacy of these bacteriocins is particularly valuable in food preservation as they inhibit common foodborne pathogens, such as L. monocytogenes, S. aureus, and certain strains of E. coli and Salmonella. Additionally, because bacteriocins are naturally produced and generally regarded as safe (GRAS), they offer a promising alternative to synthetic preservatives and antibiotics [65].
The stability of juices during cold storage is a critical aspect of guaranteeing the product’s shelf life. In this study, the antimicrobial activity was evaluated after 35 days of cold storage at 4 °C to guarantee the antibacterial effect of the juices after this period.
The antibacterial activity was maintained after 35 days of storage (Figure 3E,F). The probiotic juice maintained stable inhibition rates against both pathogens at 7 log CFU/mL: 62% (vs. 61% in dialyzed samples day 0) for E. coli and 59% (vs. 53% in dialyzed samples day 0) for S. typhimurium, demonstrating consistent efficacy over time (Figure 3E,F). This sustained efficacy may be explained by the continued fermentation activity of L. casei during cold storage, as the strain remains metabolically active at low temperatures, potentially generating additional antimicrobial compounds (e.g., bacteriocins) over 35 days [21,34].
The postbiotic juices (TS60 and TH80) retained significant antibacterial activity after 35 days of storage, demonstrating remarkable stability despite minor reductions in efficacy. TS60 maintained 57% inhibition against E. coli (vs. 65% initially) and 54% against S. typhimurium (vs. 56% initially), representing only 8–12% activity loss. TH80 showed even greater stability against E. coli (53% vs. 51% initially), with a 7% reduction against S. typhimurium (50% vs. 57%) (Figure 3E,F). These results indicate that the antibacterial activity remains effective in juices for up to 35 days with some reductions.
LAB bacteriocins exhibit exceptional thermostability and acid tolerance, making them ideal candidates for food preservation [66]. Notable examples include pentocin TV35b, enterocin A5-11, and bacteriocin EF2019, along with bacteriocins produced by intestinal strains of Pediococcus and Lactococcus spp. These antimicrobial peptides demonstrate remarkable stability under processing conditions while maintaining bioactivity [67].
4. Conclusions
This study demonstrated the feasibility of producing a stable, functional, non-dairy postbiotic beverage from acerola juice fermented with L. casei B-442. Both thermosonication (TS) and thermal processing (TH) effectively inactivated the probiotic strain (≥10.9-log reduction), while preserving and enhancing key bioactive compounds. Juices treated with thermosonication (TS, 60–65 °C) demonstrated superior retention of ascorbic acid, total phenolic content, and antioxidant activity compared to conventional thermal pasteurization. These bioactive parameters remained stable throughout 35 days of refrigerated storage at 4 °C. Notably, postbiotic juices exhibited significant antibacterial activity against E. coli and S. Typhimurium, which remained stable throughout 35 days of cold storage. Among the tested methods, thermosonication at 60 °C emerged as the most promising strategy due to its superior preservation of bioactive compounds, sustained antimicrobial efficacy, and potential for efficient industrial scalability. These findings support the application of thermosonication in developing clean-label, shelf-stable, and health-promoting postbiotic beverages from fruit-based matrices.
Author Contributions
Conceptualization S.R.; methodology, C.S.d.N. and B.N.S.; software, C.S.d.N.; validation, S.R. and T.V.F.; formal analysis, C.S.d.N. and B.N.S.; investigation, C.S.d.N.; resources, S.R.; data curation, C.S.d.N. and T.V.F.; writing—original draft preparation, C.S.d.N.; writing—review and editing, C.S.d.N., T.V.F. and S.R.; visualization, S.R. and T.V.F.; supervision, S.R.; project administration, S.R.; funding acquisition, S.R. All authors have read and agreed to the published version of the manuscript.
Funding
The authors thank the Brazilian funding agency CNPq through the National Institute of Science and Technology of Tropical Fruits for the financial support and FUNCAP for the grants and scholarships. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—finance code 001 Brasil (CAPES).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors thank the ARS Culture Collection (Peoria, IL, USA) for the L. casei strain.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-Parabiotics: The New Horizons in Microbial Biotherapy and Functional Foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An Evolving Term within the Functional Foods Field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Barros, C.P.; Guimarães, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and Postbiotics: Concepts and Potential Applications in Dairy Products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef]
- Yeşilyurt, N.; Yılmaz, B.; Ağagündüz, D.; Capasso, R. Involvement of Probiotics and Postbiotics in the Immune System Modulation. Biologics 2021, 1, 89–110. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Kadum, H.; Meor Hussin, A.S. Metabolomics Profiling of Fermented Cantaloupe Juice and the Potential Application to Extend the Shelf Life of Fresh Cantaloupe Juice for Six Months at 8 °C. Food Control 2021, 120, 107555. [Google Scholar] [CrossRef]
- Algboory, H.L.; Muhialdin, B.J. Novel Peptides Contribute to the Antimicrobial Activity of Camel Milk Fermented with Lactobacillus plantarum IS10. Food Control 2021, 126, 108057. [Google Scholar] [CrossRef]
- İncili, G.K.; Karatepe, P.; Akgöl, M.; Güngören, A.; Koluman, A.; İlhak, O.İ.; Kanmaz, H.; Kaya, B.; Hayaloğlu, A.A. Characterization of Lactic Acid Bacteria Postbiotics, Evaluation in-Vitro Antibacterial Effect, Microbial and Chemical Quality on Chicken Drumsticks. Food Microbiol. 2022, 104, 104001. [Google Scholar] [CrossRef]
- Shirkhan, F.; Mirdamadi, S.; Mirzaei, M.; Akbari-adergani, B.; Nasoohi, N. The Role of Lactic Acid Bacteria in Production of Bioactive Peptides in Fermented Milk with Antioxidant and Antidiabetic Properties. J. Food Meas. Charact. 2023, 17, 4727–4738. [Google Scholar] [CrossRef]
- Nunes, B.V.; da Silva, C.N.; Bastos, S.C.; de Souza, V.R. Microbiological Inactivation by Ultrasound in Liquid Products. Food Bioproc. Tech. 2022, 15, 2185–2209. [Google Scholar] [CrossRef]
- Oliveira, G.A.R.; Guimarães, J.T.; Ramos, G.L.P.A.; Esmerino, E.A.; Pimentel, T.C.; Neto, R.P.C.; Tavares, M.I.B.; Sobral, L.A.; Souto, F.; Freitas, M.Q.; et al. Benefits of Thermosonication in Orange Juice Whey Drink Processing. Innov. Food Sci. Emerg. Technol. 2022, 75, 102876. [Google Scholar] [CrossRef]
- Fan, L.; Ismail, B.B.; Gao, L.; Liu, D. Comparison of High- and Low- Frequency Thermosonication and Carvacrol Treatments of Carrot Juice: Microbial Inactivation and Quality Retention. Appl. Food Res. 2022, 2, 100162. [Google Scholar] [CrossRef]
- Jafarpour, D. The Effect of Heat Treatment and Thermosonication on the Microbial and Quality Properties of Green Olive. J. Food Meas. Charact. 2022, 16, 2172–2180. [Google Scholar] [CrossRef]
- Rodrigues, V.C.d.C.; da Silva, L.G.S.; Simabuco, F.M.; Venema, K.; Antunes, A.E.C. Survival, Metabolic Status and Cellular Morphology of Probiotics in Dairy Products and Dietary Supplement after Simulated Digestion. J. Funct. Foods 2019, 55, 126–134. [Google Scholar] [CrossRef]
- Pereira, A.L.F.; Feitosa, W.S.C.; Abreu, V.K.G.; Lemos, T.d.O.; Gomes, W.F.; Narain, N.; Rodrigues, S. Impact of Fermentation Conditions on the Quality and Sensory Properties of a Probiotic Cupuassu (Theobroma grandiflorum) Beverage. Food Res. Int. 2017, 100, 603–611. [Google Scholar] [CrossRef]
- Pereira, A.L.F.; Maciel, T.C.; Rodrigues, S. Probiotic Beverage from Cashew Apple Juice Fermented with Lactobacillus casei. Food Res. Int. 2011, 44, 1276–1283. [Google Scholar] [CrossRef]
- Fonteles, T.V.; Costa, M.G.M.; de Jesus, A.L.T.; Fontes, C.P.M.L.; Fernandes, F.A.N.; Rodrigues, S. Stability and Quality Parameters of Probiotic Cantaloupe Melon Juice Produced with Sonicated Juice. Food Bioproc Tech. 2013, 6, 2860–2869. [Google Scholar] [CrossRef]
- de Godoy Alves Filho, E.; Rodrigues, T.H.S.; Fernandes, F.A.N.; Pereira, A.L.F.; Narain, N.; de Brito, E.S.; Rodrigues, S. Chemometric Evaluation of the Volatile Profile of Probiotic Melon and Probiotic Cashew Juice. Food Res. Int. 2017, 99, 461–468. [Google Scholar] [CrossRef]
- dos Santos Filho, A.L.; Freitas, H.V.; Rodrigues, S.; Abreu, V.K.G.; Lemos, T.d.O.; Gomes, W.F.; Narain, N.; Pereira, A.L.F. Production and Stability of Probiotic Cocoa Juice with Sucralose as Sugar Substitute during Refrigerated Storage. LWT 2019, 99, 371–378. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to Optimize the Drop Plate Method for Enumerating Bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Silva do Nascimento, C.; Santos, B.N.; Rodrigues, S. High-intensity Ultrasound Processed Acerola Juice Containing Oligosaccharides and Dextran Promotes Lacticaseibacillus casei NRRL B-442 Growth. Int. J. Food Sci. Technol. 2022, 57, 5186–5194. [Google Scholar] [CrossRef]
- Almada, C.N.; Almada-Érix, C.N.; Bonatto, M.S.; Pradella, F.; dos Santos, P.; Abud, Y.K.D.; Farias, A.S.; Martínez, J.; Sant’Anna Filho, C.B.; Lollo, P.C.; et al. Obtaining Paraprobiotics from Lactobacilus acidophilus, Lacticaseibacillus casei and Bifidobacterium animalis Using Six Inactivation Methods: Impacts on the Cultivability, Integrity, Physiology, and Morphology. J. Funct. Foods 2021, 87, 104826. [Google Scholar] [CrossRef]
- Oliveira, A.F.A.; Mar, J.M.; Santos, S.F.; da Silva Júnior, J.L.; Kluczkovski, A.M.; Bakry, A.M.; Bezerra, J.d.A.; Nunomura, R.d.C.S.; Sanches, E.A.; Campelo, P.H. Non-Thermal Combined Treatments in the Processing of Açai (Euterpe oleracea) Juice. Food Chem. 2018, 265, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Fonteles, T.V.; dos Santos, A.Y.S.; Linhares, M.d.F.D.; Miguel, T.B.A.R.; Miguel, E.d.C.; Rodrigues, S. Metabolic Responses of Kombucha Consortium Fermentation upon Ultrasound-Processing. Food Chem. Adv. 2024, 4, 100646. [Google Scholar] [CrossRef]
- Tournas, V.H.; Heeres, J.; Burgess, L. Moulds and Yeasts in Fruit Salads and Fruit Juices. Food Microbiol. 2006, 23, 684–688. [Google Scholar] [CrossRef]
- Fernandes Pereira, A.L.; Rodrigues, S. Turning Fruit Juice Into Probiotic Beverages. In Fruit Juices: Extraction, Composition, Quality and Analysis; Academic Press: San Diego, CA, USA, 2018; pp. 279–287. [Google Scholar] [CrossRef]
- Rasika, D.M.; Vidanarachchi, J.K.; Rocha, R.S.; Balthazar, C.F.; Cruz, A.G.; Sant’Ana, A.S.; Ranadheera, C.S. Plant-Based Milk Substitutes as Emerging Probiotic Carriers. Curr. Opin. Food Sci. 2021, 38, 8–20. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation Transforms the Phenolic Profiles and Bioactivities of Plant-Based Foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Xu, H.; Feng, L.; Deng, Y.; Chen, L.; Li, Y.; Lin, L.; Liang, M.; Jia, X.; Wang, F.; Zhang, X.; et al. Change of Phytochemicals and Bioactive Substances in Lactobacillus Fermented Citrus Juice during the Fermentation Process. LWT 2023, 180, 114715. [Google Scholar] [CrossRef]
- Tran, A.M.; Nguyen, T.B.; Nguyen, V.D.; Bujna, E.; Dam, M.S.; Nguyen, Q.D. Changes in Bitterness, Antioxidant Activity and Total Phenolic Content of Grapefruit Juice Fermented by Lactobacillus and Bifidobacterium Strains. Acta Aliment. 2020, 49, 103–110. [Google Scholar] [CrossRef]
- Dzandu, B.; Chotiko, A.; Sathivel, S. Antioxidant Activity and Viability of Lacticaseibacillus rhamnosus, Lacticaseibacillus casei, and Co-Culture in Fermented Tomato Juice during Refrigerated Storage. Food Biosci. 2022, 50, 102085. [Google Scholar] [CrossRef]
- Maciel da Silva, R.; Henrique Campelo, P.; Rodrigues, S. In Vitro Viability of L. Casei B-442 and Fructooligosaccharides Integrity in Amazonian Sapota-Do-Solimões Functional Juice. Food Res. Int. 2022, 154, 111036. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I.G. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Adebayo-Tayo, B.C.; Olomitutu, F.O.; Adebami, G.E. Production and Evaluation of Probioticated Mango Juice Using Pediococcus pentosaceus and Pediococcus acidilactici during Storage at Different Temperature. J. Agric. Food Res. 2021, 6, 100202. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, L.; Liu, X.; Hasan, K.M.F.; Li, H.; Zhou, S.; Zhang, Q.; Zhou, Y. Effect of Thermosonication Treatment on Blueberry Juice Quality: Total Phenolics, Flavonoids, Anthocyanin, and Antioxidant Activity. LWT 2021, 150, 112021. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Jafarpour, D.; Soto, E.R.; Barba, F.J. Ultrasound-Assisted Lactic Acid Fermentation of Bakraei (Citrus reticulata cv. Bakraei) Juice: Physicochemical and Bioactive Properties. Fermentation 2023, 9, 37. [Google Scholar] [CrossRef]
- Alcántara-Zavala, A.E.; Figueroa-Cárdenas, J.d.D.; Pérez-Robles, J.F.; Arámbula-Villa, G.; Miranda-Castilleja, D.E. Thermosonication as an Alternative Method for Processing, Extending the Shelf Life, and Conserving the Quality of Pulque: A Non-Dairy Mexican Fermented Beverage. Ultrason. Sonochem. 2021, 70, 105290. [Google Scholar] [CrossRef]
- Anaya-Esparza, L.M.; Velázquez-Estrada, R.M.; Roig, A.X.; García-Galindo, H.S.; Sayago-Ayerdi, S.G.; Montalvo-González, E. Thermosonication: An Alternative Processing for Fruit and Vegetable Juices. Trends Food Sci. Technol. 2017, 61, 26–37. [Google Scholar] [CrossRef]
- Evelyn; Milani, E.; Silva, F.V.M. Comparing High Pressure Thermal Processing and Thermosonication with Thermal Processing for the Inactivation of Bacteria, Moulds, and Yeasts Spores in Foods. J. Food Eng. 2017, 214, 90–96. [Google Scholar] [CrossRef]
- Peluzio, M.d.C.G.; Martinez, J.A.; Milagro, F.I. Postbiotics: Metabolites and Mechanisms Involved in Microbiota-Host Interactions. Trends Food Sci. Technol. 2021, 108, 11–26. [Google Scholar] [CrossRef]
- Prajapati, N.; Patel, J.; Singh, S.; Yadav, V.K.; Joshi, C.; Patani, A.; Prajapati, D.; Sahoo, D.K.; Patel, A. Postbiotic Production: Harnessing the Power of Microbial Metabolites for Health Applications. Front. Microbiol. 2023, 14, 1306192. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Jiang, A. Effect of Ultrasound Treatment on Quality and Microbial Load of Carrot Juice. Food Sci. Technol. 2016, 36, 111–115. [Google Scholar] [CrossRef]
- Muñoz, A.; Caminiti, I.M.; Palgan, I.; Pataro, G.; Noci, F.; Morgan, D.J.; Cronin, D.A.; Whyte, P.; Ferrari, G.; Lyng, J.G. Effects on Escherichia Coli Inactivation and Quality Attributes in Apple Juice Treated by Combinations of Pulsed Light and Thermosonication. Food Res. Int. 2012, 45, 299–305. [Google Scholar] [CrossRef]
- Qiu, X.; Su, J.; Nie, J.; Zhang, Z.; Ren, J.; Wang, S.; Pei, Y.; Li, X. Effects of Thermosonication on the Antioxidant Capacity and Physicochemical, Bioactive, Microbiological, and Sensory Qualities of Blackcurrant Juice. Foods 2024, 13, 809. [Google Scholar] [CrossRef]
- Xia, M.; LI, C.; Wu, D.; Wu, F.; Kong, L.; Jia, Z.; Han, W.; Chen, S.; Fang, W.; Liu, Y.; et al. Benefits of Heat-Killed Lactobacillus acidophilus on Growth Performance, Nutrient Digestibility, Antioxidant Status, Immunity, and Cecal Microbiota of Rabbits. Front. Vet. Sci. 2024, 11, 1361908. [Google Scholar] [CrossRef]
- Scudino, H.; Pimentel, T.C.; Guimarães, J.T.; Moura, R.S.; Mársico, E.T.; Esmerino, E.A.; Freitas, M.Q.; Souza, A.A.; Nogueira, F.C.S.; Cruz, A.G. Effect of Thermosonication on the Bioactive Peptide Profile, Volatile Compounds, and Fatty Acid Profile of Minas Frescal Cheese. Int. Dairy. J. 2024, 157, 106009. [Google Scholar] [CrossRef]
- Pavlova, A.S.; Ozhegov, G.D.; Arapidi, G.P.; Butenko, I.O.; Fomin, E.S.; Alemasov, N.A.; Afonnikov, D.A.; Yarullina, D.R.; Ivanov, V.T.; Govorun, V.M.; et al. Identification of Antimicrobial Peptides from Novel Lactobacillus fermentum Strain. Protein J. 2020, 39, 73–84. [Google Scholar] [CrossRef]
- He, J.F.; Jin, D.X.; Luo, X.G.; Zhang, T.C. LHH1, a Novel Antimicrobial Peptide with Anti-Cancer Cell Activity Identified from Lactobacillus casei HZ1. AMB Express 2020, 10, 204. [Google Scholar] [CrossRef]
- Tian, X.; Hu, W.; Chen, J.; Zhang, W.; Li, W. The Supplement of Vitamin C Facilitates L-Lactic Acid Biosynthesis in Lactobacillus thermophilus A69 from Sweet Sorghum Juice Coupled with Soybean Hydrolysate as Feedstocks. Ind. Crops Prod. 2020, 146, 112159. [Google Scholar] [CrossRef]
- Prakash, A.; Baskaran, R. Acerola, an Untapped Functional Superfruit: A Review on Latest Frontiers. J. Food Sci. Technol. 2018, 55, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Peng, Z.; Zhao, Y.; Wu, K.; Zhou, N.; Yan, Y.; Ramaswamy, H.S.; Sun, J.; Bai, W. The Impact of Ultrasonic Treatment on Blueberry Wine Anthocyanin Color and Its In-Vitro Anti-Oxidant Capacity. Food Chem. 2020, 333, 127455. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Sackey, A.S.; Wu, M.; Xiao, L. Impact of Ultrasonication and Pulsed Light Treatments on Phenolics Concentration and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. LWT 2018, 92, 61–66. [Google Scholar] [CrossRef]
- Wang, J.; Vanga, S.K.; Raghavan, V. High-Intensity Ultrasound Processing of Kiwifruit Juice: Effects on the Ascorbic Acid, Total Phenolics, Flavonoids and Antioxidant Capacity. LWT 2019, 107, 299–307. [Google Scholar] [CrossRef]
- Aguilar, K.; Garvín, A.; Ibarz, A.; Augusto, P.E.D. Ascorbic Acid Stability in Fruit Juices during Thermosonication. Ultrason. Sonochem. 2017, 37, 375–381. [Google Scholar] [CrossRef]
- Szutowska, J. Functional Properties of Lactic Acid Bacteria in Fermented Fruit and Vegetable Juices: A Systematic Literature Review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of Culture Media, PH and Temperature on Growth and Bacteriocin Production of Bacteriocinogenic Lactic Acid Bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Qu, X. Genetic Mechanisms of Prebiotic Carbohydrate Metabolism in Lactic Acid Bacteria: Emphasis on Lacticaseibacillus casei and Lacticaseibacillus paracasei as Flexible, Diverse and Outstanding Prebiotic Carbohydrate Starters. Trends Food Sci. Technol. 2021, 115, 486–499. [Google Scholar] [CrossRef]
- Aneja, K.R.; Dhiman, R.; Aggarwal, N.K.; Kumar, V.; Kaur, M. Microbes Associated with Freshly Prepared Juices of Citrus and Carrots. Int. J. Food Sci. 2014, 2014, 408085. [Google Scholar] [CrossRef]
- Li, P.; Xia, J.; Nie, Z.; Shan, Y. Pectic Oligosaccharides Hydrolyzed from Orange Peel by Fungal Multi-Enzyme Complexes and Their Prebiotic and Antibacterial Potentials. LWT-Food Sci. Technol. 2016, 69, 203–210. [Google Scholar] [CrossRef]
- Majkut, M.; Kwiecińska-Piróg, J.; Wszelaczyńska, E.; Pobereżny, J.; Gospodarek-Komkowska, E.; Wojtacki, K.; Barczak, T. Antimicrobial Activity of Heat-Treated Polish Honeys. Food Chem. 2021, 343, 128561. [Google Scholar] [CrossRef] [PubMed]
- Ullah, N.; Wang, X.; Wu, J.; Guo, Y.; Ge, H.; Li, T.; Khan, S.; Li, Z.; Feng, X. Purification and Primary Characterization of a Novel Bacteriocin, LiN333, from Lactobacillus casei, an Isolate from a Chinese Fermented Food. LWT 2017, 84, 867–875. [Google Scholar] [CrossRef]
- Xue, X.; Gao, Y.; Liu, F.; Du, P.; Li, C.; Liu, Y.; Yu, W.; Liu, L. Purification, Characterization, and Identification of a Novel Bacteriocin Produced by Lacticaseibacillus casei KLS1, and Its Antimicrobial Mechanism against Staphylococcus aureus. LWT 2024, 200, 116207. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, Z.; Liang, Z.; Fu, X.; Li, D.; Zhu, C.; Kong, Q.; Mou, H. Current Challenges and Development Strategies of Bacteriocins Produced by Lactic Acid Bacteria Applied in the Food Industry. Compr. Rev. Food Sci. Food Saf. 2025, 24, e70038. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, A.; Asadi, A.; Mahdizade Ari, M.; Ohadi, E.; Talebi, M.; Halaj Zadeh, M.; Darb Emamie, A.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and Potential Use as Antimicrobials. J. Clin. Lab. Anal. 2022, 36, e24093. [Google Scholar] [CrossRef]
- Barbosa, A.A.T.; Mantovani, H.C.; Jain, S. Bacteriocins from Lactic Acid Bacteria and Their Potential in the Preservation of Fruit Products. Crit. Rev. Biotechnol. 2017, 37, 852–864. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).