


Fumaric Acid Production by R. arrhizus NRRL 1526 Using Apple Pomace Enzymatic Hydrolysates: Kinetic Modelling

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Wastes
2.2. Enzymes and Reagents
2.3. Enzymatic Hydrolysis
2.4. Detoxification
2.5. Microorganism and Stock Elaboration
2.6. Media and Culture Conditions
2.7. Biomass and Ammonium Tracing
2.8. Sample Analysis
2.9. Mathematical Methods
2.10. Kinetic Model
3. Results and Discussion
3.1. Enzymatic Hydrolysis
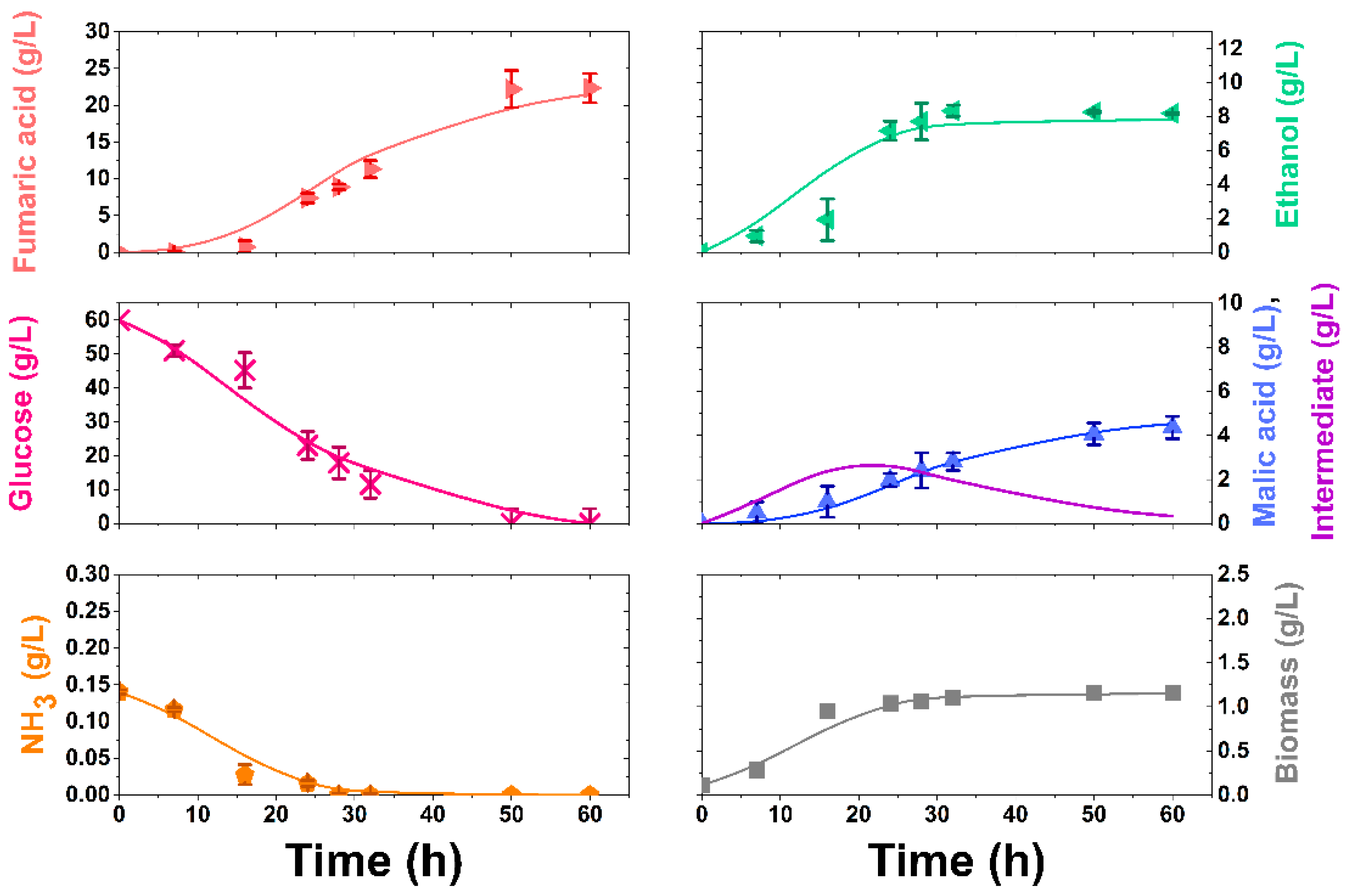
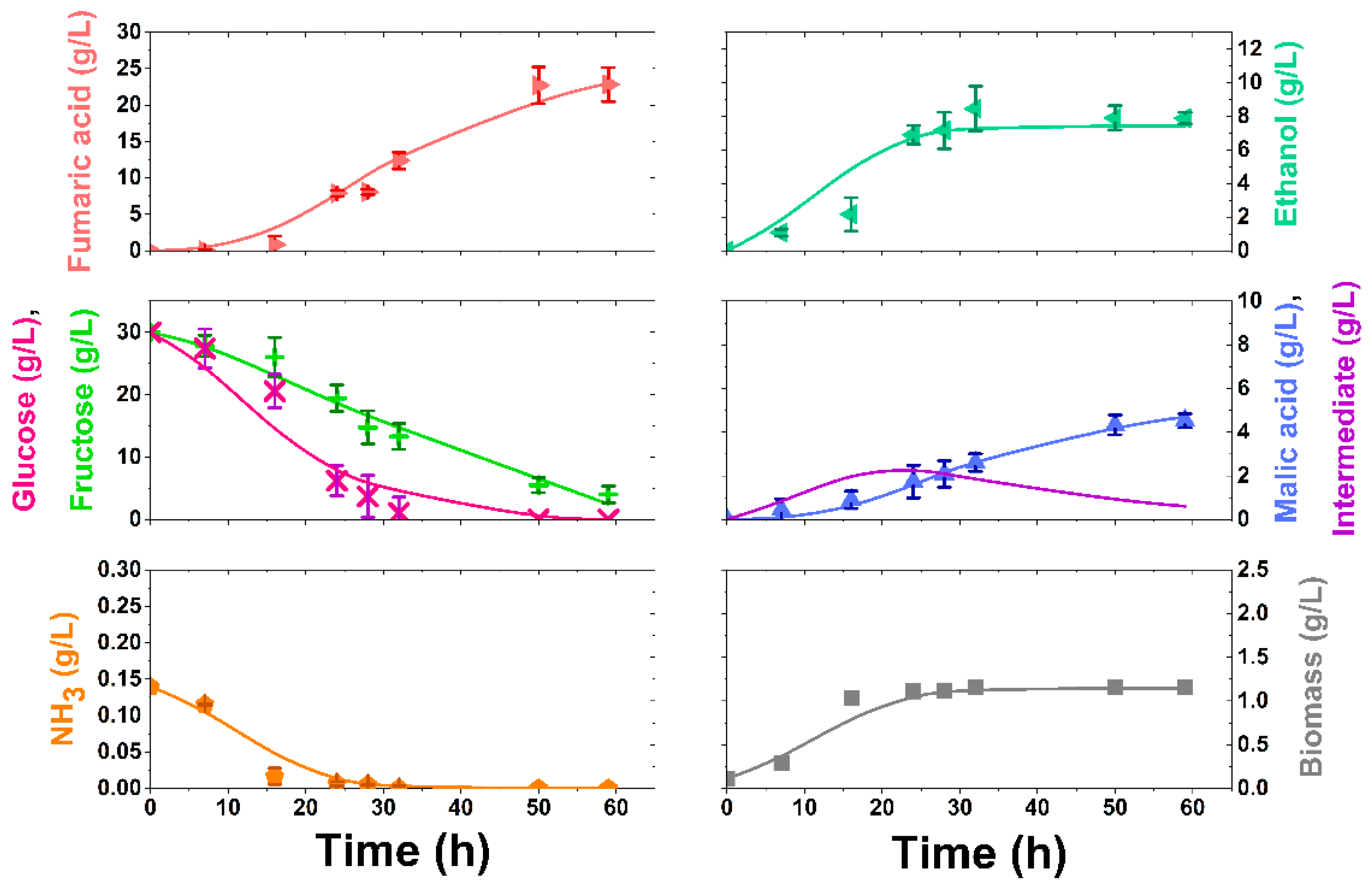
3.2. Synthetic Media Experiments
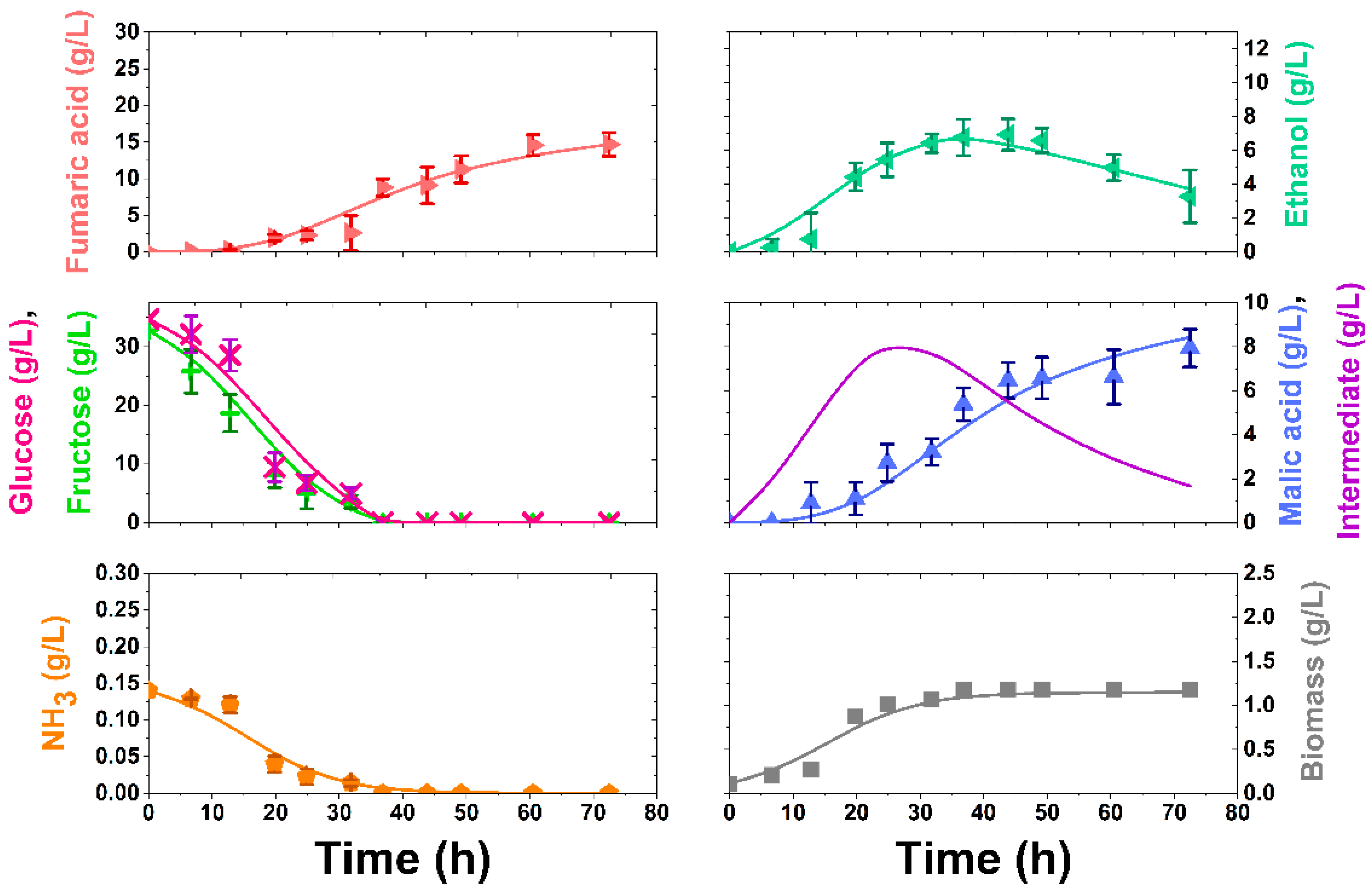
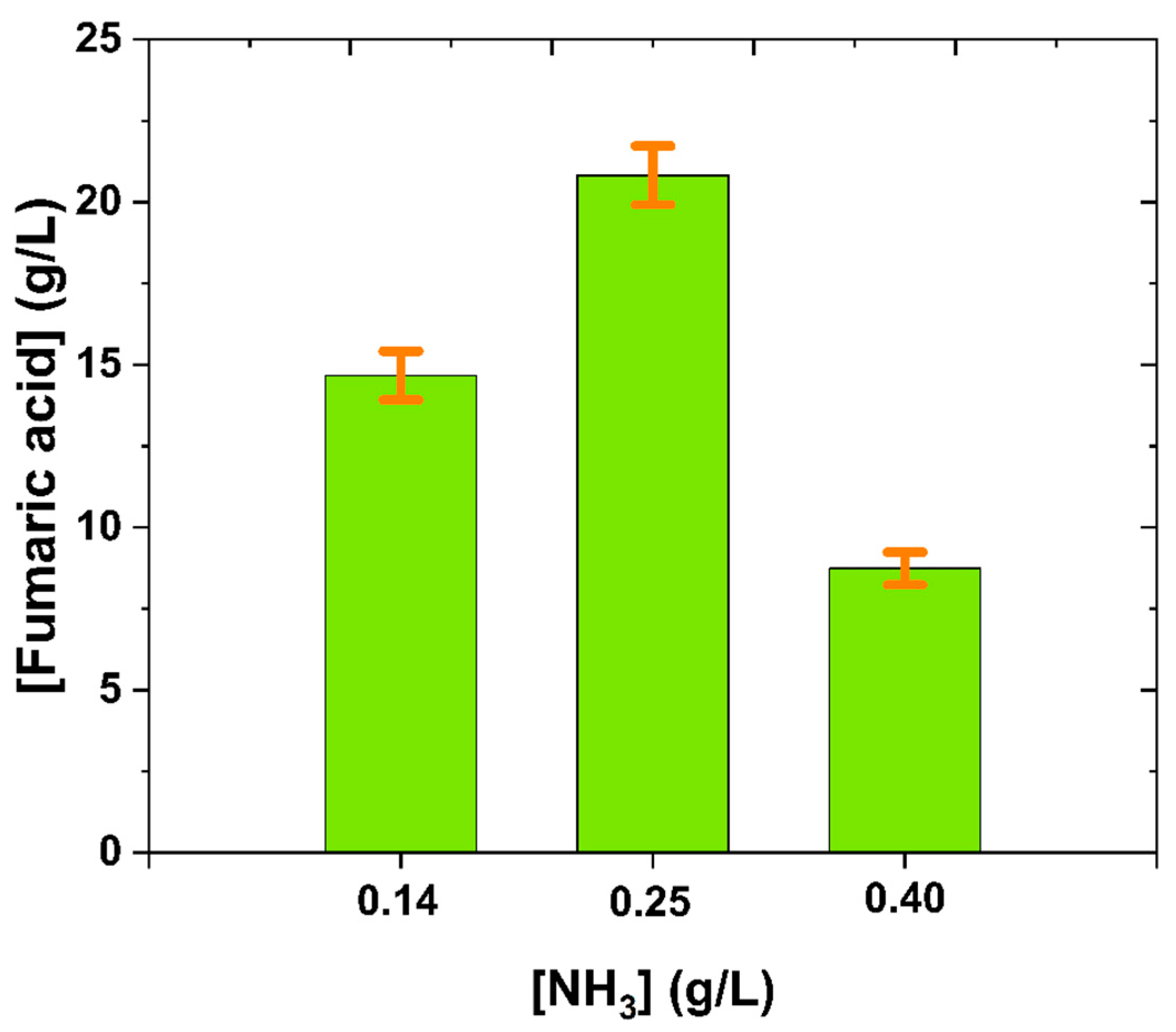
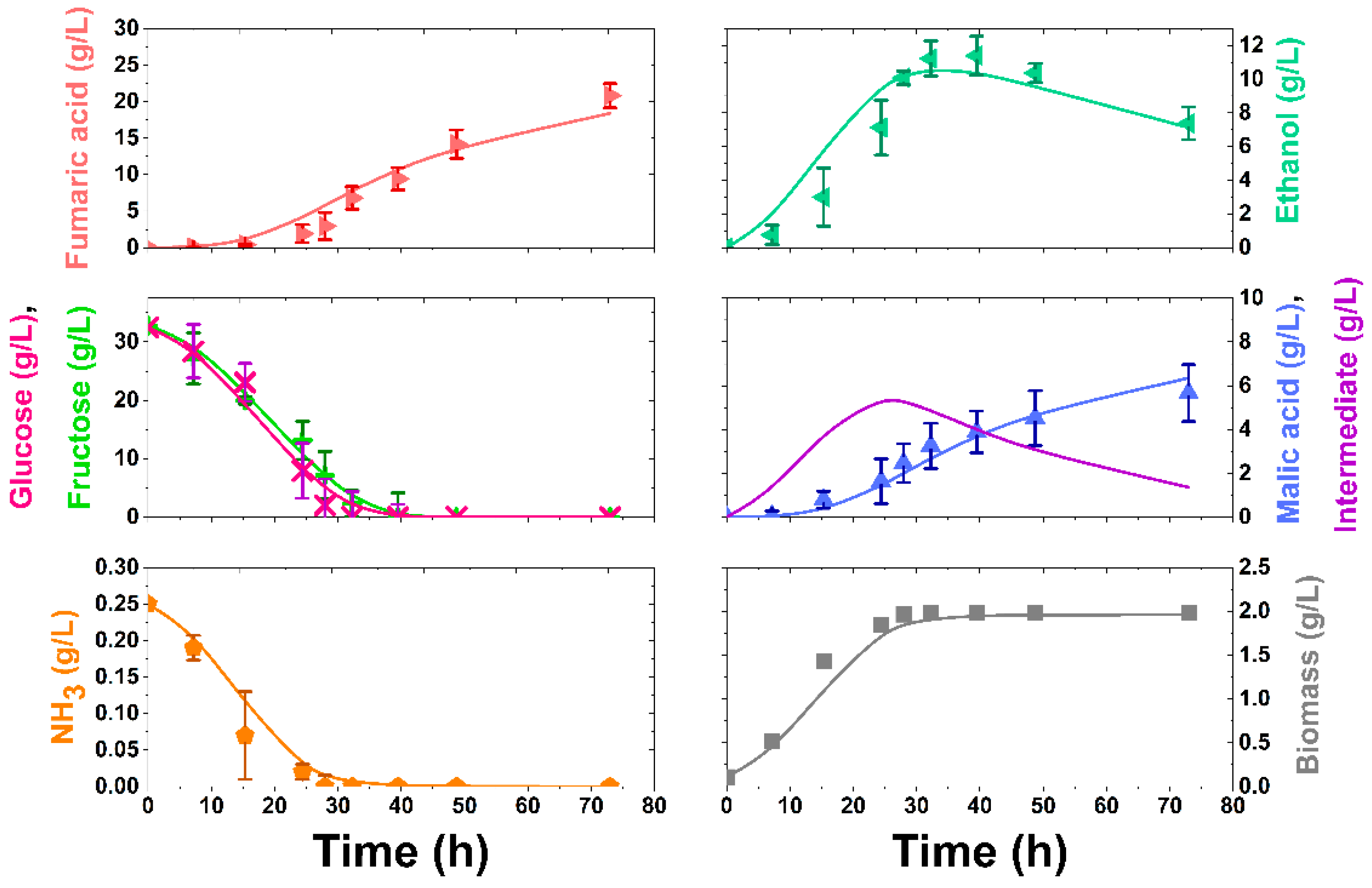
3.3. APH Application
3.4. Kinetic Modelling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portada—Desarrollo Sostenible. Available online: https://www.un.org/sustainabledevelopment/es/ (accessed on 17 May 2022).
- Mat Aron, N.S.; Khoo, K.S.; Chew, K.W.; Show, P.L.; Chen, W.H.; Nguyen, T.H.P. Sustainability of the four generations of biofuels—A review. Int. J. Energy Res. 2020, 44, 9266–9282. [Google Scholar] [CrossRef]
- Batten, S. Climate Change and the Macro-Economy: A Critical Review. SSRN Electron. J. 2018. Staff Working Paper No. 706. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3104554 (accessed on 17 May 2022). [CrossRef]
- Moral-Zamorano, M.; Quijada-Garrido, I.; San-Miguel, V.; Serrano, B.; Baselga, J.; Hashmi, S.; Stadler, F.J.; García-Peñas, A. Concentration effect over thermoresponse derived from organometallic compounds of functionalized poly(N-isopropylacrylamide-co-dopamine methacrylamide). Polymers 2021, 13, 3921. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, V.; Rodríguez, A.; Ladero, M.; Santos, V.E. High 2,3-butanediol production from glycerol by Raoultella terrigena CECT 4519. Bioprocess Biosyst. Eng. 2020, 43, 685–692. [Google Scholar] [CrossRef]
- Esteban, J.; Ladero, M. Food waste as a source of value-added chemicals and materials: A biorefinery perspective. Int. J. Food Sci. Technol. 2018, 53, 1095–1108. [Google Scholar] [CrossRef]
- Nanda, S.; Rana, R.; Sarangi, P.K.; Dalai, A.K.; Kozinski, J.A. A Broad Introduction to First-, Second-, and Third-Generation Biofuels. In Recent Advancements in Biofuels and Bioenergy Utilization; Sarangi, P., Nanda, S., Mohanty, P., Eds.; Springer: Singapore, 2018; pp. 1–25. [Google Scholar] [CrossRef]
- Straathof, A.J.J.; Wahl, S.A.; Benjamin, K.R.; Takors, R.; Wierckx, N.; Noorman, H.J. Grand research challenges for sustainable industrial biotechnology. Trends Biotechnol. 2019, 37, 1042–1050. [Google Scholar] [CrossRef]
- Garcia-Montalvo, J.; Garcia-Martin, A.; Ibañez-Bujan, J.; Santos, V.E.; Yustos, P.; Bolivar, J.M.; Ladero, M. Extraction of antioxidants from grape and apple pomace: Solvent selection and process kinetics. Appl. Sci. 2022, 12, 4901. [Google Scholar] [CrossRef]
- Arias, A.; Feijoo, G.; Moreira, M.T. Exploring the potential of antioxidants from fruits and vegetables and strategies for their recovery. Innov. Food Sci. Emerg. Technol. 2022, 77, 102974. [Google Scholar] [CrossRef]
- Perussello, C.A.; Zhang, Z.; Marzocchella, A.; Tiwari, B.K. Valorization of apple pomace by extraction of valuable compounds. Compr. Rev. Food Sci. Food Saf. 2017, 16, 776–796. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab.: Golden, CO, USA, 2004. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, J.; Tan, T.; Liu, L. Production of fumaric acid from l-malic acid by solvent engineering using a recombinant thermostable fumarase from Thermus thermophilus HB8. Appl. Biochem. Biotechnol. 2015, 175, 2823–2831. [Google Scholar] [CrossRef] [PubMed]
- El-Enshasy, H.A. Filamentous Fungal Cultures-Process Characteristics, Products, and Applications. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.-T., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2007; pp. 225–261. [Google Scholar] [CrossRef]
- Zhang, K. Fumaric Acid Fermentation by Rhizopus oryzae with Integrated Separation Technologies. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2012; pp. 1–210. [Google Scholar]
- Martin-Dominguez, V.; Estevez, J.; De Borja Ojembarrena, F.; Santos, V.E.; Ladero, M. Fumaric acid production: A biorefinery perspective. Fermentation 2018, 4, 33. [Google Scholar] [CrossRef]
- Martin-Dominguez, V.; Bouzas-Santiso, L.; Martinez-Peinado, N.; Santos, V.E.; Ladero, M. Kinetic modelling of the coproduction process of fumaric and malic acids by Rhizopus arrhizus NRRL 1526. Processes 2020, 8, 188. [Google Scholar] [CrossRef]
- Xu, Q.; Li, S.; Huang, H.; Wen, J. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol. Adv. 2012, 30, 1685–1696. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process. Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Jiménez-Quero, A.; Pollet, E.; Zhao, M.; Marchioni, E.; Avérous, L.; Phalip, V. Fungal fermentation of lignocellulosic biomass for itaconic and fumaric acid production. J. Microbiol. Biotechnol. 2017, 27, 1–8. [Google Scholar] [CrossRef]
- Gołebiewska, E.; Kalinowska, M.; Yildiz, G. Sustainable use of apple pomace (AP) in different industrial sectors. Materials 2022, 15, 1788. [Google Scholar] [CrossRef]
- Vukušić, J.L.; Millenautzki, T.; Cieplik, R.; Obst, V.; Saaid, A.M.; Clavijo, L.; Zlatanovic, S.; Hof, J.; Mösche, M.; Barbe, S. Reshaping apple juice production into a zero discharge biorefinery process. Waste Biomass Valorization 2021, 12, 3617–3627. [Google Scholar] [CrossRef]
- de la Torre, I.; Ladero, M.; Santos, V.E. D-lactic acid production from orange waste enzymatic hydrolysates with L. delbrueckii cells in growing and resting state. Ind. Crop. Prod. 2020, 146, 112176. [Google Scholar] [CrossRef]
- Senit, J.J.; Velasco, D.; Gomez Manrique, A.; Sanchez-Barba, M.; Toledo, J.M.; Santos, V.E.; Garcia-Ochoa, F.; Yustos, P.; Ladero, M. Orange peel waste upstream integrated processing to terpenes, phenolics, pectin and monosaccharides: Optimization approaches. Ind. Crop. Prod. 2019, 134, 370–381. [Google Scholar] [CrossRef]
- Krull, S.; Eidt, L.; Hevekerl, A.; Kuenz, A.; Prüße, U. Itaconic acid production from wheat chaff by Aspergillus terreus. Process. Biochem. 2017, 63, 169–176. [Google Scholar] [CrossRef]
- Grande-Tovar, C.D.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan coatings enriched with essential oils: Effects on fungi involve in fruit decay and mechanisms of action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Chandel, A.K.; Kapoor, R.K.; Singh, A.; Kuhad, R.C. Detoxification of sugarcane bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501. Bioresour. Technol. 2007, 98, 1947–1950. [Google Scholar] [CrossRef] [PubMed]
- Martin-Dominguez, V.; Aleman-Cabrera, P.I.; Eidt, L.; Pruesse, U.; Kuenz, A.; Ladero, M.; Santos, V.E. Production of fumaric acid by Rhizopus arrhizus NRRL 1526: A Simple Production Medium and the Kinetic Modelling of the Bioprocess. Fermentation 2022, 8, 64. [Google Scholar] [CrossRef]
- Rhodes, R.A.; Moyer, A.J.; Smith, M.L.; Kelley, S.E. Production of fumaric acid by Rhizopus arrhizus. Appl. Microbiol. 1959, 7, 74–80. [Google Scholar] [CrossRef]
- Ling, L.; Ng, T. Fermentation Process for Carboxylic Acids. U.S. Patent 4,877,731, 31 October 1989. [Google Scholar]
- Ribeiro, L.F.; Ribani, R.H.; Francisco, T.M.G.; Soares, A.A.; Pontarolo, R.; Haminiuk, C.W.I. Profile of bioactive compounds from grape pomace (Vitis vinifera and Vitis labrusca) by spectrophotometric, chromatographic and spectral analyses. J. Chromatogr. B 2015, 1007, 72–80. [Google Scholar] [CrossRef]
- Ozturk, B.; Parkinson, C.; Gonzalez-Miquel, M. Extraction of polyphenolic antioxidants from orange peel waste using deep eutectic solvents. Sep. Purif. Technol. 2018, 206, 2384–2393. [Google Scholar] [CrossRef]
- Dennis, J.E.; Gay, D.M.; Welsch, R.E. An adaptative nonlinear least-squares algorithm. ACM Trans. Math. Softw. 1981, 7, 348–368. [Google Scholar] [CrossRef]
- Yepes, C.; Estévez, J.; Arroyo, M.; Ladero, M. Immobilization of an industrial β-glucosidase from Aspergillus fumigatus and its use for cellobiose hydrolysis. Processes 2022, 10, 1225. [Google Scholar] [CrossRef]
- Esteban, J.; Fuente, E.; Blanco, A.; Ladero, M.; Garcia-Ochoa, F. Phenomenological kinetic model of the synthesis of glycerol carbonate assisted by focused beam reflectance measurements. Chem. Eng. J. 2015, 260, 434–443. [Google Scholar] [CrossRef]
- Ichikawa, S.; Iino, T.; Sato, S.; Nakahara, T.; Mukataka, S. Improvement of production rate and yield of fumaric acid from maleic acid by heat treatment of Pseudomonas alcaligenes strain XD-1. Biochem. Eng. J. 2003, 13, 7–13. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Improved continuous fumaric acid production with immobilised Rhizopus oryzae by implementation of a revised nitrogen control strategy. New Biotechnol. 2018, 44, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Swart, R.M.; Brink, H.; Nicol, W. Rhizopus oryzae for fumaric acid production: Optimising the use of a synthetic lignocellulosic hydrolysate. Fermentation 2022, 8, 278. [Google Scholar] [CrossRef]
- Swart, R.M.; Ronoh, D.K.; Brink, H.; Nicol, W. Continuous production of fumaric acid with immobilised Rhizopus oryzae: The role of ph and urea addition. Catalysts 2022, 12, 82. [Google Scholar] [CrossRef]
- Swart, R.M.; Roux, F.; Naude, A.; de Jongh, N.W.; Nicol, W. Fumarate production with Rhizopus oryzae: Utilising the Crabtree effect to minimise ethanol by-product formation. Biotechnol. Biofuels 2020, 13, 22. [Google Scholar] [CrossRef]
- Rana, B.K.; Singh, U.P.; Taneja, V. Antifungal activity and kinetics of inhibition by essential oil isolated from leaves of Aegle marmelos. J. Ethnopharmacol. 1997, 57, 29–34. [Google Scholar] [CrossRef]
- Raj, T.; Chandrasekhar, K.; Morya, R.; Kumar Pandey, A.; Jung, J.H.; Kumar, D.; Singhania, R.R.; Kim, S.H. Critical challenges and technological breakthroughs in food waste hydrolysis and detoxification for fuels and chemicals production. Bioresour. Technol. 2022, 360, 127512. [Google Scholar] [CrossRef]
- Tavares, A.P.M.; Gonçalves, M.J.A.; Brás, T.; Pesce, G.R.; Xavier, A.M.R.B.; Fernandes, M.C. Cardoon hydrolysate detoxification by activated carbon or membranes system for bioethanol production. Energies 2022, 15, 1993. [Google Scholar] [CrossRef]
- Yin, X.; Li, J.; Shin, H.-D.; Du, G.; Liu, L.; Chen, J. Metabolic engineering in the biotechnological production of organic acids in the tricarboxylic acid cycle of microorganisms: Advances and prospects. Biotechnol. Adv. 2015, 33, 830–841. [Google Scholar] [CrossRef]
- Tsen, H.Y.; Tsai, S.Y. Comparison of the kinetics and factors affecting the stabilities of chitin-immobilized naringinases from two fungal sources. J. Ferment. Technol. 1988, 66, 193–198. [Google Scholar] [CrossRef]
- Ding, Y.; Li, S.; Dou, C.; Yu, Y.; He, H. Production of fumaric acid by Rhizopus oryzae: Role of carbon-nitrogen ratio. Appl. Biochem. Biotechnol. 2011, 164, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, Z.; Deng, L.; Tan, T.; Wang, F.; Yan, Y. Pull-in urea cycle for the production of fumaric acid in Escherichia coli. Appl. Microbiol. Biotechnol. 2015, 99, 5033–5044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, S.T. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing fumR gene on cell growth and fumaric acid biosynthesis from glucose. Process. Biochem. 2012, 47, 2159–2165. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, S.T. In situ recovery of fumaric acid by intermittent adsorption with IRA-900 ion exchange resin for enhanced fumaric acid production by Rhizopus oryzae. Biochem. Eng. J. 2015, 96, 38–45. [Google Scholar] [CrossRef]
- Sebastian, J.; Rouissi, T.; Brar, S.K. Miscanthus sp.—Perennial lignocellulosic biomass as feedstock for greener fumaric acid bioproduction. Ind. Crop. Prod. 2022, 175, 114248. [Google Scholar] [CrossRef]
- Deng, F.; Aita, G.M. Fumaric acid production by Rhizopus oryzae ATCC® 20344TM from lignocellulosic syrup. Bioenergy Res. 2018, 11, 330–340. [Google Scholar] [CrossRef]
- Papadaki, A.; Papapostolou, H.; Alexandri, M.; Kopsahelis, N.; Papanikolaou, S.; de Castro, A.M.; Freire, D.M.G.; Koutinas, A.A. Fumaric acid production using renewable resources from biodiesel and cane sugar production processes. Environ. Sci. Pollut. Res. 2018, 25, 35960–35970. [Google Scholar] [CrossRef]
- Groff, M.C.; Scaglia, G.; Gaido, M.; Kassuha, D.; Ortiz, O.A.; Noriega, S.E. Kinetic modeling of fungal biomass growth and lactic acid production in Rhizopus oryzae fermentation by using grape stalk as a solid substrate. Biocatal. Agric. Biotechnol. 2022, 39, 102255. [Google Scholar] [CrossRef]
- Ajala, E.O.; Ajala, M.A.; Onoriemu, O.O.; Akinpelu, S.G.; Bamidele, S.H. Lactic acid production: Utilization of yam peel hydrolysate as a substrate using Rhizopus orysae in kinetic studies. Biofuels Bioprod. Biorefin. 2021, 15, 1031–1045. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Before Detoxification | Detoxified APH | Units |
|---|---|---|---|
| [Sugars] | 68.12 | 65.29 | g/L |
| [Glucose] | 35.11 | 32.48 | g/L |
| [Fructose] | 33.01 | 32.81 | g/L |
| Total phenolic content | 587.86 | 336.32 | mgeq.gallic acid/L |
| Antioxidant activity | 67.69 | 53.77 | TEAC/L |
| [NH4+] | 105.17 | <10 * | mg/L |
| [Soluble protein] | − | 0.42 | g/L |
| Experiment | Medium | [Fumaric Acid] (g/L) | Yield (gFumaric acid/gconsumed sugar) | Specific Productivity (gFumaric acid/(gBiomass·h)) |
|---|---|---|---|---|
| 1 | Glucose: 60 g/L NH3: 0.14 g/L | 22.20 | 0.37 | 0.32 |
| 2 | Glucose: 30 g/L Fructose: 30 g/L NH3: 0.14 g/L | 22.70 | 0.38 | 0.33 |
| 3 | APH NH3: 0.14 g/L | 14.67 | 0.23 | 0.17 |
| 4 | APH NH3: 0.25 g/L | 20.82 | 0.32 | 0.14 |
| Reference | Microorganism | Medium | [Fumaric Acid] (g/L) | Production Yield (gfumaric acid/gsugar) |
|---|---|---|---|---|
| [52] | Rhizopus oryzae NRRL 2582 | Silver grass hydrolysate: Sugars ≈ 55 g/L; Yeast extract: 0.5 g/L | 9.26 | 0.17 |
| [53] | Rhizopus oryzae ATCC 20344 | Lignocellulosic syrup: Sugars ≈ 80 g/L; Urea: 0.2 g/L | 34.20 | 0.43 |
| [54] | Rhizopus oryzae NRRL 2582 | Sugar cane molasses with soybean cake hydrolysate: Sugars ≈ 25 g/L; free amino nitrogen: 0.2 g/L | 7.90 | 0.32 |
| Present work | Rhizopus arrhizus NRRL 1526 | Apple pomace hydrolysate: Sugars ≈ 60 g/L; (NH4)2SO4: 0.25 g/L | 20.82 | 0.32 |
| Parameter | Units | Experiment 1 | Experiment 2 | Experiment 3 | Experiment 4 |
|---|---|---|---|---|---|
| μm | 1.20 ± 0.07 | 1.28 ± 0.08 | 0.96 ± 0.04 | 0.76 ± 0.05 | |
| kF | (2.64 ± 0.40)·10−1 | (2.87 ± 0.34)·10−1 | (0.74 ± 0.05)·10−1 | (0.48 ± 0.05)·10−1 | |
| kE | − | − | (8.14 ± 1.42)·10−2 | (5.14 ± 2.20)·10−2 | |
| mS | (5.94 ± 2.67)·10−1 | (3.14 ± 0.38)·10−1 | (5.36 ± 0.26)·10−1 | (3.25 ± 2.81)·10−1 | |
| YFrEn | − | 1.25 ± 0.13 | 1.93 ± 0.10 | 1.27 ± 1.06 | |
| YMF | (2.11 ± 0.16)·10−1 | (2.04 ± 0.16)·10−1 | (5.61 ± 0.34)·10−1 | (3.45 ± 0.79)·10−1 | |
| YEX | 7.52 ± 0.36 | 7.17 ± 0.34 | 6.88 ± 0.29 | 5.86 ± 0.39 | |
| YGX | 30.40 ± 8.85 | 20.57 ± 2.95 | 21.95 ± 1.19 | 10.57 ± 4.47 | |
| YFrX | − | 5.97 ± 2.84 | 9.47 ± 1.19 | 8.17 ± 4.41 | |
| YIX | 4.86 ± 0.68 | 3.49 ± 0.46 | 6.93 ± 0.80 | 4.19 ± 0.47 | |
| YIF | (2.19 ± 0.16)·10−1 | (1.31 ± 0.32)·10−1 | (3.49 ± 0.88)·10−1 | (3.45 ± 0.79)·10−1 |
| Parameter | Experiment 1 | Experiment 2 | Experiment 3 | Experiment 4 |
|---|---|---|---|---|
| RMSE | 0.088 | 0.086 | 0.089 | 0.127 |
| F95 | 195430 | 95366 | 114079 | 37696 |
| %VE | 96.28 | 95.50 | 96.55 | 96.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Dominguez, V.; Garcia-Montalvo, J.; Garcia-Martin, A.; Ladero, M.; Santos, V.E. Fumaric Acid Production by R. arrhizus NRRL 1526 Using Apple Pomace Enzymatic Hydrolysates: Kinetic Modelling. Processes 2022, 10, 2624. https://doi.org/10.3390/pr10122624
Martin-Dominguez V, Garcia-Montalvo J, Garcia-Martin A, Ladero M, Santos VE. Fumaric Acid Production by R. arrhizus NRRL 1526 Using Apple Pomace Enzymatic Hydrolysates: Kinetic Modelling. Processes. 2022; 10(12):2624. https://doi.org/10.3390/pr10122624
Chicago/Turabian StyleMartin-Dominguez, Victor, Jorge Garcia-Montalvo, Alberto Garcia-Martin, Miguel Ladero, and Victoria E. Santos. 2022. "Fumaric Acid Production by R. arrhizus NRRL 1526 Using Apple Pomace Enzymatic Hydrolysates: Kinetic Modelling" Processes 10, no. 12: 2624. https://doi.org/10.3390/pr10122624
APA StyleMartin-Dominguez, V., Garcia-Montalvo, J., Garcia-Martin, A., Ladero, M., & Santos, V. E. (2022). Fumaric Acid Production by R. arrhizus NRRL 1526 Using Apple Pomace Enzymatic Hydrolysates: Kinetic Modelling. Processes, 10(12), 2624. https://doi.org/10.3390/pr10122624