
Evaluation of the Effect of Betulin and Its Alkynyl Derivatives on the Profile of Changes in Gene Expression of the Inflammatory Process of Colorectal Adenocarcinoma Cells (HT-29 Cell Line)


Abstract
:1. Introduction
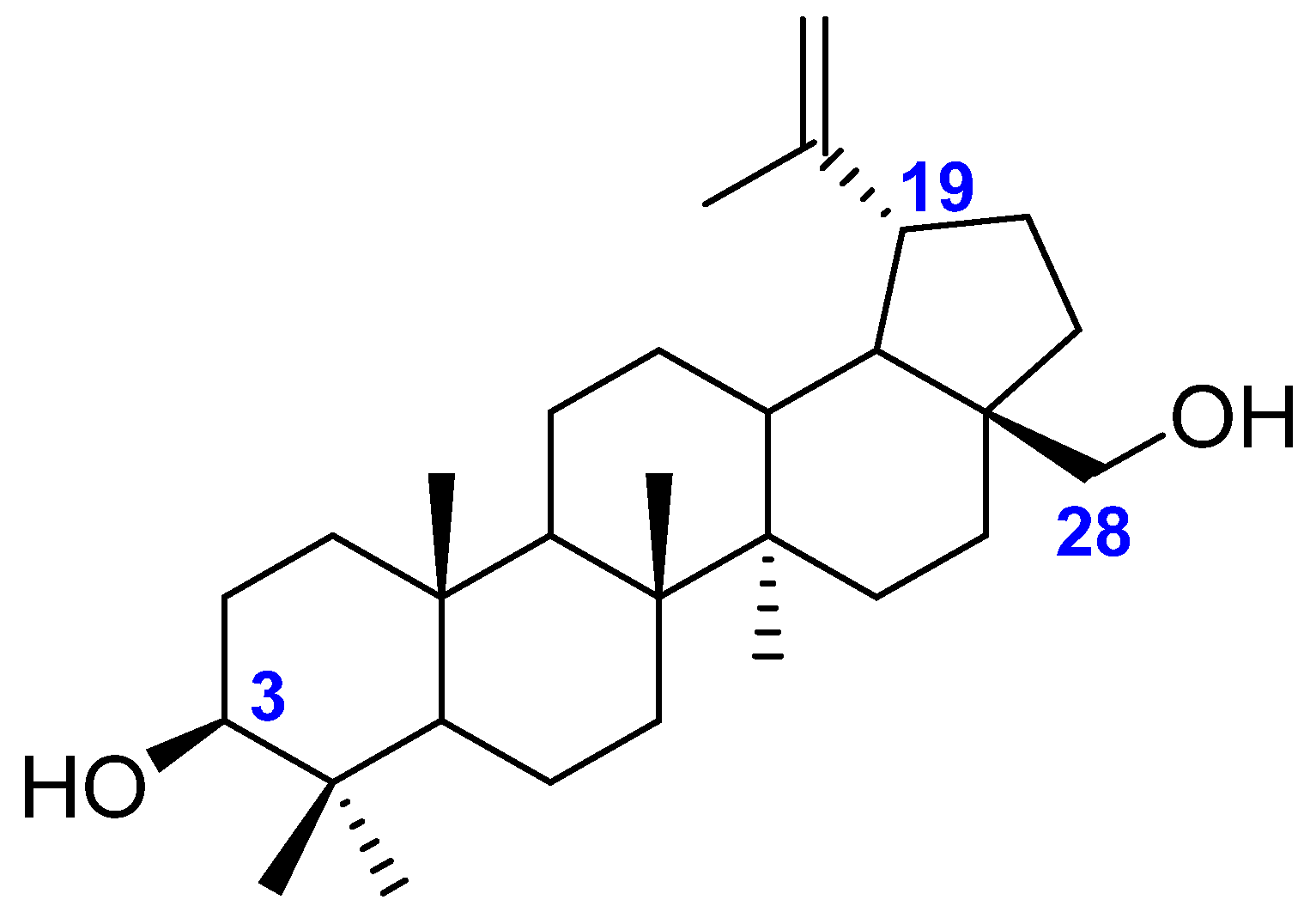
1.1. Betulin
1.2. Antitumor Activity
1.3. Anti-Inflammatory Action
1.4. Lipophilicity
1.5. Colorectal Cancer and Inflammation
2. Materials and Methods
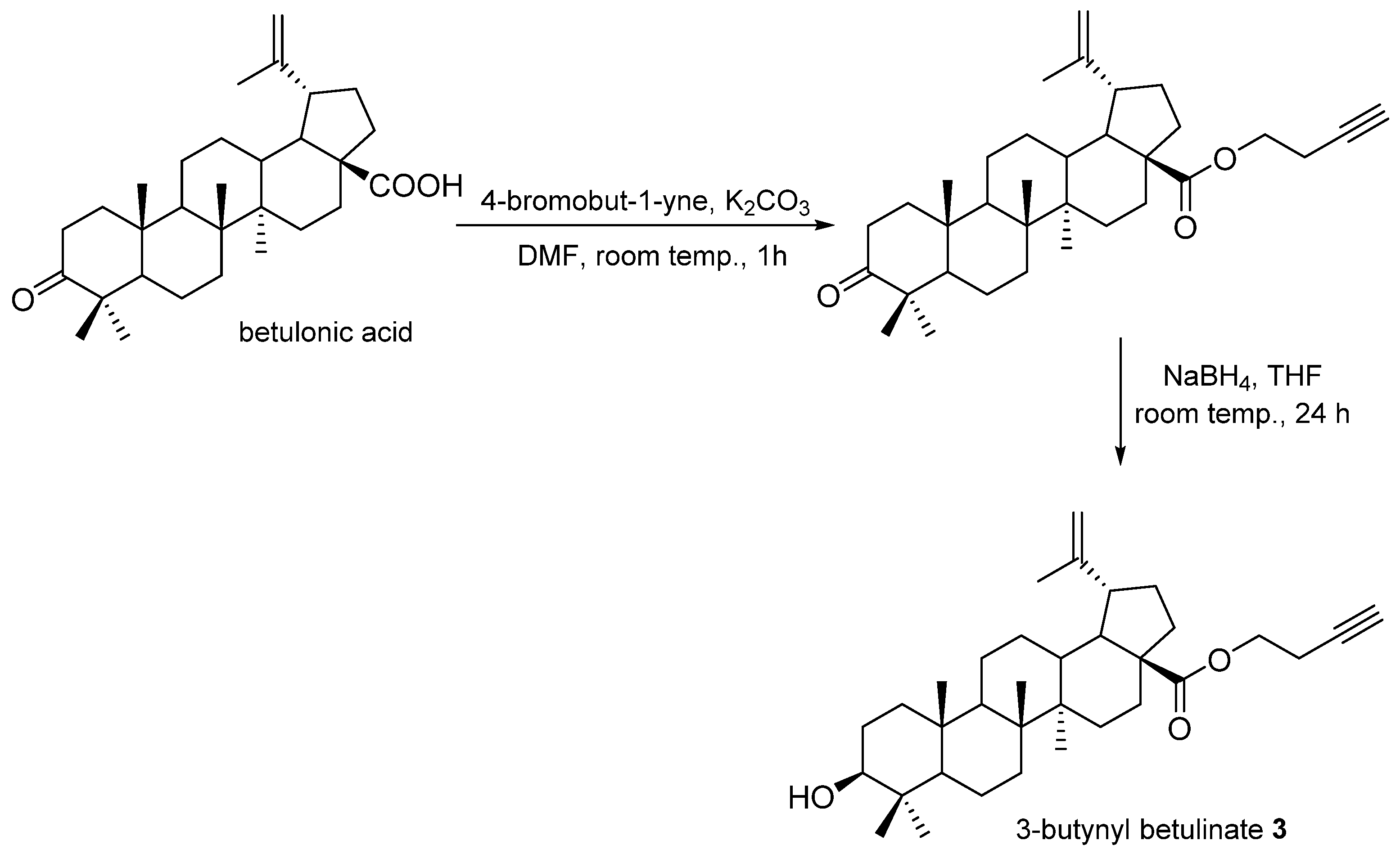
2.1. Compound Synthesis
2.2. Cell Lines
2.3. Cytotoxicity
2.4. Lipophilicity
2.5. Transcriptome Determination Using Oligonucleotide Microarray Technology
2.6. Statistical analysis
3. Results
3.1. Cytotoxicity Evaluation
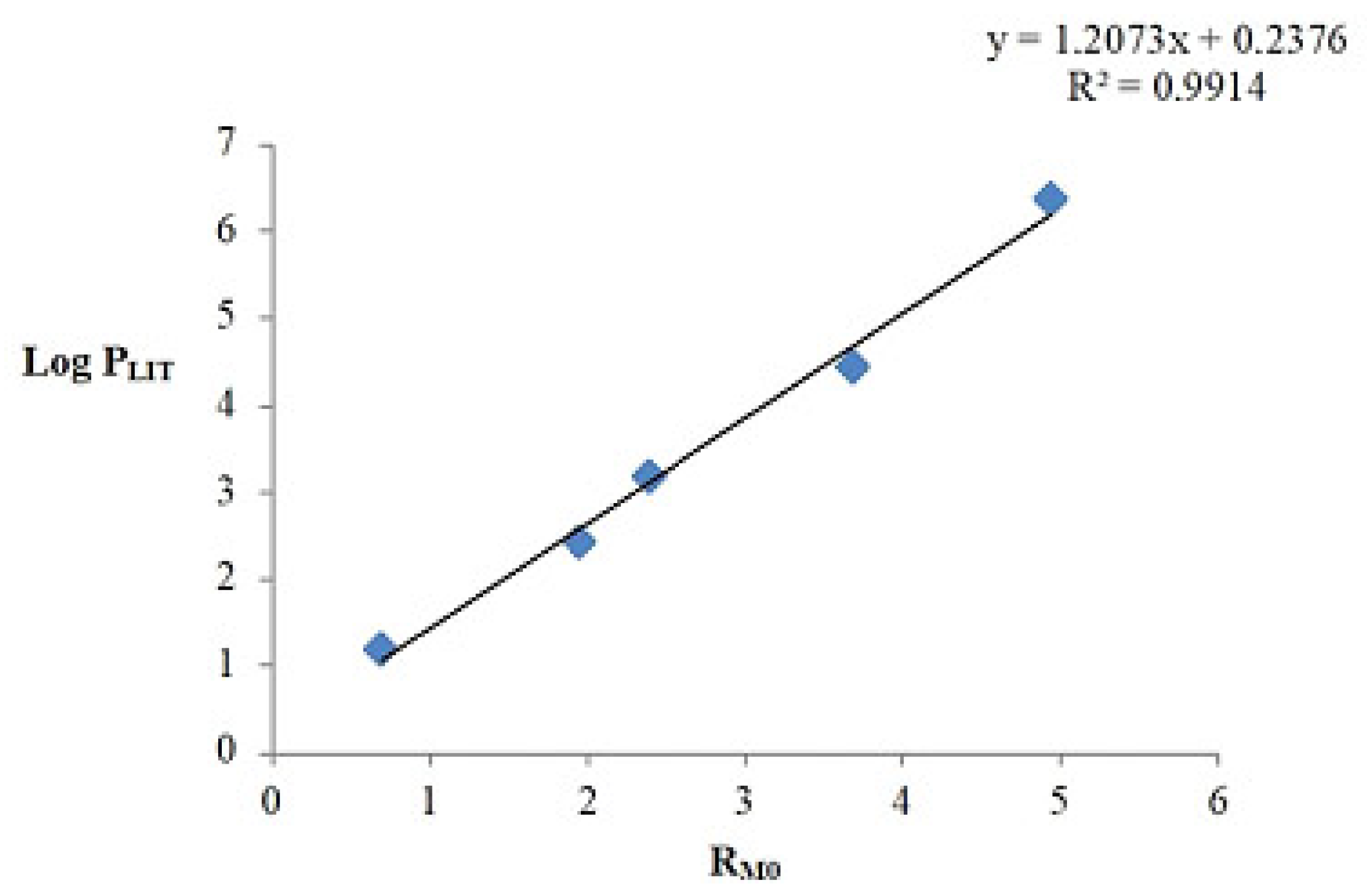
3.2. Lipophilicity
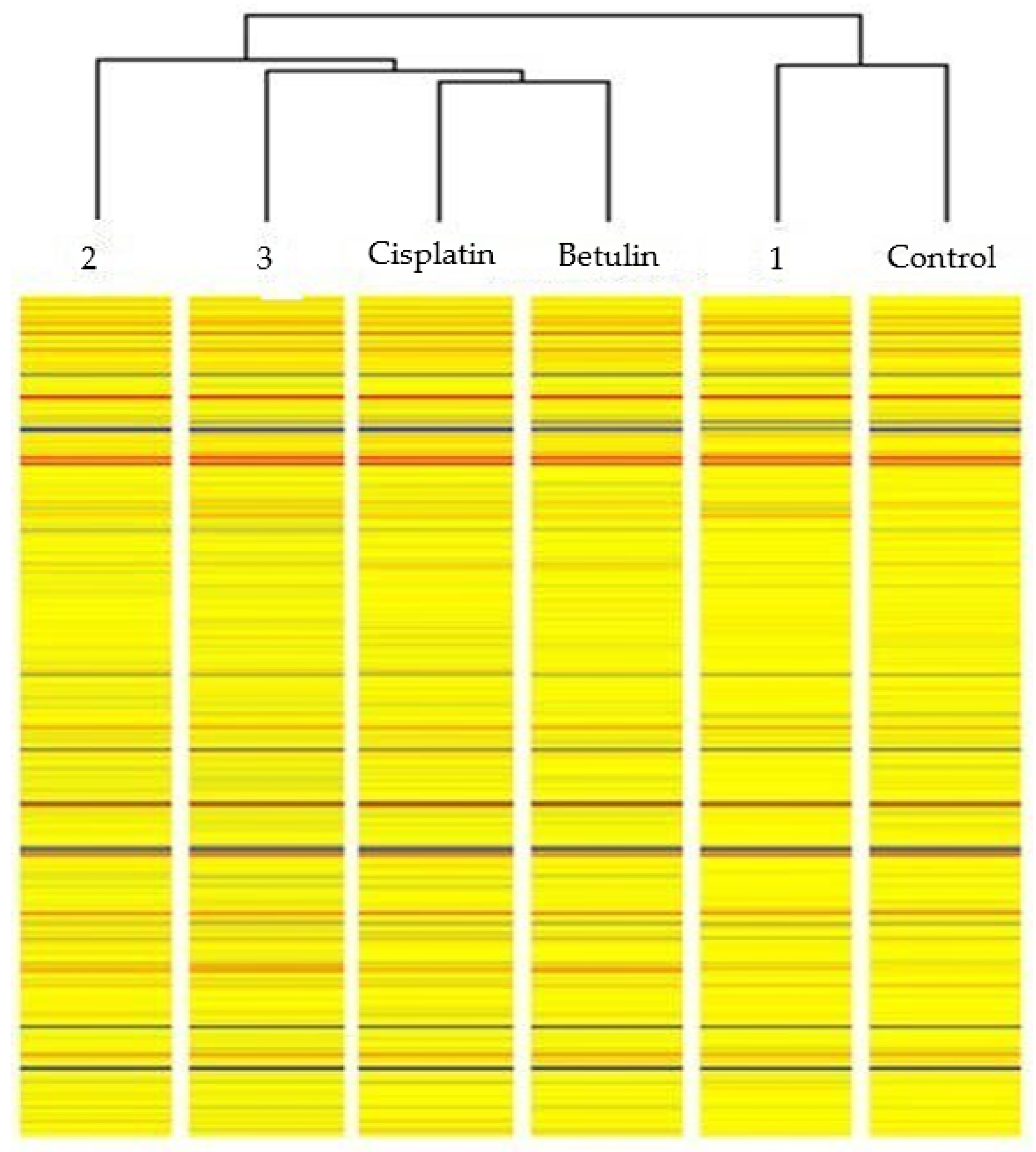
3.3. Evaluation of Changes in the Gene Expression Profile of Proteins Involved in Inflammatory Processes in Colorectal Cancer Cells
- HT-29 cells treated with betulin vs. control NHDF cells;
- HT-29 cells treated with derivative 1 vs. control NHDF cells;
- HT-29 cells treated with derivative 2 vs. control NHDF cells;
- HT-29 cells treated with derivative 3 vs. control NHDF cells;
- HT-29 cells treated with cisplatin vs. control NHDF cells.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef]
- Król, S.K.; Kiełbus, M.; Rivero-Müller, A.; Stepulak, A. Comprehensive review on betulin as a potent anticancer agent. Biomed. Res. Int. 2015, 2015, 584189. [Google Scholar] [CrossRef]
- Alakurtti, S.; Mäkelä, T.; Koskimies, S.; Yli-Kauhaluoma, J. Pharmacological properties of the ubiquitous natural product betulin. Eur. J. Pharm. Sci. 2006, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boryczka, S.; Bębenek, E.; Wietrzyk, J.; Kempińska, K.; Jastrzębska, M.; Kusz, J.; Nowak, M. Synthesis, structure and cytotoxic activity of new acetylenic derivatives of betulin. Molecules 2013, 18, 4526–4543. [Google Scholar] [CrossRef]
- Tolstikov, G.A.; Flekhter, O.B.; Shultz, E.; Baltina, L.A.; Tolstikov, A.G. Betulin and Its Derivatives. Chemistry and Biological Activity. Chem. Sustain. Dev. 2005, 13, 1–29. [Google Scholar]
- Li, Y.; He, K.; Huang, Y.; Zheng, D.; Gao, C.; Cui, L.; Jin, Y.H. Betulin induces mitochondrial cytochrome c release associated apoptosis in human cancer cells. Mol. Carcinog. 2010, 49, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Dehelean, C.A.; Soica, C.; Ledeţi, I.; Aluaş, M.; Zupko, I.G.; Luşcan, A.; Munteanu, M. Study of the betulin enriched birch bark extracts effects on human carcinoma cells and ear inflammation. Chem. Cent. J. 2012, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Yim, N.H.; Jung, Y.P.; Kim, A.; Kim, T.; Ma, J.Y. Induction of apoptotic cell death by betulin in multidrug-resistant human renal carcinoma cells. Oncol. Rep. 2015, 34, 1058–1064. [Google Scholar] [CrossRef]
- Oh, S.H.; Choi, J.E.; Lim, S.C. Protection of betulin against cadmium-induced apoptosis in hepatoma cells. Toxicology 2006, 220, 1–12. [Google Scholar] [CrossRef]
- Yang, L.; Taiyi, S.J.; Chang, G.; Qing, L.; Ying-hua, J. Caspase-9 Activation—Critical for Betulin-induced Apoptosis of Human Hepatoma Cells. Chem. Res. Chin. Univ. 2010, 26, 792–797. [Google Scholar]
- Li, Y.; Liu, X.; Jiang, D.; Lin, Y.; Wang, Y.; Li, Q.; Jin, Y.H. Betulin induces reactive oxygen species-dependent apoptosis in human gastric cancer SGC7901 cells. Arch. Pharm. Res. 2016, 39, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhu, C.; Cai, Z.; Zhao, F.; He, L.; Lou, X.; Qi, X. Betulin induces cytochrome c release and apoptosis in colon cancer cells via NOXA. Oncol. Lett. 2018, 15, 7319–7327. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Prasad, S.; Sung, B.; Kannappan, R.; Aggarwal, B.B. Targeting inflammatory pathways by triterpenoids for prevention and treatment of cancer. Toxins 2010, 2, 2428–2466. [Google Scholar] [CrossRef]
- Bernard, P.; Scior, T.; Didier, B.; Hiber, M.; Berthon, J.Y. Ethnopharmacology and bioinformatic combination for leads discovery: Application to phospholipase A(2) inhibitors. Phytochemistry 2001, 58, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Nennig, S.E.; Schank, J.R. The Role of NFkB in Drug Addiction: Beyond Inflammation. Alcohol Alcohol. 2017, 52, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Zhao, Q.F.; Fang, N.N.; Yu, J.G. Betulin inhibits pro-inflammatory cytokines expression through activation STAT3 signaling pathway in human cardiac cells. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 455–460. [Google Scholar]
- Wan, Y.; Jiang, S.; Lian, L.H.; Bai, T.; Cui, P.H.; Sun, X.T.; Nan, J.X. Betulinic acid and betulin ameliorate acute ethanol-induced fatty liver via TLR4 and STAT3 in vivo and in vitro. Int. Immunopharmacol. 2013, 17, 184–190. [Google Scholar] [CrossRef]
- Zatorski, H.; Sałaga, M.; Zielińska, M. Czynniki genetyczne w patogenezie, przebiegu i leczeniu nieswoistych chorób zapalnych jelit. Postepy Hig. Med. Dosw. 2015, 69, 335–344. [Google Scholar] [CrossRef]
- Laavola, M.; Haavikko, R.; Hämäläinen, M.; Leppänen, T.; Nieminen, R.; Alakurtti, S.; Moilanen, E. Betulin Derivatives Effectively Suppress Inflammation in Vitro and in Vivo. J. Nat. Prod. 2016, 79, 274–280. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Jóźwiak, K.; Szumiło, H.; Soczewiński, E. Lipofilowość, metody wyznaczania i rola w działaniu biologicznym substancji chemicznych. Wiadomości Chem. 2001, 55, 1048–1074. [Google Scholar]
- Alqahtani, S. In silico ADME-Tox modeling: Progress and prospects. Expert. Opin. Drug Metab. Toxicol. 2017, 13, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Tsopelas, F.; Giaginis, C.; Tsantili-Kakoulidou, A. Lipophilicity and biomimetic properties to support drug discovery. Expert. Opin. Drug Discov. 2017, 12, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Płużański, A. Kryteria oceny odpowiedzi na leczenie RECIST 1.1. J. Oncol. 2014, 64, 331–335. [Google Scholar] [CrossRef]
- Platts, J. Insulin therapy and cancer risk in diabetes mellitus. Clin. Med. 2010, 10, 509–512. [Google Scholar] [CrossRef]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef]
- Protti, M.P.; De Monte, L. Cross-talk within the tumor microenvironment mediates Th2-type inflammation in pancreatic cancer. Oncoimmunology 2012, 1, 89–91. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Bębenek, E.; Chrobak, E.; Wietrzyk, J.; Kadela, M.; Chrobak, A.; Kusz, J.; Książek, M.; Jastrzębska, M.; Boryczka, S. Synthesis, structure and cytotoxic activity of acetylenic derivatives of betulonic and betulinic acids. J. Mol. Struct. 2016, 1106, 210–219. [Google Scholar] [CrossRef]
- Kaps, A.; Chodurek, E.; Orchel, A.; Jaworska-Kik, M.; Bębenek, E.; Boryczka, S.; Kasperczyk, J. Influence of 28-O-propynoylbetulin on proliferation and apoptosis of melanotic and amelanotic human melanoma cells. Postepy. Hig. Med. Dosw. 2016, 70, 1404–1408. [Google Scholar]
- Banerjee, A.; Pathak, S.; Subramanium, V.D.; Murugesan, R.; Verma, R.S. Strategies for targeted drug delivery in treatment of colon cancer: Current trends and future perspectives. Drug Discov. Today 2017, 22, 1224–1232. [Google Scholar] [CrossRef]
- Drąg-Zalesińska, M.; Wysocka, T.; Borska, S.; Drąg, M.; Poręba, M.; Choromańska, A.; Saczko, J. The new esters derivatives of betulin and betulinic acid in epidermoid squamous carcinoma treatment—In vitro studies. Biomed. Pharmacother. 2015, 72, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Bębenek, E.; Chodurek, E.; Orchel, A.; Dzierżewicz, Z.; Boryczka, S. Antiproliferative Activity of Novel Acetylenic Derivatives of Betulin against G-361 Human Melanoma Cells. Acta Pol. Pharm. 2015, 72, 699–703. [Google Scholar]
- Bębenek, E.; Kadela-Tomanek, M.; Chrobak, E.; Wietrzyk, J.; Sadowska, J.; Boryczka, S. New acetylenic derivatives of betulin and betulone, synthesis and cytotoxic activity. Med. Chem. Res. 2017, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Szoka, L.; Karna, E.; Hlebowicz-Sarat, K.; Karaszewski, J.; Boryczka, S.; Palka, J.A. Acetylenic derivative of betulin induces apoptosis in endometrial adenocarcinoma cell line. Biomed. Pharmacother. 2017, 95, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Lalani, R.; Bardoliwala, D.; Ghosh, S.; Misra, A. Lipid-Based Oral Formulation Strategies for Lipophilic Drugs. AAPS PharmSciTech 2018, 19, 3609–3630. [Google Scholar] [CrossRef] [PubMed]
- Luu Hoang, K.N.; Anstee, J.E.; Arnold, J.N. The Diverse Roles of Heme Oxygenase-1 in Tumor Progression. Front. Immunol. 2021, 12, 658315. [Google Scholar] [CrossRef]
- Gamage, S.M.K.; Nanayakkara, S.; Macfarlane, L.; Hewage, D.; Cheng, T.; Aktar, S.; Lu, C.T.; Dissabandara, L.; Islam, F.; Lam, A.K.; et al. Heme oxygenase-1 & 2 and their potential contribution in heme induced colorectal carcinogenesis. Pathol. Res. Pract. 2022, 233, 153885. [Google Scholar] [CrossRef]
- Biasci, D.; Smoragiewicz, M.; Connell, C.M.; Wang, Z.; Gao, Y.; Thaventhiran, J.E.D.; Basu, B.; Magiera, L.; Johnson, T.I.; Bax, L.; et al. CXCR4 inhibition in human pancreatic and colorectal cancers induces an integrated immune response. Proc. Natl. Acad. Sci. USA 2020, 117, 28960–28970. [Google Scholar] [CrossRef]
- Qiu, L.; Xu, Y.; Xu, H.; Yu, B. The clinicopathological and prognostic value of CXCR4 expression in patients with lung cancer: A meta-analysis. BMC Cancer 2022, 22, 681. [Google Scholar] [CrossRef]
- Bjørge, L.; Vedeler, C.A.; Ulvestad, E.; Matre, R. Expression and function of CD59 on colonic adenocarcinoma cells. Eur. J. Immunol. 1994, 24, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, Q.; Liao, Q.; Zhao, Y. CD59: A promising target for tumor immunotherapy. Future Oncol. 2018, 14, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Jinna, N.; Rida, P.; Su, T.; Gong, Z.; Yao, S.; LaBarge, M.; Natarajan, R.; Jovanovic-Talisman, T.; Ambrosone, C.; Seewaldt, V. The DARC Side of Inflamm-Aging: Duffy Antigen Receptor for Chemokines (DARC/ACKR1) as a Potential Biomarker of Aging, Immunosenescence, and Breast Oncogenesis among High-Risk Subpopulations. Cells 2022, 11, 3818. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Liu, M.; Hu, Y.; An, W.; Liang, X.; Yu, W.; Piao, F. Expression of Duffy antigen receptor for chemokines (DARC) is down-regulated in colorectal cancer. J. Recept. Signal Transduct. Res. 2015, 35, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Gowhari Shabgah, A.; Amir, A.; Gardanova, Z.R.; Olegovna Zekiy, A.; Thangavelu, L.; Ebrahimi Nik, M.; Ahmadi, M.; Gholizadeh Navashenaq, J. Interleukin-25: New perspective and state-of-the-art in cancer prognosis and treatment approaches. Cancer Med. 2021, 10, 5191–5202. [Google Scholar] [CrossRef]
- Lim, E.J.; Kang, J.H.; Kim, Y.J.; Kim, S.; Lee, S.J. ICAM-1 promotes cancer progression by regulating SRC activity as an adapter protein in colorectal cancer. Cell Death Dis. 2022, 13, 417. [Google Scholar] [CrossRef]
- Hassanzadeh, P. Colorectal cancer and NF-κB signaling pathway. Gastroenterol. Hepatol. Bed Bench 2011, 4, 127–132. [Google Scholar]
- Warr, N.; Siggers, P.; Carré, G.A.; Wells, S.; Greenfield, A. Genetic Analyses Reveal Functions for MAP2K3 and MAP2K6 in Mouse Testis Determination. Biol. Reprod. 2016, 94, 103. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 Value ± SD (µg/mL) | |
|---|---|---|
| HT-29 | NHDF | |
| Betulin | 0.1 ± 0.03 | >100 |
| 28-O-Propynoylbetulin 1 | 9.31 ± 0.59 | 0.85 ± 0.07 |
| 28-O-Propargyloxycarbonylbetulin 2 | 9.7 ± 0.42 | >100 |
| 3-Butynyl betulinate 3 | 8.57 ± 0.97 | >100 |
| Cisplatin | 33.74 ± 5.03 | 72.91 ± 5.34 |
| Compound | RM0 | log PTLC |
|---|---|---|
| Betulin | 3.765 | 4.781 |
| 28-O-Propynoylbetulin 1 | 5.763 | 7.193 |
| 28-O-Propargyloxycarbonylbetulin 2 | 6.413 | 7.977 |
| 3-Butynyl betulinate 3 | 6.197 | 7.717 |
| One-Way ANOVA Analysis | ||||||
|---|---|---|---|---|---|---|
| p < 0.05 | p < 0.02 | p < 0.01 | p < 0.005 | p < 0.001 | ||
| Transcripts | 973 | 68 | 32 | 16 | 8 | 1 |
| Group Name | Control | Betulin | Cisplatin | 1 | 3 | 2 |
|---|---|---|---|---|---|---|
| Control | 68 | 5 | 10 | 6 | 14 | 18 |
| Betulin | 63 | 68 | 3 | 13 | 4 | 12 |
| Cisplatin | 58 | 65 | 68 | 11 | 3 | 7 |
| 1 | 62 | 55 | 57 | 68 | 18 | 20 |
| 3 | 54 | 64 | 65 | 50 | 68 | 3 |
| 2 | 50 | 56 | 61 | 48 | 65 | 68 |
| number of ID mRNAs determined by ANOVA test | ||||||
| number of non-representative ID mRNAs in the differentiation of compared groups | ||||||
| number of representative ID mRNAs for differentiation between the compared transcriptome groups at p < 0.05 | ||||||
| mRNA | p-Value | FC (log2) | Change of Expression | |
|---|---|---|---|---|
| ID | Gene Symbol | |||
| Betulin vs. K | ||||
| 200984_s_at | CD59 | 0.022 | 1.288 | increase |
| 208335_s_at | DARC | 0.004 | 1.285 | reduction |
| 209666_s_at | CHUK | 0.002 | 1.400 | increase |
| 212501_at | CEBPB | 0.007 | 1.398 | increase |
| 220971_at | IL25 | 0.024 | 1.157 | reduction |
| Compound 1 vs. K | ||||
| 200989_at | HIF1A | 0.028 | 1.430 | increase |
| 202637_s_at | ICAM1 | 0.008 | 1.332 | reduction |
| 203665_at | HMOX1 | 0.0001 | 2.842 | increase |
| 207667_s_at | MAP2K3 | 0.004 | 1.178 | reduction |
| 208023_at | TNFRSF4 | 0.003 | 1.263 | reduction |
| 217028_at | CXCR4 | 0.018 | 1.518 | reduction |
| Compound 2 vs. K | ||||
| 266_s_at | CD24 | 0.003 | 1.412 | increase |
| 202625_at | LYN | 0.012 | 1.353 | increase |
| 202637_s_at | ICAM1 | 0.008 | 1.399 | reduction |
| 203381_s_at | APOE | 0.007 | 1.942 | increase |
| 204463_s_at | EDNRA | 0.013 | 1.106 | increase |
| 205392_s_at | CCL14/CCL15 | 0.006 | 1.234 | reduction |
| 205729_at | OSMR | 0.003 | 1.127 | reduction |
| 206150_at | CD27 | 0.008 | 1.321 | reduction |
| 206541_at | KLKB1 | 0.011 | 1.246 | increase |
| 209341_s_at | IKBKB | 0.036 | 1.193 | reduction |
| 209666_s_at | CHUK | 0.002 | 1.298 | increase |
| 210168_at | C6 | 0.012 | 1.219 | reduction |
| 213095_x_at | AIF1 | 0.007 | 1.328 | reduction |
| 215760_s_at | SBNO2 | 0.024 | 1.277 | increase |
| 216042_at | TNFRSF25 | 0.018 | 1.377 | reduction |
| 216379_x_at | CD24 | 0.021 | 1.198 | increase |
| 217563_at | CLOCK | 0.015 | 1.261 | reduction |
| 220971_at | IL25 | 0.024 | 1.169 | reduction |
| Compound 3 vs. K | ||||
| 266_s_at | CD24 | 0.003 | 1.322 | increase |
| 200989_at | HIF1A | 0.028 | 1.480 | increase |
| 202637_s_at | ICAM1 | 0.008 | 1.272 | reduction |
| 204490_s_at | CD44 | 0.021 | 1.635 | increase |
| 205392_s_at | CCL14/CCL15 | 0.006 | 1.210 | reduction |
| 207785_s_at | RBPJ | 0.029 | 1.455 | increase |
| 208023_at | TNFRSF4 | 0.003 | 1.297 | reduction |
| 208303_s_at | CRLF2 | 0.020 | 1.456 | reduction |
| 208650_s_at | CD24 | 0.004 | 1.477 | increase |
| 209666_s_at | CHUK | 0.002 | 1.496 | increase |
| 210423_s_at | SLC11A1 | 0.047 | 1.226 | reduction |
| 214658_at | TMED7 | 0.010 | 2.103 | increase |
| 216042_at | TNFRSF25 | 0.018 | 1.465 | reduction |
| 220246_at | CAMK1D | 0.013 | 1.247 | reduction |
| Cisplatin vs. K | ||||
| 202685_s_at | AXL | 0.040 | 1.272 | reduction |
| 203915_at | CXCL9 | 0.023 | 1.320 | reduction |
| 204490_s_at | CD44 | 0.021 | 1.664 | increase |
| 206991_s_at | CCR5 | 0.036 | 1.424 | reduction |
| 207667_s_at | MAP2K3 | 0.004 | 1.101 | reduction |
| 209666_s_at | CHUK | 0.002 | 1.405 | increase |
| 211269_s_at | IL2RA | 0.034 | 1.547 | reduction |
| 213095_x_at | AIF1 | 0.007 | 1.440 | reduction |
| 215760_s_at | SBNO2 | 0.024 | 1.262 | increase |
| 220133_at | ODAM | 0.049 | 1.325 | reduction |
| Biological Process | Homo Sapiens (REF) | Differentiating Genes | p-Value | |
|---|---|---|---|---|
| Number | Gene Symbol | |||
| Response to oxidative stress | 12 | 1 | HMOX1 | 2.26 * 10−2 |
| Response to bacterial lipopolysaccharide | 12 | 1 | SLC11A1 | 2.26 * 10−2 |
| Anti-microbial immune response | 399 | 2 | CCR5, MAP2K3, CXCR4 | 4.03 * 10−2 |
| Immune cell chemotaxis | 94 | 3 | CCR5, CXCR4, CXCL9 | 7.83 * 10−4 |
| Defense response | 231 | 3 | DARC, MAP2K3, CXCL9 | 9.71 * 10−3 |
| Cytokine-mediated signaling pathway | 107 | 2 | CHUK, CXCL9 | 1.78 * 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubczyńska, A.; Bębenek, E.; Garncarczyk, A.; Wcisło-Dziadecka, D. Evaluation of the Effect of Betulin and Its Alkynyl Derivatives on the Profile of Changes in Gene Expression of the Inflammatory Process of Colorectal Adenocarcinoma Cells (HT-29 Cell Line). Processes 2023, 11, 2676. https://doi.org/10.3390/pr11092676
Lubczyńska A, Bębenek E, Garncarczyk A, Wcisło-Dziadecka D. Evaluation of the Effect of Betulin and Its Alkynyl Derivatives on the Profile of Changes in Gene Expression of the Inflammatory Process of Colorectal Adenocarcinoma Cells (HT-29 Cell Line). Processes. 2023; 11(9):2676. https://doi.org/10.3390/pr11092676
Chicago/Turabian StyleLubczyńska, Agnieszka, Ewa Bębenek, Agnieszka Garncarczyk, and Dominika Wcisło-Dziadecka. 2023. "Evaluation of the Effect of Betulin and Its Alkynyl Derivatives on the Profile of Changes in Gene Expression of the Inflammatory Process of Colorectal Adenocarcinoma Cells (HT-29 Cell Line)" Processes 11, no. 9: 2676. https://doi.org/10.3390/pr11092676
APA StyleLubczyńska, A., Bębenek, E., Garncarczyk, A., & Wcisło-Dziadecka, D. (2023). Evaluation of the Effect of Betulin and Its Alkynyl Derivatives on the Profile of Changes in Gene Expression of the Inflammatory Process of Colorectal Adenocarcinoma Cells (HT-29 Cell Line). Processes, 11(9), 2676. https://doi.org/10.3390/pr11092676