
Optimization of L-Citrulline Operon in Corynebacterium glutamicum for L-Citrulline Production

1
School of Biological and Food Engineering, Changzhou University, Changzhou 213164, China
2
Zaozhuang Key Laboratory of Corn Bioengineering, Zaozhuang Science and Technology Collaborative Innovation Center of Enzyme, Shandong Hengren Gongmao Co., Ltd., Zaozhuang 277533, China
3
School of Petrochemical Engineering, Changzhou University, Changzhou 213164, China
*
Authors to whom correspondence should be addressed.
Processes 2022, 10(10), 2153; https://doi.org/10.3390/pr10102153
Submission received: 23 September 2022
/
Revised: 13 October 2022
/
Accepted: 19 October 2022
/
Published: 21 October 2022
(This article belongs to the Special Issue Progress in Biorefinery of Lignocellulosic Biomass to Bio-Energies and Bio-Based Chemicals)
Abstract
:L-citrulline plays important roles in many physiological processes, and its application range is expanding rapidly. Corynebacterium glutamicum strains have the potential to be efficient L-citrulline producers. In this study, we performed optimization of L-citrulline biosynthesis operon in C. glutamicum ATCC13032 for L-citrulline production. Chromosomal integration of the integral argBEc gene from Escherichia coli (encoding natively insensitive N-acetylglutamate kinase), the deletion of the argR gene (encoding repressor ArgR), and the deletion of the argG gene (encoding argininosuccinate synthase) were achieved simultaneously by one-step genome modification and by obtaining the L-citrulline-producing strain. Then, plasmid-based overexpression of the optimized L-citrulline operon was carried out and the L-citrulline production was further improved. In fed-batch fermentation, the L-citrulline production and yield from glucose of the final strain reached 26.7 g/L and 0.18 g/g, respectively. These results indicate that optimization of L-citrulline operon in C. glutamicum is effective to construct the L-citrulline over-producing strain.
1. Introduction
L-citrulline plays important roles in many physiological processes, such as the regulation of nitrogen homeostasis, immune-system stimulation, the antioxidant effect, and nitric oxide metabolism. Due to its multiple functions, L-citrulline is widely used in the pharmaceutical, health-care food, and cosmetics industries, and its application range is expanding rapidly [1,2,3].
In recent years, methods for L-citrulline production by enzymatic synthesis and microbial fermentation have been reported. The enzymatic synthesis uses arginine deiminase for the hydrolysis of the arginine guanidinium group [2,3]. In the enzymatic method for L-citrulline production, L-arginine is the substrate or raw material. However, L-arginine is produced by microbial fermentation at present, and the L-arginine production strains are ordinarily derived from Corynebacterium glutamicum and its related species [4,5,6]. In C. glutamicum strains, L-citrulline is an intermediate metabolite in the L-arginine biosynthetic pathway (Figure 1). During L-arginine production by the microbial fermentation process, in the cells of the L-arginine producing strain, L-arginine is synthesized from L-citrulline by introducing the imino group in the carbamoyl of L-citrulline [4]. However, in an enzymatic method for L-citrulline production, the introduced imino group in L-arginine is hydrolyzed. Therefore, in consideration of the acquisition of the L-arginine substrate, the enzymatic method for L-citrulline production is uneconomical. In comparison, the one-step production of L-citrulline by microbial fermentation is more economical.
C. glutamicum and its related species have been employed for the industrial production of amino acids for several decades, and it is especially known for its ability to synthesis and secrete glutamate [7,8,9,10]. On this basis, C. glutamicum is considered an excellent host for the production of glutamate-derived amino acids, such as L-ornithine and L-arginine [9,10]. The genome of C. glutamicum reveals a rich repertoire of pathways, suggesting attractive properties for industrial production purposes; moreover, a wide range of genetic engineering tools for C. glutamicum engineering are established, and the omics data of C. glutamicum are constantly expanding. Therefore, metabolic engineering on C. glutamicum is likely to obtain a promising cell factory [7,9,10]. As mentioned above, in C. glutamicum strains, L-citrulline is an intermediate metabolite in the L-arginine biosynthetic pathway (Figure 1). In C. glutamicum, the genes involved in L-arginine biosynthesis are organized as the gene cluster argCJBDFRGH and are divided into argCJBDFR and argGH operons [5,11,12]. Additionally, the argCJBDFR operon encodes all of the enzymes required to convert L-glutamate to L-citrulline (Figure 1). The argR gene encodes repressor ArgR of the argCJBDFR and argGH operons. The argG gene encodes argininosuccinate synthase, which catalyzes the conversion of L-citrulline to argininosuccinate (Figure 1). The argB gene encodes N-acetylglutamate kinase (NAGK), which is feedback-inhibited by L-arginine, and the intracellular activity of NAGK is the limitation for L-arginine and L-citrulline synthesis by C. glutamicum [4,13,14]. However, in Escherichia coli, the NAGKEc encoded by argBEc is natively insensitive to L-arginine. In C. glutamicum, the replacement of the chromosomal argB by argBEc could effectively improve L-arginine and L-citrulline production [13]. In C. glutamicum, introducing mutant NAGK that was less sensitive to L-arginine was also beneficial for L-ornithine production [8,15].
In C. glutamicum strains, L-citrulline is an intermediate metabolite between L-ornithine and L-arginine in the L-arginine biosynthetic pathway (Figure 1). Metabolically engineered C. glutamicum could produce 51.5 g/L of L-ornithine with the productivity of 1.29 g/L/h in fed-batch fermentation [15]. Systems metabolic engineering in combination with random mutagenesis on C. glutamicum could result in the efficient production of L-arginine to 92.5 g/L in fed-batch fermentation [5]. Therefore, C. glutamicum strains have the potential to be efficient L-citrulline producers. The inactivation of argininosuccinate synthase (encoded by argG) and repressor ArgR combined with the overexpression of argJ encoding ornithine acetyltransferase in C. glutamicum ATCC13032 resulted in the L-citrulline production of 8.51 g/L [16]. The deletion of argR and argG genes combined with the overexpression of argBfbr (encoding feedback resistant NAGK) and argF (encoding ornithine carbamoylphosphate transferase) in C. glutamicum MB001 resulted in the accumulation of 7.7 g/L of L-citrulline in minimal medium [17].
In this study, we performed the optimization of L-citrulline biosynthesis operon in C. glutamicum for L-citrulline production. Firstly, we inserted the argBEc gene at an appropriate site in gene cluster argCJBDFRGH to achieve the expression of feedback-resistant NAGKEc and the deletion of argR and argG genes. Then, the plasmid-based overexpression of the optimized L-citrulline operon argCJBDFBEc was carried out to further improve L-citrulline biosynthesis.
2. Materials and Methods
2.1. Microorganism and Cultivation Conditions
All strains plasmids [18,19] used in this study are listed in Table 1. E. coli strain JM109 was used as the host for plasmid construction and was cultured in Luria–Bertani (LB) medium at 37 °C. The precultivations of C. glutamicum strains were carried out in LBG medium (LB medium supplemented with 5 g/L glucose) at 30 °C. For L-arginine auxotroph strain precultivation, 0.5 g/L L-arginine was added to the LBG medium. Where appropriate, kanamycin (25 mg/L for C. glutamicum strains, 50 mg/L for E. coli strains) was added to the cultures.
For L-citrulline fermentation in shake flasks, the C. glutamicum strains from agar slants were inoculated in 20 mL seed medium (LBG medium), and 0.5 g/L L-arginine was added for the L-arginine auxotroph strain. Seed cultivation was carried out at 30 °C for 18 h in 250-mL shake flasks with a rotational speed of 220 r/min. The seed culture (1 mL) was transferred into 250-mL shake flasks containing 20 mL fermentation medium. The fermentation medium contained glucose 80 g/L, yeast extract 8 g/L, (NH4)2SO4 40 g/L, KH2PO4 1.5 g/L, MgSO4·7H2O 0.5 g/L, FeSO4·7H2O 0.02 g/L, MnSO4 0.02 g/L, and CaCO3 20 g/L; 0.5 g/L L-arginine was added for the L-arginine auxotroph strain, and the fermentation was carried out at 30 °C with a rotational speed of 220 r/min.
For L-citrulline fed-batch fermentation in the bioreactor, the C. glutamicum strain from agar slants was inoculated in a 50 mL seed medium (LBG medium supplemented with 0.5 g/L L-arginine) and cultured at 30 °C for 18 h in 500-mL shake flasks with a rotational speed of 220 r/min. The seed culture (100 mL) was transferred into 5-L stirred fermenter (BIOTECH-5BG, Baoxing Co., Shanghai, China) containing 1.5 L fermentation medium. The fermentation medium contained glucose 60 g/L, yeast extract 8 g/L, (NH4)2SO4 20 g/L, KH2PO4 1.5 g/L, MgSO4·7H2O 0.5 g/L, FeSO4·7H2O 0.02 g/L, MnSO4 0.02 g/L, and L-arginine 0.5 g/L. The feed medium contained glucose 700 g/L, yeast extract 8 g/L, KH2PO4 1.5 g/L, and MgSO4·7H2O 0.5 g/L and was fed into the fermenter when the residual glucose concentration was below 10 g/L. The temperature was maintained at 30 °C, and the pH was maintained at 7.0 by addition of 30% NH4OH solution. The air flow rate was maintained at 2 L/min, and the agitation speed was 600 r/min.
2.2. Analytical Methods
Cell growth was monitored by measuring the OD600 using a spectrophotometer (UNICO™-UV2000, UNICO (Shanghai) Instrument Co., Ltd., Shanghai, China). For monitoring the cell growth during L-citrulline fermentation in shake flasks, the culture fluid was diluted by 0.2 mol/L HCl to dissolve CaCO3. The glucose was measured using a bioanalyzer (SBA-40C, Biology Institute of Shandong Academy of Sciences, Jinan, China). The amino acids were measured by high-pressure liquid chromatography [20].
2.3. Transcriptional Analysis
The total RNA of C. glutamicum was extracted at the exponential phase using the RNAiso Plus reagent (Takara, Dalian, China), and the synthesis of cDNA was performed using the PrimeScript RT reagent kit (Takara, Dalian, China). Real-time PCR (RT-PCR) was carried out as described previously [21]. The primers used for the RT-PCR analysis of genes in the argCJBDF cluster were referred to the published study [22], and the primers EcBF (5′-GCCGTAGGTTTGTTTCTCGG-3′) and EcBR (5′-CGTTCATCAGTTGCCCTTC-3′) were used for the RT-PCR analysis of the argBEc gene. The 16S rRNA relative abundance was used as an internal standard to standardize the results.
2.4. Construction of Plasmids and Strains
All of the plasmids used in this study are listed in Table 1, and the primers are listed in Table 2. The insertion of argBEc into the C. glutamicum genome was performed via two-step homologous recombination using vector pK18mobsacB [18]. The plasmid pK18RB was used for the insertion of argBEc into the argR gene. To obtain the argRF-argBEc-argRR fragment, the upstream and downstream regions of the argR gene (argRF and argRR) were amplified using the primer combinations RB-RF1/RB-RR1 and RB-RF2/RB-RR2, respectively, from genomic DNA of C. glutamicum ATCC13032; the integral argBEc gene was amplified using the primers RB-BF and RB-BR from the genomic DNA of E. coli BL21; then, the argRF fragment and the argBEc gene were fused in an overlap-extension PCR using the primers RB-RF1 and RB-BR, and the argRF-argBEc fragment was obtained; and finally, the argRF-argBEc and argRR fragments were fused using the primers RB-RF1 and RB-RR2, and the argRF-argBEc-argRR fragment was obtained. This argRF-argBEc-argRR fragment was cloned into pK18mobsacB, resulting in plasmid pK18RB. This plasmid was electro-transformed into C. glutamicum ATCC13032, and the modified strain CRB was screened as described previously [11].
The plasmid pK18RBG was used for the insertion of argBEc between the argR and argG genes. The primers RBG-RF and RBG-RR were used for the amplification of the upstream region of the argR gene (argRF’), the primers RBG-GF and RBG-GR were used for the amplification of downstream region of the argG gene (argGR’), and the primers RBG-BF and RBG-BR were used for amplification of the integral argBEc gene. Overlap-extension PCR was used for obtaining the argRF’-argBEc-argGR’ fragment as described above. This argRF’-argBEc-argGR’ fragment was cloned into pK18mobsacB, resulting in plasmid pK18RBG. Additionally, plasmid pK18RBG was used for CRBG strain construction as described above.
Plasmid pDXW-10 [19] was used for the constitutive expression of the argCJBDF and argCJBDFBEc gene cluster. The argCJBDF gene cluster was amplified using the primers CF and CR from the genomic DNA of C. glutamicum ATCC13032. The resulting DNA fragment was cloned into pDXW-10, resulting in plasmid pDXW-10-argCJBDF. This plasmid was used to transform the CRBG strain using electroporation and to obtain the CRBGC strain. The argCJBDFBEc gene cluster was amplified using the primers CF and CoR from the genomic DNA of the CRBG strain. The resulting DNA fragment was cloned into pDXW-10, and the restriction enzyme NheI was used to validate the direction of gene ligation. The correct ligating plasmid was named pDXW-10-argCJBDFBEc. This plasmid was used to transform the CRBG strain using electroporation and to obtain the CRBGCo strain.
3. Results and Discussion
3.1. Effects of argR/argG Deletion and argBEc Expression on L-Citrulline Biosynthesis of C. glutamicum
In the CRB strain, the heterologous argBEc gene encoding feedback-resistant NAGKEc from E. coli was inserted into the argR gene in the genome (Figure 1). Therefore, the argR gene encoding the repressor ArgR of the argCJBDFR and argGH operons was disrupted. The expression of the genes in the argCJBDF and argGH operons was strengthened in the CRB strain at the transcriptional level (Figure 2). This result indicated that the expression of the argCJBDF and argGH operons in the CRB strain was derepressed by the inactivation of the ArgR repressor. Meanwhile, the transcriptional analysis revealed that the argBEc gene was expressed along with the operons in the CRB strain (Figure 2).
As reported [13,17], C. glutamicum ATCC13032 could not accumulate L-citrulline and L-arginine (Figure 3a). Batch fermentation in the shake flasks revealed that the CRB strain could produce 3.6 g/L of L-citrulline and 1.1 g/L of L-arginine (Figure 3b). Previous research has shown that individually alleviating the feedback inhibition of NAGK by L-arginine or individually alleviating the transcriptional repression of the argCJBDF and argGH operons by the ArgR repressor could not make C. glutamicum ATCC13032 produce L-arginine [13]. Only simultaneously alleviating the feedback inhibition and transcriptional repression could give rise to excessive synthesis and the accumulation of L-arginine by C. glutamicum ATCC13032 [13]. The strain CRB could produce L-arginine; therefore, the insertion of argBEc into the argR gene led to the expression of heterologous, feedback-resistant NAGKEc and the inactivation of the ArgR repressor simultaneously in the CRB strain. In other words, the inactivation of the ArgR repressor and the expression of feedback-resistant NAGKEc were achieved in one-step genome modification.
In the CRBG strain, the argBEc gene was inserted into the genome between the argR and argG genes; meanwhile, the downstream region of the argR gene and the upstream region of the argG gene were deleted (Figure 1). Therefore, the ArgR repressor and argininosuccinate synthase were inactivated simultaneously. The expression of genes in the argCJBDF operon was strengthened in the CRBG strain at the transcriptional level (Figure 2). Meanwhile, the transcriptional analysis also revealed that the argBEc gene was expressed along with the argCJBDF operon, in the CRBG strain (Figure 2).
Batch fermentation in shake flasks showed that the strain CRBG could produce 5.7 g/L of L-citrulline with a yield of 0.13 g/g glucose, and there was no accumulation of L-arginine in fermentation broth (Figure 3c). The results indicated that the conversion of L-citrulline to L-arginine was blocked in the CRBG strain by deleting the argG gene. Therefore, the inactivation of the ArgR repressor, the expression of feedback-resistant NAGKEc, and the blocking of L-citrulline conversion were achieved simultaneously in the CRBG strain by one-step genome modification.
3.2. Further Enhancing of L-Citrulline Biosynthesis by Plasmid-Based Overexpression of the Optimized L-Citrulline Operon
In C. glutamicum, the argCJBDF operon encodes all of the enzymes for L-citrulline synthesis from L-glutamate. After the elimination of the transcriptional repression of argCJBDF operon by ArgR, the plasmid-based expression of L-citrulline operon can further increase the expression level. Thus, the plasmid-based overexpression of the L-citrulline operon strain CRBGC and CRBGCo was constructed. The batch fermentation of the CRBGC (Figure 4a) and CRBGCo (Figure 4b) strain in shake flasks resulted in 8.2 g/L and 9.1 g/L of L-citrulline, respectively, with a yield of 0.16 g/g glucose and 0.18 g/g glucose, respectively, which were obviously higher than those of the CRBG strain (Figure 3c). In addition, the CRBGCo strain obtained higher L-citrulline production than the CRBGC strain. The results indicated that the L-citrulline biosynthesis in the CRBGC and CRBGCo strains was further strengthened by the plasmid-based overexpression of the L-citrulline operon, and the plasmid-based overexpression of the optimized L-citrulline operon was more beneficial to the L-citrulline biosynthesis.
In L-ornithine-producing C. glutamicum, the plasmid-based overexpression of the argCJBD genes (Figure 1) could significantly increase the L-ornithine production [15]. Similarly, in L-arginine producing C. crenatum, the plasmid-based overexpression of the argCJBDFRGH cluster (Figure 1) could also strengthen the L-arginine synthesis [23]. In this study, the L-citrulline biosynthesis was also strengthened by the plasmid-based overexpression of the L-citrulline operon. Therefore, the intensification of the whole biosynthetic pathway is effective to increase the biosynthesis efficiency of the target product.
3.3. Fed-Batch Fermentation for L-Citrulline Production
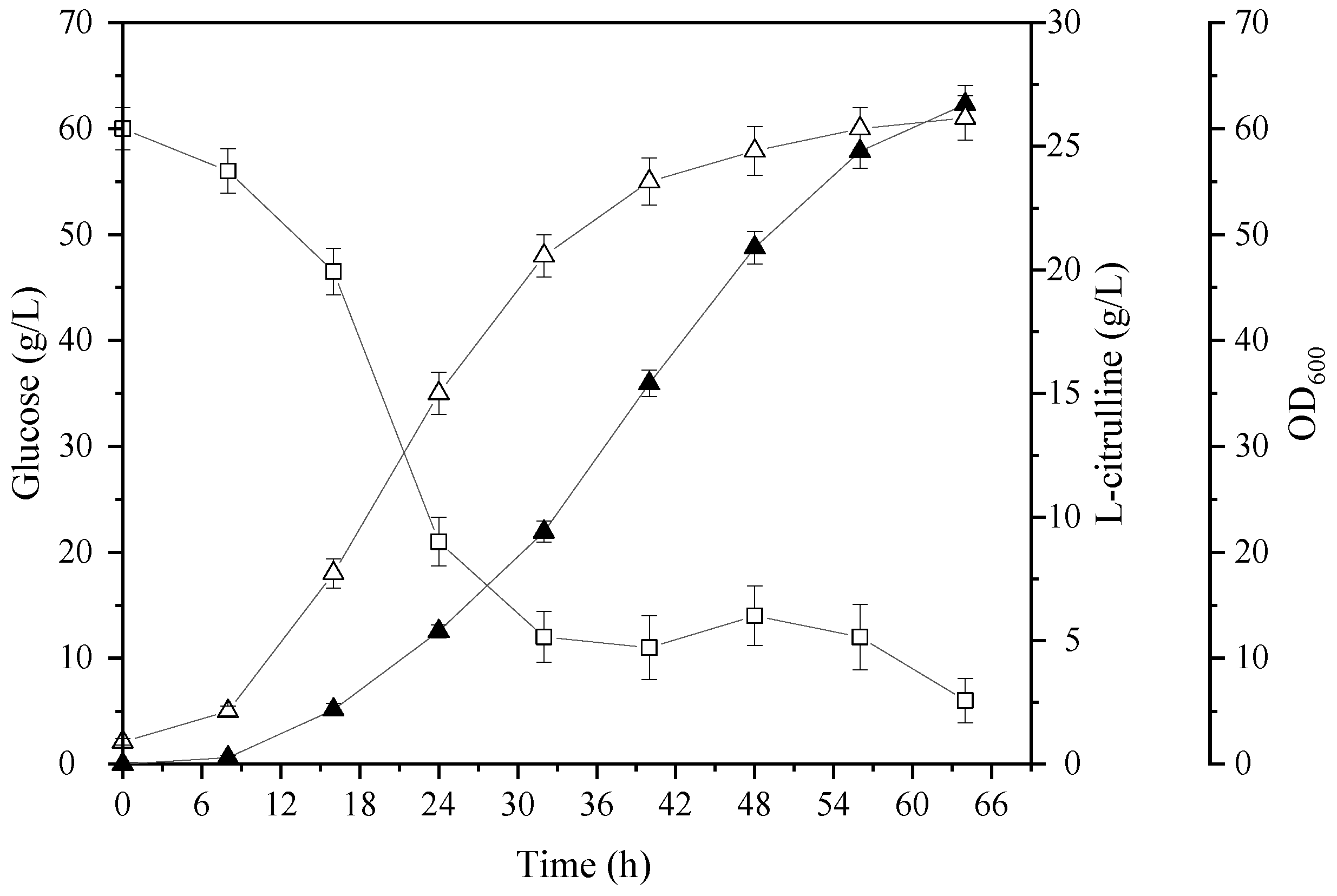
The production performance of the CRBGCo strain was investigated in a fed-batch process, and the fermentation process is shown in Figure 5. The fed-batch fermentation of the CRBGCo strain resulted in 26.7 g/L of L-citrulline with a yield of 0.18 g/g glucose. The results indicate that the optimization of the L-citrulline operon in C. glutamicum is effective to construct the L-citrulline over-producing strain. In fed-batch fermentation, the L-citrulline yield from the glucose of the CRBGCo strain was the same as the batch fermentation in shake flasks, but the productivity (0.42 g/L/h) was higher than that of the batch fermentation (0.25 g/L/h). In the next work, the production of L-citrulline by the CRBGCo strain can be improved through the optimization of the fermentation conditions.
4. Discussion
In this study, we performed preliminary metabolic engineering on C. glutamicum for L-citrulline production, and the work was focused on the optimization of the L-citrulline operon. The resulting CRBGCo strain could produce 26.7 g/L of L-citrulline in fed-batch fermentation. Therefore, as reported previously [16,17], the results in this study also prove that C. glutamicum can be used for L-citrulline fermentation production. Compared to the CIT0(pVWEx1-argFBfbr) strain derived from C. glutamicum MB001 [17], the L-citrulline productivity and yield from the glucose of the CRBGCo strain in batch fermentation were relatively low, but the L-citrulline production of the CRBGCo strain was relatively high. Compared to the CIT strain derived from C. glutamicum ATCC13032 [16], the CRBGCo strain had higher L-citrulline production, productivity, and yield from glucose in the batch fermentation. Therefore, the metabolic engineering strategies used in this study are effective for constructing L-citrulline-producing C. glutamicum. The feedback-insensitive NAGK is critical for L-citrulline synthesis in C. glutamicum [8,17]. Therefore, the L-citrulline synthesis pathway can be further fine-tuned by regulating the expression level of the argBEc gene through the integration of different promoters [5,15]. C. glutamicum MB001, derived from C. glutamicum ATCC13032, a prophage-free C. glutamicum strain with a genome reduced by 6%, is more efficient for plasmid-based gene overexpression [17,24]. Therefore, the C. glutamicum MB001 is perhaps more suitable for the tuning of natural or synthetic metabolic pathways required for small-molecule production [24]. In the next work, the metabolic engineering strategies used in this study can be performed in C. glutamicum MB001, and more efficient L-citrulline production is possible to realize.
Similar to the previous study [17], in batch fermentation, the biomass of the L-citrulline-producing strains was decreased when L-citrulline production was increased (Figure 3 and Figure 4). This is because more carbon flux and energy were used for L-citrulline production when the L-citrulline synthesis pathway was strengthened, and the cell growth was negatively affected. In batch fermentation, the initial concentration of glucose was set at a high level (80 g/L) to avoid the depletion of glucose, and the residual glucose concentration was around 30 g/L. Meanwhile, the productivity of L-citrulline in the fed-batch fermentation was obviously enhanced compared with that in the batch fermentation. Therefore, the L-citrulline production potential of the CRBGCo strain can be further unleashed through the optimization of the fermentation medium and conditions.
Similar to L-arginine, the biosynthesis of L-citrulline in C. glutamicum requires ATP, NADPH, and carbamoyl phosphate (Figure 1) [5,11,12,25]. Systems metabolic engineering, including the overexpression of biosynthetic genes, the removal of metabolism regulation [5], the increase in the supply of the L-glutamate precursor [12], cofactors [5,15], energy [11], the building block (carbamoyl phosphate) [5], and the improvement of the ammonia assimilation [25,26], can be carried out to optimize the metabolic network and obtain a more efficient L-citrulline-producing strain. C. glutamicum can use a variety of carbon sources for cell growth and product synthesis; more importantly, C. glutamicum can simultaneously utilize different carbon sources present in mixtures such as lignocellulosic hydrolysates [17]. In the next work, the investigation of using lignocellulosic hydrolysates as a carbon source for L-citrulline production can be performed.
5. Conclusions
In this study, the optimization of the L-citrulline operon in C. glutamicum, including the chromosomal integration of the heterologous argBEc gene, the chromosomal deletion of the argR and argG genes, and the plasmid-based overexpression of the optimized L-citrulline operon, was performed for L-citrulline production. In C. glutamicum ATCC13032, the chromosomal integration of the integral argBEc gene could introduce the natively insensitive NAGK, the deletion of the argR gene could relieve the repression of the L-citrulline operon by the ArgR repressor, and the deletion of the argG gene could block the conversion of L-citrulline to L-arginine. These three strain-engineering strategies were simultaneously achieved by one-step genome modification, and the L-citrulline-producing strain was obtained. Further enhancing the expression level of the optimized L-citrulline operon by plasmid-based overexpression could further improve the L-citrulline production. In the next work, systems metabolic engineering and fermentation-process optimization can be performed to increase the L-citrulline production, and using a non-food-derived carbon source such as lignocellulosic hydrolysates for L-citrulline production can also be investigated.
Author Contributions
Conceptualization, Z.M., J.L. and H.C.; data curation, Z.M. and J.G.; methodology, Z.M., J.L. and H.C.; software, Z.M.; supervision, Z.C. and J.G.; writing—original draft, Z.M., J.L. and H.C.; and writing—review and editing, Z.C. and J.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31700075); the Key Research and Development Program of Shandong Province, China (2019JZZY020605); the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (19KJB180001); and the Scientific Research Foundation of Changzhou University (ZMF19020298; ZMF17020115).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Breuillard, C.; Cynober, L.; Moinard, C. Citrulline and nitrogen homeostasis: An overview. Amino Acids 2015, 47, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Sun, X.; Chen, X.; Liu, D.; Liu, L. Enzymatic production of L-citrulline by hydrolysis of the guanidinium group of L-arginine with recombinant arginine deiminase. J. Biotechnol. 2015, 208, 37–43. [Google Scholar] [CrossRef]
- Su, L.; Ma, Y.; Wu, J. Extracellular expression of natural cytosolic arginine deiminase from Pseudomonas putida and its application in the production of L-citrulline. Bioresour. Technol. 2015, 196, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Lee, S.Y. Metabolic engineering of microorganisms for the production of L-arginine and its derivatives. Microb. Cell Fact. 2014, 13, 166. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, H.U.; Kim, T.Y.; Park, J.S.; Kim, S.S.; Lee, S.Y. Metabolic engineering of Corynebacterium glutamicum for L-arginine production. Nat. Commun. 2014, 5, 4618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Wendisch, V.F. Production of amino acids—Genetic and metabolic engineering approaches. Bioresour. Technol. 2017, 245, 1575–1587. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Wendisch, V.F. Engineering microbial cell factories: Metabolic engineering of Corynebacterium glutamicum with a focus on non-natural products. Biotechnol. J. 2015, 10, 1170–1184. [Google Scholar] [CrossRef]
- Jensen, J.V.K.; Eberhardt, D.; Wendisch, V.F. Modular pathway engineering of Corynebacterium glutamicum for production of the glutamate-derived compounds ornithine, proline, putrescine, citrulline, and arginine. J. Biotechnol. 2015, 214, 85–94. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Jorge, J.M.P.; Pérez-García, F.; Sgobba, E. Updates on industrial production of amino acids using Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2016, 32, 105. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. Bio-based production of chemicals, materials and fuels—Corynebacterium glutamicum as versatile cell factory. Curr. Opin. Biotechnol. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Man, Z.; Rao, Z.; Xu, M.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Improvement of the intracellular environment for enhancing L-arginine production of Corynebacterium glutamicum by inactivation of H2O2-forming flavin reductases and optimization of ATP supply. Metab. Eng. 2016, 38, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Man, Z.; Xu, M.; Rao, Z.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Systems pathway engineering of Corynebacterium crenatum for improved L-arginine production. Sci. Rep. 2016, 6, 28629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Mitsuhashi, S.; Tanaka, K.; Hayashi, M. Reengineering of a Corynebacterium glutamicum L-arginine and L-citrulline producer. Appl. Environ. Microbiol. 2009, 75, 1635–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Rao, Z.; Dou, W.; Yang, J.; Jin, J.; Xu, Z. Site-directed mutagenesis and feedback-resistant N-acetyl-L-glutamate kinase (NAGK) increase Corynebacterium crenatum L-arginine production. Amino Acids 2012, 43, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, J.; Lee, S.Y. Metabolic engineering of Corynebacterium glutamicum for the production of L-ornithine. Biotechnol. Bioeng. 2015, 112, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Hao, N.; Mu, J.; Hu, N.; Xu, S.; Yan, M.; Li, Y.; Guo, K.; Xu, L. Improvement of L-citrulline production in Corynebacterium glutamicum by ornithine acetyltransferase. J. Ind. Microbiol. Biotechnol. 2015, 42, 307–313. [Google Scholar] [CrossRef]
- Eberhardt, D.; Jensen, J.V.K.; Wendisch, V.F. L-citrulline production by metabolically engineered Corynebacterium glutamicum from glucose and alternative carbon sources. AMB Express 2014, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Xu, D.; Tan, Y.; Shi, F.; Wang, X. An improved shuttle vector constructed for metabolic engineering research in Corynebacterium glutamicum. Plasmid 2010, 64, 85–91. [Google Scholar] [CrossRef]
- Xu, H.; Dou, W.; Xu, H.; Zhang, X.; Rao, Z.; Shi, Z.; Xu, Z. A two-stage oxygen supply strategy for enhanced L-arginine production by Corynebacterium crenatum based on metabolic fluxes analysis. Biochem. Eng. J. 2009, 43, 41–51. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Bao, T.; Yang, T.; Xu, M.; Li, H.; Xu, Z.; Rao, Z. Moderate expression of the transcriptional regulator ALsR enhances acetoin production by Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rao, Z.; Dou, W.; Xu, Z. The role of ARGR repressor regulation on L-arginine production in Corynebacterium crenatum. Appl. Biochem. Biotechnol. 2013, 170, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rao, Z.; Yang, J.; Xia, H.; Dou, W.; Jin, J.; Xu, Z. Heterologous and homologous expression of the arginine biosynthetic argC~H cluster from Corynebacterium crenatum for improvement of L-arginine production. J. Ind. Microbiol. Biotechnol. 2012, 39, 495–502. [Google Scholar] [CrossRef]
- Baumgart, M.; Unthan, S.; Rückert, C.; Sivalingam, J.; Grünberger, A.; Kalinowski, J.; Bott, M.; Noack, S.; Frunzke, J. Construction of a prophage-free variant of Corynebacterium glutamicum ATCC 13032 for use as a platform strain for basic research and industrial biotechnology. Appl. Environ. Microbiol. 2013, 79, 6006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Man, Z.; Rao, Z.; Xu, M.; Yang, T.; Zhang, X.; Xu, Z. Improvement of the ammonia assimilation for enhancing L-arginine production of Corynebacterium crenatum. J. Ind. Microbiol. Biotechnol. 2017, 44, 443–451. [Google Scholar] [CrossRef]
- Xu, M.; Li, J.; Shu, Q.; Tang, M.; Zhang, X.; Yang, T.; Xu, Z.; Rao, Z. Enhancement of L-arginine production by increasing ammonium uptake in an AmtR-deficient Corynebacterium crenatum mutant. J. Ind. Microbiol. Biotechnol. 2019, 46, 1155–1166. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the L-citrulline biosynthesis pathway in C. glutamicum and the modifications of the genome.
Figure 1.
Schematic representation of the L-citrulline biosynthesis pathway in C. glutamicum and the modifications of the genome.
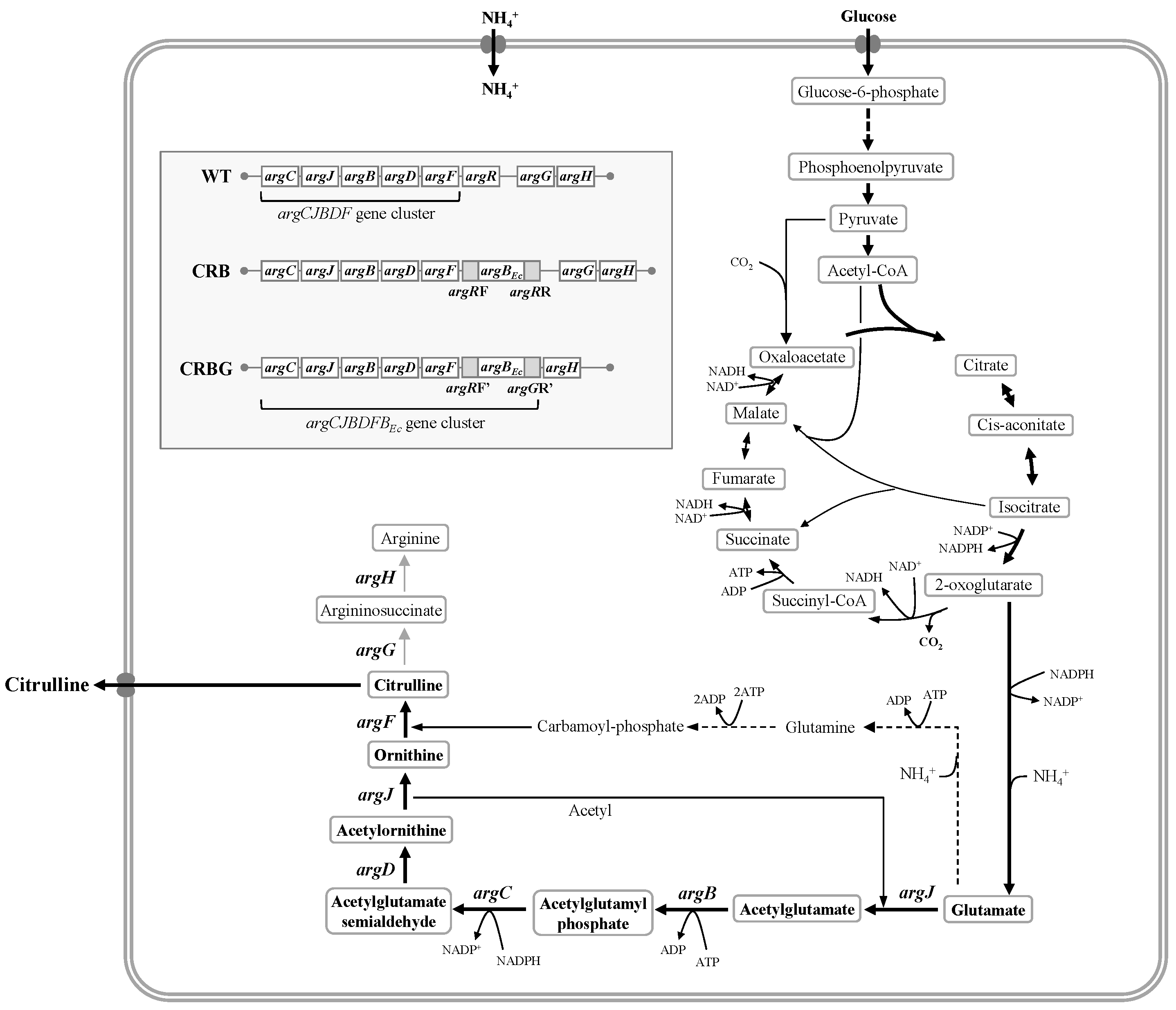
Figure 2.
Transcriptional analysis of genes involved in L-citrulline and L-arginine biosynthesis. The expression levels of the genes were calculated as the ratio of 16S rRNA expression level. The error bars represent the standard deviation of three independent replicates.
Figure 2.
Transcriptional analysis of genes involved in L-citrulline and L-arginine biosynthesis. The expression levels of the genes were calculated as the ratio of 16S rRNA expression level. The error bars represent the standard deviation of three independent replicates.
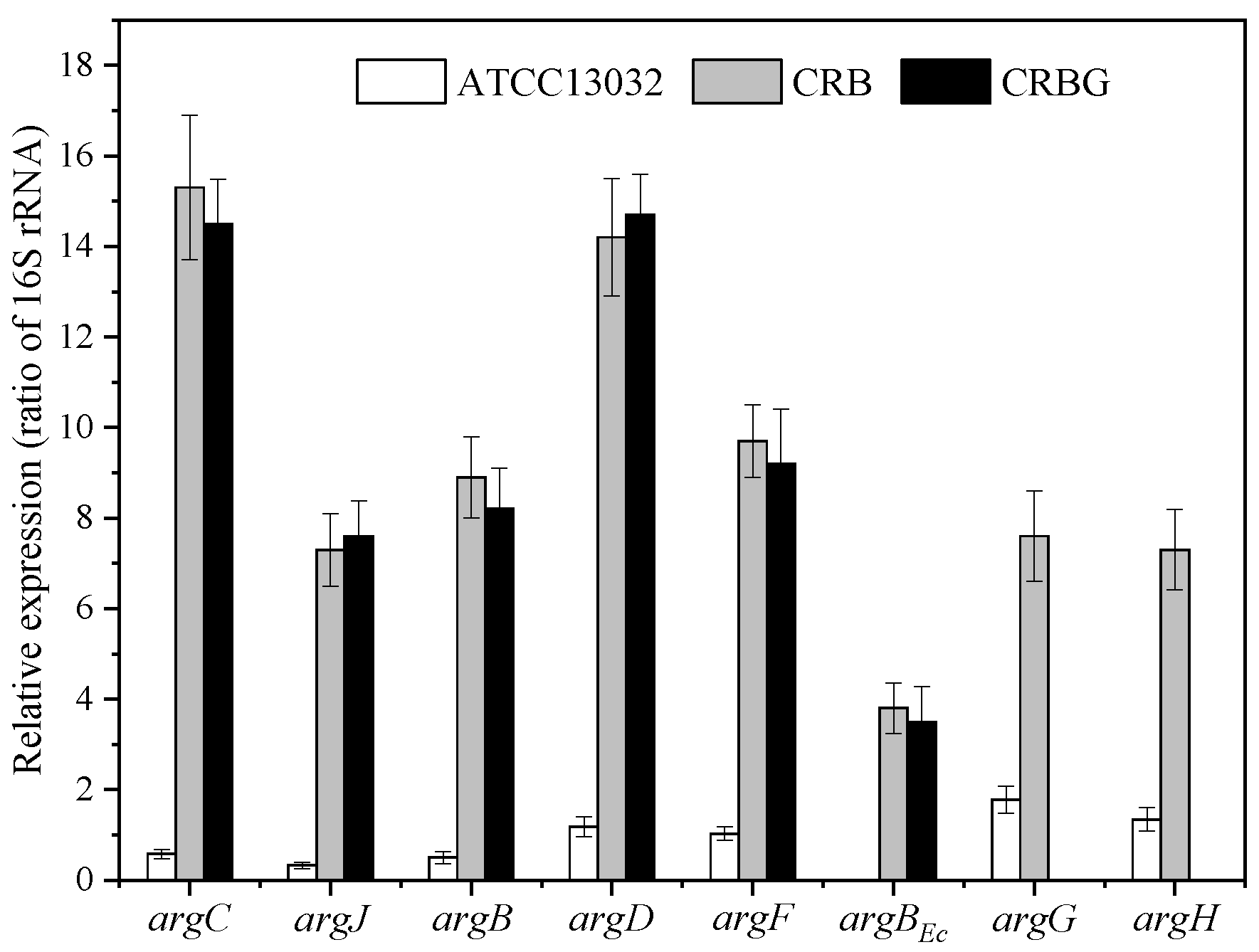
Figure 3.
Batch fermentation of C. glutamicum ATCC13032 (a), CRB (b), and CRBG (c) strains in shake flasks. Signal denotes: filled triangle, L-citrulline; filled circle, L-arginine; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.
Figure 3.
Batch fermentation of C. glutamicum ATCC13032 (a), CRB (b), and CRBG (c) strains in shake flasks. Signal denotes: filled triangle, L-citrulline; filled circle, L-arginine; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.
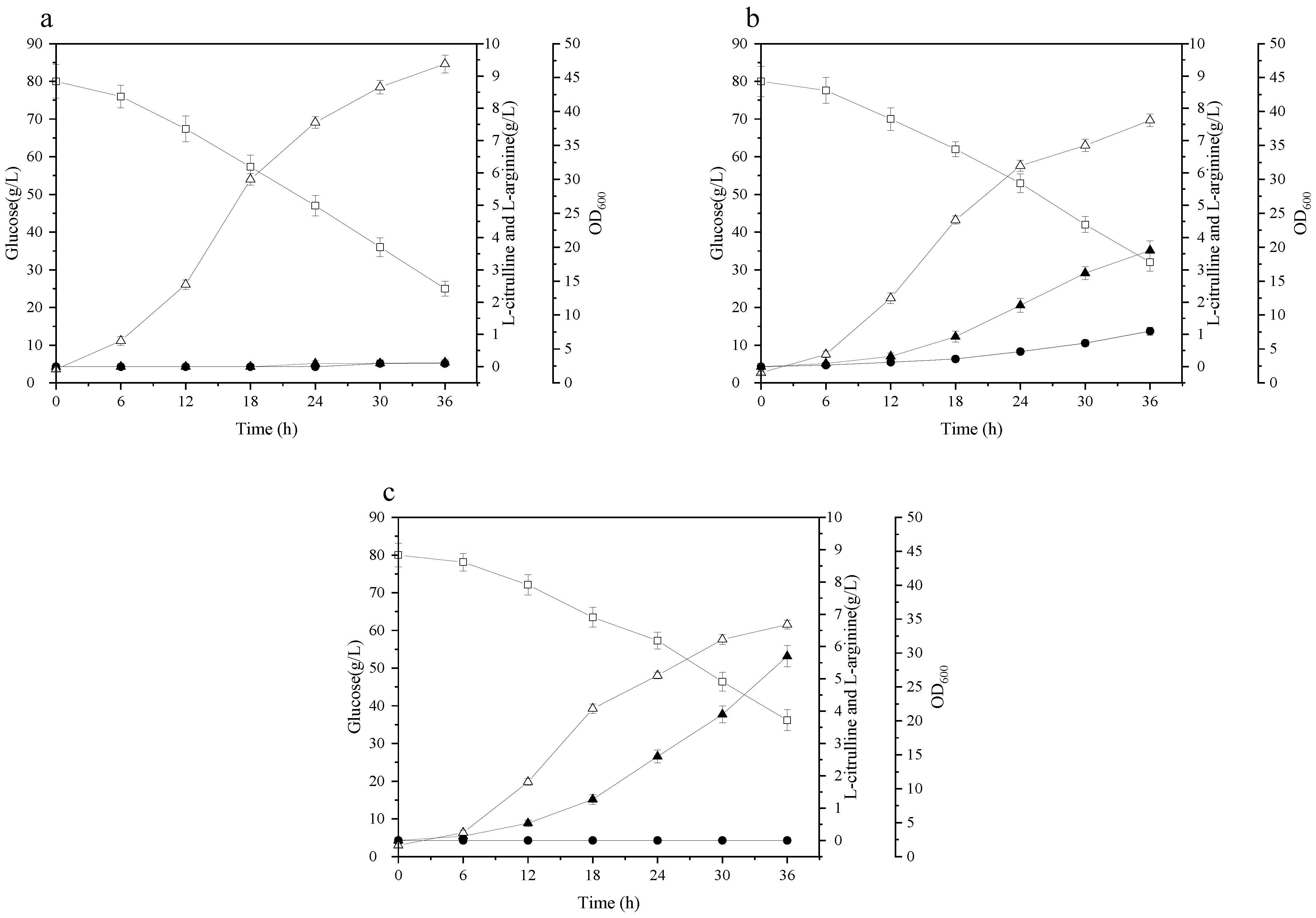
Figure 4.
Batch fermentation of CRBGC (a) and CRBGCo (b) strains in shake flasks. Signal denotes: filled triangle, L-citrulline; filled circle, L-arginine; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.
Figure 4.
Batch fermentation of CRBGC (a) and CRBGCo (b) strains in shake flasks. Signal denotes: filled triangle, L-citrulline; filled circle, L-arginine; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.

Figure 5.
Fed-batch fermentation of CRBGCo strain in 5-L stirred fermenter. Signal denotes: filled triangle, L-citrulline; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.
Figure 5.
Fed-batch fermentation of CRBGCo strain in 5-L stirred fermenter. Signal denotes: filled triangle, L-citrulline; empty square, glucose; and empty triangle, OD600. The error bars represent the standard deviation of three independent replicates.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains and plasmids used in this study.
| Strain or Plasmid | Description | Reference or Source |
|---|---|---|
| Strains | ||
| E. coli JM109 | General cloning host | TaKaRa |
| C. glutamicum ATCC13032 | Wild-type strain | ATCC |
| CRB | C. glutamicum ATCC13032 derivative with insertion of argBEc into argR gene | This work |
| CRBG | C. glutamicum ATCC13032 derivative with insertion of argBEc between argR and argG genes | This work |
| CRBGC | CRBG derivative harboring pDXW-10-argCJBDF | This work |
| CRBGCo | CRBG derivative harboring pDXW-10-argCJBDFBEc | |
| plasmids | ||
| pK18mobsacB | Kanr; vector for in-frame deletions | Lab stock [18] |
| pK18RB | Derived from pK18mobsacB, harboring argRF-argBEc-argRR fragment | This work |
| pK18RBG | Derived from pK18mobsacB, harboring argRF’-argBEc-argGR’ fragment | This work |
| pDXW-10 | Kanr; shuttle vector between E. coli and C. glutamicum | Lab stock [19] |
| pDXW-10-argCJBDF | Derived from pDXW-10, for constitutive expression of argCJBDF gene cluster | This work |
| pDXW-10-argCJBDFBEc | Derived from pDXW-10, for constitutive expression of the optimized L-citrulline operon argCJBDFBEc | This work |
Table 2.
Primers used in this study.
| Names | Sequences (5′→3′) | Restriction Sites |
|---|---|---|
| RB-RF1 | CCGGAATTCTAACGCTAACAAGCTCTCATTTTGCAGATT | EcoRI |
| RB-RR1 | TTACATTTAGCGGCGCAGCTTCTCCGACGGAC | |
| RB-RF2 | TAGCTTAAGTTTTGTTGGCCGGAGGCGCAGTGATGAACTGCTGGTTTCTACAGATC | |
| RB-RR2 | CGCGGATCCGCGCCCGCTGAGTAATTCACCTA | BamHI |
| RB-BF | AGCTGCGCCGCTAAATGTAACGAAGACAACTTACTGAAAG | |
| RB-BR | GGCCAACAAAACTTAAGCTA | |
| RBG-RF | CCGGAATTCGCACGCCAAGCTCTCATTTTGCAGATTTTG | EcoRI |
| RBG-RR | GGTGCGCAGCATCGCGATGTTGCCGG | |
| RBG-GF | GGCGCAGCTTTCCGGCATTGAATTTCGCTACAAGCGCGGCGTTGACGCACG | |
| RBG-GR | CGCGGATCCCTAAATACTTGAAGTTCTAGTCTGG | BamHI |
| RBG-BF | CCGGCAACATCGCGATGCTGCGCACCCCAATATTCGTTTCGGCTATGCGGAAAC | |
| RBG-BR | GAAATTCAATGCCGGAAAGCTGCGCC | |
| CF | AAGGAAAAAAGCGGCCGCAAATTCATGCTTTTACCCACTTGCAG | NotI |
| CR | GGAAGATCTGCAAAATGAGAGCTTGGCGTGCAGTGCG | BglII |
| CoR | AAGGAAAAAAGCGGCCGCGAAATTCAATGCCGGAAAGCTGCGCC | NotI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Man, Z.; Li, J.; Cui, H.; Cai, Z.; Guo, J. Optimization of L-Citrulline Operon in Corynebacterium glutamicum for L-Citrulline Production. Processes 2022, 10, 2153. https://doi.org/10.3390/pr10102153
AMA Style
Man Z, Li J, Cui H, Cai Z, Guo J. Optimization of L-Citrulline Operon in Corynebacterium glutamicum for L-Citrulline Production. Processes. 2022; 10(10):2153. https://doi.org/10.3390/pr10102153
Chicago/Turabian StyleMan, Zaiwei, Jin Li, Huihui Cui, Zhiqiang Cai, and Jing Guo. 2022. "Optimization of L-Citrulline Operon in Corynebacterium glutamicum for L-Citrulline Production" Processes 10, no. 10: 2153. https://doi.org/10.3390/pr10102153
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.