
Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation

 ,
, 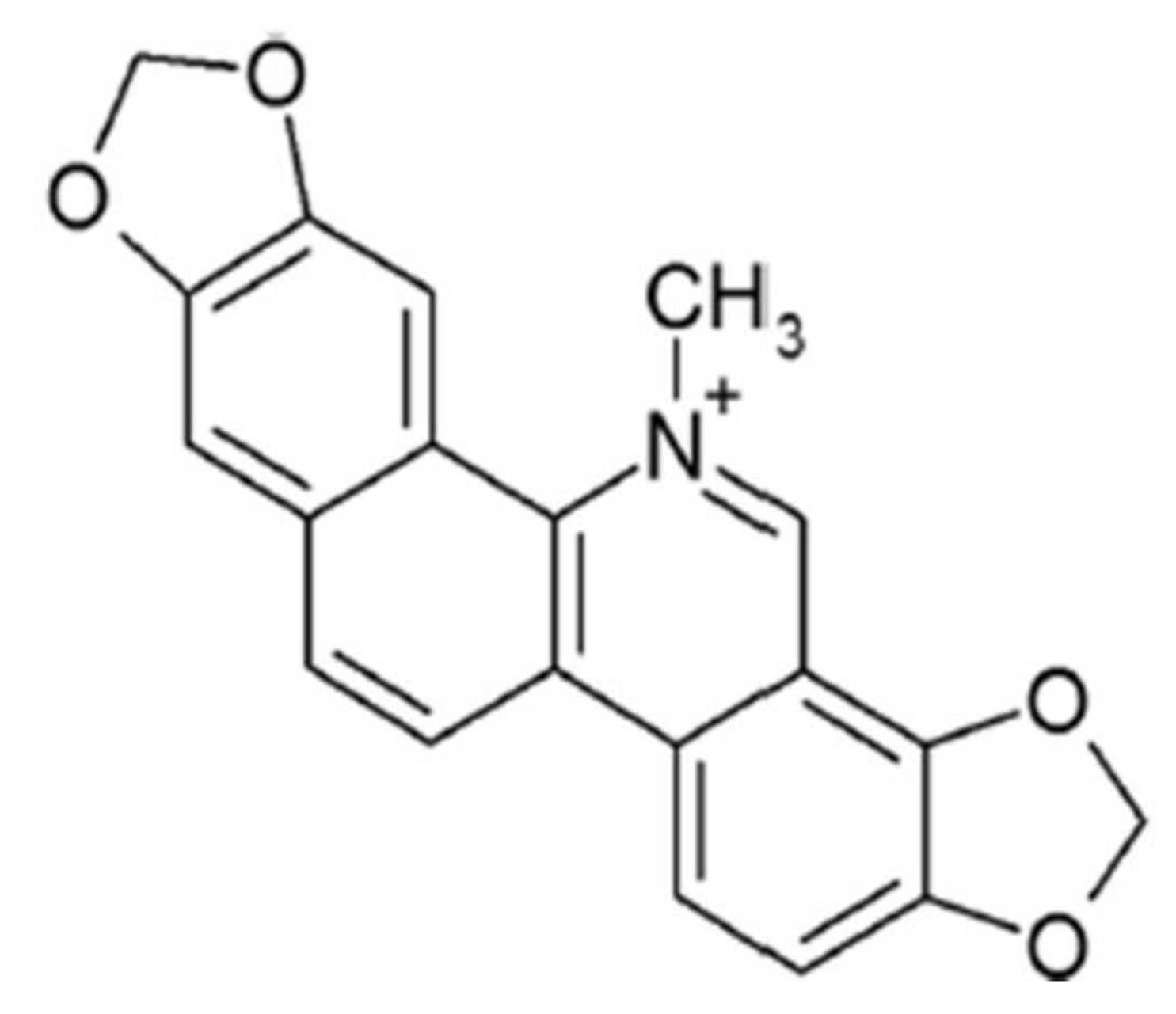{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. FeCl3-Induced Carotid Arterial Injury Thrombosis Model
2.3. Bleeding-Time Tests
2.4. Human Platelet Preparation
2.5. Platelet Aggregation and ATP Release Assay
2.6. Flow Cytometric Analysis
2.7. Platelet Adhesion on the Collagen-Coated Surface under Flow
2.8. Calcium Signaling
2.9. Platelet Spreading on Immobilized Fibrinogen
2.10. Immunoblot Analysis
2.11. Statistical Analysis
3. Results
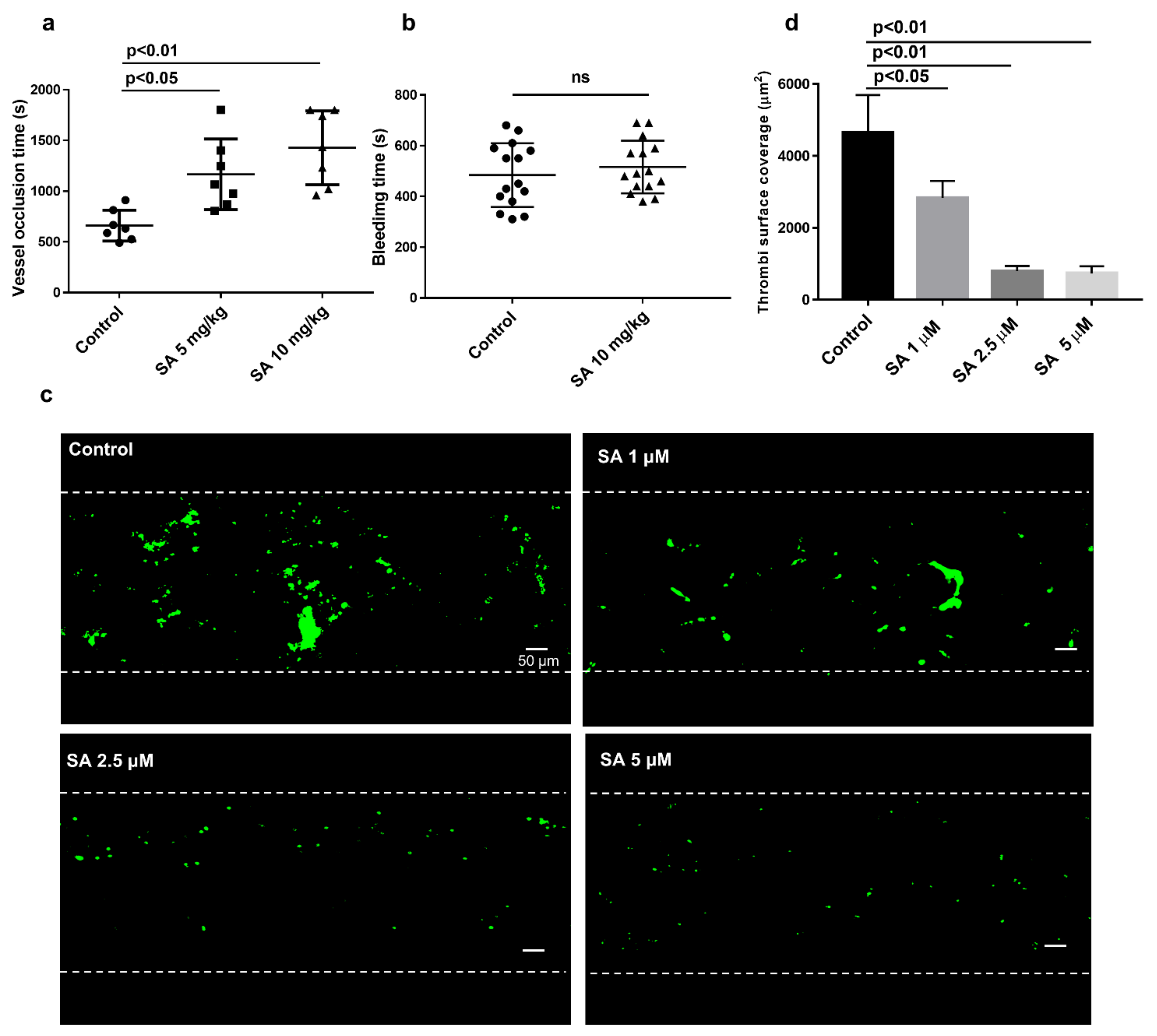
3.1. Sanguinarine Inhibits Arterial Thrombosis In Vivo and In Vitro
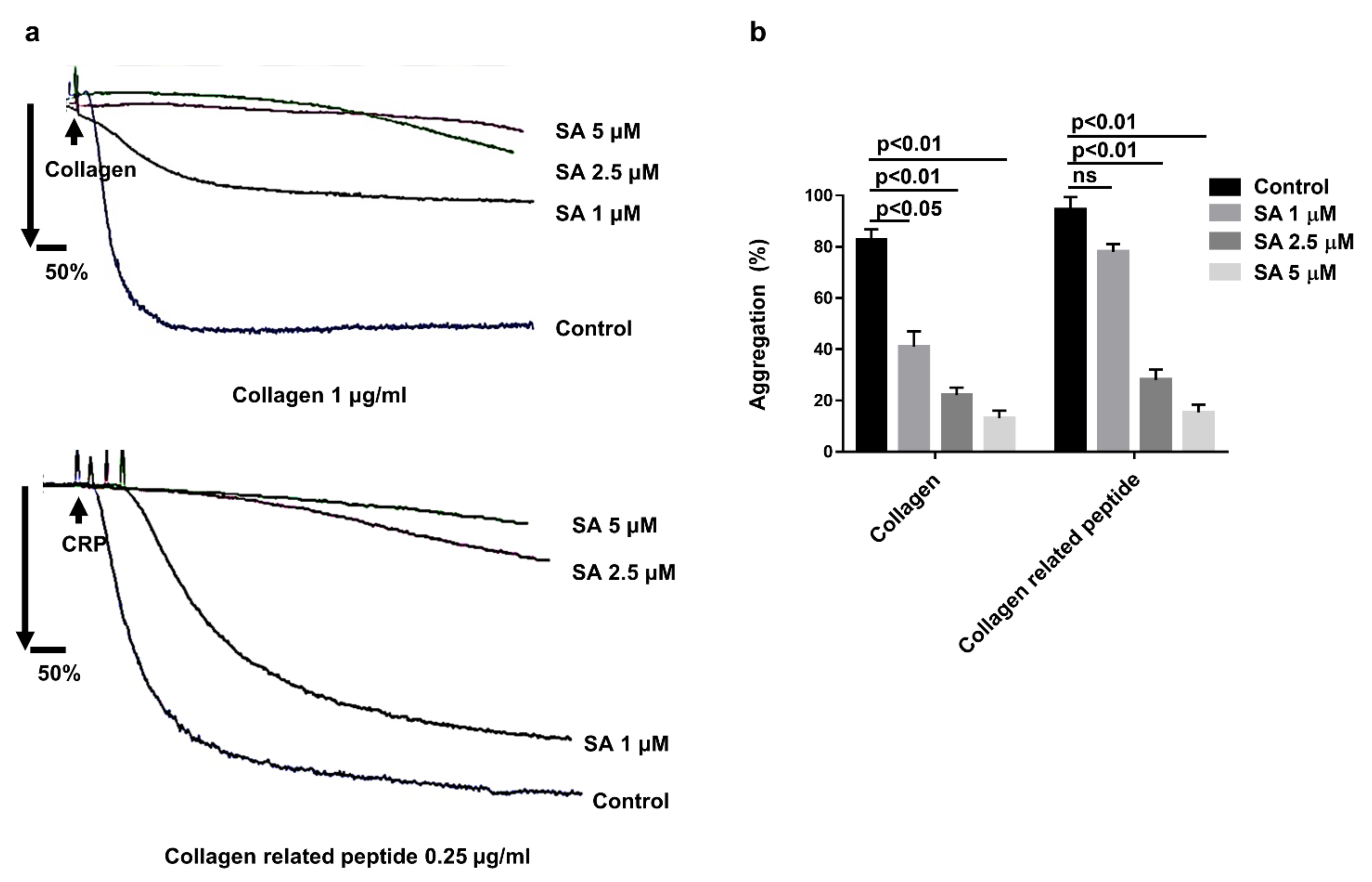
3.2. Sanguinarine Inhibits Human Platelet Aggregation Induced by Collagen and Collagen-Related Peptide
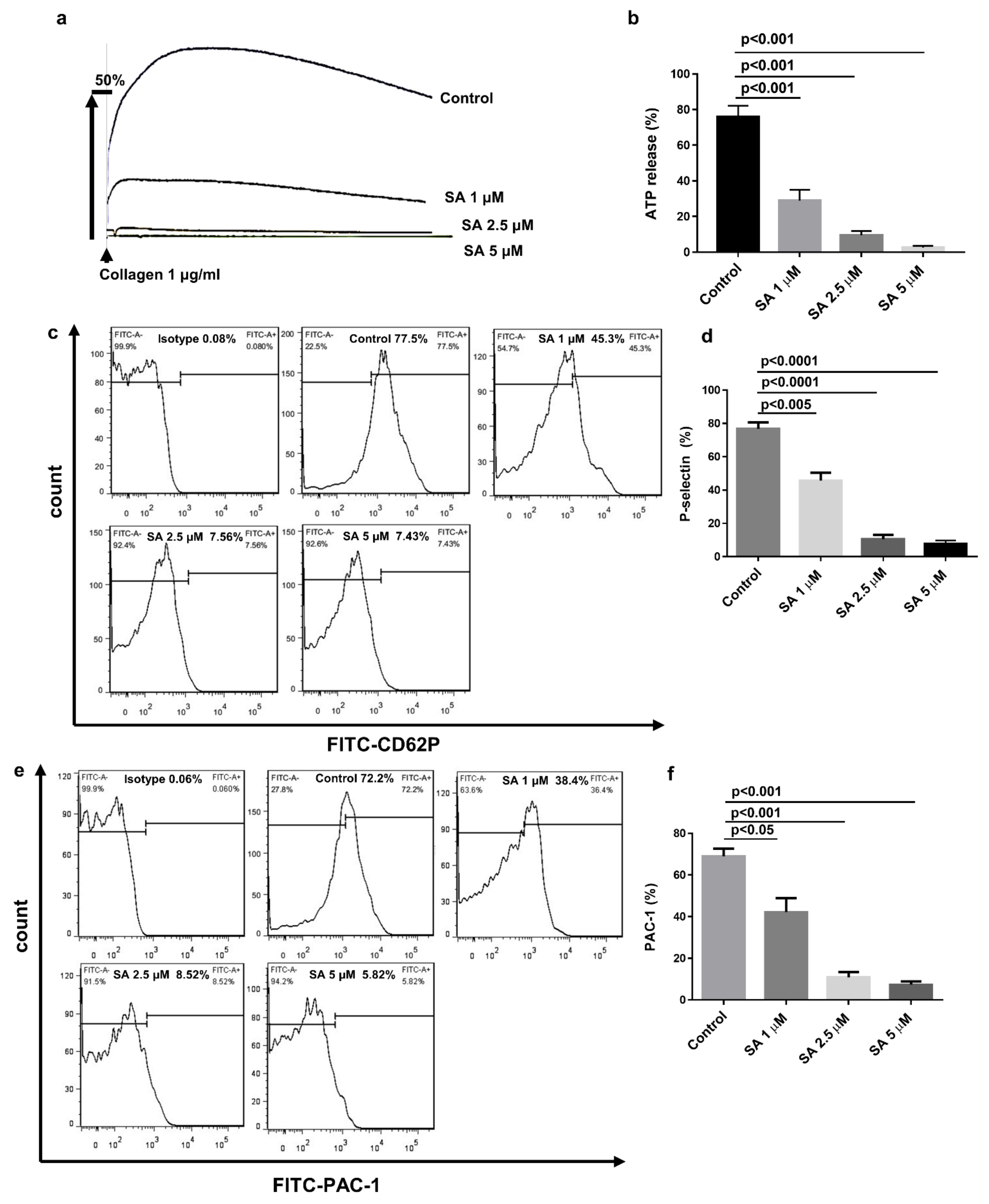
3.3. Sanguinarine Inhibits Collagen-Induced Human Platelet Granule Secretion and Integrin αIibβ3 Activation
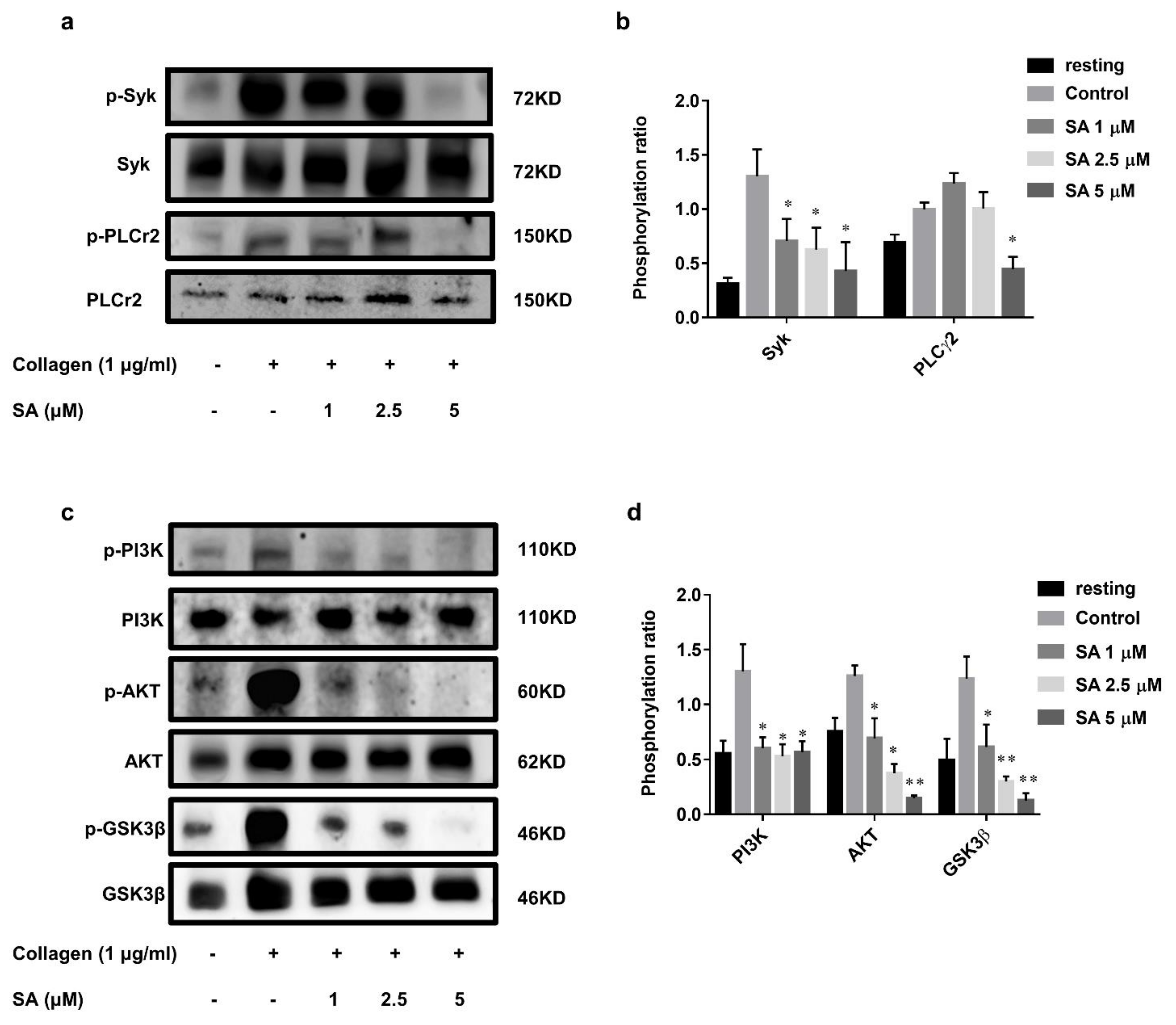
3.4. Sanguinarine Inhibits Collagen-Induced Human Platelet Intracellular Signal Transduction
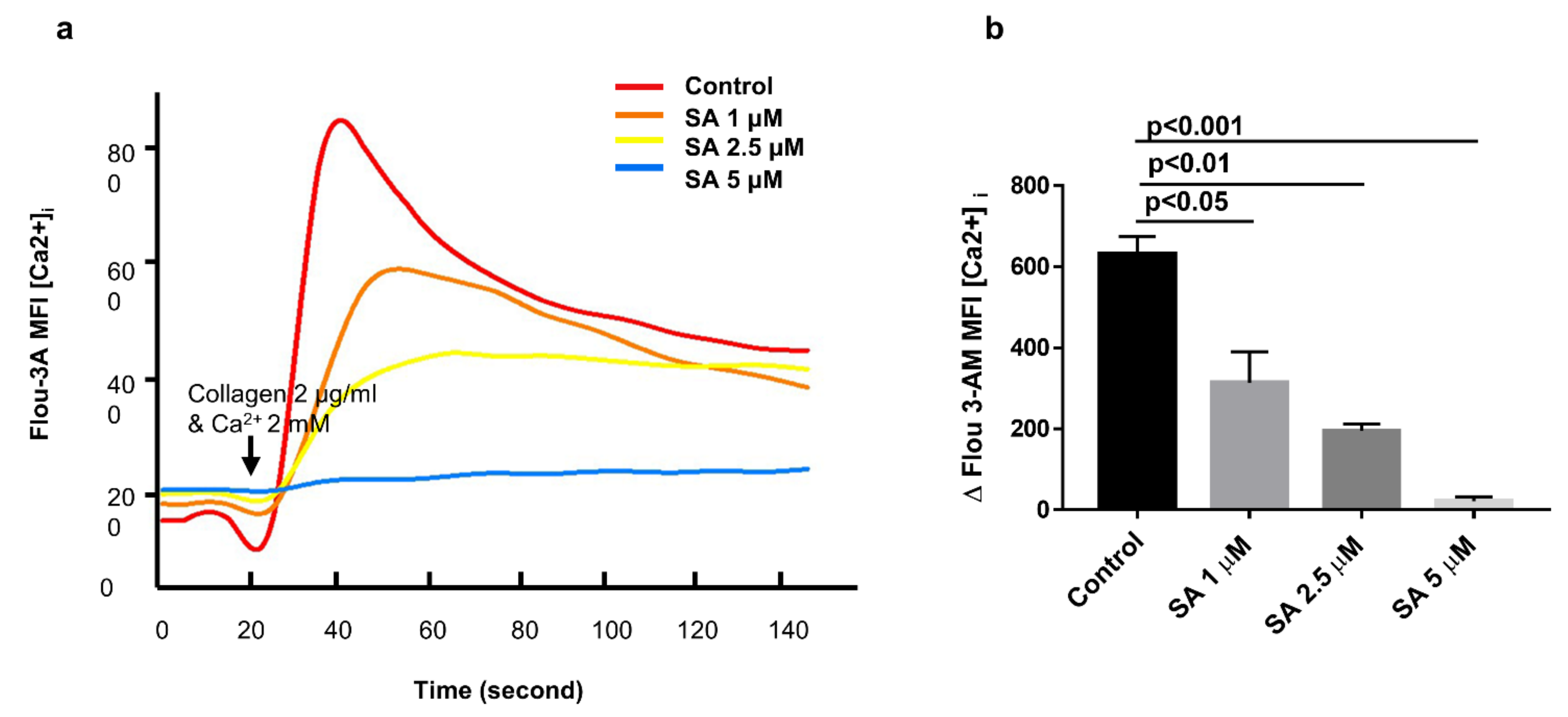
3.5. Sanguinarine Reduces Platelet Intracellular Ca2+ Concentrations ([Ca2+]i) in a Dose-Dependent Manner
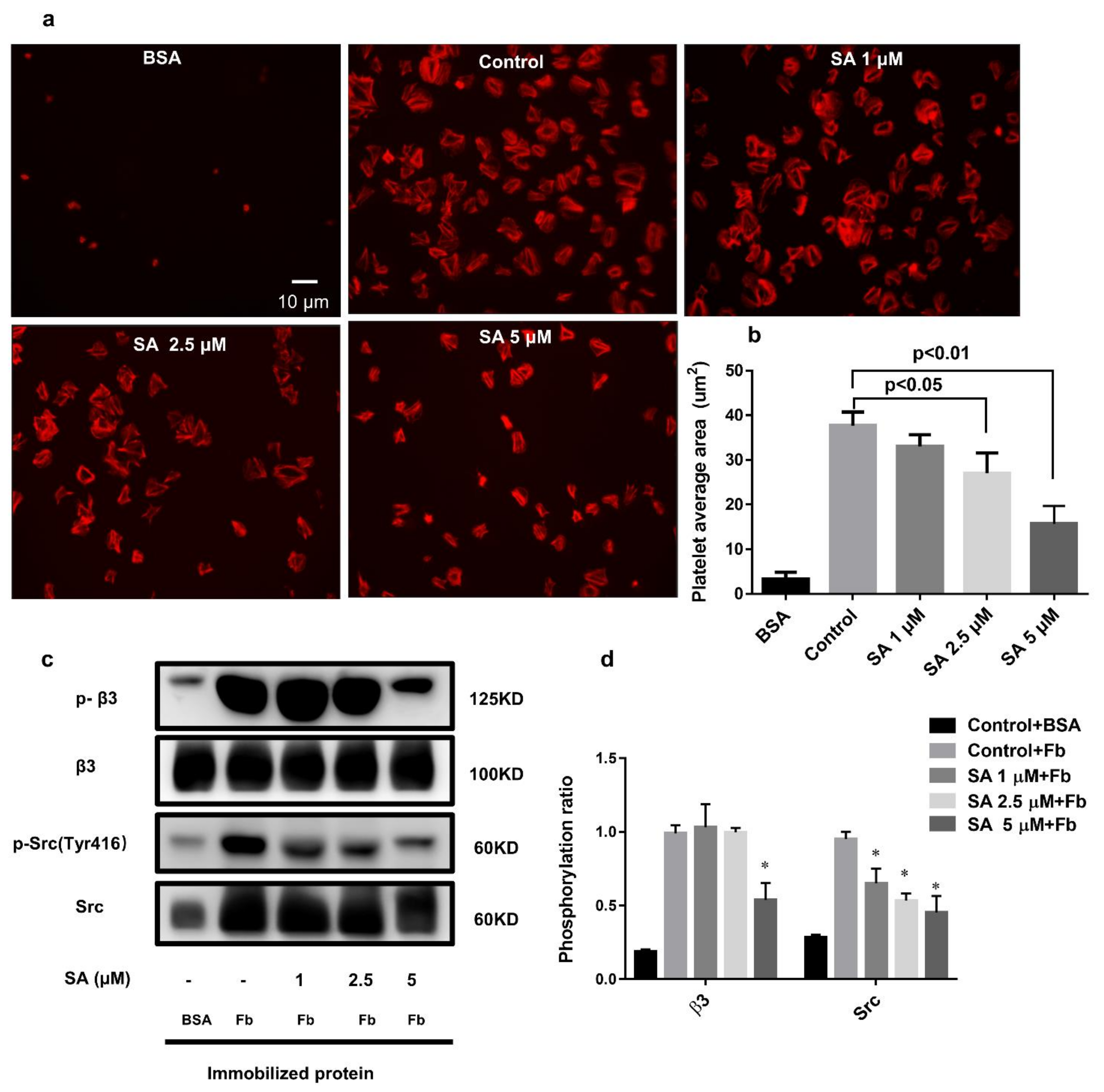
3.6. Sanguinarine Negatively Regulates the Integrin αIIbβ3 “Outside-In” Signaling
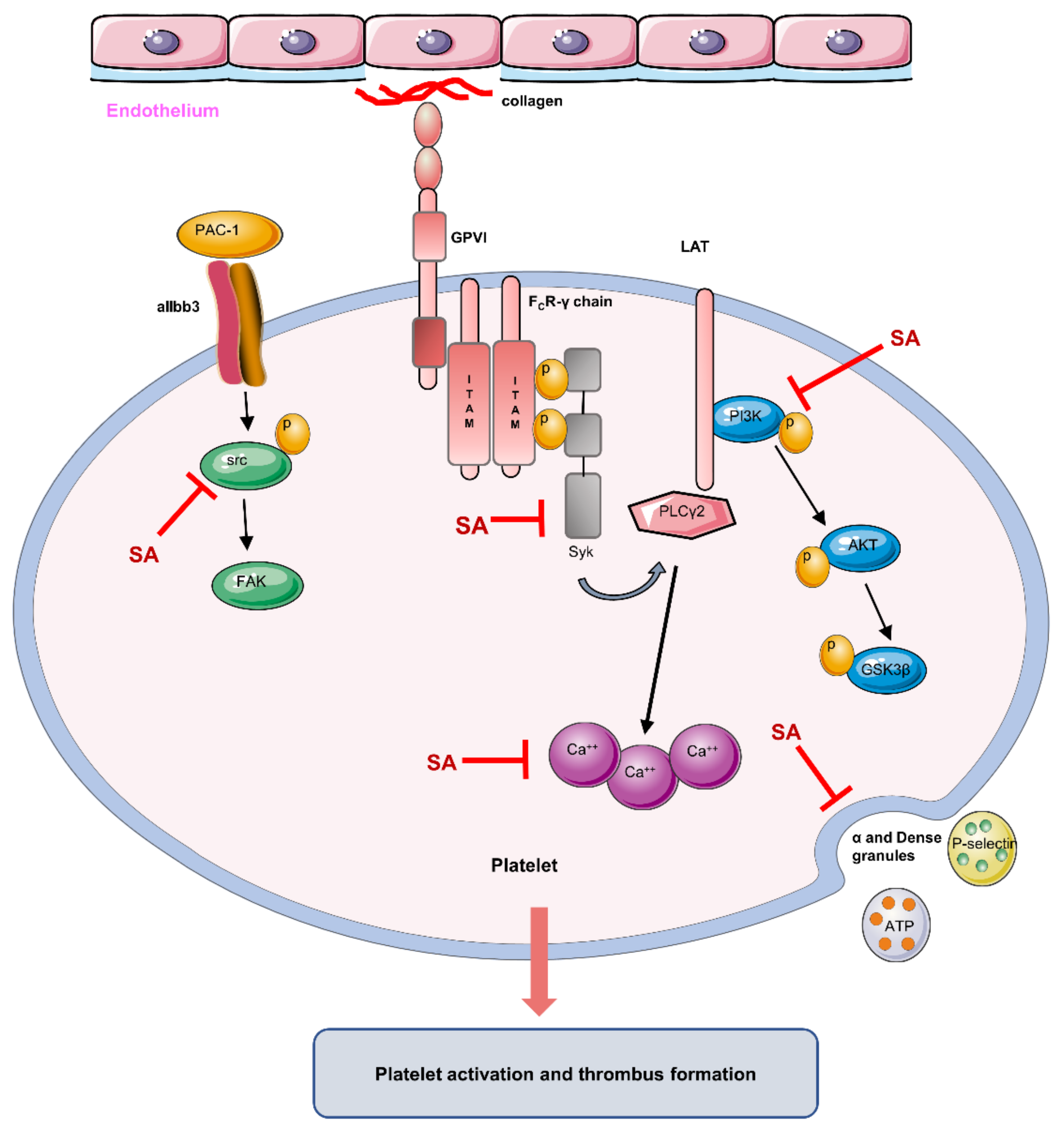
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: New concepts and clinical perspectives. Nat. Rev. Cardiol. 2019, 16, 166–179. [Google Scholar] [CrossRef]
- Cosemans, J.M.; Angelillo-Scherrer, A.; Mattheij, N.J.; Heemskerk, J.W. The effects of arterial flow on platelet activation, thrombus growth, and stabilization. Cardiovasc. Res. 2013, 99, 342–352. [Google Scholar] [CrossRef]
- Barry, F.A.; Gibbins, J.M. Protein kinase B is regulated in platelets by the collagen receptor glycoprotein VI. J. Biol. Chem. 2002, 277, 12874–12878. [Google Scholar] [CrossRef]
- Liang, M.L.; Da, X.W.; He, A.D.; Yao, G.Q.; Xie, W.; Liu, G.; Xiang, J.Z.; Ming, Z.Y. Pentamethylquercetin (PMQ) reduces thrombus formation by inhibiting platelet function. Sci. Rep. 2015, 5, 11142. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Xie, W.; He, A.D.; Da, X.W.; Liang, M.L.; Yao, G.Q.; Xiang, J.Z.; Gao, C.J.; Ming, Z.Y. Antiplatelet activity of chrysin via inhibiting platelet alphaIIbbeta3-mediated signaling pathway. Mol. Nutr. Food Res. 2016, 60, 1984–1993. [Google Scholar] [CrossRef] [PubMed]
- Authi, K.S. Orai1: A channel to safer antithrombotic therapy. Blood 2009, 113, 1872–1873. [Google Scholar] [CrossRef] [PubMed]
- Konopatskaya, O.; Matthews, S.A.; Harper, M.T.; Gilio, K.; Cosemans, J.M.; Williams, C.M.; Navarro, M.N.; Carter, D.A.; Heemskerk, J.W.; Leitges, M.; et al. Protein kinase C mediates platelet secretion and thrombus formation through protein kinase D2. Blood 2011, 118, 416–424. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Michelson, A.D. Antiplatelet therapies for the treatment of cardiovascular disease. Nat. Rev. Drug Discov. 2010, 9, 154–169. [Google Scholar] [CrossRef]
- Stolla, M.; Stefanini, L.; Roden, R.C.; Chavez, M.; Hirsch, J.; Greene, T.; Ouellette, T.D.; Maloney, S.F.; Diamond, S.L.; Poncz, M.; et al. The kinetics of alphaIIbbeta3 activation determines the size and stability of thrombi in mice: Implications for antiplatelet therapy. Blood 2011, 117, 1005–1013. [Google Scholar] [CrossRef]
- Shaito, A.; Thuan, D.T.B.; Phu, H.T.; Nguyen, T.H.D.; Hasan, H.; Halabi, S.; Abdelhady, S.; Nasrallah, G.K.; Eid, A.H.; Pintus, G. Herbal Medicine for Cardiovascular Diseases: Efficacy, Mechanisms, and Safety. Front. Pharmacol. 2020, 11, 422. [Google Scholar] [CrossRef]
- Mackraj, I.; Govender, T.; Gathiram, P. Sanguinarine. Cardiovasc. Ther. 2008, 26, 75–83. [Google Scholar] [CrossRef]
- Grenby, T.H. The use of sanguinarine in mouthwashes and toothpaste compared with some other antimicrobial agents. Br. Dent. J. 1995, 178, 254–258. [Google Scholar] [CrossRef]
- Basu, P.; Kumar, G.S. Sanguinarine and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 155–172. [Google Scholar] [CrossRef]
- Jeng, J.H.; Wu, H.L.; Lin, B.R.; Lan, W.H.; Chang, H.H.; Ho, Y.S.; Lee, P.H.; Wang, Y.J.; Wang, J.S.; Chen, Y.J.; et al. Antiplatelet effect of sanguinarine is correlated to calcium mobilization, thromboxane and cAMP production. Atherosclerosis 2007, 191, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Day, S.M.; Reeve, J.L.; Myers, D.D.; Fay, W.P. Murine thrombosis models. Thromb. Haemost. 2004, 92, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Nieman, M.; Sen Gupta, A. Ferric Chloride-induced Murine Thrombosis Models. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, T.; Ahmad, S.; Gupta, N.; Sahu, A.; Ahmad, Y.; Nair, V.; Chatterjee, T.; Bajaj, N.; Sengupta, S.; Ganju, L.; et al. Altered expression of platelet proteins and calpain activity mediate hypoxia-induced prothrombotic phenotype. Blood 2014, 123, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Xiang, J.Z.; Yuan, H.; Liu, H.; Tang, Q.; Hao, H.Z.; Yin, Z.; Wang, J.; Ming, Z.Y. Neferine exerts its antithrombotic effect by inhibiting platelet aggregation and promoting dissociation of platelet aggregates. Thromb. Res. 2013, 132, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Alluqmani, N.; Buhari, F.H.M.; Wasim, L.; Smith, L.K.; Quaile, A.T.; Shannon, M.; Hakim, Z.; Furmli, H.; Owen, D.M.; et al. Galectin-9 binds IgM-BCR to regulate B cell signaling. Nat. Commun. 2018, 9, 3288. [Google Scholar] [CrossRef]
- Xiang, K.; Liu, G.; Zhou, Y.J.; Hao, H.Z.; Yin, Z.; He, A.D.; Da, X.W.; Xiang, J.Z.; Wang, J.L.; Ming, Z.Y. 2,3,5,4′-tetrahydroxystilbene-2-O-β-D-glucoside (THSG) attenuates human platelet aggregation, secretion and spreading in vitro. Thromb. Res. 2014, 133, 211–217. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef]
- Ilkan, Z.; Wright, J.R.; Goodall, A.H.; Gibbins, J.M.; Jones, C.I.; Mahaut-Smith, M.P. Evidence for shear-mediated Ca2+ entry through mechanosensitive cation channels in human platelets and a megakaryocytic cell line. J. Biol. Chem. 2017, 292, 9204–9217. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, G.F.; Canobbio, I.; Torti, M. PI3K/Akt in platelet integrin signaling and implications in thrombosis. Adv. Biol. Regul. 2015, 59, 36–52. [Google Scholar] [CrossRef]
- Li, D.; August, S.; Woulfe, D.S. GSK3beta is a negative regulator of platelet function and thrombosis. Blood 2008, 111, 3522–3530. [Google Scholar] [CrossRef] [PubMed]
- Rink, T.J.; Sage, S.O. Calcium signaling in human platelets. Annu. Rev. Physiol. 1990, 52, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, M.H. Integrin activation. BMB Rep. 2014, 47, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Kehrel, B.E.; Corkrey, H.A.; Freedman, J.E. Thrombosis and platelets: An update. Eur. Heart J. 2017, 38, 785–791. [Google Scholar] [CrossRef]
- Eckly, A.; Hechler, B.; Freund, M.; Zerr, M.; Cazenave, J.P.; Lanza, F.; Mangin, P.H.; Gachet, C. Mechanisms underlying FeCl3-induced arterial thrombosis. J. Thromb. Haemost. 2011, 9, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Eckly, A.; Rinckel, J.Y.; Proamer, F.; Ulas, N.; Joshi, S.; Whiteheart, S.W.; Gachet, C. Respective contributions of single and compound granule fusion to secretion by activated platelets. Blood 2016, 128, 2538–2549. [Google Scholar] [CrossRef]
- Heijnen, H.; van der Sluijs, P. Platelet secretory behaviour: As diverse as the granules … or not? J. Thromb. Haemost. 2015, 13, 2141–2151. [Google Scholar] [CrossRef]
- Chen, Y.; Ju, L.A.; Zhou, F.; Liao, J.; Xue, L.; Su, Q.P.; Jin, D.; Yuan, Y.; Lu, H.; Jackson, S.P.; et al. An integrin αIIbβ3 intermediate affinity state mediates biomechanical platelet aggregation. Nat. Mater. 2019, 18, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.P.; Auger, J.M.; McCarty, O.J.; Pearce, A.C. GPVI and integrin alphaIIb beta3 signaling in platelets. J. Thromb. Haemost. 2005, 3, 1752–1762. [Google Scholar] [CrossRef]
- Watson, S.P.; Herbert, J.M.J.; Pollitt, A.Y. GPVI and CLEC-2 in hemostasis and vascular integrity. J. Thromb. Haemost. 2010, 8, 1457–1467. [Google Scholar] [CrossRef]
- Gratacap, M.P.; Payrastre, B.; Viala, C.; Mauco, G.; Plantavid, M.; Chap, H. Phosphatidylinositol 3,4,5-trisphosphate-dependent stimulation of phospholipase C-gamma2 is an early key event in FcgammaRIIA-mediated activation of human platelets. J. Biol. Chem. 1998, 273, 24314–24321. [Google Scholar] [CrossRef]
- Pasquet, J.M.; Bobe, R.; Gross, B.; Gratacap, M.P.; Tomlinson, M.G.; Payrastre, B.; Watson, S.P. A collagen-related peptide regulates phospholipase Cgamma2 via phosphatidylinositol 3-kinase in human platelets. Biochem. J. 1999, 342 Pt 1, 171–177. [Google Scholar] [CrossRef]
- Volz, J.; Kusch, C.; Beck, S.; Popp, M.; Vogtle, T.; Meub, M.; Scheller, I.; Heil, H.S.; Preu, J.; Schuhmann, M.K.; et al. BIN2 orchestrates platelet calcium signaling in thrombosis and thrombo-inflammation. J. Clin. Investig. 2020, 130, 6064–6079. [Google Scholar] [CrossRef] [PubMed]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef]
- Boylan, B.; Gao, C.; Rathore, V.; Gill, J.C.; Newman, D.K.; Newman, P.J. Identification of FcgammaRIIa as the ITAM-bearing receptor mediating alphaIIbbeta3 outside-in integrin signaling in human platelets. Blood 2008, 112, 2780–2786. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.R.; Nannizzi-Alaimo, L.; Prasad, K.S. Beta3 tyrosine phosphorylation in alphaIIbbeta3 (platelet membrane GP IIb-IIIa) outside-in integrin signaling. Thromb. Haemost. 2001, 86, 246–258. [Google Scholar] [PubMed]







 ; Induce
; Induce  )
; Induce )
)
; Induce )
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, D.; Zhu, Y.; Lu, M.; He, A.-D.; Chen, J.-B.; Ye, D.-S.; Liu, Y.; Zeng, X.-B.; Ma, R.; Ming, Z.-Y. Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation. Biomedicines 2021, 9, 444. https://doi.org/10.3390/biomedicines9050444
Shu D, Zhu Y, Lu M, He A-D, Chen J-B, Ye D-S, Liu Y, Zeng X-B, Ma R, Ming Z-Y. Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation. Biomedicines. 2021; 9(5):444. https://doi.org/10.3390/biomedicines9050444
Chicago/Turabian StyleShu, Dan, Ying Zhu, Meng Lu, Ao-Di He, Jiang-Bin Chen, Ding-Song Ye, Yue Liu, Xiang-Bin Zeng, Rong Ma, and Zhang-Yin Ming. 2021. "Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation" Biomedicines 9, no. 5: 444. https://doi.org/10.3390/biomedicines9050444
APA StyleShu, D., Zhu, Y., Lu, M., He, A.-D., Chen, J.-B., Ye, D.-S., Liu, Y., Zeng, X.-B., Ma, R., & Ming, Z.-Y. (2021). Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation. Biomedicines, 9(5), 444. https://doi.org/10.3390/biomedicines9050444