

Investigation of Synechocystis sp. CPCC 534 Motility during Different Stages of the Growth Period in Active Fluids

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture Conditions
2.2. Experimental Protocol
2.3. Calculation of Synechocystis sp. Growth Rate (r) and Doubling Time (k)
2.4. Video Microscopy and Image Acquisition
2.5. Analysis of Cell Dynamics
2.6. Statistical Analysis
3. Results and Discussion
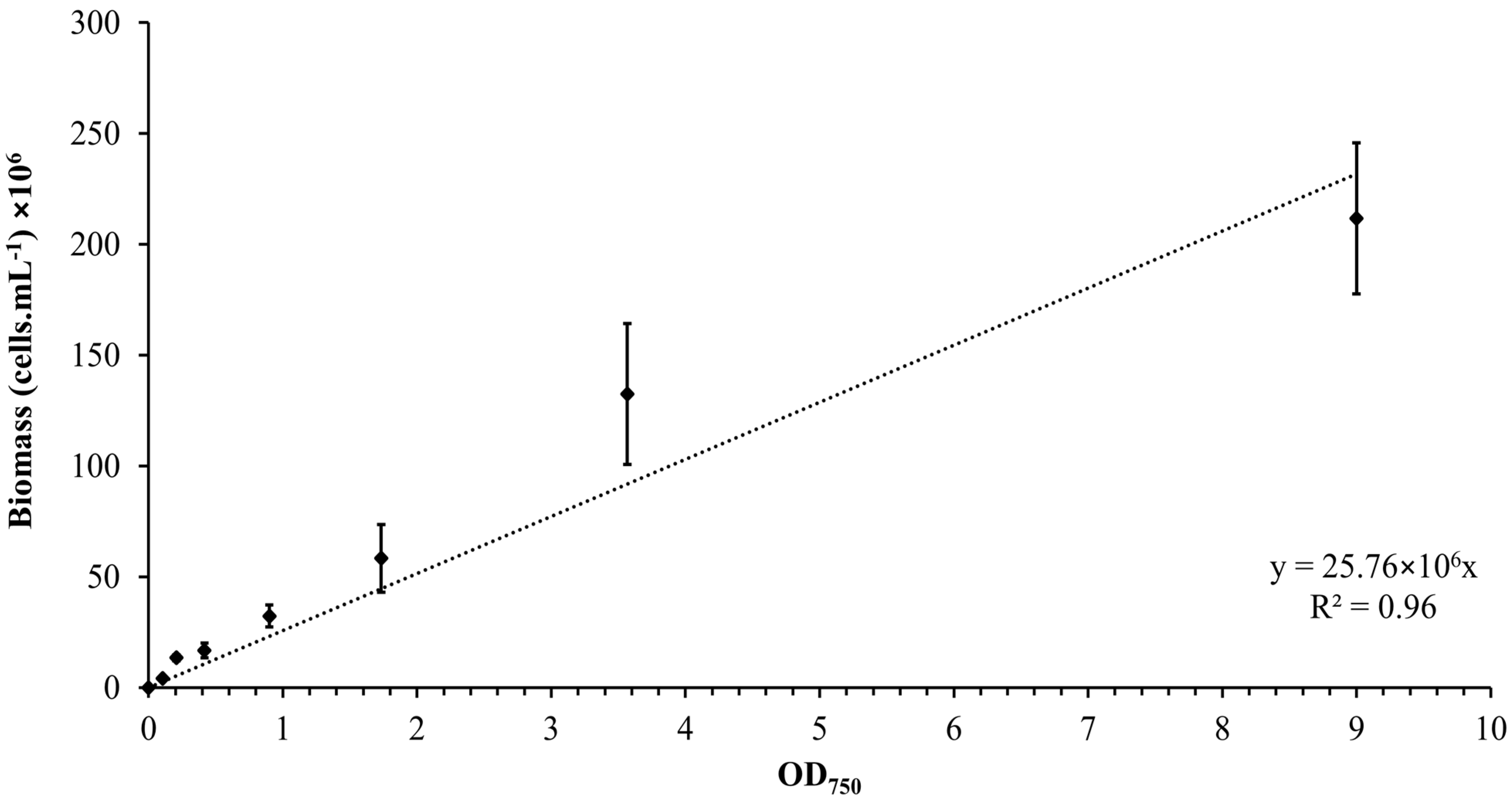
3.1. Calibration Curve
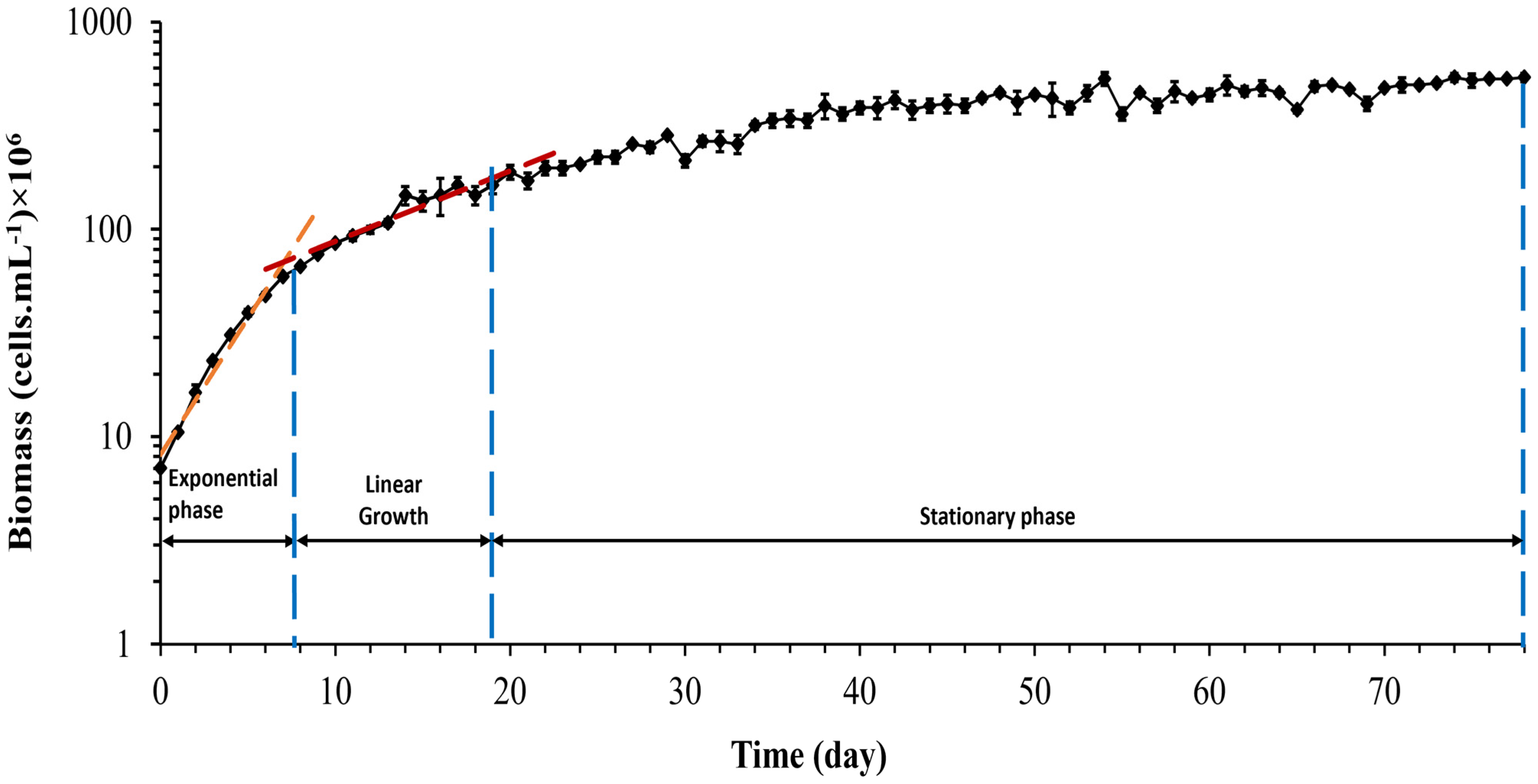
3.2. Growth Curve
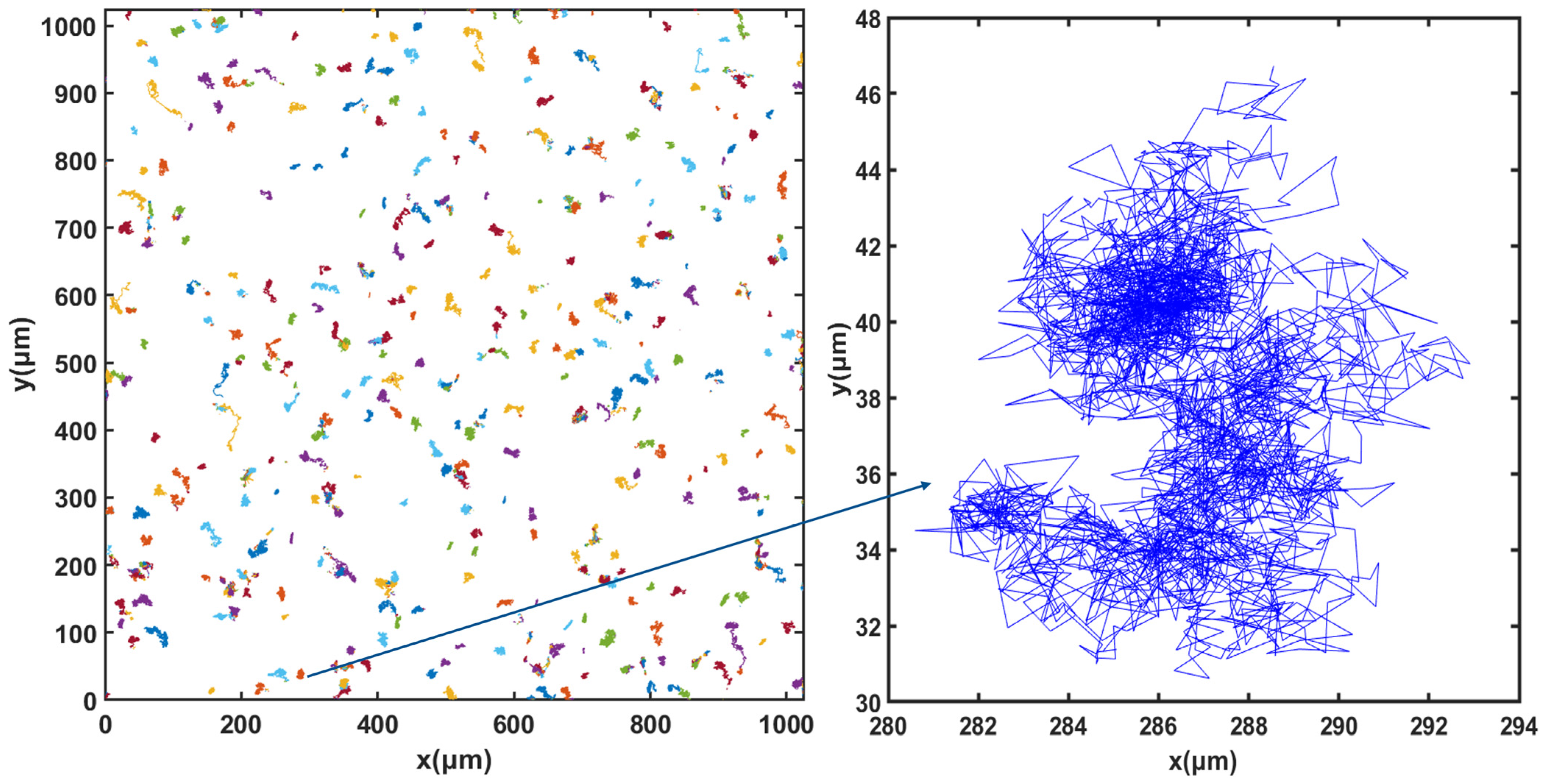
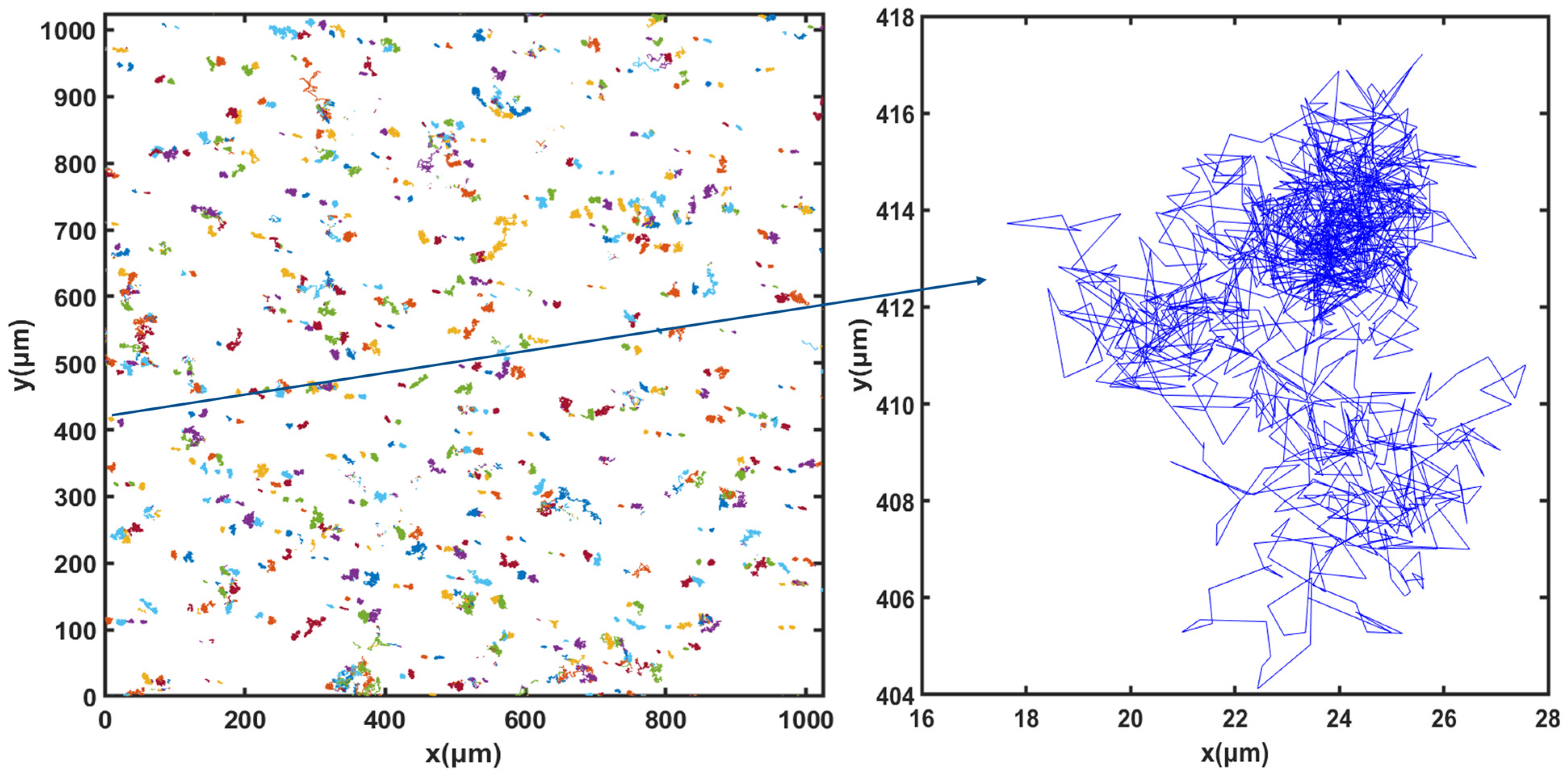
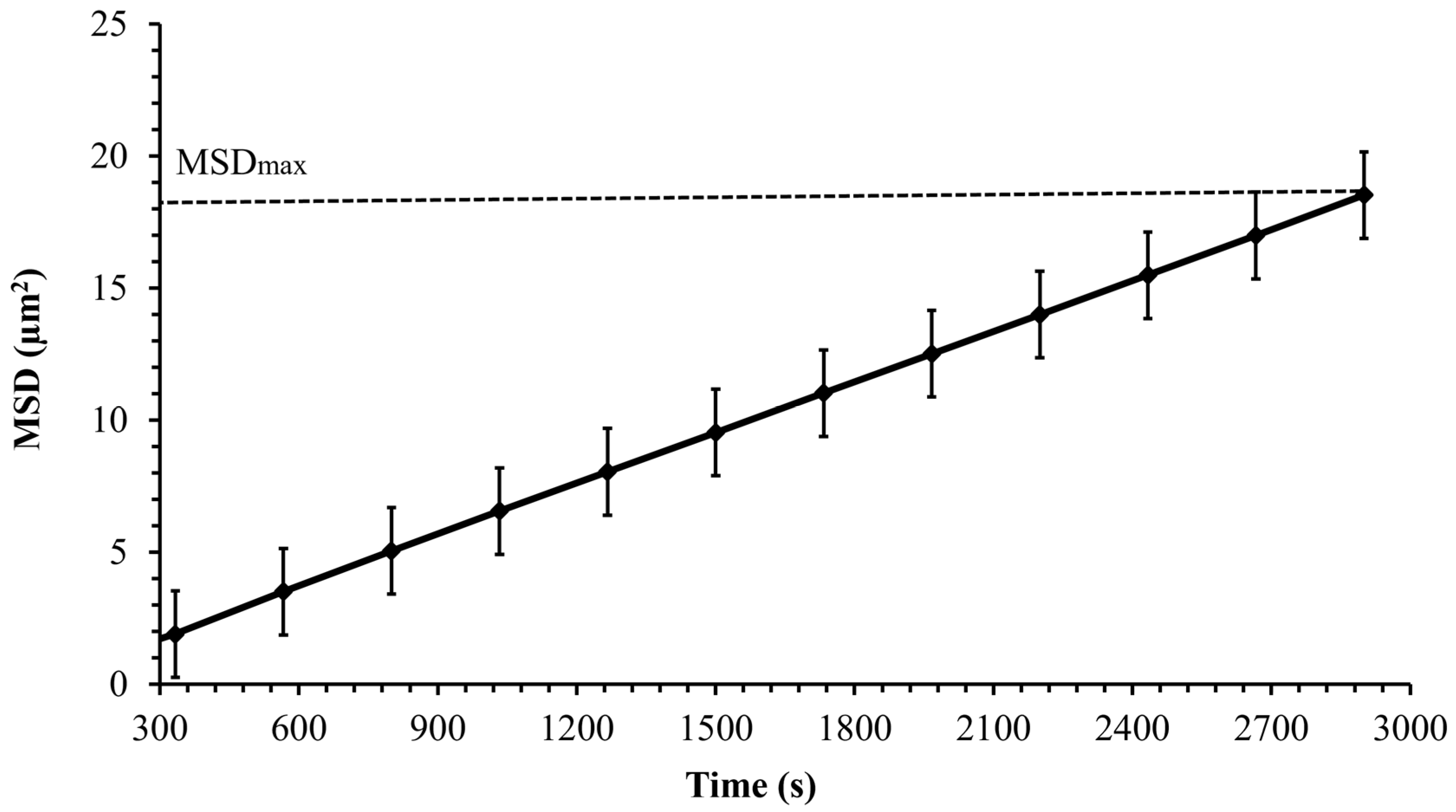
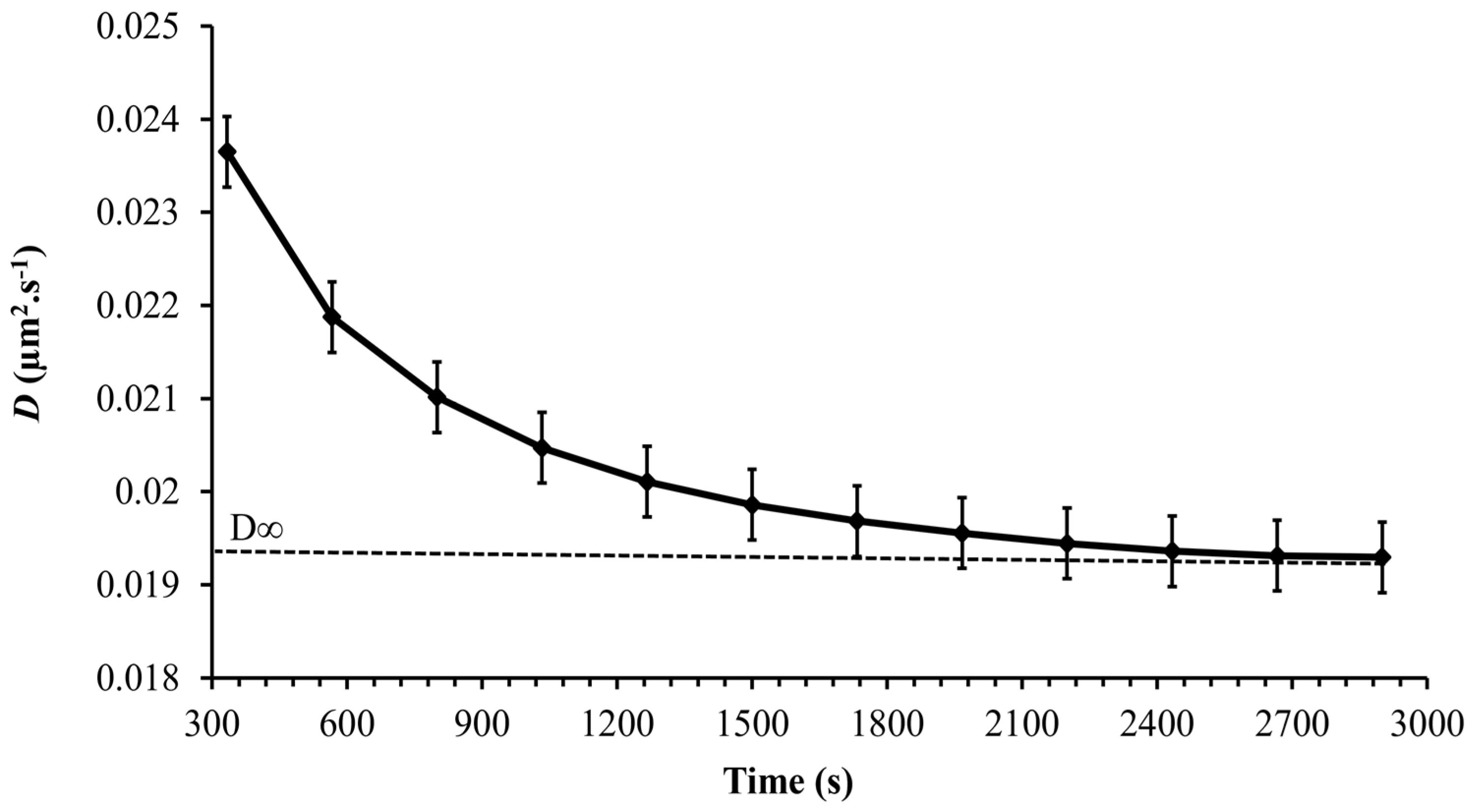
3.3. Cell Motion
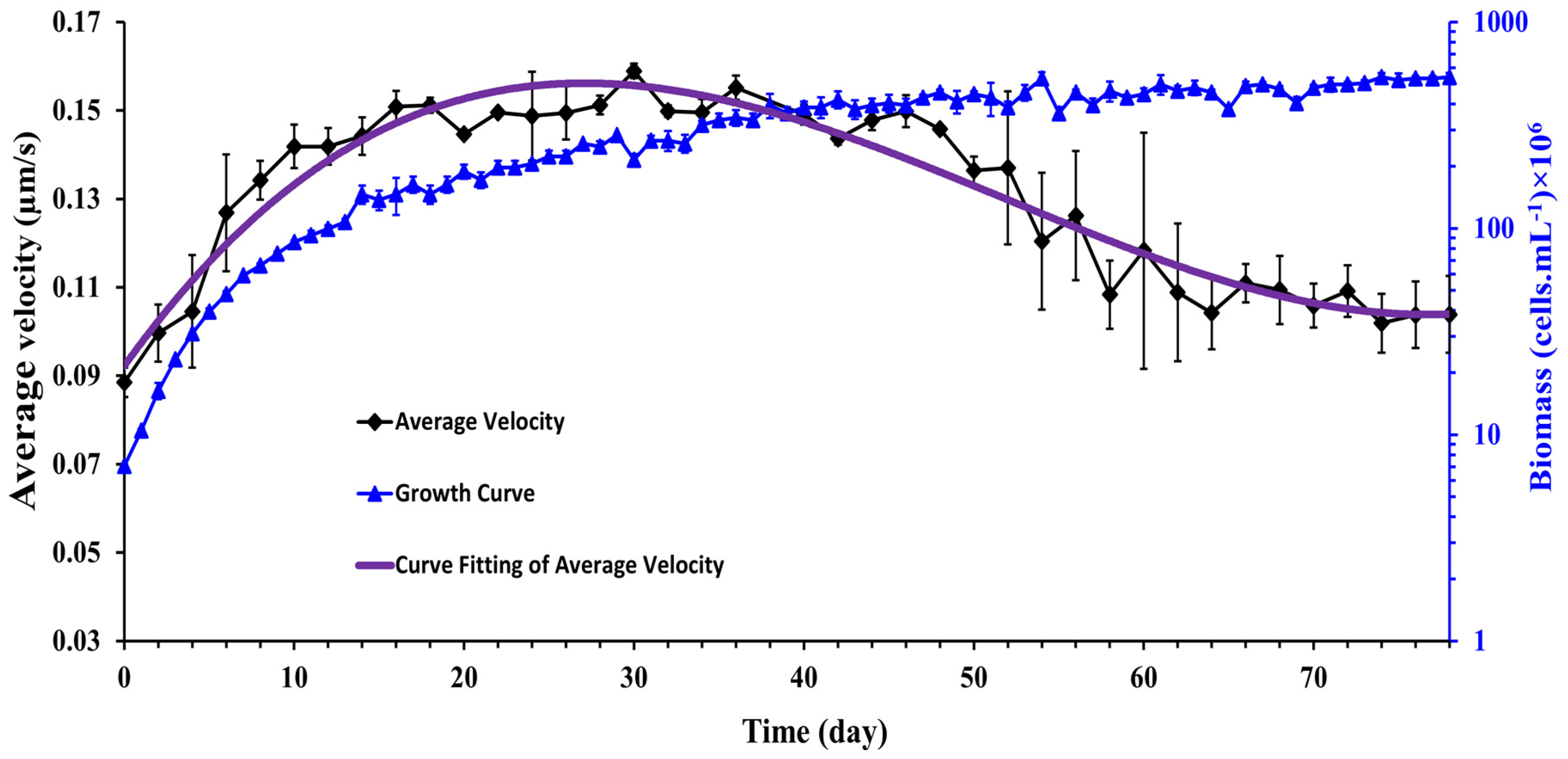
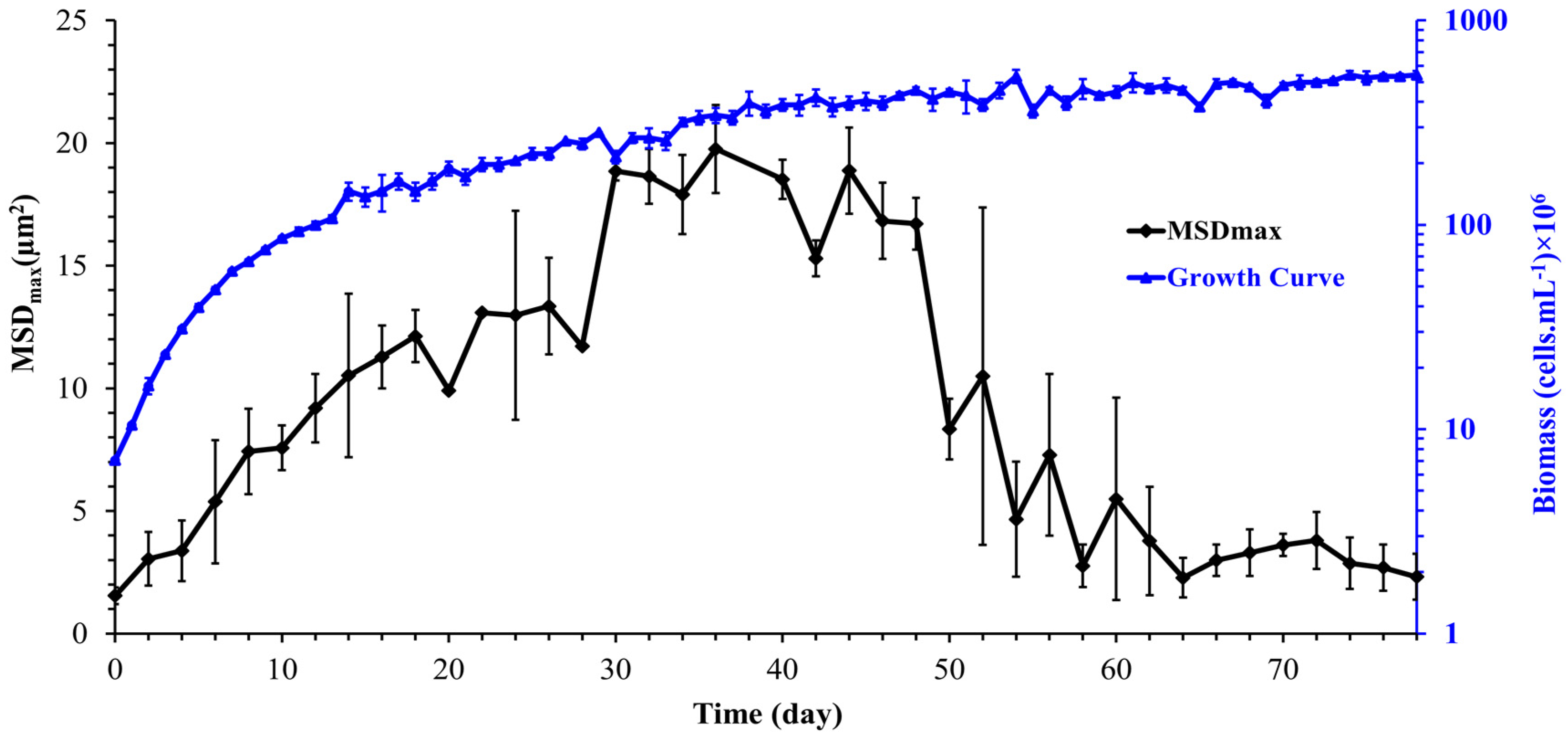
3.4. Evolution of Cell Motility during Synechocystis Growth
- a1 = 8.36 × 10−7 (6.44 × 10−7 to 1.03 × 10−6)
- a2 = −1.31 × 10−4 (−1.54 × 10−4 to −1.09 × 10−4)
- a3 = 5.30 × 10−3 (4.53 × 10−3 to 6.06 × 10−3)
- a4 = 9.25 × 10−2 (8.57 × 10−2 to 9.92 × 10−2)
3.5. Dynamics of Cell Motion during Growth
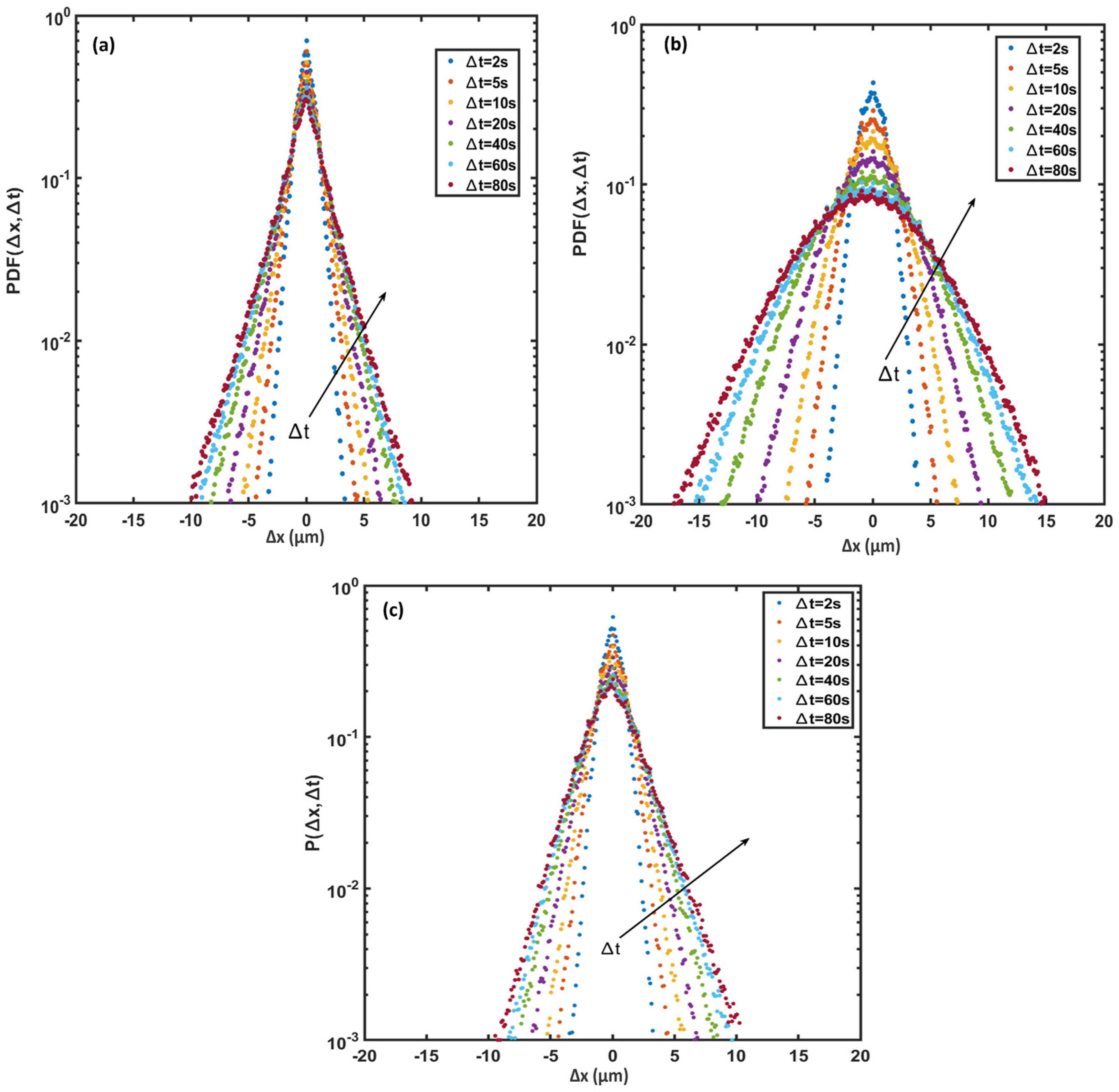
3.6. Probability Density Function (PDF)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| d | Day |
| D | Diffusion coefficient |
| EPS | Exopolysaccharide |
| k | Doubling per day |
| MSD | Mean squared displacement |
| OD | Optical density |
| PBR | Photobioreactor |
| Probability density function | |
| r | Growth rate |
| Vm | Average velocity |
References
- Vourc’h, T.; Léopoldès, J.; Peerhossaini, H. Light control of the diffusion coefficient of active fluids. J. Fluids Eng. 2020, 142, 031109. [Google Scholar] [CrossRef]
- Fadlallah, H.; Jarrahi, M.; Herbert, E.; Ferrari, R.; Mejean, A.; Peerhossaini, H. Active Fluids: Effects of Hydrodynamic Stress on Growth of Self-Propelled Fluid Particles. J. Appl. Fluid Mech. 2020, 13, 561–570. [Google Scholar] [CrossRef]
- Saintillan, D. Rheology of active fluids. Annu. Rev. Fluid Mech. 2018, 50, 563–592. [Google Scholar] [CrossRef]
- Dervaux, J.; Capellazzi Resta, M.; Brunet, P. Light-controlled flows in active fluids. Nat. Phys. 2017, 13, 306–312. [Google Scholar] [CrossRef]
- Mehdizadeh Allaf, M.; Peerhossaini, H. Cyanobacteria: Model microorganisms and beyond. Microorganisms 2022, 10, 696. [Google Scholar] [CrossRef]
- Ortiz, A.; Díez-Montero, R.; García, J.; Khalil, N.; Uggetti, E. Advanced biokinetic and hydrodynamic modelling to support and optimize the design of full-scale high-rate algal ponds. Comput. Struct. Biotechnol. J. 2022, 20, 386–398. [Google Scholar] [CrossRef]
- Solimeno, A.; Parker, L.; Lundquist, T.; García, J. Integral microalgae-bacteria model (BIO_ALGAE): Application to wastewater high rate algal ponds. Sci. Total Environ. 2017, 601, 646–657. [Google Scholar] [CrossRef]
- Hadiyanto, H.; Elmore, S.; Van Gerven, T.; Stankiewicz, A. Hydrodynamic evaluations in high-rate algae pond (HRAP) design. Chem. Eng. J. 2013, 217, 231–239. [Google Scholar] [CrossRef]
- Palma, V.; Gutiérrez, M.S.; Vargas, O.; Parthasarathy, R.; Navarrete, P. Methods to evaluate bacterial motility and its role in bacterial–host interactions. Microorganisms 2022, 10, 563. [Google Scholar] [CrossRef]
- Lauffenburger, D.; Aris, R.; Keller, K. Effects of cell motility and chemotaxis on microbial population growth. Biophys. J. 1982, 40, 209–219. [Google Scholar] [CrossRef]
- Sunda, W.G.; Graneli, E.; Gobler, C.J. Positive feedback and the development and persistence of ecosystem disruptive algal blooms. J. Phycol. 2006, 42, 963–974. [Google Scholar] [CrossRef]
- Vissers, T.; Koumakis, N.; Hermes, M.; Brown, A.T.; Schwarz-Linek, J.; Dawson, A.; Poon, W.C. Dynamical analysis of bacteria in microscopy movies. PLoS ONE 2019, 14, e0217823. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; González Barrios, A.F.; Herzberg, M.; Lee, J. Motility influences biofilm architecture in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Karimi, A.; Karig, D.; Kumar, A.; Ardekani, A.M. Interplay of physical mechanisms and biofilm processes: Review of microfluidic methods. Lab Chip 2015, 15, 23–42. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond risk: Bacterial biofilms and their regulating approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Shoesmith, J.G. The measurement of bacterial motility. Microbiology 1960, 22, 528–535. [Google Scholar] [CrossRef]
- Kim, Y.C. Diffusivity of bacteria. Korean J. Chem. Eng. 1996, 13, 282–287. [Google Scholar] [CrossRef]
- Arora, S.; Bhat, V.; Mittal, A. Correlating single cell motility with population growth dynamics for flagellated bacteria. Biotechnol. Bioeng. 2007, 97, 1644–1649. [Google Scholar] [CrossRef]
- Fadlallah, H.; Peerhossaini, H.; De Groot, C.; Jarrahi, M. Motility Response to Hydrodynamic Stress During the Growth Cycle in Active Fluid Suspensions. J. Fluids Eng. 2021, 143, 074501. [Google Scholar] [CrossRef]
- Bertrand, R.L. Lag phase is a dynamic, organized, adaptive, and evolvable period that prepares bacteria for cell division. J. Bacteriol. 2019, 201, e00697-18. [Google Scholar] [CrossRef]
- Himeoka, Y.; Kaneko, K. Theory for transitions between exponential and stationary phases: Universal laws for lag time. Phys. Rev. X 2017, 7, 021049. [Google Scholar] [CrossRef]
- Jin, D.; Kotar, J.; Silvester, E.; Leptos, K.C.; Croze, O.A. Diurnal variations in the motility of populations of biflagellate microalgae. Biophys. J. 2020, 119, 2055–2062. [Google Scholar] [CrossRef] [PubMed]
- Vourc’h, T.; Peerhossaini, H.; Léopoldès, J.; Méjean, A.; Chauvat, F.; Cassier-Chauvat, C. Slowdown of surface diffusion during early stages of bacterial colonization. Phys. Rev. E 2018, 97, 032407. [Google Scholar] [CrossRef] [PubMed]
- Samadi, Z.; Mehdizadeh Allaf, M.; Saifi, R.; De Groot, C.T.; Peerhossaini, H. Effects of turbulent mixing and orbitally shaking on cell growth and biomass production in active fluids. AJBSR 2022, 15, 396–404. [Google Scholar] [CrossRef]
- Mueller, T.J.; Welsh, E.A.; Pakrasi, H.B.; Maranas, C.D. Identifying regulatory changes to facilitate nitrogen fixation in the nondiazotroph Synechocystis sp. PCC 6803. ACS Synth. Biol. 2016, 5, 250–258. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Tabata, S. Synechocystis sp. PCC 6803—A useful tool in the study of the genetics of cyanobacteria. Photosynth. Res. 2001, 70, 73–83. [Google Scholar] [CrossRef]
- Muth-Pawlak, D.; Kreula, S.; Gollan, P.J.; Huokko, T.; Allahverdiyeva, Y.; Aro, E.M. Patterning of the Autotrophic, Mixotrophic, and Heterotrophic Proteomes of Oxygen-Evolving Cyanobacterium Synechocystis sp. PCC 6803. Front. Microbiol. 2022, 13, 891895. [Google Scholar] [CrossRef]
- Knoop, H.; Gründel, M.; Zilliges, Y.; Lehmann, R.; Hoffmann, S.; Lockau, W.; Steuer, R. Flux balance analysis of cyanobacterial metabolism: The metabolic network of Synechocystis sp. PCC 6803. PLoS Comput. Biol. 2013, 9, e1003081. [Google Scholar] [CrossRef]
- Chau, R.M.; Bhaya, D.; Huang, K.C. Emergent phototactic responses of cyanobacteria under complex light regimes. MBio 2017, 8, e02330-16. [Google Scholar] [CrossRef]
- Conradi, F.D.; Mullineaux, C.W.; Wilde, A. The role of the cyanobacterial type IV pilus machinery in finding and maintaining a favourable environment. Life 2020, 10, 252. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Mehdizadeh Allaf, M.; Habib, Z.; de Bruyn, J.R.; DeGroot, C.T.; Peerhossaini, H. Rheological and Biophysical Properties of Living Fluids Under Shear: Active Suspensions of Synechocystis sp. CPCC 534. J. Fluids Eng. 2022, 144, 021208. [Google Scholar] [CrossRef]
- Wood, A.M.; Everroad, R.C.; Wingard, L.M.; Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005; p. 269. [Google Scholar]
- Berg, H.; Darnton, N.; Jaffe, N. Object Tracking Software; Rowland Institute at Harvard University: Cambridge, MA, USA, 2003. [Google Scholar]
- Samadi, Z.; Mehdizadeh Allaf, M.; Vourc’h, T.; DeGroot, C.T.; Peerhossaini, H. Are Active Fluids Age-Dependent? In Fluids Engineering Division Summer Meeting; American Society of Mechanical Engineers: New York, NY, USA, 2022; Volume 85833, p. V001T03A037. [Google Scholar] [CrossRef]
- Mei, X.; Lee, H.C.; Diao, K.Y.; Huang, M.; Lin, B.; Liu, C.; Xie, Z.; Ma, Y.; Robson, P.M.; Chung, M.; et al. Artificial intelligence–enabled rapid diagnosis of patients with COVID-19. Nat. Med. 2020, 26, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- Leptos, K.C.; Guasto, J.S.; Gollub, J.P.; Pesci, A.I.; Goldstein, R.E. Dynamics of enhanced tracer diffusion in suspensions of swimming eukaryotic microorganisms. Phys. Rev. Lett. 2009, 103, 198103. [Google Scholar] [CrossRef] [PubMed]
- Berg, H.C.; Brown, D.A. Chemotaxis in Escherichia coli Analyzed by Three-Dimensional Tracking. In Chemotaxis: Its Biology and Biochemistry; Karger Publishers: Basel, Switzerland, 1974; Volume 19, pp. 55–78. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samadi, Z.; Mehdizadeh Allaf, M.; Vourc’h, T.; DeGroot, C.T.; Peerhossaini, H. Investigation of Synechocystis sp. CPCC 534 Motility during Different Stages of the Growth Period in Active Fluids. Processes 2023, 11, 1492. https://doi.org/10.3390/pr11051492
Samadi Z, Mehdizadeh Allaf M, Vourc’h T, DeGroot CT, Peerhossaini H. Investigation of Synechocystis sp. CPCC 534 Motility during Different Stages of the Growth Period in Active Fluids. Processes. 2023; 11(5):1492. https://doi.org/10.3390/pr11051492
Chicago/Turabian StyleSamadi, Zahra, Malihe Mehdizadeh Allaf, Thomas Vourc’h, Christopher T. DeGroot, and Hassan Peerhossaini. 2023. "Investigation of Synechocystis sp. CPCC 534 Motility during Different Stages of the Growth Period in Active Fluids" Processes 11, no. 5: 1492. https://doi.org/10.3390/pr11051492
APA StyleSamadi, Z., Mehdizadeh Allaf, M., Vourc’h, T., DeGroot, C. T., & Peerhossaini, H. (2023). Investigation of Synechocystis sp. CPCC 534 Motility during Different Stages of the Growth Period in Active Fluids. Processes, 11(5), 1492. https://doi.org/10.3390/pr11051492