Abstract
Although immunotherapy has become the standard of treatment for non-small cell lung cancer (NSCLC), only a limited number of patients benefit from it clinically because of the resistance to immunotherapeutic agents. Of late, the usefulness of herbal medicines in combination with immunotherapy has been investigated. Bojungikki-Tang (BJIKT) is a widely used traditional herbal medicine. It synergistically enhances the antitumor effects of chemotherapy and regulates the immune responses in cancer, but its antitumor effect with immunotherapy in NSCLC is unclear. In this study, we investigated the combined effects of BJIKT and an anti-PD-1 antibody in a KLN205-DBA/2 syngeneic lung cancer model. Immunohistochemistry and flow cytometry analyses were performed to analyze the changes in immune cells in the tumor microenvironment. BJIKT plus an anti-PD-1 antibody treatment significantly inhibited tumor growth, unlike the respective monotherapies. Compared to monotherapy, the combination treatment resulted in a higher population of CD8+ cytotoxic T cells and a lower number of Ki67+ cells in the tumor tissues. Furthermore, the combination treatment decreased the proportion of myeloid-derived suppressor cells but increased the proportion of M1-like macrophages compared to that observed with monotherapy. Cytokine analysis showed that the combination treatment increased the levels of T helper type 1-related cytokines. Network pharmacology analysis revealed that BJIKT might regulate multiple signaling pathways related to immune function and tumor progression in NSCLC. These findings indicate that the combination treatment with BJIKT and an anti-PD-1 antibody effectively suppresses tumor growth by regulating immune function and may be an alternative therapeutic option for the treatment of NSCLC.
1. Introduction
Lung cancer is the most prevalent malignancy worldwide. Non-small cell lung cancer (NSCLC) accounts for more than 85% of all lung cancer cases. Despite the marked progress in treatment methods for NSCLC over the past decade, it is still a major cause of cancer-related deaths [1]. Chemotherapy, radiotherapy, and targeted therapy are commonly used for treating metastatic NSCLCs. However, the emergence of resistance to chemotherapeutic agents poses a significant obstacle to the management of patients with NSCLC, demonstrating the necessity for developing other treatment options for refractory NSCLC [2]. Immune checkpoint inhibitors (ICIs) have been a significant breakthrough in the field of cancer research and have revolutionized the management of NSCLC. ICIs have shown potent activity against NSCLC in clinical trials [3]. However, they have a relatively low response rate and can cause hyperprogression of the disease. ICIs are more effective against immunogenic tumors associated with a sufficient number of tumor-infiltrating lymphocytes and the increased expression of the PD-1 ligand (PD-L1) [4].
The response to ICIs, especially ones targeting the PD-L1/PD-1 axis, depends on the cancer immunity cycle, which results in an increase in the number of tumor antigen-specific effector T cells [5]. There are various roadblocks in the tumor microenvironment (TME) that hamper the development and activation of effector T cells [6]. The main roadblock is immunosuppressive myeloid cells in tumors. This population of cells includes myeloid-derived suppressor cells (MDSCs), tumor-associated macrophages, regulatory T cells, and immature dendritic cells [7]. They express several cell surface receptors that stimulate their recruitment into the TME and promote immunosuppressive cytokine production. Therefore, multi-target combination treatments with chemotherapeutic agents, vaccines, and immunotherapies are promising strategies to overcome the low response of ICIs and increase the number of patients who benefit from PD-1/PD-L1 blockade [8].
Traditional Chinese medicine (TCM) has been a crucial part of healthcare for centuries in East Asia, including in China, Japan, and Korea. Bojungikki-Tang (BJIKT), a herbal medicine with 10 component herbs, is a well-known formula in TCM. It has been widely used for the treatment of general fatigues, the lack of appetite, and digestive disorders [9]. In particular, clinical trials demonstrated that BJIKT improved the cancer-related symptoms, such as fatigue, cachexia, and anorexia [10,11]. Moreover, experimental studies indicated that BJIKT synergistically enhances the antitumor effects of other drugs [12] and regulates the immune responses in cancer [13,14]. Recently, we have shown that BJIKT significantly improves the response to anti-PD-L1 immunotherapy and prolongs survival in colorectal tumor-bearing mice [15]. Although the ability of BJIKT to increase the therapeutic effects of anticancer drugs has been proven, the exact mechanisms of action of BJIKT plus ICIs in NSCLC are unclear. Therefore, in the present study, we investigated the therapeutic effects of the combination of BJIKT plus an anti-PD-1 antibody in the KLN205 lung cancer model. We also investigated their effects on various immune cells in tumors. Our findings will provide evidence for the antitumor effects of BJIKT in combination with an anti-PD-1 antibody against NSCLC.
2. Materials and Methods
2.1. Chemicals and Reagents
The BJIKT extract was manufactured by Hanpoong Pharmaceutical Co. (Jeonju, Korea) using herbal medicines certified by the Korea Ministry of Food and Drug Safety in accordance with the standardized manufacturing methods in Good Manufacturing Practice facilities. BJIKT is a water extract composed of 10 herbs: Astragalus membranaceus (Fisch.) Bunge (16.6%), Atractylodes macrocephala Koidz. (16.6%), Panax ginseng C. A. Mey. (16.6%), Angelica gigas Nakai (12.5%), Ziziphus jujuba var. inermis (Bunge) Rehder (8.4%), Bupleurum falcatum L. (8.4%), Citrus unshiu Marcow. (8.4%), Glycyrrhiza glabra L. (6.3%), Cimicifuga heracleifolia Kom. (4.1%), and Zingiber officinale Roscoe (2.1%). BJIKT was prepared as reported [15]. In brief, 10 herbs were extracted using water under reflux conditions at 95 °C. Then, the extract solution was filtered, evaporated under low pressure, and lyophilized in a freeze dryer. The dried extract was evaluated and passed according to the standards and test methods of Bojungikgitang Soft/Dried Extract in the Korean Herbal Pharmacopoeia. The contents of ginsenoside Rb1 and glycyrrhizic acid in the BJIKT extract were 0.77 mg/g and 5.66 mg/g, respectively, which met the KHP ingredients content standards. Eagle’s minimum essential medium (EMEM) was purchased from ATCC (Manassas, VA, USA). Dulbecco’s phosphate-buffered saline and fetal bovine serum (FBS) were obtained from Gibco (Grand Island, NY, USA). Matrigel was purchased from Corning Inc. (Corning, NY, USA). Neutralized antibody anti-mouse PD-1 (RMP1-14) and isotype control IgG2b antibody were obtained from BioXCell (West Lebanon, NH, USA). An anti-CD3 antibody (#NB600-1441) was purchased from Novus Biologicals (Littleton, CO, USA). Anti-CD8 (#98941) and anti-F4/80 (#70076) antibodies were purchased from Cell signaling (Beverly, MA, USA). Anti-CD4 (#183685), anti-granzyme B (#4059), and anti-Ki67 (#15580) antibodies were purchased from Abcam (Cambridge, MA, USA). APC/Cy7 anti-CD45 (#103116), PE/Cy7 anti-CD3 (#552774), BV711 anti-CD4 (#563050), BV605 anti-CD8 (#563152), Alexa647 anti-CD335 (NKp46) (#137608), BV421 anti-CD11b (#562605), PE/Cy7 anti-F4/80 (#123114), PE anti-CD206 (#141706), Alexa488 anti-MHC Class II (#107616), APC anti-GR-1 (#108412), and TruStain FcX™ PLUS (#156604) antibodies were purchased from Biolegend (San Diego, CA, USA).
2.2. Cell Culture
KLN205, a murine squamous lung cancer cell line, was purchased from ATCC (CRL-1453). The cells were cultured in EMEM containing 10% FBS, penicillin (100 U/mL), and streptomycin (100 μg/mL) and maintained at 37 °C in a humidified incubator with 5% CO2 and 95% air; the cells were confirmed to be free of mycoplasma contamination by using MycoStrip™ (InvivoGen, San Diego, CA, USA) before use.
2.3. Animals
Specific, pathogen-free, 6-week-old male DBA/2 mice were purchased from Orient Bio (Seongnam, Korea) and acclimated for 1 week before experimental use. The mice were housed under the following environmental conditions: temperature, 23 °C; humidity, 50%; and a 12 h light/dark cycle. The animals were fed a standard chow diet (Purina Co., Seoul, Korea) and provided with drinking water ad libitum. All experimental procedures were approved by the Animal Care and Use Committee of the Korea Institute of Oriental Medicine (Approval number: 21-038).
2.4. Establishment of Tumor-Bearing Mice and Treatment
KLN205 tumor-bearing mice were established by subcutaneously injecting 1 × 105 KLN205 cells in 100 μL EMEM containing 50% Matrigel into the right flank of DBA/2 mice. When the tumor volume reached an average of 85 mm3, the DBA/2 mice were evenly divided into the following treatment groups (five mice per group): the control group, BJIKT group, anti-PD-1 antibody group, and combination group (BJIKT and anti-PD-1 antibody). BJIKT was dissolved in distilled water and orally administered at 450 mg/kg daily for 19 days. The dose was calculated based on the human equivalent dose determined as per the dose conversion guide [16]. A dose of 200 µg of the anti-PD-1 antibody was intraperitoneally administered every three days. The IgG2b antibody was used as the isotype control. Tumor sizes were measured three times a week using a digital caliper, and the tumor volume was calculated using the following formula: tumor volume = (length × width2)/2. The body weight was monitored to determine levels of toxicity.
2.5. Immunohistochemistry (IHC)
IHC was performed following standard techniques. Briefly, the paraffin-embedded tumor sections (4 μm thick) were incubated in 3% H2O2 for 10 min after boiling in Tris–EDTA antigen retrieval buffers, blocked with Super Block (AAA125, ScyTek, UT, USA), and incubated overnight at 4 °C with the primary antibody, including CD3 (1:500), CD8 (1:800), CD4 (1:750), granzyme B (1:300), F4/80 (1:600), and Ki67 (1:600). Then, the secondary antibody reaction was performed using the Polink-2 HRP Plus Rabbit DAB Detection System (GBI Labs, Bothell, WA, USA) at room temperature. Finally, the sections were mounted on gelatin-coated slides, dehydrated, and cover-slipped using the Leica MM24 mounting medium (Leica Biosystems, Richmond, IL, USA). Their levels were assessed by the microscopic examination of the IHC-stained sections. Quantification was performed using the ImageJ software.
2.6. Flow Cytometry
Single cells from tumor tissues were obtained using a tumor dissociation kit and gentle MACS Dissociator (Miltenyi Biotec, Bergisch Gladbach, Germany). Splenocytes were isolated by gently grinding the spleen followed by treatment with RBC lysis buffer (Biolegend). The isolated cells were suspended in a cell staining buffer (Biolegend) and stained with Fixable Viability Stain 700 (BD Bioscience, San Diego, CA, USA) to exclude dead cells. Next, the cells were blocked with TruStain FcX™ PLUS and incubated with fluorescence-conjugated antibodies against CD45, CD3, CD4, CD8, CD335, CD11b, F4/80, CD206, MHC Class II, and GR-1. The data were acquired using a BD LSRFortessa™ X-20 (BD Biosciences, Franklin Lakes, NJ, USA) and analyzed using FlowJo 10.7.1 (BD Biosciences).
2.7. Measurement of Cytokine Production
The blood from KLN205-bearing mice was collected into an EDTA-containing tube, and the plasma was separated by centrifugation at 2000× g for 15 min. The number of cytokines in the plasma was determined using a Bio-Plex Pro Reagent Kit V with a Flat Bottom Plate (#12002798, Bio-Rad, Hercules, CA, USA), which included IL-2, IL-12, IFN-γ, and TNF-α. The plates were prepared according to the manufacturer’s protocol, and 50 μL of each plasma sample was tested. The provided cytokine standard was subjected to serial dilutions. The plates were read using a Bio-Plex 200 System (Bio-Rad) to measure the level of cytokines. This flow-based system identifies each specific reaction based on the bead color and quantitates it.
2.8. Network Pharmacological Analysis of BJIKT
Genes significantly related to NSCLC were identified using GeneCards based on a relevance score of 40 or higher. Information on the target genes of all 10 component herbs of BJIKT was obtained from TCMSP databases [17]. Network analysis was performed using Cytoscape (v3.9.1, Institute for Systems Biology, Seattle, WA, USA). Pathway enrichment analysis was performed in R (v4.2.1) using the enrichR package (v2.1) to obtain p-values for each pathway term. The Gene Ontology Biological Process (GO BP) and Hallmark gene set were obtained from the Molecular Signatures Database. The significance level for each pathway term was calculated as log-transformed adjusted p-values.
2.9. Statistical Analysis
All statistical analyses were conducted using GraphPad Prism 8. The data are expressed as the mean ± standard deviation (SD). The statistical comparison between different groups was based on a two-tailed Student’s t-test and one-way ANOVA followed by Tukey’s post hoc test. p < 0.05 indicated statistical significance.
3. Results
3.1. BJIKT Plus an Anti-PD-1 Antibody Inhibited Tumor Progression in the Lung Cancer Xenograft Model
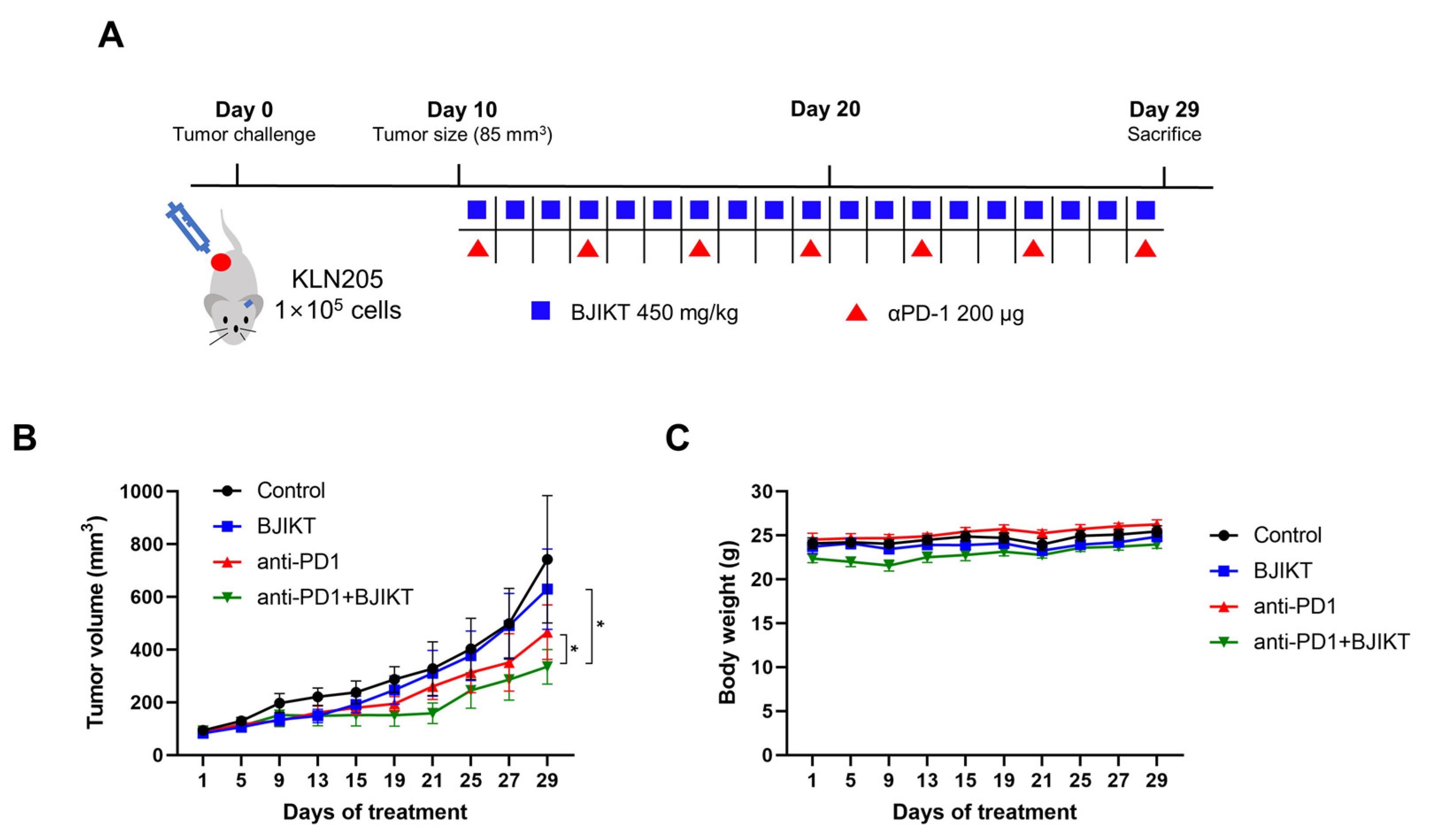
We first confirmed the inhibitory effect of the combination treatment with BJIKT and an anti-PD-1 antibody on tumor growth in KLN205 tumor-bearing mice. When the tumor size reached an average of 85 mm3, the mice were orally administered 450 mg/kg BJIKT daily and/or intraperitoneally administered 200 μg anti-PD-1 antibody every three days (Figure 1A). The tumor inhibition rates after BJIKT, anti-PD-1 antibody, and BJIKT plus anti-PD-1 treatment were 15.24%, 37.24%, and 54.85%, respectively (Figure 1B). A significant inhibition of KLN205 tumor growth was observed in the anti-PD-1 group (p = 0.0274), unlike in the control group. In particular, tumor progression was inhibited to a greater extent with the combination treatment than it was with the anti-PD-1 antibody alone (p = 0.0371). Tumor images were shown in Figure S1. There was no difference in body weight among the groups (Figure 1C). These results suggest that BJIKT inhibits tumor growth by increasing the response to anti-PD-1 immunotherapy in KLN205 tumor-bearing mice.
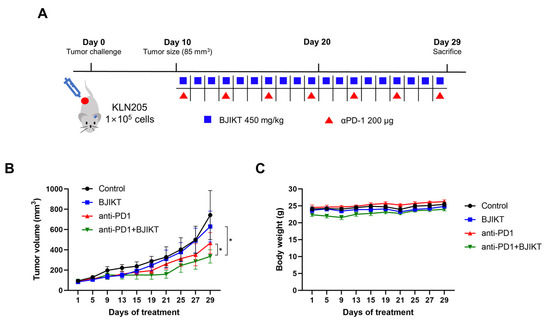
Figure 1.
BJIKT treatment enhances the response to anti-PD-1 immunotherapy in KLN205 tumor-bearing DBA/2 mice. The tumor was established through a subcutaneous injection of 1 × 105 KLN205 cells in mice. Ten days after the injection, the mice were divided into four groups: control, BJIKT, anti-PD-1 antibody, and combination (BJIKT and anti-PD-1 antibody). A dose of 450 mg/kg BJIKT was orally administrated daily, and 200 μg of anti-PD-1 antibody was intraperitoneally administered every three days. (A) Experimental scheme with the timeline of treatment in mice. (B) Tumor volume in KLN205 tumor-bearing mice. Tumor sizes were measured three times a week. (C) Body weight changes in KLN205 tumor-bearing mice. Data are represented as the mean ± SD (n = 5). * p < 0.05, compared to the combination group.
3.2. BJIKT Plus an Anti-PD-1 Antibody Increased the Infiltration of Cytotoxic T Lymphocytes (CTLs) and Macrophages
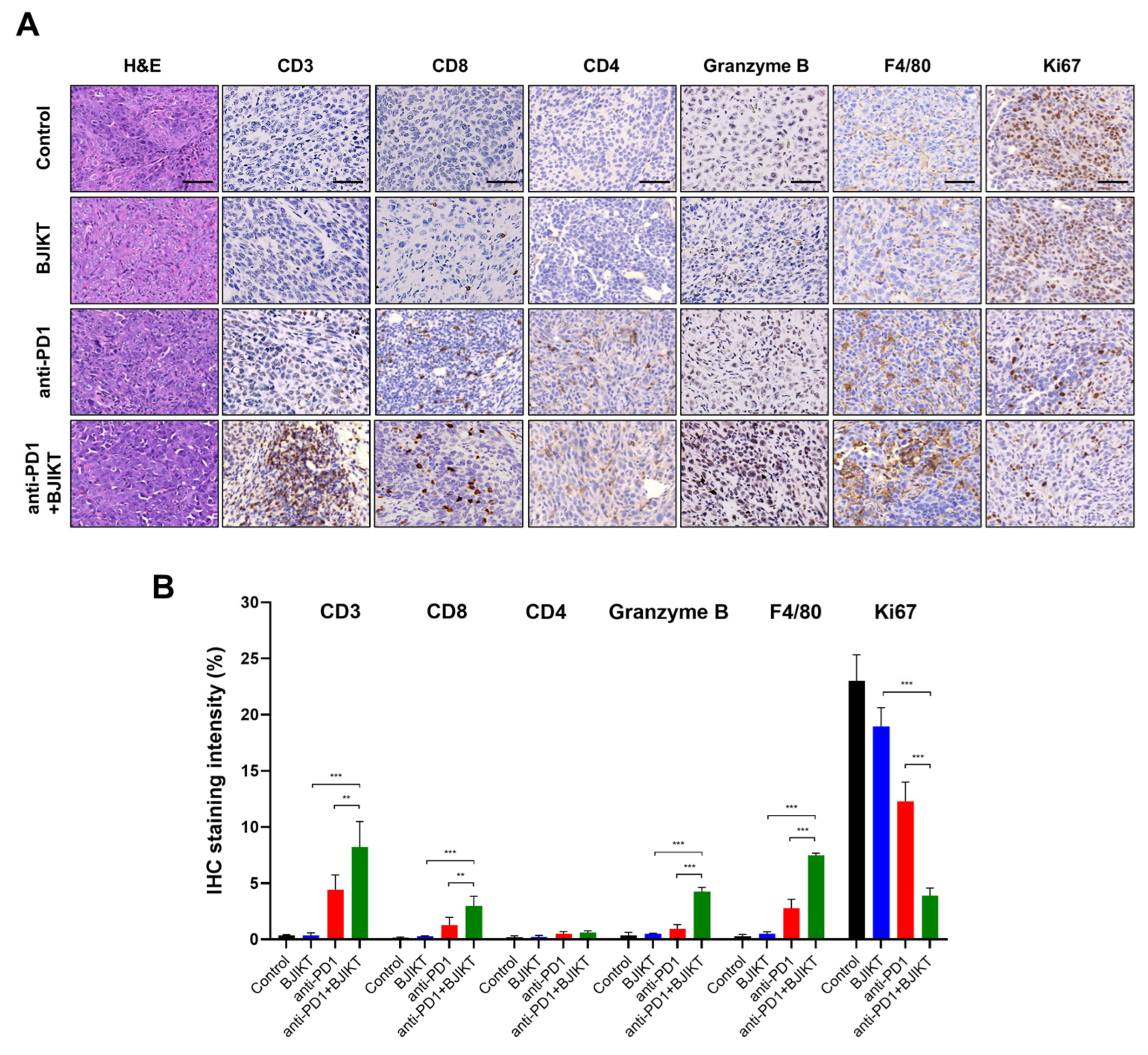
Next, we tested the infiltration of immune cells to the tumor by IHC. As shown in Figure 2A,B, BJIKT monotherapy did not affect CD3+, CD4+, and CD8+ T cell infiltration and granzyme B levels. However, the combination treatment of BJIKT and an anti-PD-1 antibody increased the expression of CD3, CD8, and granzyme B. In addition, the combination treatment increased the levels of F4/80, which is a macrophage marker, and decreased the levels of Ki67, which is a cell proliferation marker. The combination treatment did not change the expression of CD4. These results suggest that BJIKT regulates the activation of CTLs, which is closely associated with the PD-1/PD-L1 axis in the TME, thereby enhancing the response to the anti-PD-1 antibody.
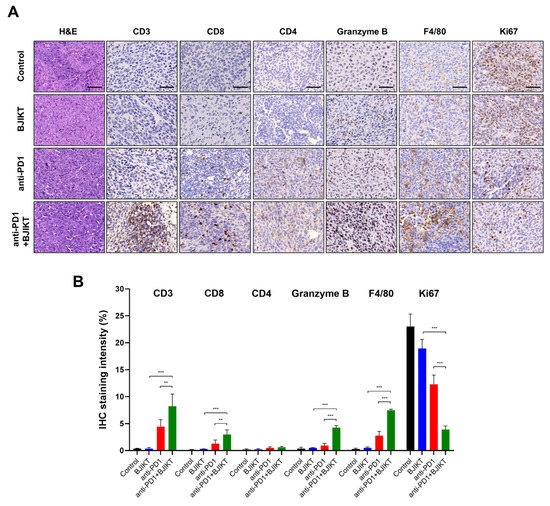
Figure 2.
BJIKT with an anti-PD-1 antibody changed the immune profiles in the tumor microenvironment. (A) Hematoxylin and eosin staining and immunohistochemistry (IHC) staining findings for the tumor infiltration of immune cells. CD3, CD8, CD4, granzyme B, F4/80, and Ki67 levels were assessed by the microscopic examination of IHC-stained sections. Representative stained sections are shown (scale bars = 50 µm). (B) Quantitative analysis of IHC staining was performed using the ImageJ software. Data are represented as the mean ± SD. ** p < 0.01 and *** p < 0.001, compared to the combination group.
3.3. BJIKT Plus an Anti-PD-1 Antibody Regulated Antitumor Immune Response through CTLs, MDSCs, and Macrophages
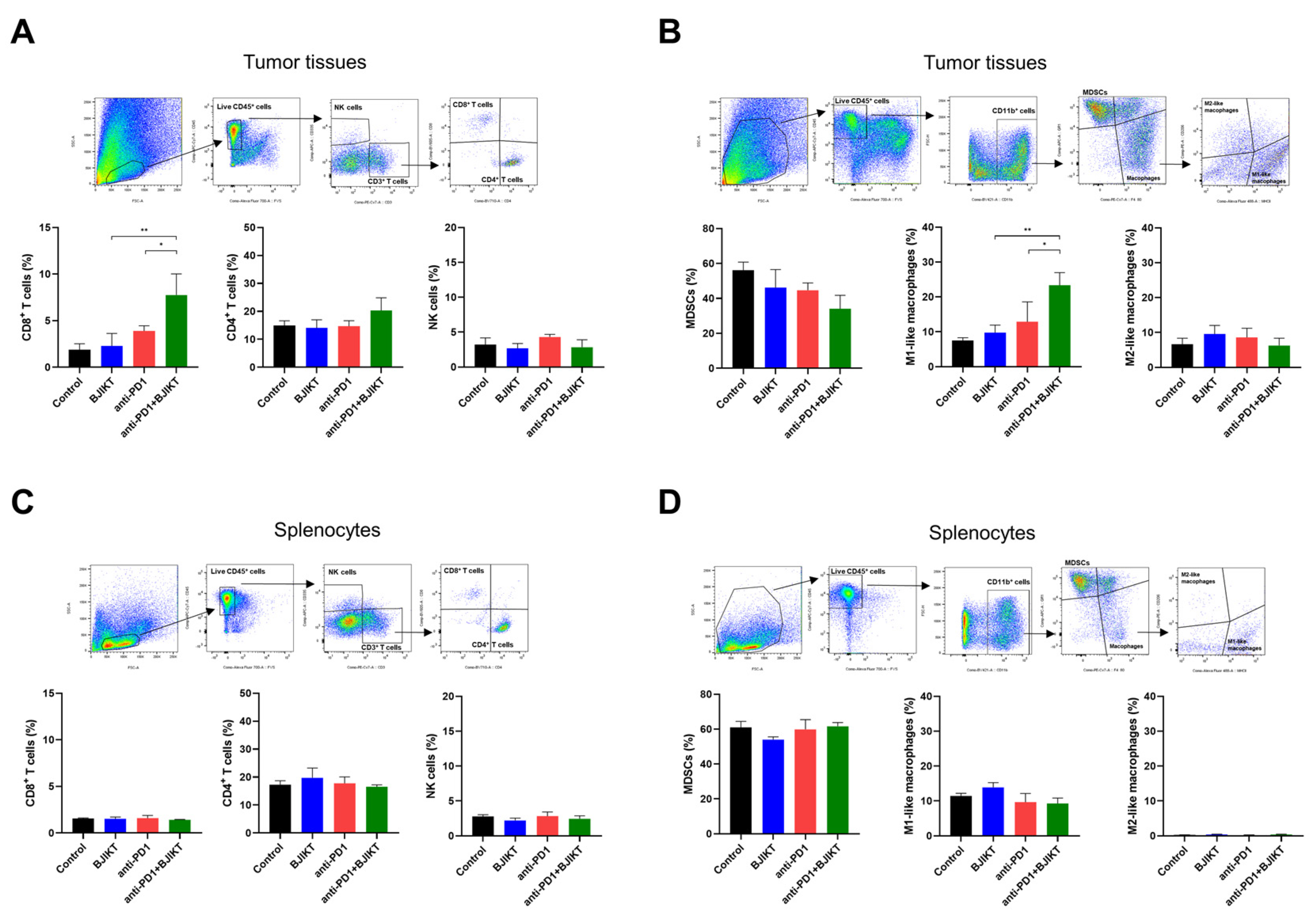
To explore the effect of combination treatment with BJIKT and an anti-PD-1 antibody on the activation of CTLs and macrophages, we analyzed the proportion of single cells, including T cells, NK cells, MDSCs, and macrophages, dissociated from tumor tissues and the spleen. In tumor tissues, BJIKT plus anti-PD-1 promoted antitumor immune response by increasing the population of CD8+ T cells (CD3+, CD8+) but not of CD4+ T cells (CD3+, CD4+). The combination treatment did not change the population of NK cells (CD3−, CD335+) (Figure 3A). MDSCs and macrophages are immunosuppressive cells that strongly inhibit the activation of T cells in the TME, contributing to the poor response to anti-PD-1 antibodies. Thus, we analyzed MDSCs (CD11b+, GR-1+) and macrophages (CD11b+, F4/80+) in tumor tissues and splenocytes. In tumor tissues, the number of MDSCs was lower in the combination treatment group than it was in the BJIKT and anti-PD-1 antibody monotherapy groups. The number of macrophages was higher in the combination treatment group than it was in the anti-PD-1 group. Interestingly, the combination treatment with BJIKT and an anti-PD-1 antibody increased the population of M1-like macrophages but did not affect the number of M2-like macrophages (Figure 3B). In splenocytes, no significant changes were observed in the population of lymphocytes (Figure 3C) and myeloid cells (Figure 3D). These results suggest that BJIKT combined with an anti-PD-1 antibody enhances the response to anti-PD-1 immunotherapy in KLN205 tumor-bearing mice by regulating the population of MDSCs and M1-like macrophages.
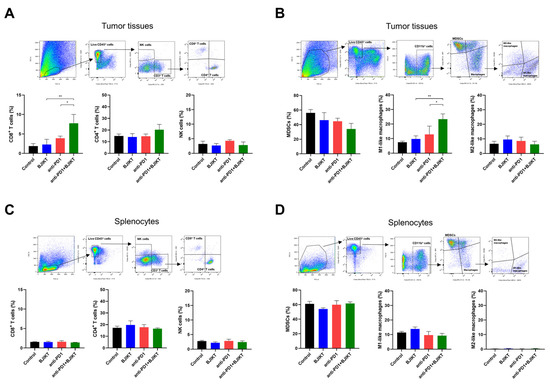
Figure 3.
Flow cytometry analysis of immune markers on tumor tissues and splenocytes in KLN205 tumor-bearing mice. The cells from tumor tissues and the spleen were dissociated and gated based on CD45+ live cells. (A) Lymphocytes were stained with anti-CD45, anti-CD3, anti-CD4, anti-CD8, and anti-CD335 (NKp46) antibodies for tumor tissue analysis. (B) Myeloid cells were stained with anti-CD45, anti-CD11b, anti-F4/80, anti-GR-1, anti-MHC Class II, and anti-CD206 antibodies for tumor tissue analysis. (C) Lymphocytes were stained with anti-CD45, anti-CD3, anti-CD4, anti-CD8, and anti-CD335 antibodies for splenocytes analysis. (D) Myeloid cells were stained with anti-CD45, anti-CD11b, anti-F4/80, anti-GR-1, anti-MHC Class II, and anti-CD206 antibodies for splenocyte analysis. * p < 0.05 and ** p < 0.01, compared to the combination group.
3.4. BJIKT Combined with an Anti-PD-1 Antibody Altered the Concentration of Th1 Cytokines in the Peripheral Blood
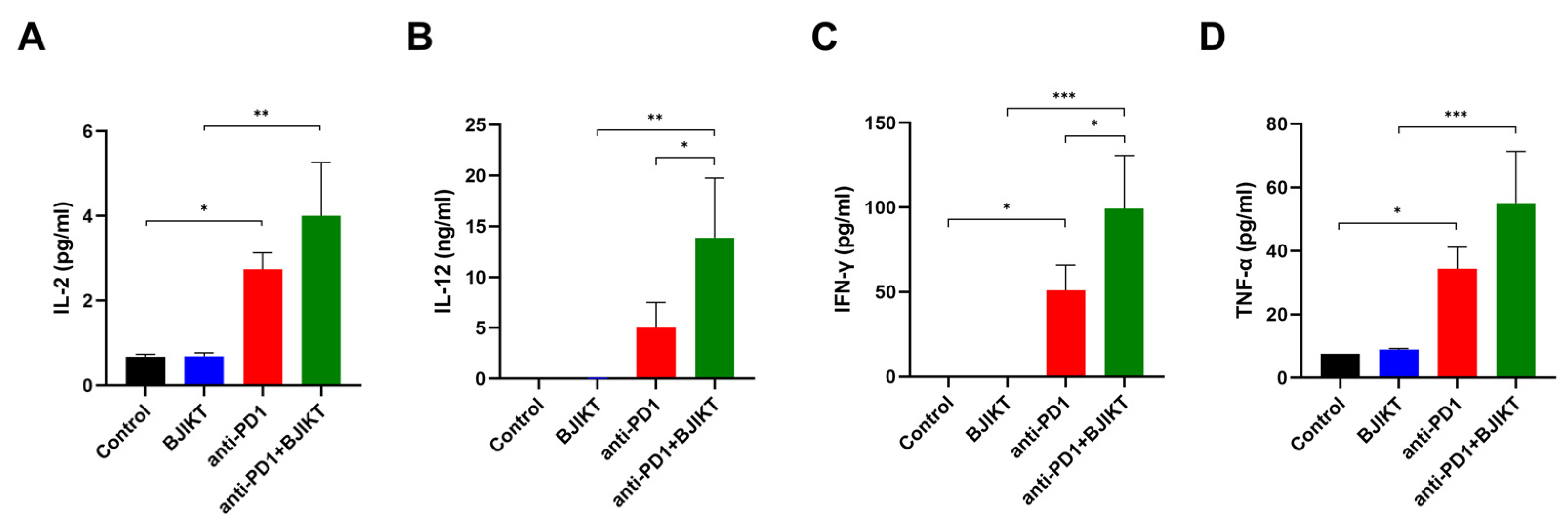
To determine the change in Th1 cytokine levels, following the combination treatment with BJIKT and an anti-PD-1 antibody, we evaluated the plasma concentrations of IL-2, IL-12, INF-γ, and TNF-α. As shown in Figure 4A–D, IL-2 (p = 0.0206), IL-12 (p = 0.2981), INF-γ (p = 0.0288), and TNF-α (p = 0.0241) concentrations in anti-PD-1 group were higher than those in the control group. The combination treatment with BJIKT and an anti-PD-1 antibody significantly increased the levels of IL-2 (p = 0.0012), IL-12 (p = 0.0032), INF-γ (p = 0.0005), and TNF-α (p = 0.0009) compared to the levels in the BJIKT group. Significant differences in the levels of IL-12 (p = 0.0376) and INF-γ (p = 0.0373) were observed between the anti-PD-1 group and the combination group. These results suggest that BJIKT combined with an anti-PD-1 antibody activates the Th1-mediated immunity response, primarily via IFN-γ.
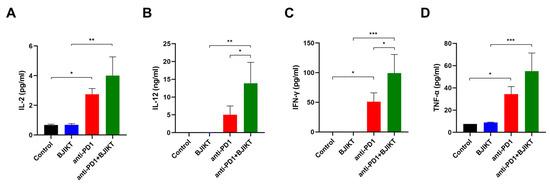
Figure 4.
Change in inflammatory cytokine levels in plasma samples collected from mice. The levels of IL-2 (A), IL-12 (B), IFN-γ (C), and TNF-α (D) were determined by the Bio-Plex Pro™ Mouse Cytokine Assay. * p < 0.05, ** p < 0.01, and *** p < 0.001, compared between each group.
3.5. Network Pharmacology Analysis Predicted the Target Pathways of BJIKT to Treat NSCLC
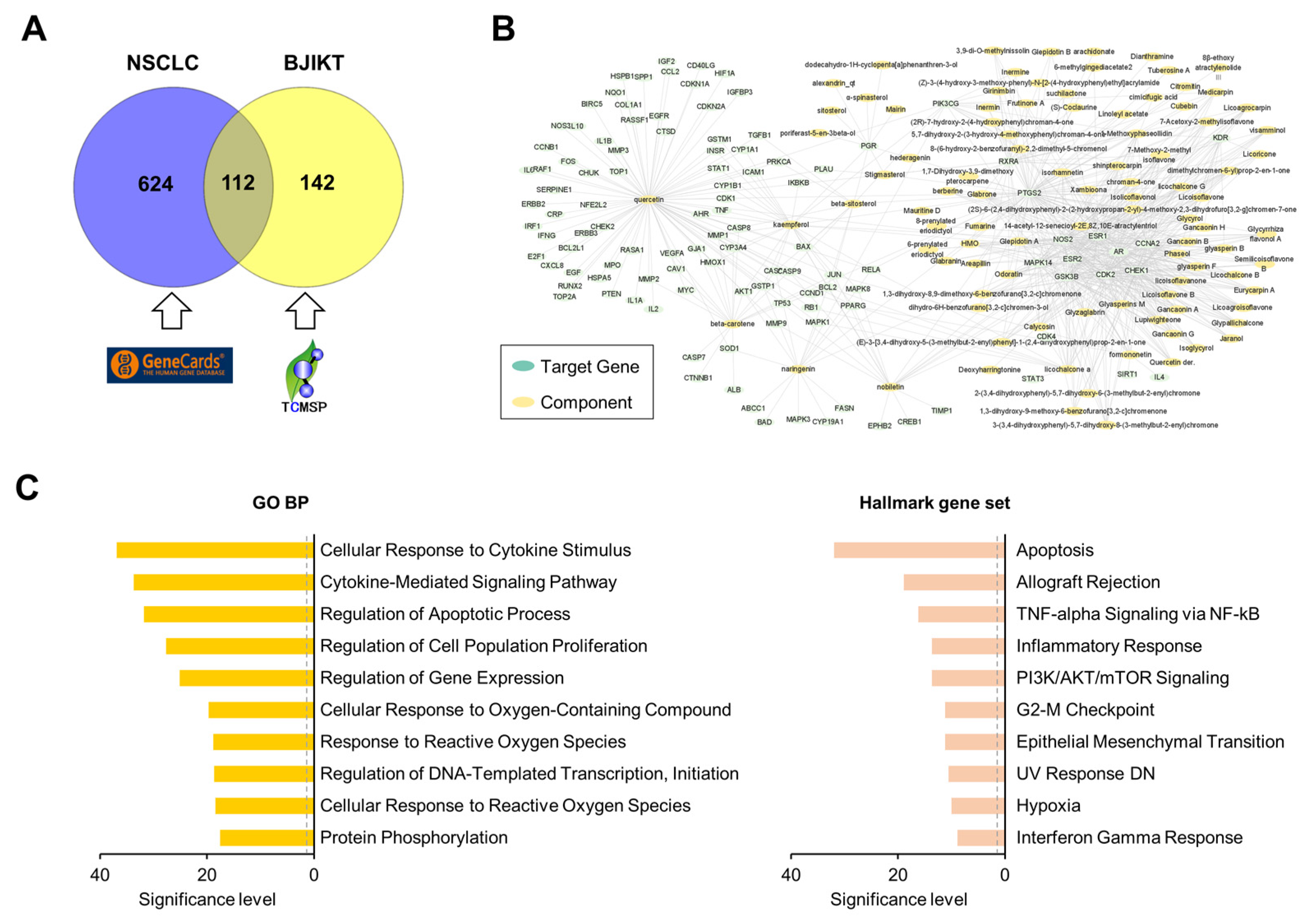
To investigate the molecular mechanism of BJIKT in NSCLC, we adopted a network pharmacology approach. A total of 736 genes significantly associated with NSCLC were extracted from GeneCards (Table S1). Compounds with an oral bioavailability ≥ 35%, a drug-likeness ≥ 0.18, a half-life ≥ 4, and a Caco-2 cell permeability ≥ −0.4 were selected as active ingredients of BJIKT (Table S2). A total of 254 target genes of 119 active ingredients in BJIKT were extracted by integrating information from the TCMSP databases (Table S3). There were 112 common genes between the NSCLC-associated genes and the target genes of BJIKT (Table S4). Its Venn diagram is shown in Figure 5A. Based on this information, we reconstructed a network for the molecular targets of BJIKT components acting in NSCLC (Figure 5B). PTGS2, ESR1, NOS2, AR, GSK3B, CDK2, CCNA2, and ESR2 were the main target genes of multiple components in BJIKT. We next performed pathway enrichment analysis on the common target genes using the GO BP and Hallmark gene sets (Figure 5C). Consistent with the experimental results, cytokine-mediated signaling, INF-γ response, and TNF-α and NF-κB signaling were significantly enriched in the target pathways of BJIKT. In addition, BJIKT may inhibit NSCLC progression by regulating the pathways associated with apoptosis and cell proliferation.
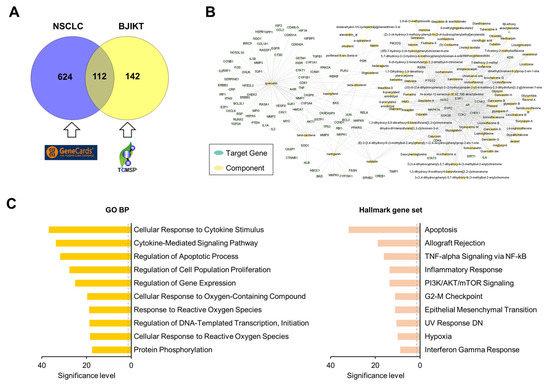
Figure 5.
Network pharmacological analysis of BJIKT on NSCLC. (A) Venn diagram for 736 NSCLC-related genes obtained from GeneCard and 254 target genes of BJIKT obtained from TCMSP; 112 genes were common. (B) Reconstructed network for molecular targets of BJIKT components acting on NSCLC. (C) Top 10 enriched pathways for the common 112 target genes using GO BP (left) and the Hallmark gene set (right). The significance level of each pathway term was calculated as −log10 (adjusted p-value). The dashed gray line denotes a p-value of 0.05.
4. Discussion
In the present study, we investigated the antitumor effect of BJIKT combined with an anti-PD-1 antibody in KLN205 lung tumor-bearing mice. Although immunotherapy is the most promising treatment for lung cancer, the immune-competent lung cancer animal models are substantially limited. There are several murine lung cancer cell lines: LL/2, CMT64, CMT167, LA-4, and KLN205. Among them, we used the KLN205 syngeneic tumor model by subcutaneously injecting KLN205 cells into syngeneic DBA/2 mice, since it provides a moderate immune response in a mouse with a functional immune system [18,19]. Here, we found that the antitumor effects of the combination treatment with BJIKT plus an anti-PD-1 antibody were significantly higher than those with BJIKT or an anti-PD-1 antibody monotherapy. The combination treatment increased the infiltration of tumor lymphocytes and regulated the antitumor immune response by regulating MDSCs and M1-like macrophages, suggesting that BJIKT enhances the effects of PD-1 immunotherapy for NSCLC. The present study is the first to demonstrate the immune-mediated antitumor activity of BJIKT against NSCLC by enhancing the tumor response to the anti-PD-1 antibody.
ICIs have greatly altered the landscape of patient care in multiple cancer types [20]. Since pembrolizumab and nivolumab were approved by the US Food and Drug Administration as PD-1 blockers in 2015, ICIs have brought in a new era of immunotherapy in NSCLC [21]. Although PD-1 inhibitors are now approved as standard anticancer treatments for patients with advanced NSCLC, their low response rate to this therapy remains to be resolved [22]. This limitation of PD-1 blockade has made it necessary to develop combination regimens with PD-1 inhibitors that can promote antitumor immunity and enhance the treatment response [23]. The combination strategies effectively regulate multiple steps in the cancer immunity cycle, remove immunosuppressive factors, and restore immune function to the immune-supportive condition in the TME [24]. Therefore, the combination of natural products with an immunotherapeutic agent can be advantageous during immunotherapy [25]. Recently, polysaccharides have emerged as promising immune-modulating agents in cancer immunotherapy due to their immunity improvement and antitumor effects. The immunomodulating effect of polysaccharides includes the activation of immune cells, such as tumor-infiltrating lymphocytes, dendritic cells, and macrophages, and the release of cytokines, such as IFN-γ, TNF-α, and interleukins [26,27]. Astragalus polysaccharides and ginseng polysaccharides enhance the antitumor response of immunotherapies [28,29]. In addition, the polysaccharides of A. macrocephala, A. gigas, and licorice regulate the antitumor immunity of macrophages and CTLs in the tumor model [30,31,32]. The herbs mentioned above are the main components of BJIKT, suggesting that the increased response of anti-PD-1 by BJIKT is likely the result of the polysaccharides in BJIKT. However, the detailed mechanism of immune response needs to be further investigated. Nevertheless, the combination of BJIKT and an anti-PD-1 antibody significantly improved antitumor immunity and enhanced the response to immunotherapy.
In the TME, various cells are involved in the immune regulatory networks. It is a major barrier to the infiltration and activation of effector immune cells [33]. MDSCs and macrophages are key immunosuppressive cells in TME. MDSCs are known to induce intense immunosuppression by inhibiting the functions of CTLs and NK cells [34]. Macrophages are classified into proinflammatory M1 (classically activated)-like and anti-inflammatory M2 (alternatively activated)-like macrophages. Recent studies have shown that M1-like macrophages are dominant in the early stages of lung cancer and undergo the transition to M2-like macrophages [35]. Therefore, the regulation of MDSCs and the polarization of M1-like macrophages by herbal medicine in TME can be effective as an adjuvant treatment for NSCLC. In our study, the combination treatment with BJIKT and an anti-PD-1 antibody significantly decreased the population of MDSCs but increased the population of M1-like macrophages in tumor tissues, suggesting that the combination treatment suppressed KLN205-tumor growth by inducing the polarization of M1-like macrophages and subsequently activating CTLs in the TME. Further studies are required to explore the components of BJIKT involved in the polarization of macrophages.
Systemic pharmacology is useful for understanding the mode of action of TCMs. In this study, we undertook a comprehensive approach to investigate the pharmacological effects of BJIKT in antitumor immunity. The main target genes of BJIKT, including PTGS2, ESR1, NOS2, AR, GSK3B, CDK2, CCNA2, and ESR2, were selectively related to specific cancer types, such as lung, breast, and prostate cancer, or generally related across types. Especially, ESR2 was revealed to promote NSCLC progression [36]. CCNA2 was correlated with overall survival in patients with NSCLC [37]. In addition, PTGS2 could be a therapeutic target for improving the response to the therapeutic agents in resistant NSCLC [38]. The molecular targets of BJIKT in NSCLC were enriched in pathways regulating apoptosis and cell proliferation as well as cytokine-mediated signaling, INF-γ response, and TNF-α and NF-κB signaling. Therefore, we speculated that the targets of BJIKT were associated with the antitumor immune response and increased the response to anti-PD-1 antibody immunotherapy. Previous research demonstrated that BJIKT exerts anticancer effects to induce apoptosis in human lung cancer H460 cells [39]. In addition, BJIKT inhibited the proliferation of NSCLC A549/DDP cells [40] and hepatoma cell lines by inducing apoptosis [41]. These results support the idea that BJIKT may inhibit NSCLC progression by regulating the pathways associated with apoptosis and cell proliferation. Although network pharmacology provides drug target prediction, the mechanism by which it affects NSCLC is quite complex because the action of TCM is not only based on combining the active compounds. In addition, the analysis of the TCM target based on network pharmacology is still in an early stage [42]. Network pharmacology mainly depends on databases to acquire original data. Additionally, active components are associated with various targets regarding other diseases. Therefore, the results obtained from network pharmacology may be inconsistent with expectations or experimental models. Further studies on the main components of BJIKT and their related signaling pathways using the integrated databases are needed to overcome these limitations and verify targets in an experimental model. Nevertheless, our results provide the rationale for the clinical evaluation of BJIKT for NSCLC in combination with immunotherapies.
Although the subcutaneous microenvironment differs from the actual lung, the KLN205 tumor syngeneic model provides the immune-competent environment that is necessary for the study of immune responses, and it is useful for testing the immune response of an anti-PD-1 antibody in NSCLC [18,19]. Poczobutt et al. employed an orthotopic immunocompetent model of lung adenocarcinoma, in which LL/2 cells are directly implanted into the C57BL/6 mice [43]. However, they are negative to cancer cell-specific MHC class II and resistant to immunotherapies [44], which might not be useful for the research of immunotherapies. Further studies are needed to investigate the effect of the combination treatment with BJIKT and an anti-PD-1 antibody in an orthotopic lung model or a humanized mouse model with lung cancer immune system.
5. Conclusions
Our study indicated that the combination treatment with BJIKT and an anti-PD-1 antibody significantly improved antitumor effects in vivo, suggesting that this therapeutic strategy may be a promising treatment option for NSCLC. However, further studies are needed to fully understand the detailed mechanism of action of this combination treatment.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pr10091683/s1, Table S1: Genes significantly associated with NSCLC. Table S2: Compounds selected as active ingredients of BJIKT. Table S3: Target genes of active ingredients. Table S4: The common genes between the NSCLC-associated genes and target genes of BJIKT. Figure S1: Tumor images from DBA/2 mice.
Author Contributions
Conceptualization, J.C. and M.-K.J.; investigation, J.C., H.N.K., J.-M.Y. and S.H.H.; methodology, J.C. and S.-M.P.; writing—original draft preparation, J.C., H.N.K. and S.-M.P.; writing—review and editing, J.-M.Y. and M.-K.J.; visualization, J.C., H.N.K. and S.-M.P.; project administration, M.-K.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Korea Institute of Oriental Medicine, grant number KSN2022240.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of the Korea Institute of Oriental Medicine (Approval number: 21-038).
Data Availability Statement
All data used and analyzed to support the current study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yuan, M.; Huang, L.-L.; Chen, J.-H.; Wu, J.; Xu, Q. The emerging treatment landscape of targeted therapy in non-small-cell lung cancer. Signal Transduct. Target. Ther. 2019, 4, 61. [Google Scholar] [CrossRef]
- Liu, W.-J.; Du, Y.; Wen, R.; Yang, M.; Xu, J. Drug resistance to targeted therapeutic strategies in non-small cell lung cancer. Pharmacol. Ther. 2020, 206, 107438. [Google Scholar] [CrossRef]
- Assi, H.I.; Kamphorst, A.O.; Moukalled, N.M.; Ramalingam, S.S. Immune checkpoint inhibitors in advanced non-small cell lung cancer. Cancer 2018, 124, 248–261. [Google Scholar] [CrossRef]
- Bodor, J.N.; Boumber, Y.; Borghaei, H. Biomarkers for immune checkpoint inhibition in non-small cell lung cancer (NSCLC). Cancer 2020, 126, 260–270. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef]
- Haist, M.; Stege, H.; Grabbe, S.; Bros, M. The Functional Crosstalk between Myeloid-Derived Suppressor Cells and Regulatory T Cells within the Immunosuppressive Tumor Microenvironment. Cancers 2021, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Vasievich, E.A.; Huang, L. The suppressive tumor microenvironment: A challenge in cancer immunotherapy. Mol. Pharmacol. 2011, 8, 635–641. [Google Scholar] [CrossRef]
- Kumari, S.; Advani, D.; Sharma, S.; Ambasta, R.K.; Kumar, P. Combinatorial therapy in tumor microenvironment: Where do we stand? Biochim. Biophys. Acta 2021, 1876, 188585. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.E.; Kim, J.N.; Kwon, M.J.; Lee, J.R.; Kim, S.C.; Nam, J.H.; Kim, B.J. The traditional medicine bojungikki-tang increases intestinal motility. Phcog. Mag. 2021, 17, 1–8. [Google Scholar] [CrossRef]
- Jeong, J.S.; Ryu, B.H.; Kim, J.S.; Park, J.W.; Choi, W.C.; Yoon, S.W. Bojungikki-tang for cancer-related fatigue: A pilot randomized clinical trial. Integr. Cancer Ther. 2010, 9, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.-J.; Kim, K.-I.; Choi, C.-W.; Kim, J.Y.; Lee, J.-H. Long-term progression-free survival in a patient with advanced non-small-cell lung cancer treated with low-dose gefitinib and traditional herbal medicine: A case report. Medicine 2021, 100, e24292. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kita, K.; Sato, C.; Kaneda, A. Hochuekkito (Buzhongyiqitang), a herbal medicine, enhances cisplatin-induced apoptosis in HeLa cells. Mol. Med. Report. 2015, 12, 6215–6220. [Google Scholar] [CrossRef] [PubMed]
- Utsuyama, M.; Seidlar, H.; Kitagawa, M.; Hirokawa, K. Immunological restoration and anti-tumor effect by Japanese herbal medicine in aged mice. Mech. Ageing Dev. 2001, 122, 341–352. [Google Scholar] [CrossRef]
- Li, T.; Tamada, K.; Abe, K.; Tada, H.; Onoe, Y.; Tatsugami, K.; Harada, M.; Kubo, C.; Nomoto, K. The restoration of the antitumor T cell response from stress-induced suppression using a traditional Chinese herbal medicine Hochu-ekki-to (TJ-41:Bu-Zhong-Yi-Qi-Tang). Immunopharmacology 1999, 43, 11–21. [Google Scholar] [CrossRef]
- Chun, J.; Park, S.-M.; Yi, J.-M.; Ha, I.J.; Kang, H.N.; Jeong, M.-K. Bojungikki-Tang improves response to PD-L1 immunotherapy by regulating the tumor microenvironment in MC38 tumor-bearing mice. Front. Pharmacol. 2022, 13, 901563. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef]
- Du, W.; Huang, H.; Sorrelle, N.; Brekken, R.A. Sitravatinib potentiates immune checkpoint blockade in refractory cancer models. JCI Insight 2018, 3, 124184. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, K.; Xiao, Y.; Feng, B.; Mikule, K.; Ma, X.; Feng, N.; Vellano, C.P.; Federico, L.; Marszalek, J.R.; et al. Niraparib activates interferon signaling and potentiates anti-PD-1 antibody efficacy in tumor models. Sci. Rep. 2019, 9, 1853. [Google Scholar] [CrossRef]
- Rizvi, H.; Sanchez-Vega, F.; La, K.; Chatila, W.; Jonsson, P.; Halpenny, D.; Plodkowski, A.; Long, N.; Sauter, J.L.; Rekhtman, N.; et al. Molecular Determinants of Response to Anti-Programmed Cell Death (PD)-1 and Anti-Programmed Death-Ligand 1 (PD-L1) Blockade in Patients with Non-Small-Cell Lung Cancer Profiled With Targeted Next-Generation Sequencing. J. Clin. Oncol. 2018, 36, 633–641. [Google Scholar] [CrossRef]
- Sui, H.; Ma, N.; Wang, Y.; Li, H.; Liu, X.; Su, Y.; Yang, J. Anti-PD-1/PD-L1 Therapy for Non-Small-Cell Lung Cancer: Toward Personalized Medicine and Combination Strategies. J. Immunol. Res. 2018, 2018, 6984948. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Yi, M.; Li, N.; Luo, S.; Wu, K. Predictive biomarkers of anti-PD-1/PD-L1 therapy in NSCLC. Exp. Hematol. Oncol. 2021, 10, 18. [Google Scholar] [CrossRef]
- Yi, M.; Zheng, X.; Niu, M.; Zhu, S.; Ge, H.; Wu, K. Combination strategies with PD-1/PD-L1 blockade: Current advances and future directions. Mol. Cancer 2022, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Pio, R.; Ajona, D.; Ortiz-Espinosa, S.; Mantovani, A.; Lambris, J.D. Complementing the Cancer-Immunity Cycle. Front. Immunol. 2019, 10, 774. [Google Scholar] [CrossRef]
- Deng, L.-J.; Qi, M.; Li, N.; Lei, Y.-H.; Zhang, D.-M.; Chen, J.-X. Natural products and their derivatives: Promising modulators of tumor immunotherapy. J. Leukoc. Biol. 2020, 108, 493–508. [Google Scholar] [CrossRef]
- Yang, Y.; Li, N.; Wang, T.-M.; Di, L. Natural Products with Activity against Lung Cancer: A Review Focusing on the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 10827. [Google Scholar] [CrossRef]
- Liu, L.; Nie, S.; Xie, M. Tumor Microenvironment as a New Target for Tumor Immunotherapy of Polysaccharides. Crit. Rev. Food Sci. Nutr. 2016, 56 (Suppl. S1), S85–S94. [Google Scholar] [CrossRef]
- Hwang, J.; Zhang, W.; Dhananjay, Y.; An, E.-K.; Kwak, M.; You, S.; Lee, P.C.; Jin, J.-O. Astragalus membranaceus polysaccharides potentiate the growth-inhibitory activity of immune checkpoint inhibitors against pulmonary metastatic melanoma in mice. Int. J. Biol. Macromol. 2021, 182, 1292–1300. [Google Scholar] [CrossRef]
- Huang, J.; Liu, D.; Wang, Y.; Liu, L.; Li, J.; Yuan, J.; Jiang, Z.; Jiang, Z.; Hsiao, W.W.; Liu, H.; et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 2022, 71, 734–745. [Google Scholar] [CrossRef]
- Feng, Z.; Yang, R.; Wu, L.; Tang, S.; Wei, B.; Guo, L.; He, L.; Feng, Y. Atractylodes macrocephala polysaccharides regulate the innate immunity of colorectal cancer cells by modulating the TLR4 signaling pathway. Onco Targets Ther. 2019, 12, 7111–7121. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lee, S.W.; Park, H.J.; Lee, S.H.; Im, W.K.; Kim, Y.D.; Kim, K.H.; Park, S.J.; Hong, S.; Jeon, S.H. Anti-cancer activity of Angelica gigas by increasing immune response and stimulating natural killer and natural killer T cells. BMC Complement. Altern. Med. 2018, 18, 218. [Google Scholar] [CrossRef] [PubMed]
- Ayeka, P.A.; Bian, Y.; Githaiga, P.M.; Zhao, Y. The immunomodulatory activities of licorice polysaccharides (Glycyrrhiza uralensis Fisch.) in CT 26 tumor-bearing mice. BMC Complement. Altern. Med. 2017, 17, 536. [Google Scholar] [CrossRef]
- Duan, Q.; Zhang, H.; Zheng, J.; Zhang, L. Turning Cold into Hot: Firing up the Tumor Microenvironment. Trends Cancer 2020, 6, 605–618. [Google Scholar] [CrossRef]
- Shi, H.; Li, K.; Ni, Y.; Liang, X.; Zhao, X. Myeloid-Derived Suppressor Cells: Implications in the Resistance of Malignant Tumors to T Cell-Based Immunotherapy. Front. Cell Dev. Biol. 2021, 9, 707198. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, X.; Sun, Q.; Zhang, W.; Liu, C.; Ma, W.; Sun, C. Natural compounds: A new perspective on targeting polarization and infiltration of tumor-associated macrophages in lung cancer. Biomed. Pharmacother. 2022, 151, 113096. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cai, Y.; Wang, Z.; He, W.; Cao, S.; Xu, R.; Chen, H. Estrogen receptors promote NSCLC progression by modulating the membrane receptor signaling network: A systems biology perspective. J. Transl. Med. 2019, 17, 308. [Google Scholar] [CrossRef]
- Gong, K.; Zhou, H.; Liu, H.; Xie, T.; Luo, Y.; Guo, H.; Chen, J.; Tan, Z.; Yang, Y.; Xie, L. Identification and Integrate Analysis of Key Biomarkers for Diagnosis and Prognosis of Non-Small Cell Lung Cancer Based on Bioinformatics Analysis. Technol. Cancer Res. Treat. 2021, 20, 15330338211060202. [Google Scholar] [CrossRef]
- Lin, X.M.; Luo, W.; Wang, H.; Li, R.Z.; Huang, Y.S.; Chen, L.K.; Wu, X.P. The Role of Prostaglandin-Endoperoxide Synthase-2 in Chemoresistance of Non-Small Cell Lung Cancer. Front. Pharmacol. 2019, 10, 836. [Google Scholar] [CrossRef]
- Lee, S.-E.; Hong, J.-E.; Lee, S.-H.; Shin, J.-Y.; Ro, S.-S. Study on Apoptosis Effect and Mechanism by Bojungikki-tang on Human Cancer Cell Line H460. J. Int. Kor. Med. 2004, 25, 274–288. [Google Scholar]
- Yu, N.; Xiong, Y.; Wang, C. Bu-Zhong-Yi-Qi Decoction, the Water Extract of Chinese Traditional Herbal Medicine, Enhances Cisplatin Cytotoxicity in A549/DDP Cells through Induction of Apoptosis and Autophagy. BioMed Res. Int. 2017, 2017, 3692797. [Google Scholar] [CrossRef]
- Kao, S.T.; Yeh, C.C.; Hsieh, C.C.; Yang, M.D.; Lee, M.R.; Liu, H.S.; Lin, J.G. The Chinese medicine Bu-Zhong-Yi-Qi-Tang inhibited proliferation of hepatoma cell lines by inducing apoptosis via G0/G1 arrest. Life Sci. 2001, 69, 1485–1496. [Google Scholar] [CrossRef]
- Dong, Y.; Duan, L.; Chen, H.W.; Liu, Y.M.; Zhang, Y.; Wang, J. Network Pharmacology-Based Prediction and Verification of the Targets and Mechanism for Panax Notoginseng Saponins against Coronary Heart Disease. Evid. Based Complement. Alternat. Med. 2019, 2019, 6503752. [Google Scholar] [CrossRef] [PubMed]
- Poczobutt, J.M.; De, S.; Yadav, V.K.; Nguyen, T.T.; Li, H.; Sippel, T.R.; Weiser-Evans, M.C.; Nemenoff, R.A. Expression Profiling of Macrophages Reveals Multiple Populations with Distinct Biological Roles in an Immunocompetent Orthotopic Model of Lung Cancer. J. Immunol. 2016, 196, 2847–2859. [Google Scholar] [CrossRef] [PubMed]
- Neuwelt, A.J.; Kimball, A.K.; Johnson, A.M.; Arnold, B.W.; Bullock, B.L.; Kaspar, R.E.; Kleczko, E.K.; Kwak, J.W.; Wu, M.H.; Heasley, L.E.; et al. Cancer cell-intrinsic expression of MHC II in lung cancer cell lines is actively restricted by MEK/ERK signaling and epigenetic mechanisms. J. Immunother. Cancer 2020, 8, e000441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).