
Modeling and Optimal Control of an Electro-Fermentation Process within a Batch Culture

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
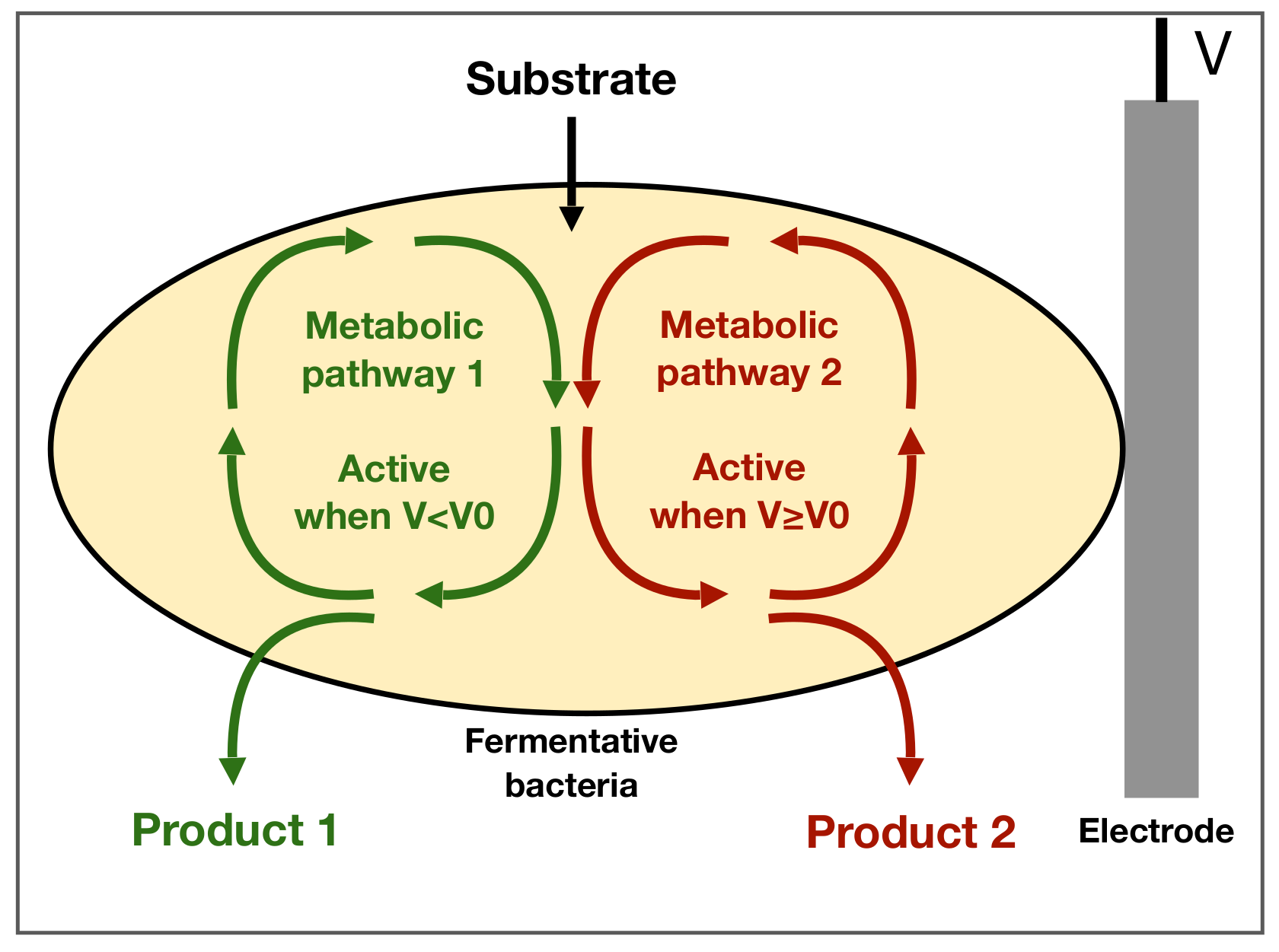
2. Model Description and Optimization Problem
3. Sufficient Condition for the Absence of Singular Arc
4. Optimal Synthesis with Identical Growth Functions
5. Optimal Synthesis with Constant Growth Functions
- If , then is optimal on .
- If , then the control:is optimal on .
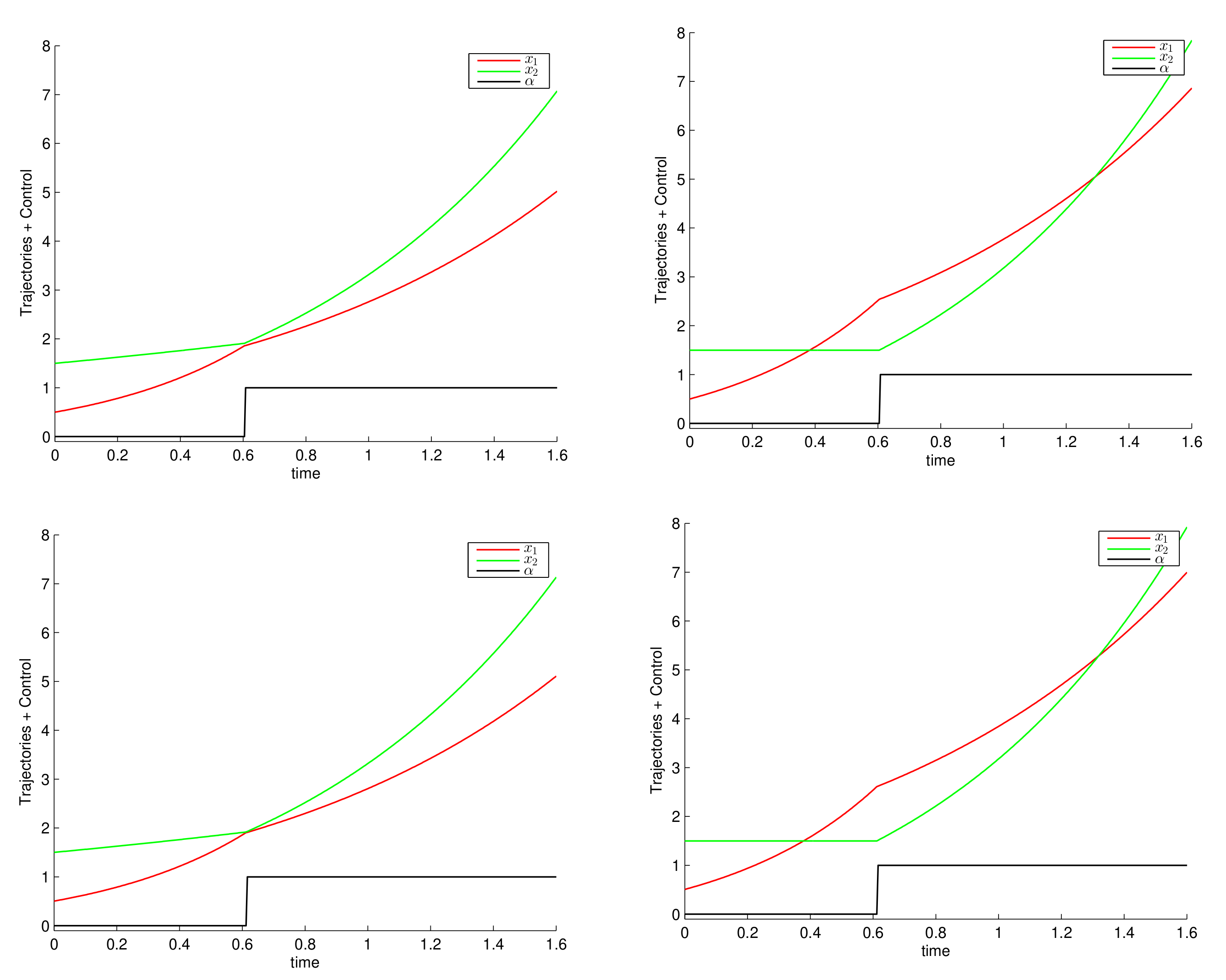
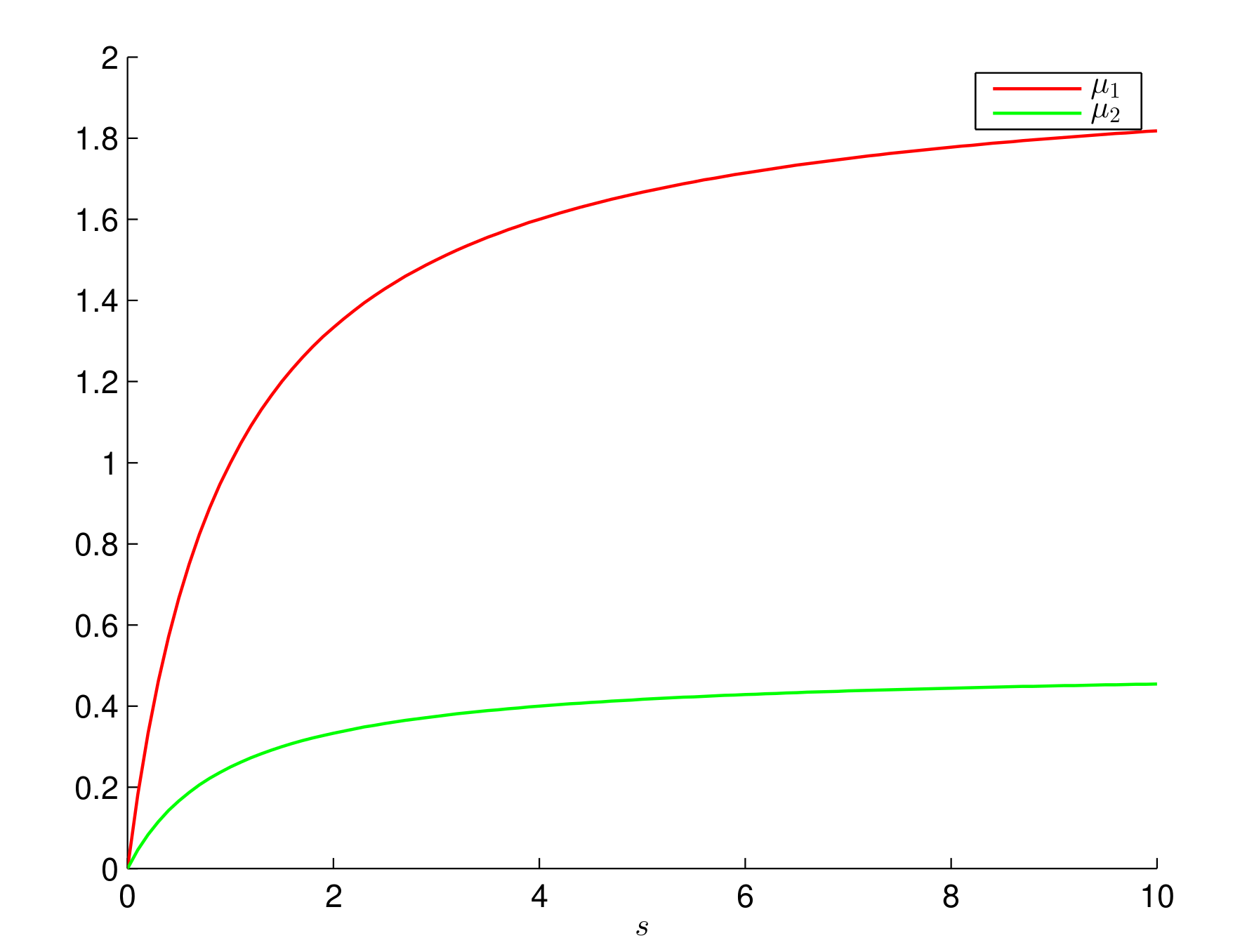
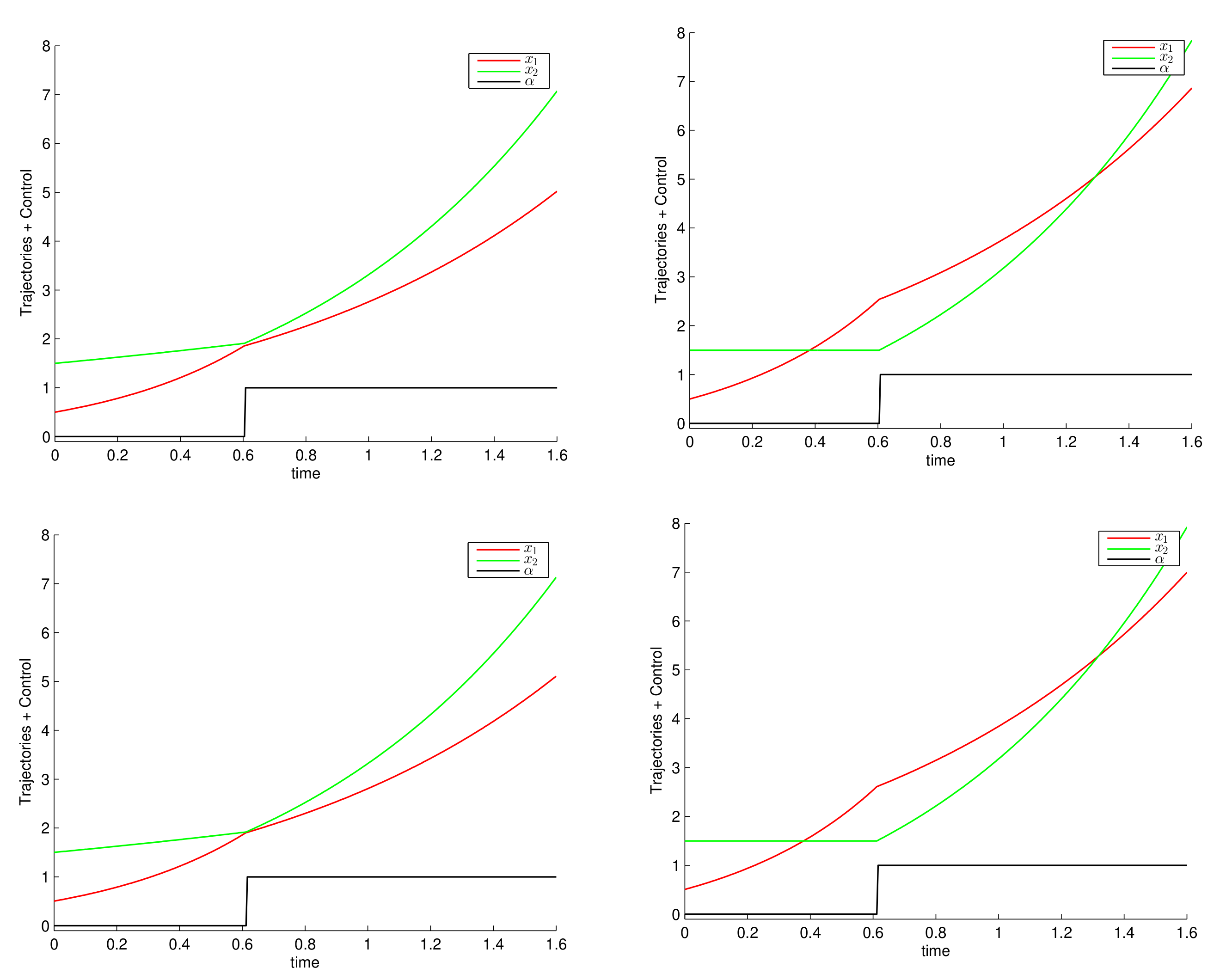
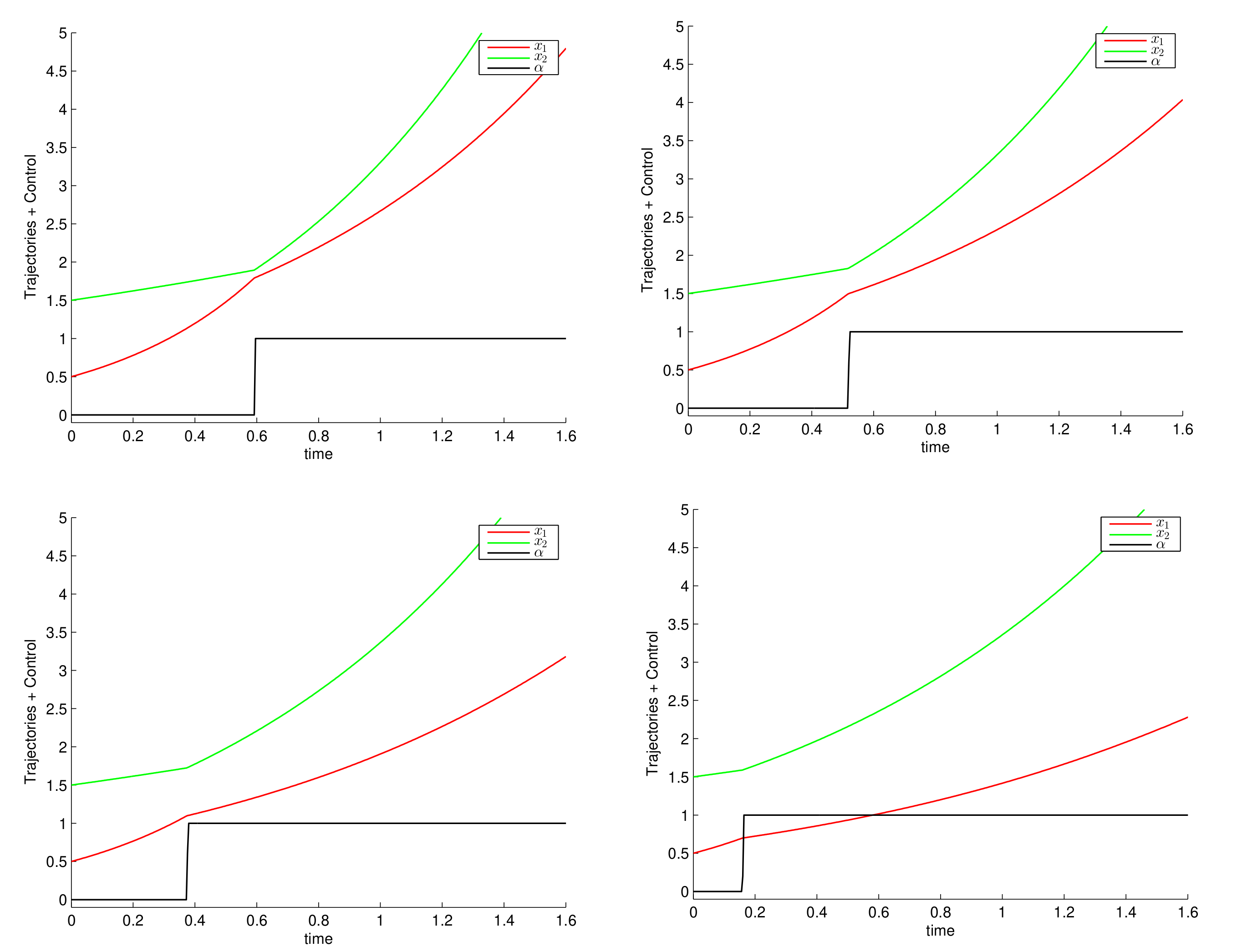
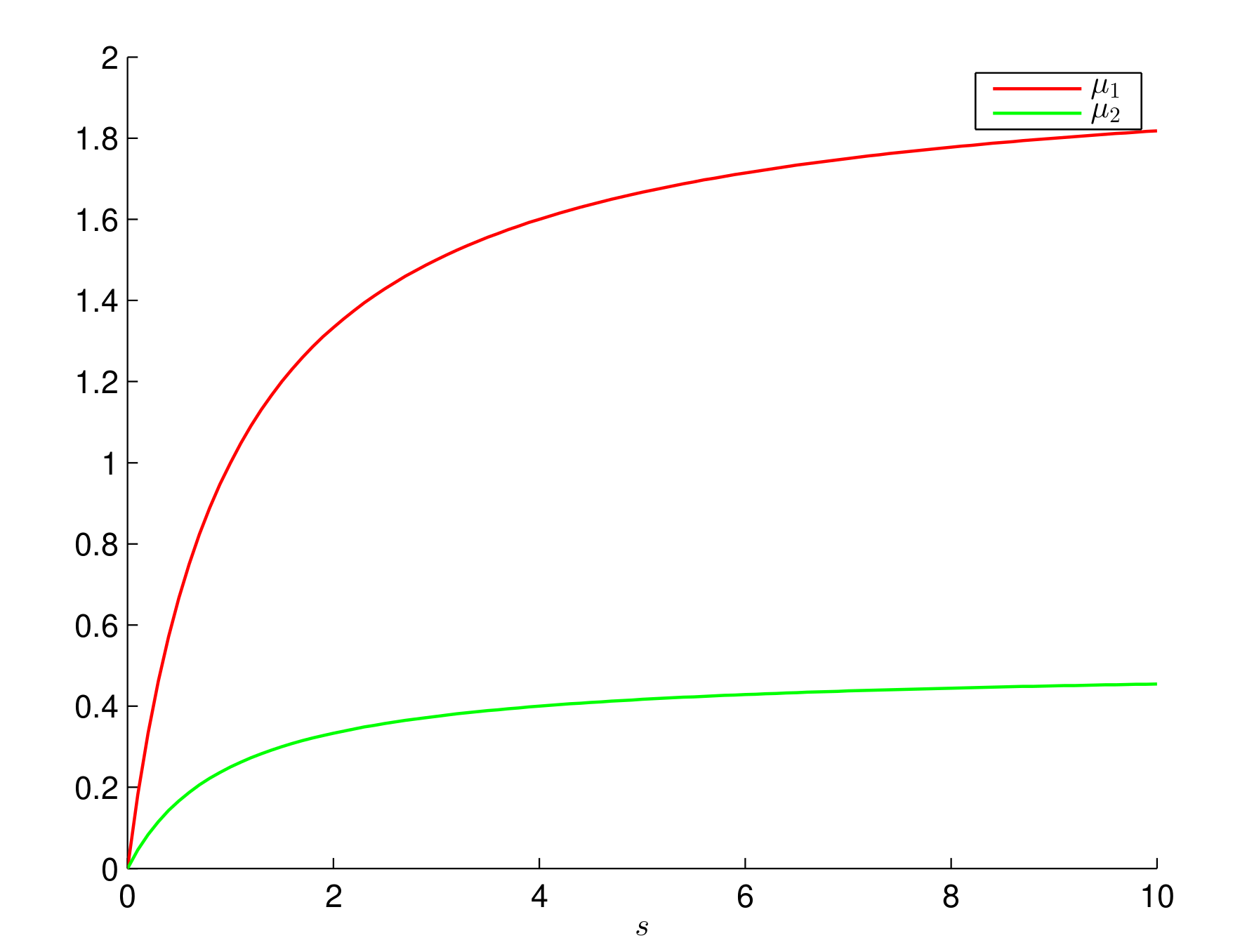
6. Numerical Simulations and Discussion
6.1. Constant Growth Rates
6.2. Monod Growth Rates
6.3. Consequences for the Control of Electro-Fermentation Experiments
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Logan, B.E.; Rabaey, K. Conversion of Wastes into Bioelectricity and Chemicals by Using Microbial Electrochemical Technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Moscoviz, R.; Toledo-Alarcon, J.; Trably, E.; Bernet, N. Electro-Fermentation: How To Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Schievano, A.; Pepé Sciarria, T.; Vanbroekhoven, K.; De Wever, H.; Puig, S.; Andersen, S.J.; Rabaey, K.; Pant, D. Electro-Fermentation—Merging Electrochemistry with Fermentation in Industrial Applications. Trends Biotechnol. 2016, 34, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Freguia, S.; Dennis, P.G.; Keller, J.; Rabaey, K. Development of bioelectrocatalytic activity stimulates mixed-culture reduction of glycerol in a bioelectrochemical system. Microb. Biotechnol. 2015, 8, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Xafenias, N.; Anunobi, M.O.; Mapelli, V. Electrochemical startup increases 1,3-propanediol titers in mixed-culture glycerol fermentations. Process Biochem. 2015, 50, 1499–1508. [Google Scholar] [CrossRef] [Green Version]
- Moscoviz, R.; Trably, E.; Bernet, N. Electro-fermentation triggering population selection in mixed-culture glycerol fermentation. Microb. Biotechnol. 2018, 11, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Alarcón, J.; Moscoviz, R.; Trably, E.; Bernet, N. Glucose electro-fermentation as main driver for efficient H2-producing bacteria selection in mixed cultures. Int. J. Hydrog. Energy 2019, 44, 2230–2238. [Google Scholar] [CrossRef]
- Paiano, P.; Premier, G.; Guwy, A.; Kaur, A.; Michie, I.; Majone, M.; Villano, M. Simplified Reactor Design for Mixed Culture-Based Electrofermentation toward Butyric Acid Production. Processes 2021, 9, 417. [Google Scholar] [CrossRef]
- Emde, R.; Schink, B. Oxidation of glycerol, lactate, and propionate by Propionibacterium freudenreichii in a poised-potential amperometric culture system. Arch. Microbiol. 1990, 153, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Choi, O.; Um, Y.; Sang, B.I. Butyrate production enhancement by Clostridium tyrobutyricum using electron mediators and a cathodic electron donor. Biotechnol. Bioeng. 2012, 109, 2494–2502. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Kim, T.; Woo, H.M.; Um, Y. Electricity-driven metabolic shift through direct electron uptake by electroactive heterotroph Clostridiumpasteurianum. Sci. Rep. 2015, 4, 6961. [Google Scholar] [CrossRef] [PubMed]
- Kracke, F.; Virdis, B.; Bernhardt, P.V.; Rabaey, K.; Krömer, J.O. Redox dependent metabolic shift in Clostridium autoethanogenum by extracellular electron supply. Biotechnol. Biofuels 2016, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.Y.; Kim, C.; Ainala, S.K.; Bae, H.; Jeon, B.H.; Park, S.; Kim, J.R. Metabolic shift of Klebsiella pneumoniae L17 by electrode-based electron transfer using glycerol in a microbial fuel cell. Bioelectrochemistry 2019, 125, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Holtmann, D.; Ulber, R.; Tippkötter, N. Increased Biobutanol Production by Mediator-Less Electro-Fermentation. Biotechnol. J. 2019, 14, 1800514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Meng, J.; Sun, K.; Yan, H. A neutral red mediated electro-fermentation system of Clostridium beijerinckii for effective co-production of butanol and hydrogen. Bioresour. Technol. 2021, 332, 125097. [Google Scholar] [CrossRef] [PubMed]
- Harmand, J.; Lobry, C.; Rapaport, A.; Sari, T. Optimal Control in Bioprocesses: Pontryagin’s Maximum Principle in Practice; Chemical Engineering Series—Chemostat and Bioprocesses; Wiley: Hoboken, NJ, USA, 2019; Volume 3, p. 244. [Google Scholar]
- Smith, H.; Waltman, P. The Theory of the Chemostat: Dynamics of Microbial Competition; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Pontryagin, L.; Boltyanski, V.; Gamkrelidze, R.; Michtchenko, E. The Mathematical Theory of Optimal Processes; The Macmillan Company: New York, NY, USA, 1964. [Google Scholar]
- Haidar, I.; Rapaport, A.; Gérard, F. Effects of spatial structure and diffusion on the performances of the chemostat. Math. Biosci. Eng. 2011, 8, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.B.; Markus, L. Foundations of Optimal Control Theory; John Wiley: Hoboken, NJ, USA, 1967. [Google Scholar]
- Zelikin, M.I.; Borisov, V.F. Theory of Chattering Control: With Applications to Astronautics, Robotics, Economics, and Engineering; Systems & Control: Foundations & Applications; Birkhaüser: Basel, Switzerland, 1994. [Google Scholar]
- Bonnans, F.; Martinon, P.; Giorgi, D.; Grélard, V.; Maindrault, S.; Tissot, O.; Liu, J. BOCOP 2.0. 5—User Guide. 2017, p. 480. Available online: http://bocop.org (accessed on 28 January 2022).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | ||||||
|---|---|---|---|---|---|---|
| 2 | 1/2 | 1 | 0.1–0.5 | 1 | 1 | 2 |
| T | ||||||||
|---|---|---|---|---|---|---|---|---|
| 2 | 1/2 | 1 | 1 | 1 | 0.1 | 1 | 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haidar, I.; Desmond-Le Quéméner, E.; Barbot, J.-P.; Harmand, J.; Rapaport, A. Modeling and Optimal Control of an Electro-Fermentation Process within a Batch Culture. Processes 2022, 10, 535. https://doi.org/10.3390/pr10030535
Haidar I, Desmond-Le Quéméner E, Barbot J-P, Harmand J, Rapaport A. Modeling and Optimal Control of an Electro-Fermentation Process within a Batch Culture. Processes. 2022; 10(3):535. https://doi.org/10.3390/pr10030535
Chicago/Turabian StyleHaidar, Ihab, Elie Desmond-Le Quéméner, Jean-Pierre Barbot, Jérôme Harmand, and Alain Rapaport. 2022. "Modeling and Optimal Control of an Electro-Fermentation Process within a Batch Culture" Processes 10, no. 3: 535. https://doi.org/10.3390/pr10030535
APA StyleHaidar, I., Desmond-Le Quéméner, E., Barbot, J.-P., Harmand, J., & Rapaport, A. (2022). Modeling and Optimal Control of an Electro-Fermentation Process within a Batch Culture. Processes, 10(3), 535. https://doi.org/10.3390/pr10030535