
An Investigation into the Potential of a Penicillium Commune Strain to Eliminate Aromatic Compounds

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Culture Conditions
2.2. Analytical Methods
2.2.1. Determination of the Concentration of Phenol and Phenolic Derivatives
2.2.2. Extraction of PAHs and GC–MS Analyses
2.2.3. Determination of the Enzyme Activities
2.2.4. Isolation of DNA from Molds and PCR Sequencing Analyses
3. Results
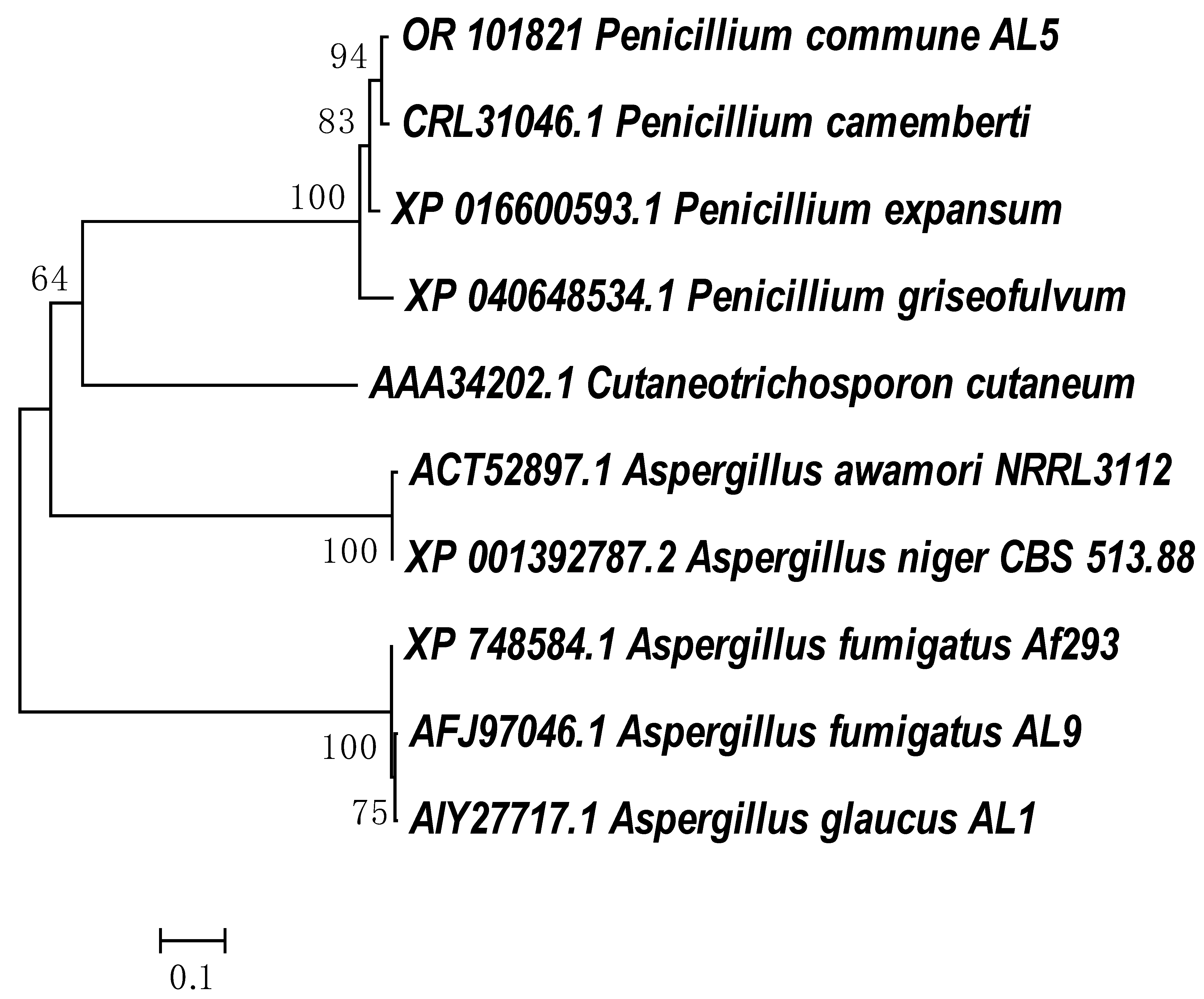
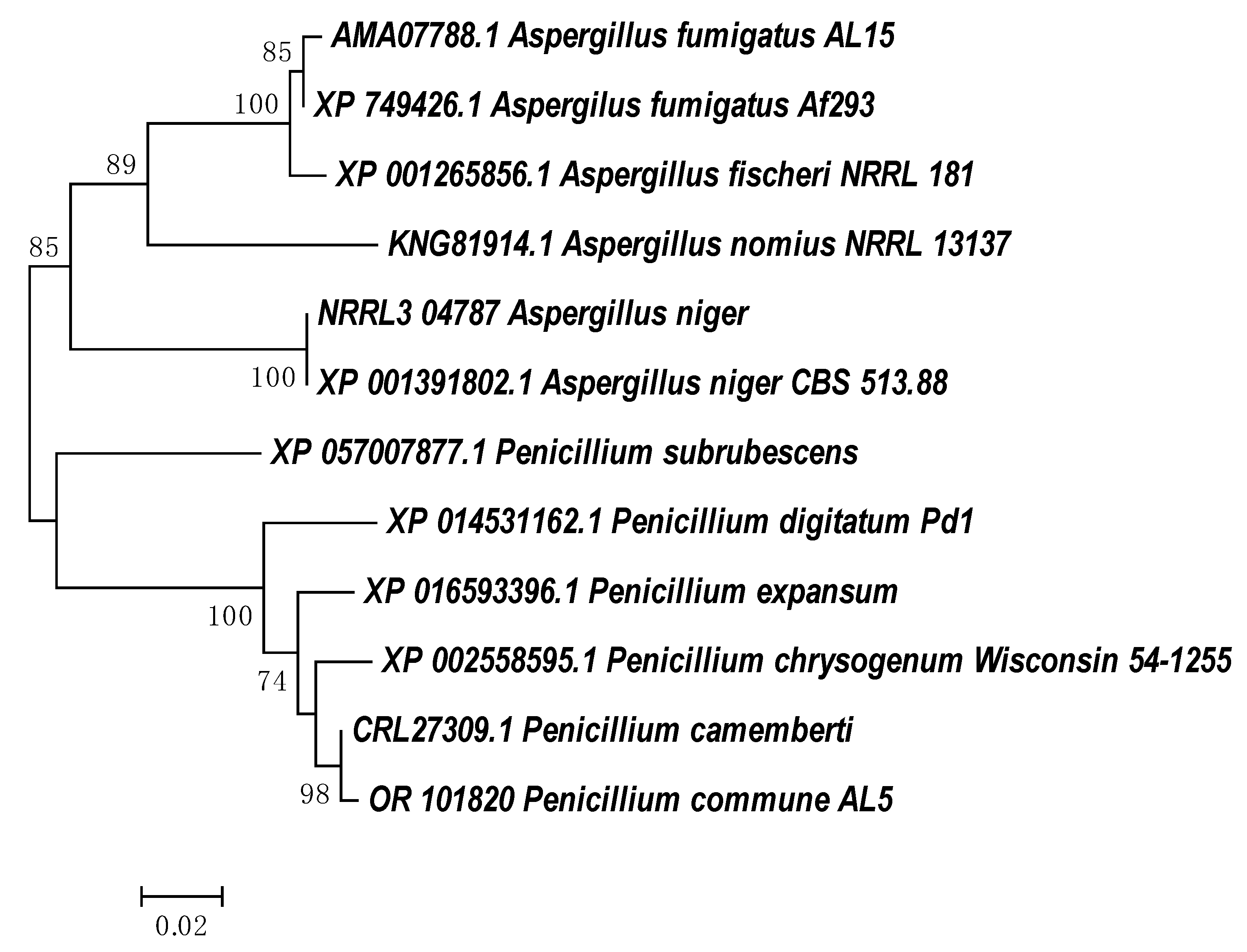
3.1. Molecular Identification
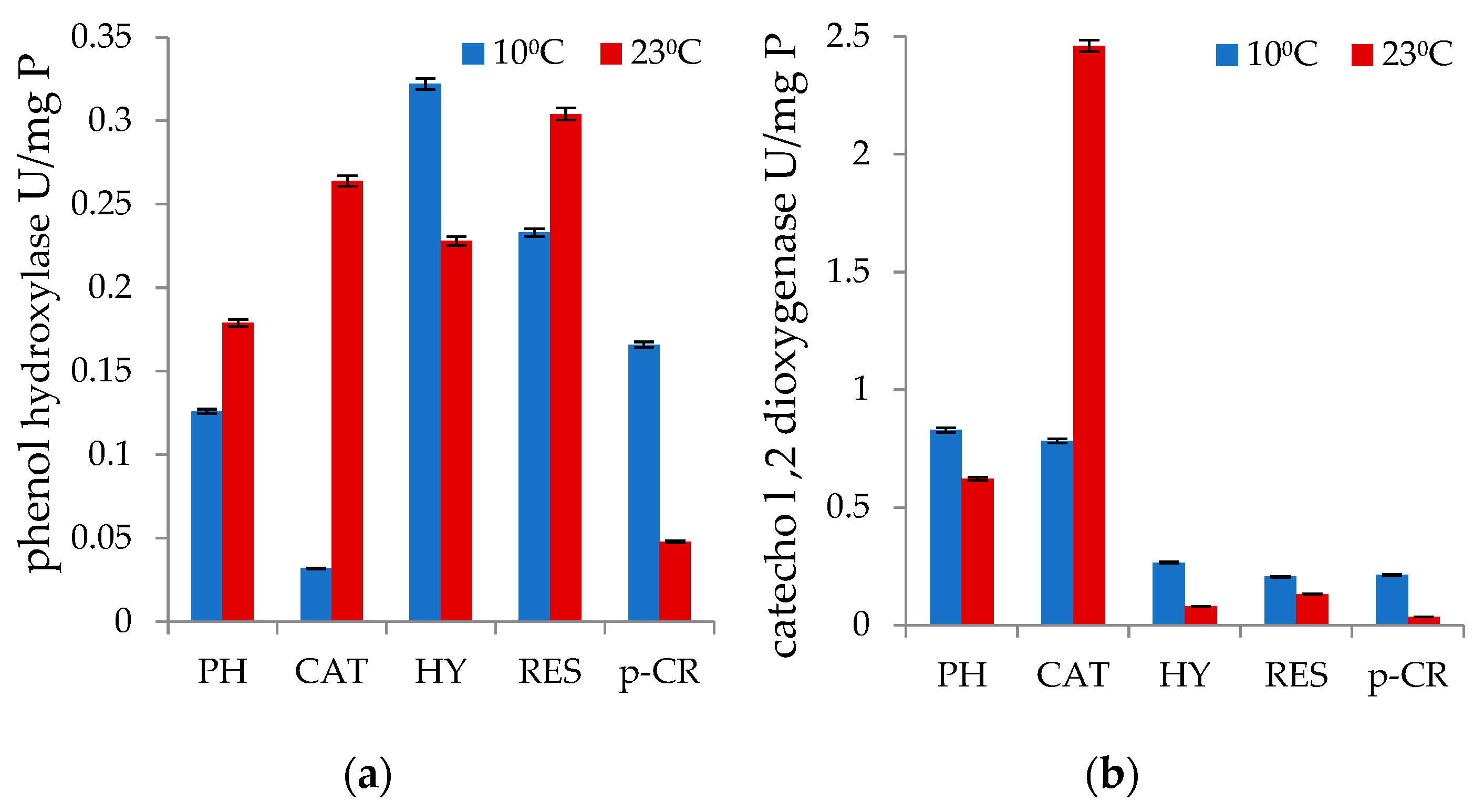
3.2. Degradation of Monophenols and Enzyme Analyses
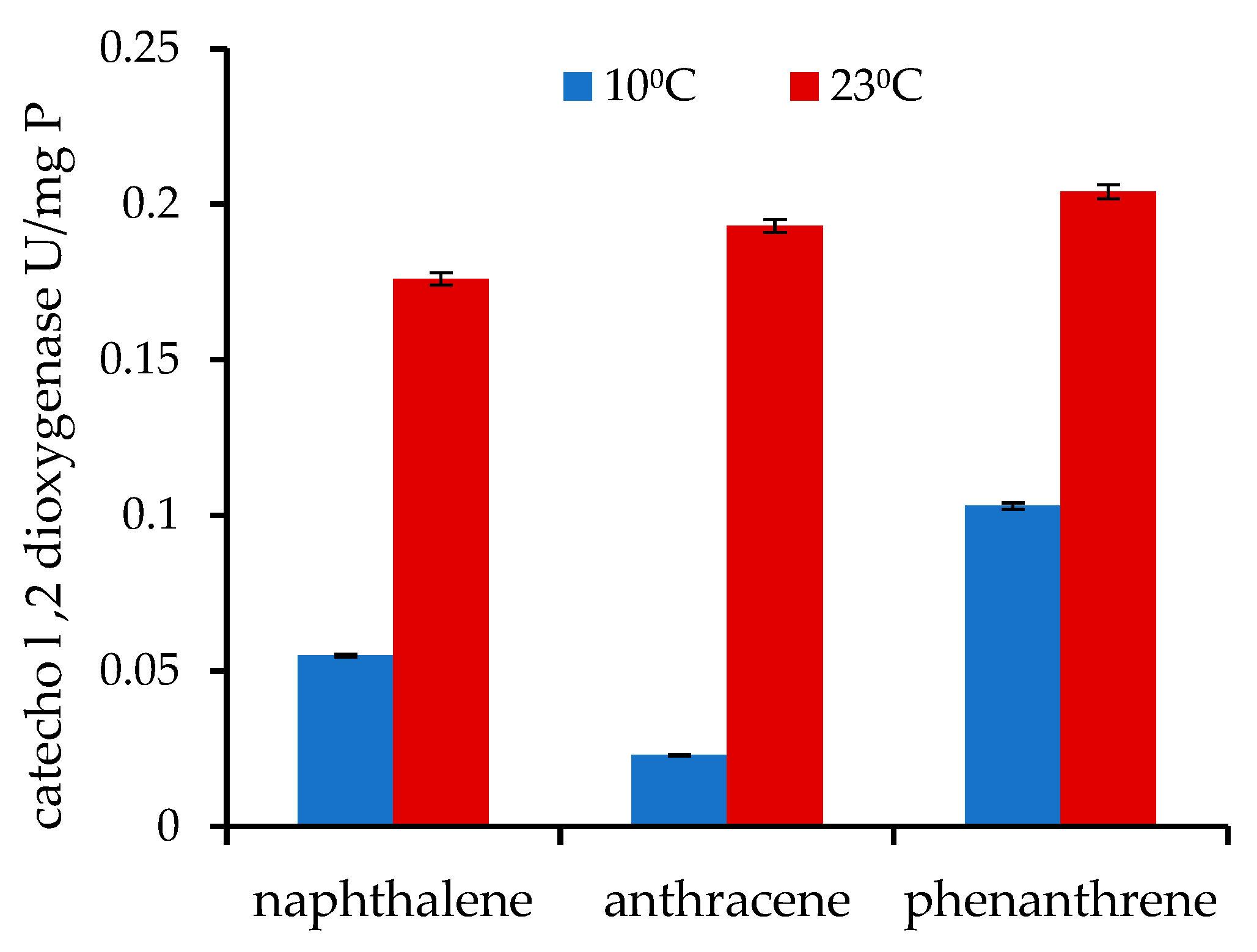
3.3. Degradation of PAHs and Catechol 1,2-Dioxygenase Activity
Intermediate Metabolites
3.4. Molecular Analyses of Phenol Hydroxylase and Catechol 1,2-Dioxigenase Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leahy, J.; Tracy, K.; Eley, M. Degradation of mixtures of aromatic and chloroaliphatic hydrocarbons by aromatic hydrocarbon-degrading bacteria. FEMS Microbiol. Ecol. 2006, 43, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Alexieva, Z.; Gerginova, M.; Zlateva, P.; Manasiev, J.; Ivanova, D.; Dimova, N. Monitoring of aromatic pollutants biodegradation. Biochem. Eng. J. 2008, 40, 233–240. [Google Scholar] [CrossRef]
- Alexieva, Z.; Yemendzhiev, H.; Zlateva, P. Cresols utilization by Trametes versicolor and substrate interactions in the mixture with phenol. Biodegradation 2010, 21, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Bhatiya, A.K.; Mishra, P.K.; Srivastava, N. An effective approach for the degradation of phenolic waste: Phenols and cresols. In Abatement of Environmental Pollutants: Trends and Strategies; Singh, P., Kumar, A., Borthakur, A., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 203–243. [Google Scholar]
- Vasileva, E.; Parvanova-Mancheva, T.; Beschkov, V.; Alexieva, Z.; Gerginova, M.; Peneva, N. Effects of constant electric field on biodegradation of phenol by free and immobilized cells of Bradyrhizobium japonicum 273. ChemEngineering 2021, 5, 75. [Google Scholar] [CrossRef]
- Zhou, Y.; Nemati, M. Co-biodegradation of phenol, o-cresol, and p-cresol in binary and ternary mixtures: Evaluation of bioreactor performance and toxicity of treated effluents. Water Air Soil Pollut. 2022, 233, 133. [Google Scholar] [CrossRef]
- Hadibarata, T.; Yusoff, A.; Aris, A.; Kristanti, R. Identification of naphthalene metabolism by white rot fungus Armillaria sp. F022. J. Environ. Sci. 2012, 24, 728–732. [Google Scholar] [CrossRef]
- Kozak, J.; Włodarczyk-Makuła, M.; Popenda, A. Impact of aerobic stabilization of sewage sludge on PAHs concentration in reject waters. J. Ecol. Eng. 2021, 22, 27–35. [Google Scholar]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Litova, K.; Gerginova, M.; Peneva, N.; Manasiev, J.; Alexieva, Z. Growth of Antarctic fungal strains on phenol at low temperatures. J. BioSci. Biotechnol. 2014, 43–46. [Google Scholar]
- Tengku-Mazuki, T.A.; Subramaniam, K.; Zakaria, N.N.; Convey, P.; Abdul, K.; Lee, G.L.Y.; Zulkhaenain, A.; Shaharuddin, N.; Ahmad, S.A. Optimization of phenol degradation by antarctic bacterium Rhodococcus sp. Antarct. Sci. 2020, 32, 486–495. [Google Scholar] [CrossRef]
- Yap, H.S.; Zakaria, N.N.; Zulkharnain, A.; Sabri, S.; Gomez-Fuentes, C.; Ahmad, S.A. Bibliometric analysis of hydrocarbon bioremediation in cold regions and a review on enhanced soil bioremediation. Biology 2021, 10, 354. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Rafiq, M.; Hassan, N.; Rehman, M.; Hasan, F. Adaptation mechanisms and applications of psychrophilic fungi. In Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Tiquia-Arashiro, S., Grube, M., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2019; pp. 157–174. [Google Scholar] [CrossRef]
- Vaksmaa, A.; Guerrero-Cruz, S.; Ghosh, P.; Zeghal, E.; Hernando-Morales, V.; Niemann, H. Role of fungi in bioremediation of emerging pollutants. Front. Mar. Sci. 2023, 10, 1070905. [Google Scholar] [CrossRef]
- Krivobok, S.; Miriouchkine, E.; Seigle-Murandi, F.; Benoit-Guyod, J.-L. Biodegradation of anthracene by soil fungi. Chemosphere 1998, 37, 523–530. [Google Scholar] [CrossRef]
- Cao, H.; Wang, C.; Liu, H.; Jia, W.; Sun, H. Enzyme activities during Benzo[a]pyrene degradation by the fungus Lasiodiplodia theobromae isolated from a polluted soil. Sci. Rep. 2020, 10, 865. [Google Scholar] [CrossRef]
- Stoilova, I.; Krastanov, A.; Stanchev, V.; Daniel, D.; Gerginova, M.; Alexieva, Z. Biodegradation of high amounts of phenol, catechol, 2,4-dichlorophenol and 2,6-dimethoxyphenol by Aspergillus awamori cells. Enzyme Microb. Technol. 2006, 39, 1036–1041. [Google Scholar] [CrossRef]
- Cai, W.; Li, J.; Zhang, Z. The characteristics and mechanisms of phenol biodegradation by Fusarium sp. J. Hazard. Mater. 2007, 148, 38–42. [Google Scholar] [CrossRef]
- Morales, P.; Cáceres, M.; Scott, F.; Díaz-Robles, L.; Aroca, G.; Vergara-Fernández, A. Biodegradation of benzo[α]pyrene, toluene, and formaldehyde from the gas phase by a consortium of Rhodococcus erythropolis and Fusarium solani. Appl. Microbiol. Biotechnol. 2017, 101, 6765–6777. [Google Scholar] [CrossRef]
- Álvarez-Barragán, J.; Cravo-Laureau, C.; Wick, L.Y.; Duran, R. Fungi in PAH-contaminated marine sediments: Cultivable diversity and tolerance capacity towards PAH. Mar. Pollut. Bull. 2021, 164, 112082. [Google Scholar] [CrossRef]
- Govarthanan, M.; Fuzisawa, S.; Hosogai, T.; Chang, Y. Biodegradation of aliphatic and aromatic hydrocarbons using the filamentous fungus Penicillium sp. CHY-2 and characterization of its manganese peroxidase activity. RSC Adv. 2017, 7, 20716–20723. [Google Scholar] [CrossRef]
- El-Gendi, H.; Saleh, A.K.; Badierah, R.; Redwan, E.M.; El-Maradny, Y.A.; El-Fakharany, E. A comprehensive insight into fungal enzymes: Structure, classification, and their role in mankind’s challenges. J. Fungi 2022, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Rafin, C.; Potin, O.; Veignie, E.; Sahraoui, A.L.H.; Sancholle, M. Degradation of benzo[a]pyrene as sole carbon source by a non white rot fungus, Fusarium solani. Pol. Aromat. Comp. 2006, 21, 311–329. [Google Scholar] [CrossRef]
- Gerginova, M.; Manasiev, J.; Yemendzhiev, H.; Terziyska, A.; Peneva, N.; Alexieva, Z. Biodegradation of phenol by antarctic strains of Aspergillus fumigates. Z. Naturforschung 2013, 68C, 384–393. [Google Scholar]
- Aranda, E.; Godoy, P.; Reina, R.; Badia-Fabregat, M.; Rosell, M.; Marco-Urrea, E.; García-Romera, I. Isolation of Ascomycota fungi with capability to transform PAHs: Insights into the biodegradation mechanisms of Penicillium oxalicum. Int. Biodeterior. Biodegrad. 2017, 122, 141–150. [Google Scholar] [CrossRef]
- Gerginova, M.; Stoyanova, K.; Peneva, N.; Alexieva, Z. Biodegradation of anthracene by an antarctic fungal strain a member of genus Alternaria. In Proceedings of the 17th International Conference on Environmental Science and Technology, Athens, Greece, 1–4 September 2021. [Google Scholar] [CrossRef]
- Stoyanova, K.; Gerginova, M.; Dincheva, I.; Peneva, N.; Alexieva, Z. Biodegradation of naphthalene and anthracene by Aspergillus glaucus strain isolated from Antarctic soil. Processes 2022, 10, 873. [Google Scholar] [CrossRef]
- Kukor, J.; Wawrik, B.; Zylstra, G. Metabolism of aromatic compounds. In Methods for General and Molecular Microbiology, 3rd ed.; Reddy, C.A., Beveridge, T.J., Breznak, J.A., Marzluf, G.A., Schmidt, T.M., Snyder, L.R., Eds.; ASM Press: Washington, DC, USA, 2007. [Google Scholar] [CrossRef]
- Tian, M.; Du, D.; Zhou, W.; Zeng, X.; Cheng, G. Phenol degradation and genotypic analysis of dioxygenase genes in bacteria isolated from sediments. Braz. J. Microbiol. 2017, 48, 2. [Google Scholar] [CrossRef]
- Semana, P.; Powlowski, J. Four aromatic intradiol ring cleavage dioxygenases from Aspergillus niger. Appl. Environ. Microbiol. 2019, 85, e01786-19. [Google Scholar] [CrossRef]
- Del Cerro, C.; Erickson, E.; Dong, T.; Wong, A.R.; Eder, E.K.; Purvine, S.O.; Mitchell, H.D.; Weitz, K.K.; Markillie, L.M.; Burnet, M.C.; et al. Intracellular pathways for lignin catabolism in white-rot fungi. Proc. Natl. Acad. Sci. USA 2021, 118, e2017381118. [Google Scholar] [CrossRef]
- Kacprzak, M.; Neczaj, E.; Okoniewska, E. The comparative mycological analysis of wastewater and sewage sludges from selected wastewater treatment plants. Desalination 2005, 185, 363–370. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Seifert, K.A.; Overy, D.P.; Tuthill, D.M.; Valdez, J.G.; Samson, R.A. New penicillin-producing Penicillium species and an overview of section Chrysogena. Persoonia-Mol. Phylogeny Evol. Fungi 2012, 29, 78–100. [Google Scholar] [CrossRef]
- Vrabl, P.; Siewert, B.; Winkler, J.; Schöbel, H.; Schinagl, C.W.; Knabl, L.; Orth-Höller, D.; Fiala, J.; Meijer, M.S.; Bonnet, S.; et al. Xanthoepocin, a photolabile antibiotic of Penicillium ochrochloron CBS 123823 with high activity against multiresistant gram-positive bacteria. Microb. Cell Factories 2022, 21, 1. [Google Scholar] [CrossRef]
- Nazareth, S.; Marbaniang, T. Effect of heavy metals on cultural and morphological growth characteristics of halotolerant Penicillium morphotypes. J. Basic Microbiol. 2008, 48, 363–369. [Google Scholar] [CrossRef]
- Yamada-Onodera, K.; Mukumoto, H.; Katsuyaya, Y.; Saiganji, A.; Tani, Y. Degradation of polyethylene by a fungus, Penicillium simplicissimum YK. Polym. Degrad. Stab. 2001, 72, 323–327. [Google Scholar] [CrossRef]
- Hofrichter, M.; Scheibner, K. Utilization of aromatic compounds by the Penicillium strain Bi. J. Basic Microbiol. 1993, 33, 227–232. [Google Scholar] [CrossRef]
- Leitão, A.; Duarte, M.; Santos Oliveira, J. Degradation of phenol by a halotolerant strain of Penicillium chrysogenum. Int. Biodeterior. Biodegrad. 2007, 59, 220–225. [Google Scholar] [CrossRef]
- Garon, D.; Sage, L.; Seigle-Murandi, F. Effects of fungal bioaugmentationand cyclodextrin amendment on fluorene degradation in soil slurry. Biodegradation 2004, 15, 1–8. [Google Scholar] [CrossRef]
- Saraswathy, A.; Hallberg, R. Mycelial pellet formation by Penicillium ochrochloron species due to exposure to pyrene. Microbiol. Res. 2005, 160, 375–383. [Google Scholar] [CrossRef]
- Meléndez-Estrada, J.; Amezcua-Allieri, M.A.; Alvarez, P.J.; Rodríguez-Vázquez, R. Phenanthrene removal by Penicillium frequentans grown on a solid-state culture: Effect of oxygen concentration. Environ. Technol. 2006, 10, 1073–1080. [Google Scholar] [CrossRef]
- Tossi, S.; Kostadinova, N.; Krumova, E.; Pashova, S.; Dishliiska, V.; Spassova, B.; Vassilev, S.; Angelova, M. Antioxidant enzyme activity of filamentous fungi isolated from Livingston Island, Maritime Antarctica. Polar Biol. 2010, 33, 1227–1237. [Google Scholar] [CrossRef]
- Kostadinova, N.; Krumova, E.; Tosi, S.; Pashova, S.; Angelova, M. Isolation and identifi cation of filamentous fungi from island Livingston, Antarctica. Biotechnol. Biotechnol. Equip. 2009, 23SE, 267–270. [Google Scholar] [CrossRef]
- Hristov, A. Change in the processes of microbial respiration in Black Sea ecosystem in the presence of phenol. Comptes Rendus Acad. Bulg. Sci. 1997, 50, 101–104. [Google Scholar]
- Jones, K.; Trudgill, P.; Hopper, D. Evidence of two pathways for the metabolism of phenol by Aspergillus fumigatus. Arch. Microbiol. 1995, 163, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Dincheva, I.; Badjakov, I.; Kondakova, V.; Batchvarova, R. Metabolic profiling of red raspberry (Robus idaeus) during fruit development and ripening. Int. J. Agric. Sci. 2013, 3, 81–88. [Google Scholar]
- Neujahr, H.; Gaal, A. Phenol Hydroxylase from Yeast. Purification and properties of the enzyme from yeast Trichosporon cutaneum. Eur. J. Biochem. 1973, 35, 386–400. [Google Scholar] [CrossRef]
- Varga, J.; Neujah, H. Purification and Properties of catechol 1,2-oxygenase from Trichosporon cutaneum. Eur. J. Biochem. 1970, 12, 4. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Liu, D.; Coloe, S.; Baird, R.; Pederson, J. Rapid mini-preparation of fungal DNA for PCR. J. Clin. Microbiol. 2000, 38, 471. [Google Scholar] [CrossRef]
- Jaeger, E.; Carroll, N.; Choudhury, S.; Dunlop, A.; Towler, H.; Matheson, M.; Adamson, P.; Okhravi, N. Rapid detection and identification of Candida, Aspergillus, and Fusarium species in ocular samples using nested PCR. J. Clin. Microbiol. 2000, 38, 2902–2908. [Google Scholar] [CrossRef]
- White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ballester, A.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef]
- Cheeseman, K.; Ropars, J.; Renault, P.; Dupont, J.; Gouzy, J.; Branca, A.; Abraham, A.; Ceppi, M.; Conseiller, E.; Debuchy, R.; et al. Multiple recent horizontal transfers of a large genomic region in cheese making fungi. Nat. Commun. 2014, 5, 2876. [Google Scholar] [CrossRef]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef]
- Harzallah, B.; Grama, S.B.; Bousseboua, H.; Jouanneau, Y.; Yang, J.; Li, J. Isolation and characterization of Indigenous Bacilli strains from an oil refinery wastewater with potential applications for phenol/cresol bioremediation. J. Environ. Manag. 2023, 332, 117322. [Google Scholar] [CrossRef]
- Krastanov, A.; Alexieva, Z.; Yemendzhiev, H. Microbial degradation of phenol and phenolic derivatives. Eng. Life Sci. 2013, 13, 76–87. [Google Scholar] [CrossRef]
- Bokade, P.; Bajaj, A. Molecular advances in mycoremediation of polycyclic aromatic hydrocarbons: Exploring fungal bacterial interactions. J. Basic Microbiol. 2023, 63, 239–256. [Google Scholar] [CrossRef]
- Wolski, E.A. The versatility of Penicillium species to degrade organic pollutants and its use for wastewater treatment. Stud. Fungi 2023, 8, 2. [Google Scholar] [CrossRef]
- Santos, V.; Linardi, V. Biodegradation of phenol by a filamentous fungi isolated from industrial effluents—Identification and degradation potential. Proc. Biochem. 2004, 39, 1001–1006. [Google Scholar] [CrossRef]
- Leitão, A.L. Potential of Penicillium species in the bioremediation field. Int. J. Environ. Res. Public Health 2009, 6, 1393–1417. [Google Scholar] [CrossRef]
- Eriksson, M.; Ka, J.; Mohn, W. Effects of low temperature and freeze-thaw cycles on hydrocarbon biodegradation in Arctic tundra soil. Appl. Environ. Microbiol. 2001, 67, 5107–5112. [Google Scholar] [CrossRef]
- Struvay, C.; Feller, G. Optimization to low temperature activity in psychrophilic enzymes. Int. J. Mol. Sci. 2012, 13, 11643–11665. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Protein stability and enzyme activity at extreme biological temperatures. J. Phys. Condens. Matter. 2010, 22, 323101. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.; Sodersten, E.; Yu, Z.; Dalhammar, G.; Mohn, W. Degradation of polycyclic aromatic hydrocarbons at low temperature under aerobic and nitrate-reducing conditions in enrichment cultures from northern soils. Appl. Environ. Microbiol. 2003, 69, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.; Viale, A.; Forchiassin, A. Degradation of organic pollutants by the white rot basidiomycete Trametes trogii. Int. Biodeterior. Biodegrad. 2003, 52, 1–5. [Google Scholar] [CrossRef]
- Uyttebroek, M.; Ortega-Calvo, J.; Breugelmans, P.; Springael, D. Comparison of mineralization of solid-sorbed phenanthrene by polycyclic aromatic hydrocarbon (PAH)-degrading Mycobacterium spp. and Sphingomonas spp. Appl. Microbiol. Biotechnol. 2006, 72, 829–836. [Google Scholar] [CrossRef]
- Guo, C.; Dang, Z.; Wong, Y.; Tam, N.F. Biodegradation ability and dioxgenase genes of PAH-degrading Sphingomonas and Mycobacterium strains isolated from mangrove sediments. Int. Biodeterior. Biodegrad. 2010, 64, 419–426. [Google Scholar] [CrossRef]
- Wu, Y.; Luo, Z.; Vrijmoed, L. Biodegradation of anthracene and benz[a]anthracene by two Fusarium solani strains isolated from mangrove sediments. Bioresour. Technol. 2010, 101, 9666–9672. [Google Scholar] [CrossRef]
- Cerniglia, C.; Kelly, D.; Freeman, J.; Miller, D. Microbial metabolism of pyrene. Chem.-Biol. Interact. 1986, 57, 203–216. [Google Scholar] [CrossRef]
- Hesham, A.E.-L.; Mawad, A.M.; Mostafa, Y.M.; Shoreit, A. Biodegradation ability and catabolic genes of petroleum-degrading Sphingomonas koreensis strain ASU-06 isolated from Egyptian oily soil. Biomed. Res. Int. 2014, 2014, 127674. [Google Scholar] [CrossRef]
- Dharmasiri, R.; Undugoda, L.; Nilmini, A.; Nugara, N.; Udayanga, D.; Manage, P. Polyaromatic hydrocarbons (PAHs) degradation ability of Penicillium spp. isolated from phyllosphere of urban areas in Sri Lanka. In Proceedings of the 26th International Forestry and Environment Symposium, Colombo, Sri Lanka, 20–21 January 2022; Volume 26. [Google Scholar] [CrossRef]
- Haritash, A.; Kaushik, C. Biodegradation aspects of Polycyclic Aromatic Hydrocarbons (PAHs): A review. J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef]
- Silva, A.; Jacques, R.; Andreazza, R.; Bento, F.; Roesch, L.; Camargo, F. Properties of catechol 1,2-dioxygenase in the cell free extract and immobilized extract of Mycobacterium fortuitum. Braz. J. Microbiol. 2013, 44, 291–297. [Google Scholar] [CrossRef]
- Zhou, Y.; ZHANG, J.; Su, E.; Wei, G.; Ma, Y.; Wei, D. Phenanthrene biodegradation by an indigenous Pseudomonas sp. ZJF08 with TX100 as surfactant. Ann. Microbiol. 2008, 58, 439–442. [Google Scholar] [CrossRef]
- Peng, R.; Xiong, A.; Xue, Y.; Fu, X.; Gao, F.; Zhao, W.; Tian, Y.; Yao, Q. Microbial biodegradation of polyaromatic hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Pitt, J.; Hocking, A. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Schreuder, H.; Prick, P.; Wierenga, R.; Vriend, G.; Wilson, K.; Hol, W.; Drenth, J. Crystal structure of the p-hydroxybenzoate hydroxylase-substrate complex refined at 1.9 Å resolution: Analysis of the enzyme-substrate and enzyme-product complexes. J. Mol. Biol. 1989, 208, 679–696. [Google Scholar] [CrossRef]
- Enroth, C.; Neujahr, H.; Schneider, G.; Lindqvist, Y. The crystal structure of phenol hydroxylase in complex with FAD and phenol provides evidence for a concerted conformational change in the enzyme and its cofactor during catalysis. Structure 1998, 6, 605–617. [Google Scholar] [CrossRef]
- Eppink, M.; Van Berkel, W.; Schreuder, H. Identification of a novel conserved sequence motif in flavoprotein hydroxylases with a putative dual function in FAD/NAD(P)H binding. Prot. Sci. 1997, 6, 2454–2458. [Google Scholar] [CrossRef]
- Berkel, W.; Kamerbeek, N.; Fraaije, M. Flavoprotein monooxygenases, a diverse class of oxidative biocatalysts. J. Biotechnol. 2006, 124, 670–689. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ → 3′) | Annealing T °C | Source |
|---|---|---|---|
| PFf | AGGGATGTATTTATTAGATAAAAAATCAA | 58 °C | [52] |
| PFr | CGCAGTAGTTAGTCTTCAGTAAATC | ||
| ITS 1 | TCCGTAGGTGAACCTGCGG | 60 °C | [53] |
| ITS 4 | TCCTCCGCTTATTGATATGC |
| Substrate | 23 °C | 10 °C | ||
|---|---|---|---|---|
| % Degradation | Time, h | % Degradation | Time, h | |
| phenol | 100 | 188 | 100 | 480 |
| resorcinol | 100 | 26 | 100 | 72 |
| catechol | 100 | 24 | 100 | 36 |
| hydroquinon | 100 | 100 | 100 | 120 |
| p-cresol | 50 | 144 | 100 | 144 |
| Substrate | 23 °C | 10 °C | ||
|---|---|---|---|---|
| % Degradation | Time, days | % Degradation | Time, days | |
| naphthalene | 77.5 | 17 | 54 | 17 |
| anthracene | 93.8 | 17 | 8.5 | 17 |
| phenanthrene | 75.1 | 17 | 22 | 17 |
| Compound | m/z of Fragment Ions (Relative Abundance, %) | Structure Confirmed with a Standard |
|---|---|---|
| Naphthalene | 160(100), 131(9), 115(4), 114(5), 113(6), 88(5), 80(8), 77(9), 51(6) | 1,2-Dihydroxynaphthalene |
| 122(100), 121(94), 104(16), 93(25), 76(26), 66(14), 65(42), 63(12), 39(41), 38(12) | Salicylaldehyde | |
| 268(8), 267(34), 135(10), 91(8), 75(8), 74(9), 73(100), 45(25), 44(4), 43(10) 254(19), 239(8), 166(4), 151(6), 135(7), 75(8),74(10), 73(100), 45(14) | Salicylic acid, TMS derivative Catechol, TMS derivative | |
| Anthracene | 324(3), 251(7), 236(6), 235(3), 178(14), 75(4), 74(7), 73(100), 45(12), 43(3) | 9,10-Dihydroanthracene, TMS derivative |
| 208(100), 181(14),180(98), 152(78), 151(33), 150(15), 76(44), 75(17), 50(19) | 9,10-Anthraquinone | |
| Phenanthrene | 188(40), 170(100), 142(42), 115(31), 114(66), 88(14), 71(17), 63(15), 57(10), 45(7) 216(100), 201(98), 185(60), 141(22), 127(21), 115(36), (73(48), 45(17) 295(16), 221(6), 149 (9), 148(16), 147(100), 141(7), 140(8), 75(6), 73(63), 45(17) 371(13), 370(38), 355(22), 311(11), 281(8), 194(13), 193(9), 74(9), 73(100), 45(22) | 1-Hydroxy-2-naphtoic acid 1-Naphthol, TMS derivative Phthalic acid, TMS derivative Protocatechuic acid, TMS derivative |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerginova, M.; Stoyanova, K.; Peneva, N.; Dincheva, I.; Alexieva, Z. An Investigation into the Potential of a Penicillium Commune Strain to Eliminate Aromatic Compounds. Processes 2023, 11, 2402. https://doi.org/10.3390/pr11082402
Gerginova M, Stoyanova K, Peneva N, Dincheva I, Alexieva Z. An Investigation into the Potential of a Penicillium Commune Strain to Eliminate Aromatic Compounds. Processes. 2023; 11(8):2402. https://doi.org/10.3390/pr11082402
Chicago/Turabian StyleGerginova, Maria, Katya Stoyanova, Nadejda Peneva, Ivayla Dincheva, and Zlatka Alexieva. 2023. "An Investigation into the Potential of a Penicillium Commune Strain to Eliminate Aromatic Compounds" Processes 11, no. 8: 2402. https://doi.org/10.3390/pr11082402
APA StyleGerginova, M., Stoyanova, K., Peneva, N., Dincheva, I., & Alexieva, Z. (2023). An Investigation into the Potential of a Penicillium Commune Strain to Eliminate Aromatic Compounds. Processes, 11(8), 2402. https://doi.org/10.3390/pr11082402