

Denaturation and Digestion Increase the Antioxidant Capacity of Proteins




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. ABTS● Decolorization Assay
2.3. FRAP Assay
2.4. CUPRAC Assay
2.5. Protein Denaturation
2.6. Protein Digestion
2.7. Statistics
3. Results
3.1. Amino Acids Contributing to the Antioxidant Activity/Capacity of Proteins
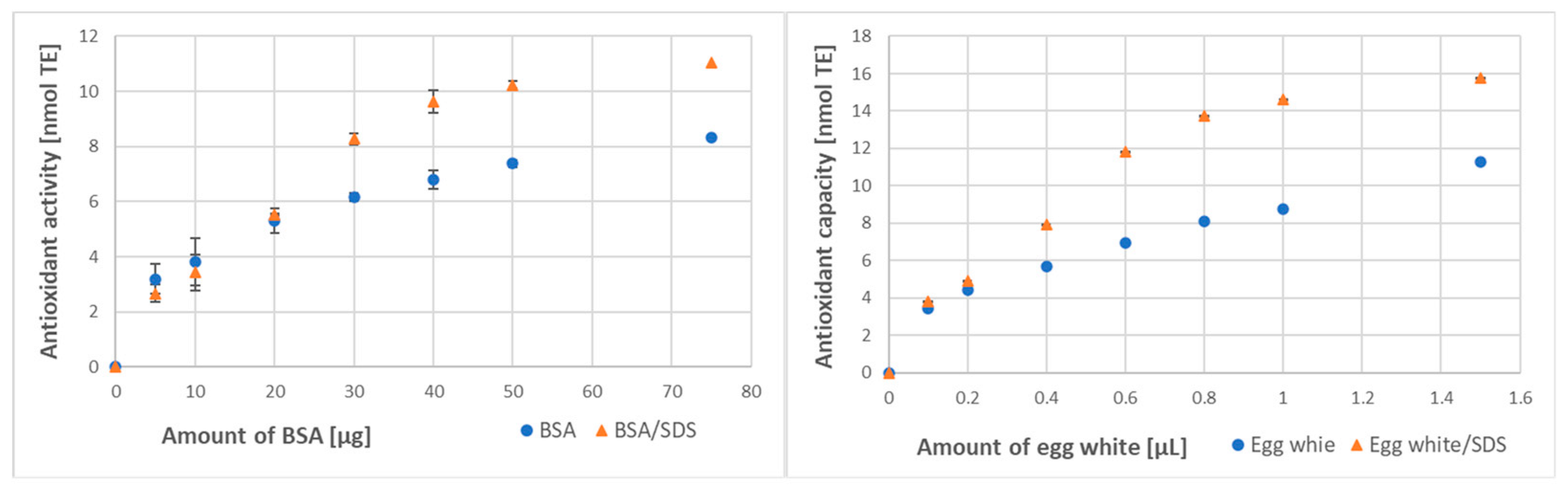
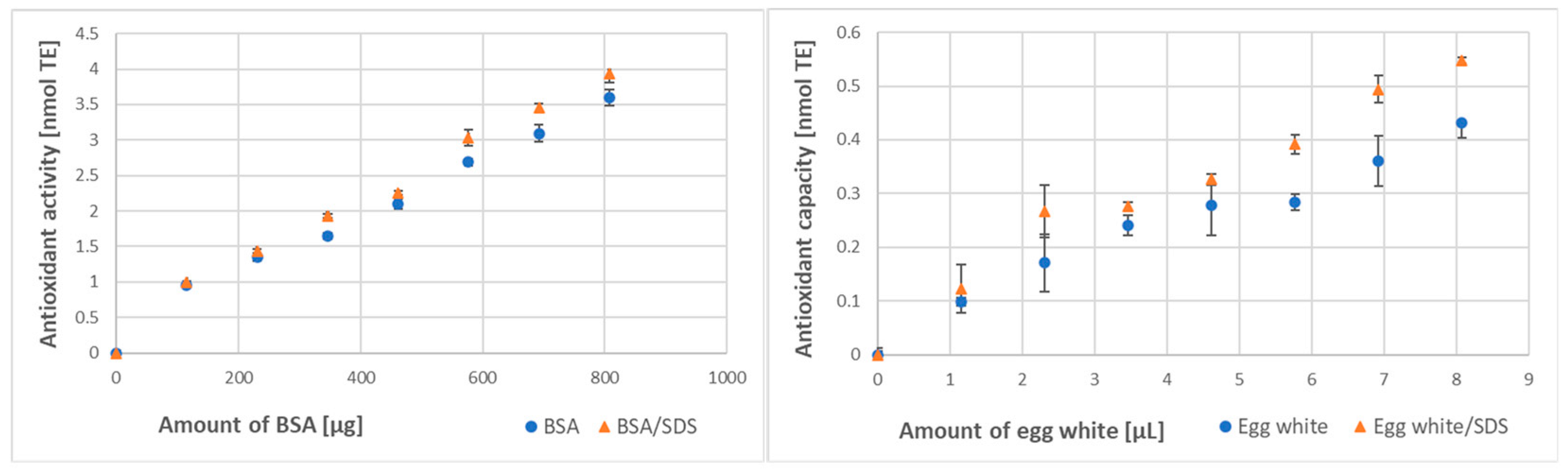
3.2. Effect of Denaturation on Protein Antioxidant Activity/Capacity
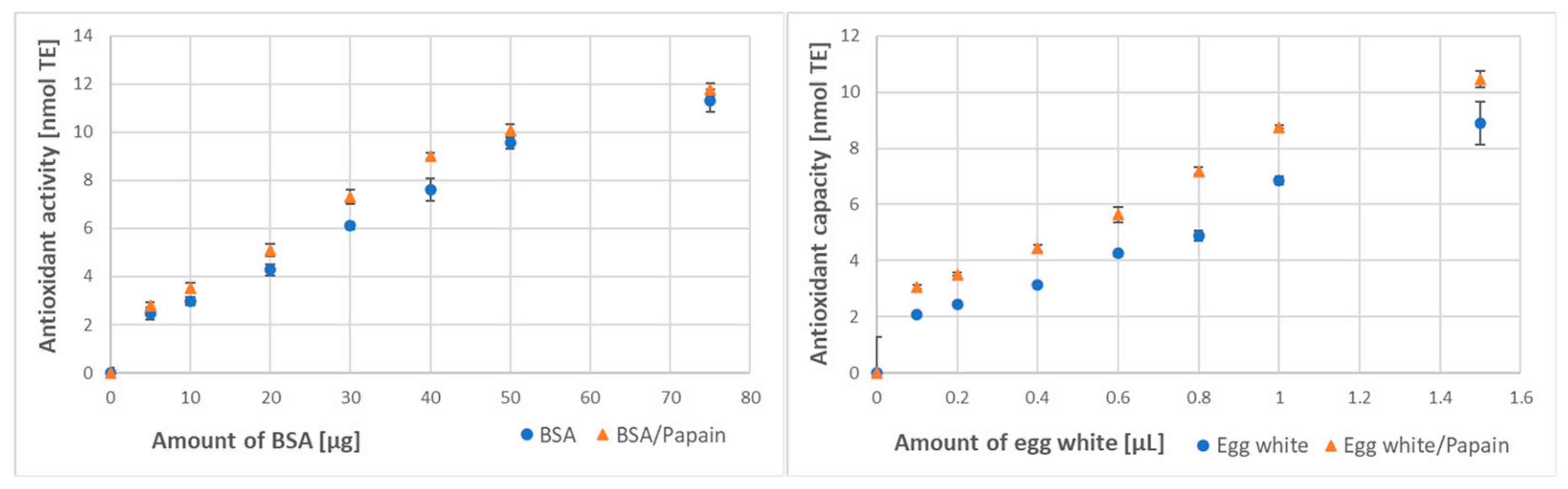
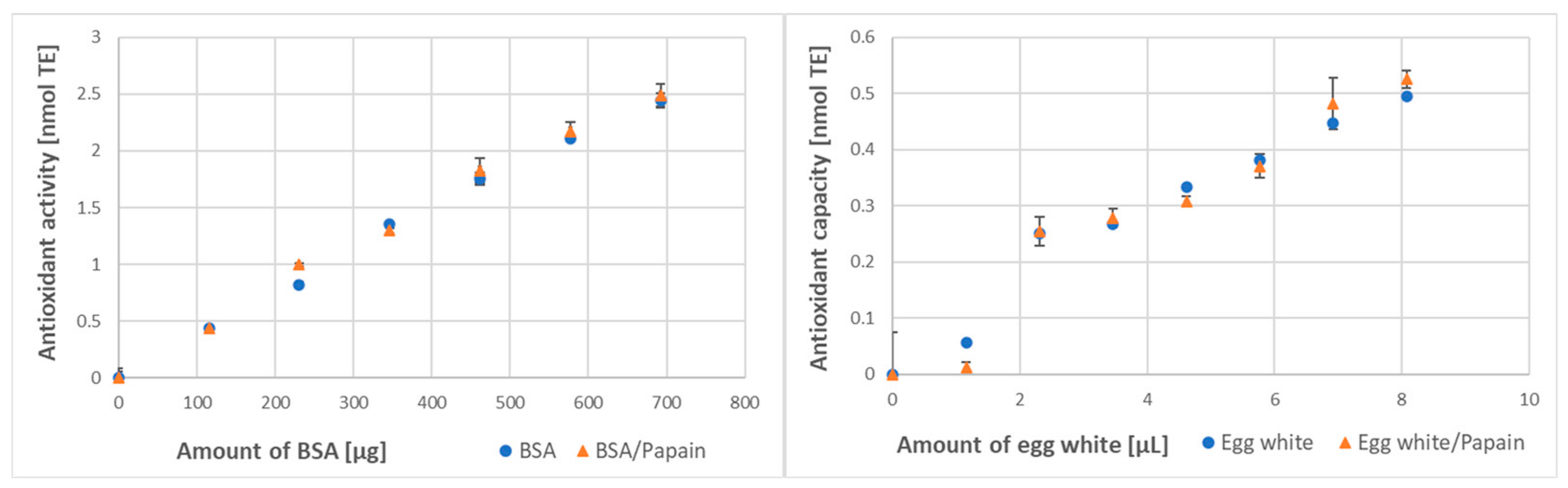
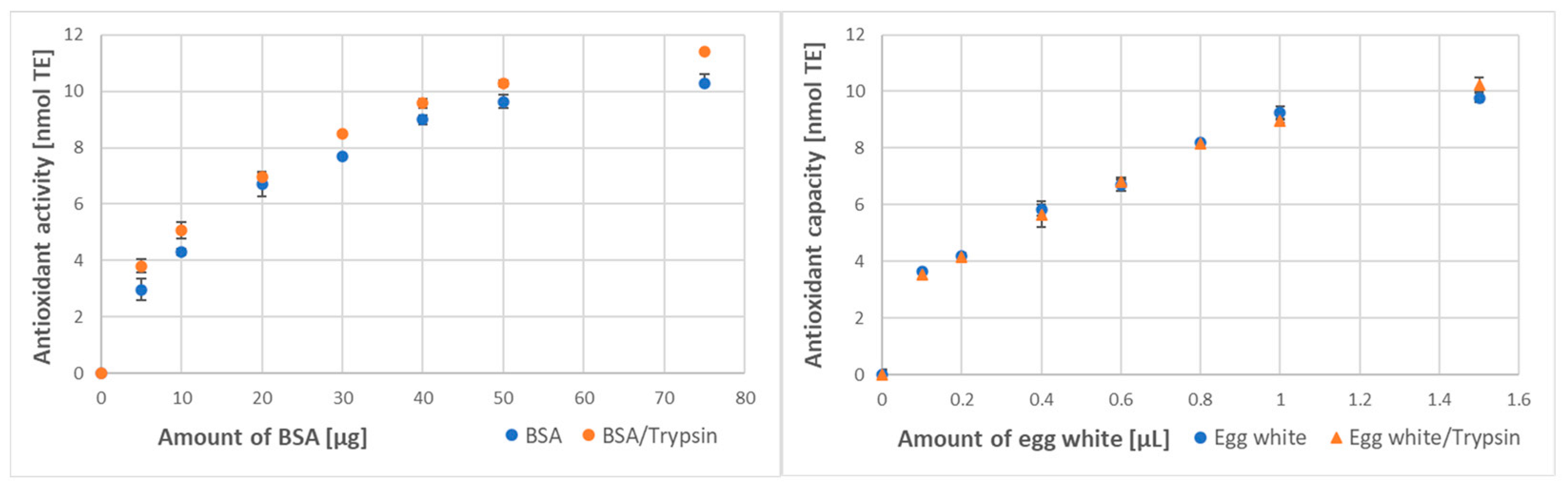
3.3. Effect of Digestion on the Protein Antioxidant Activity/Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, X.; Gu, L.; Holden, J.; Haytowitz, D.B.; Gebhardt, S.E.; Beecher, G.; Prior, R.L. Development of a database for total antioxidant capacity in foods: A preliminary study. J. Food Composit. Anal. 2004, 17, 407–422. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Pellegrini, N.; Vitaglione, P.; Granato, D.; Fogliano, V. Twenty-five years of total antioxidant capacity measurement of foods and biological fluids: Merits and limitations. J. Sci. Food Agric. 2020, 100, 5064–5078. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Composit. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Song, W.O.; Fernandez, M.L.; Bruno, R.S.; Koo, S.I.; Chun, O.K. Development and validation of an algorithm to establish a total antioxidant capacity database of the US diet. Int. J. Food Sci. Nutr. 2010, 61, 600–623. [Google Scholar] [CrossRef] [PubMed]
- Pompella, A.; Sies, H.; Wacker, R.; Brouns, F.; Grune, T.; Biesalski, H.K.; Frank, J. The use of total antioxidant capacity as surrogate marker for food quality and its effect on health is to be discouraged. Nutrition 2014, 30, 791–793. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X. Diet Antioxidant Capacity: Relationships to Oxidative Stress and Health. Am. J. Biomed. Sci. 2013, 5, 126–139. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Alvarez, J.A.; Fernández-López, J. In vitro digestion models suitable for foods: Opportunities for new fields of application and challenges. Food Res. Int. 2018, 107, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Soymilk phenolic compounds, isoflavones and antioxidant activity as affected by in vitro gastrointestinal digestion. Food Chem. 2013, 136, 206–212. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Delgado-Andrade, C. Effect of digestive process on Maillard reaction indexes and antioxidant properties of breakfast cereals. Food Res. Int. 2009, 42, 394–400. [Google Scholar] [CrossRef]
- Ng, Z.X.; Rosman, N.F. In vitro digestion and domestic cooking improved the total antioxidant activity and carbohydrate-digestive enzymes inhibitory potential of selected edible mushrooms. J. Food Sci. Technol. 2019, 56, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In Vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols-A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Reginio, F.C., Jr.; Thuengtung, S.; Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 4684–4705. [Google Scholar] [CrossRef]
- Koehnlein, E.A.; Koehnlein, É.M.; Corrêa, R.C.G.; Nishida, V.S.; Correa, V.G.; Bracht, A.; Peralta, R.M. Analysis of a whole diet in terms of phenolic content and antioxidant capacity: Effects of a simulated gastrointestinal digestion. Int. J. Food Sci. Nutr. 2016, 67, 614–623. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S.; Rupasinghe, H.P.V. Change of phenolics, carotenoids, and antioxidant capacity following simulated gastrointestinal digestion and dialysis of selected edible green leaves. Food Chem. 2018, 245, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Faller, A.L.; Fialho, E.; Liu, R.H. Cellular antioxidant activity of feijoada whole meal coupled with an in vitro digestion. J. Agric. Food Chem. 2012, 60, 4826–4832. [Google Scholar] [CrossRef]
- Corrochano, A.R.; Sariçay, Y.; Arranz, E.; Kelly, P.M.; Buckin, V.; Giblin, L. Comparison of antioxidant activities of bovine whey proteins before and after simulated gastrointestinal digestion. J. Dairy Sci. 2019, 102, 54–67. [Google Scholar] [CrossRef]
- Delgado, M.C.O.; Galleano, M.; Añón, M.C.; Tironi, V.A. Amaranth peptides from simulated gastrointestinal digestion: Antioxidant activity against reactive species. Plant Foods Hum. Nutr. 2015, 70, 27–34. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. Changes in the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates during a simulated gastrointestinal digestion. Food Chem. 2010, 120, 810–816. [Google Scholar] [CrossRef]
- Ahn, C.B.; Kim, J.G.; Je, J.Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef]
- Navajas-Porras, B.; Pérez-Burillo, S.; Valverde-Moya, Á.; Hinojosa-Nogueira, D.; Pastoriza, S.; Rufián-Henares, J.Á. Effect of Cooking Methods on the Antioxidant Capacity of Foods of Animal Origin Submitted to In Vitro Digestion-Fermentation. Antioxidants 2021, 10, 445. [Google Scholar] [CrossRef]
- Navajas-Porras, B.; Pérez-Burillo, S.; Valverde-Moya, Á.J.; Hinojosa-Nogueira, D.; Pastoriza, S.; Rufián-Henares, J.Á. Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion-Fermentation. Antioxidants 2020, 9, 1312. [Google Scholar] [CrossRef] [PubMed]
- Kut, K.; Cieniek, B.; Stefaniuk, I.; Bartosz, G.; Sadowska-Bartosz, I. A Modification of the ABTS• Decolorization Method and an Insight into Its Mechanism. Processes 2022, 10, 1288. [Google Scholar] [CrossRef]
- Bartosz, G. Total antioxidant capacity. Adv. Clin. Chem. 2003, 37, 219–292. [Google Scholar] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Özyürek, M.; Güçlü, K.; Tütem, E.; Başkan, K.S.; Erçağ, E.; Çelik, S.E.; Baki, S.; Leyla, Y.; Karaman, S.; Apak, R. A comprehensive review of CUPRAC methodology. Anal. Meth. 2011, 3, 2439–2453. [Google Scholar] [CrossRef]
- Finley, J.W.; Wheeler, E.L.; Walker, H.G., Jr.; Finlayson, A.J. Effect of cystine oxidation on lysinoalanine formation in proteins. J. Agric. Food Chem. 1982, 30, 818–820. [Google Scholar] [CrossRef]
- Karimi, M.; Crossett, B.; Cordwell, S.J.; Pattison, D.I.; Davies, M.J. Characterization of disulfide (cystine) oxidation by HOCl in a model peptide: Evidence for oxygen addition, disulfide bond cleavage and adduct formation with thiols. Free Radic. Biol. Med. 2020, 154, 62–74. [Google Scholar] [CrossRef]
- Yi, H.; Cheng, Y.; Zhang, Y.; Xie, Q.; Yang, X. Potentiometric and UV-Vis spectrophotometric titrations for evaluation of the antioxidant capacity of chicoric acid. RSC Adv. 2020, 10, 11876–11882. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef]
- Atanasiu, R.L.; Stea, D.; Mateescu, M.A.; Vergely, C.; Dalloz, F.; Briot, F.; Maupoil, V.; Nadeau, R.; Rochette, L. Direct evidence of caeruloplasmin antioxidant properties. Mol. Cell. Biochem. 1998, 189, 127–135. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Maiani, G.; Azzini, E.; Ferro-Luzzi, A. A fluorescence-based method for measuring total plasma antioxidant capability. Free Radic. Biol. Med. 1995, 18, 29–36. [Google Scholar] [CrossRef]
- Cui, J.; Chong, B.; Rutherfurd, S.M.; Wilkinson, B.; Singh, H.; Moughan, P.J. Gross and true ileal digestible amino acid contents of several animal body proteins and their hydrolysates. Meat Sci. 2013, 94, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Siriwardana, K.; Wang, A.; Gadogbe, M.; Collier, W.E.; Fitzkee, N.C.; Zhang, D. Studying the effects of cysteine residues on protein interactions with silver nanoparticles. J. Phys. Chem. C 2015, 119, 2910–2916. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.B.; Saroff, H.A. Decrease in Sulfhydryl Titer of Serum Albumin1. J. Am. Chem. Soc. 1958, 80, 2129–2131. [Google Scholar] [CrossRef]
- Michnik, A.; Michalik, K.; Drzazga, Z. Stability of bovine serum albumin at different pH. J. Therm. Anal. Calorim. 2005, 80, 399–406. [Google Scholar] [CrossRef]
- Robic, S.; Linscott, K.B.; Aseem, M.; Humphreys, E.A.; McCartha, S.R. Bile acids as modulators of enzyme activity and stability. Protein J. 2011, 30, 539–545. [Google Scholar] [CrossRef]
- Reynolds, J.A.; Tanford, C. Binding of dodecyl sulfate to proteins at high binding ratios. Possible implications for the state of proteins in biological membranes. Proc. Natl. Acad. Sci. USA 1970, 66, 1002–1007. [Google Scholar] [CrossRef]
- Mann, K. The chicken egg white proteome. Proteomics 2007, 7, 3558–3568. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, Y.Q. An insight on egg white: From most common functional food to biomaterial application. J. Biomed. Materials Res. B Appl. Biomater. 2021, 109, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.M.; Andersen, K.K.; Westh, P.; Otzen, D.E. Unfolding of beta-sheet proteins in SDS. Biophys. J. 2007, 92, 674–685. [Google Scholar] [CrossRef]
- van der Hoorn, R.A. Plant proteases: From phenotypes to molecular mechanisms. Annu. Rev. Plant Biol. 2008, 59, 191–223. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.F.; Zanette, D.; Fischer, H.; Itri, R. A systematic study of bovine serum albumin (BSA) and sodium dodecyl sulfate (SDS) interactions by surface tension and small angle X-ray scattering. J. Coll. Interface Sci. 2003, 262, 400–408. [Google Scholar] [CrossRef]
- Rao, P.S.; Nolasco, E.; Handa, A.; Naldrett, M.J.; Alvarez, S.; Majumder, K. Effect of pH and Heat Treatment on the Antioxidant Activity of Egg White Protein-Derived Peptides after Simulated In-Vitro Gastrointestinal Digestion. Antioxidants 2020, 9, 1114. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid/Assay | ABTS• Decolorization [mol TE/mol] | FRAP [mol TE/mol] | CUPRAC [mol TE/mol] |
|---|---|---|---|
| Cysteine | 2.07 ± 0.24 | 0.725 ± 0.065 | 1.775 ± 0.110 |
| Tyrosine | 4.07 ± 0.86 | 0.260 ± 0.022 | 1.656 ± 0.215 |
| Tryptophan | 3.32 ± 0.52 | 0.385 ± 0.007 | 0.401 ± 0.042 |
| Cystine | 0.15 ± 0.02 | 0.045 ± 0.003 | No reaction |
| Histidine | 0.50 ± 0.10 | No reaction | No reaction |
| Arginine | 0.40 ± 0.04 | No reaction | No reaction |
| Method of Assay | BSA Antioxidant Activity [μmol/g] | Egg White Antioxidant Capacity [μmol/mL] | ||
|---|---|---|---|---|
| Control | Treated | Control | Treated | |
| SDS denaturation | ||||
| ABTS● decolorization | 174 ± 12 | 250 ± 23 ** (144%) | 6.82 ± 0.09 | 11.39 ± 1.89 *** (167%) |
| FRAP | 35.2 ± 2.6 | 40.8 ± 3.0 * (116%) | 48.6 ± 4.5 | 63.1 ± 7.2 * (130%) |
| Papain digestion | ||||
| ABTS● decolorization | 173 ± 14 | 216 ± 10 * (125%) | 6.91 ± 0.55 | 8.71 ± 0.86 * (126%) |
| FRAP | 36.7 ± 2.1 | 41.2 ± 1.8 * (112%) | 46.3 ± 2.1 | 53.2 ± 1.5 * (115%) |
| Trypsin digestion | ||||
| ABTS● decolorization | 176 ± 11 | 199 ± 10 * (113%) | 6.79 ± 0.51 | 7.10 ± 0.38 NS (105%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kut, K.; Bartosz, G.; Sadowska-Bartosz, I. Denaturation and Digestion Increase the Antioxidant Capacity of Proteins. Processes 2023, 11, 1362. https://doi.org/10.3390/pr11051362
Kut K, Bartosz G, Sadowska-Bartosz I. Denaturation and Digestion Increase the Antioxidant Capacity of Proteins. Processes. 2023; 11(5):1362. https://doi.org/10.3390/pr11051362
Chicago/Turabian StyleKut, Kacper, Grzegorz Bartosz, and Izabela Sadowska-Bartosz. 2023. "Denaturation and Digestion Increase the Antioxidant Capacity of Proteins" Processes 11, no. 5: 1362. https://doi.org/10.3390/pr11051362
APA StyleKut, K., Bartosz, G., & Sadowska-Bartosz, I. (2023). Denaturation and Digestion Increase the Antioxidant Capacity of Proteins. Processes, 11(5), 1362. https://doi.org/10.3390/pr11051362