Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study

 and
and
Abstract
1. Introduction
2. Experimental Section
2.1. Study Group
2.2. RNA Extraction and RT-qPCR
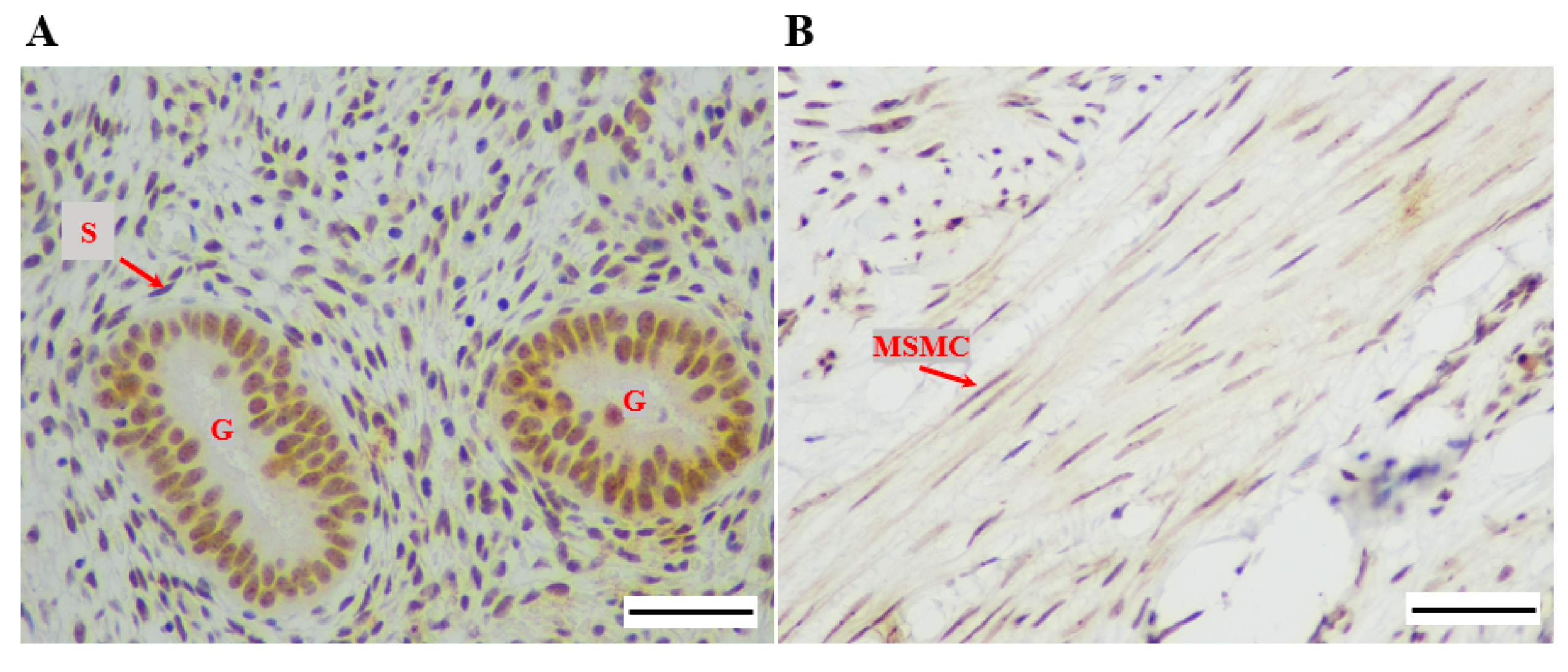
2.3. Immunohistochemistry (IHC) Staining and Analysis
2.4. Telomeric Repeat Amplification Protocol (TRAP) Assay
2.5. Statistical Analysis of the Ex-Vivo Study
2.6. In Silico Analysis of Endometrial Datasets and Identification of Differentially Regulated Genes in Mid-Secretory Phases
2.7. Identification of Key Transcription Factors
3. Results
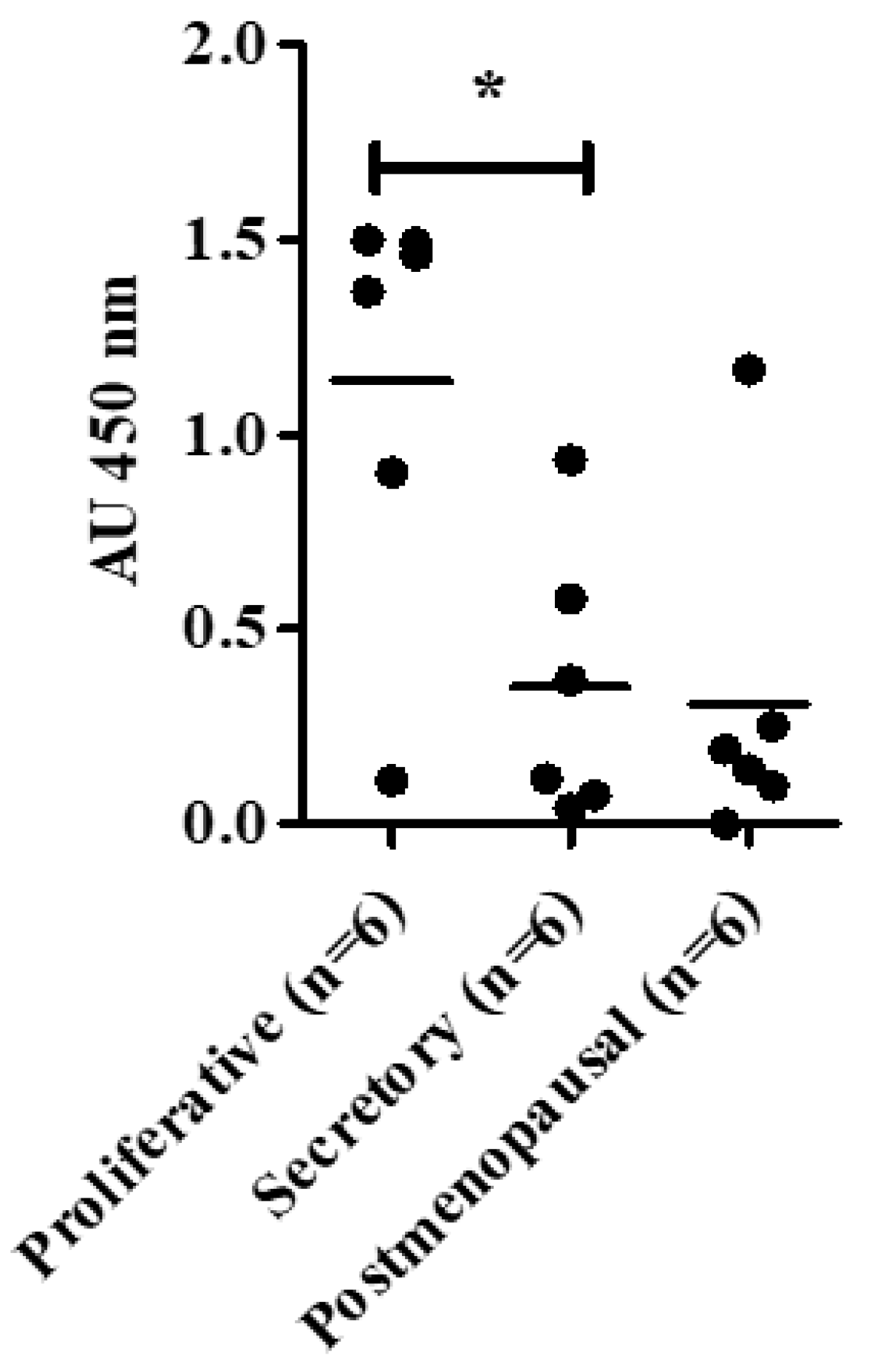
3.1. Telomerase Holoenzyme Component (hTERC and DKC1) Levels Demonstrate a Dynamic Expression Pattern in Healthy Endometrium
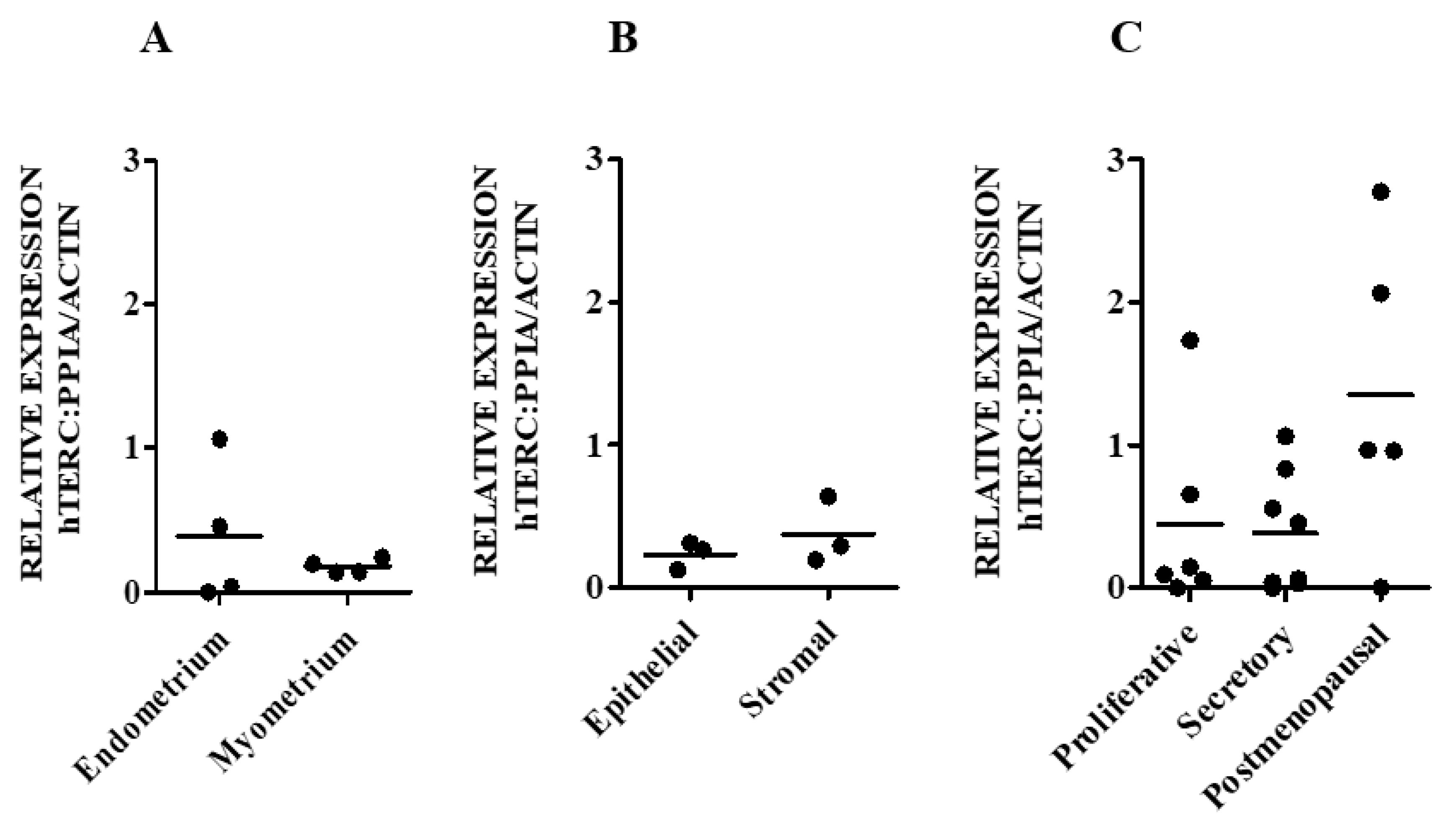
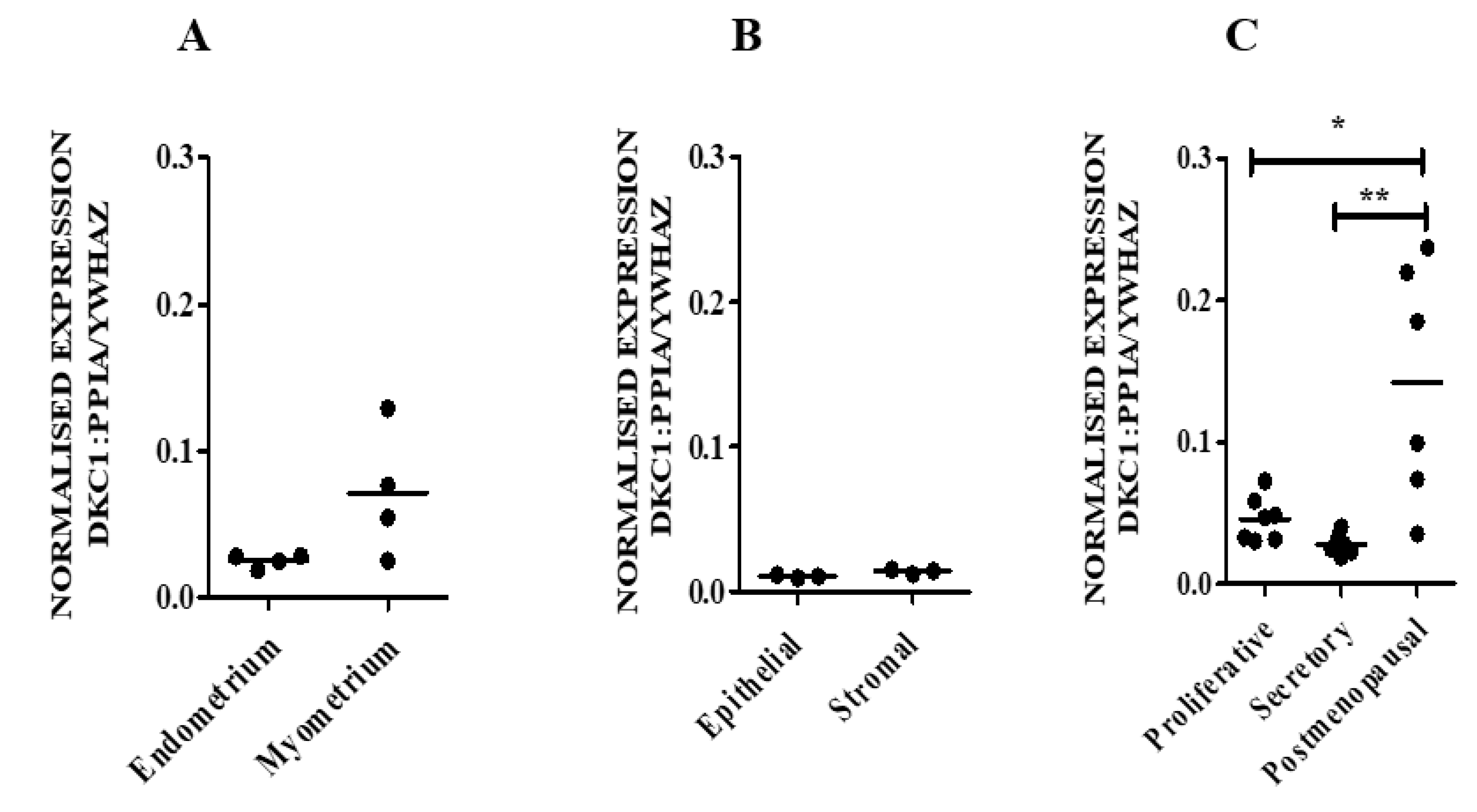
3.1.1. hTERC and DKC1 RNA
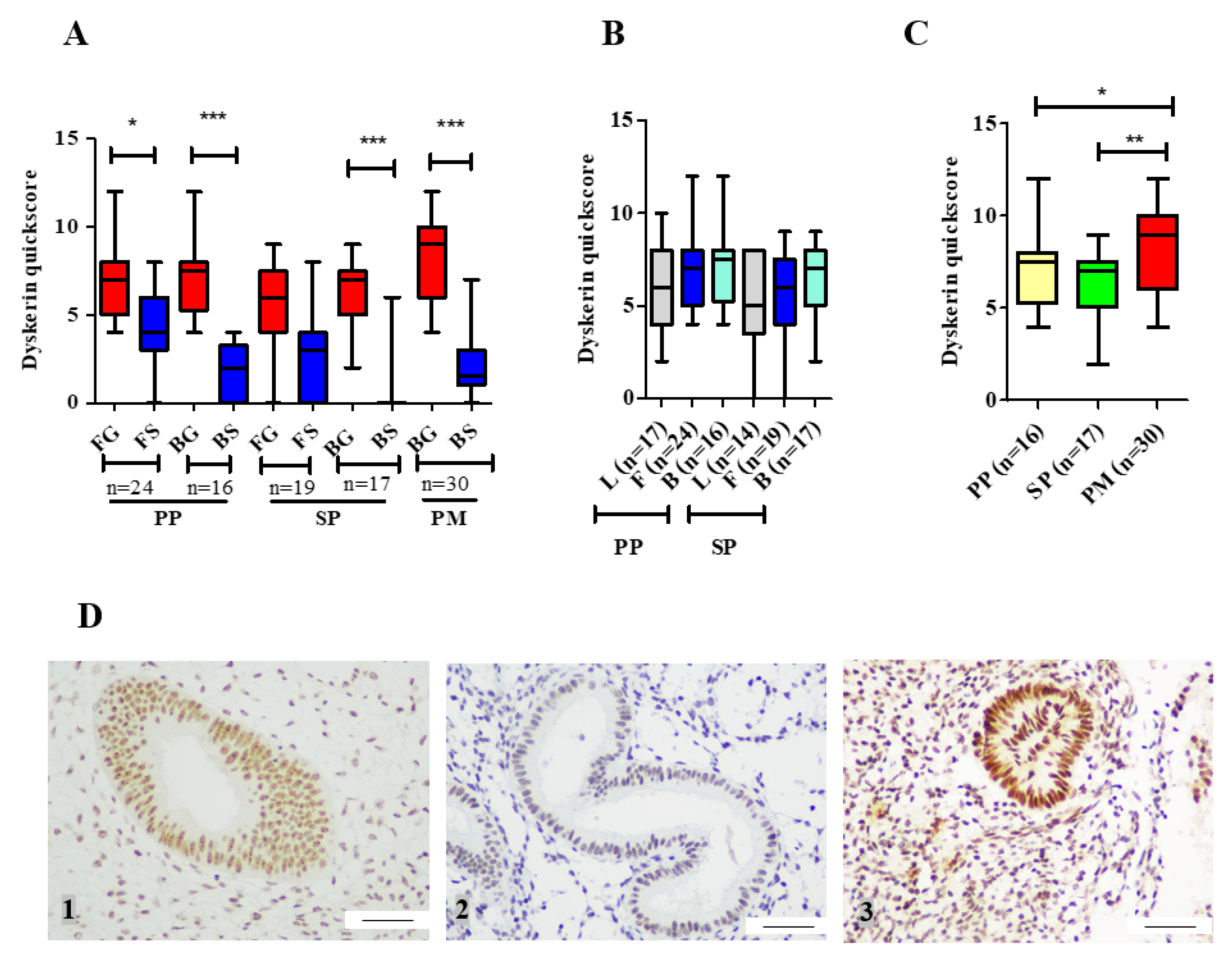
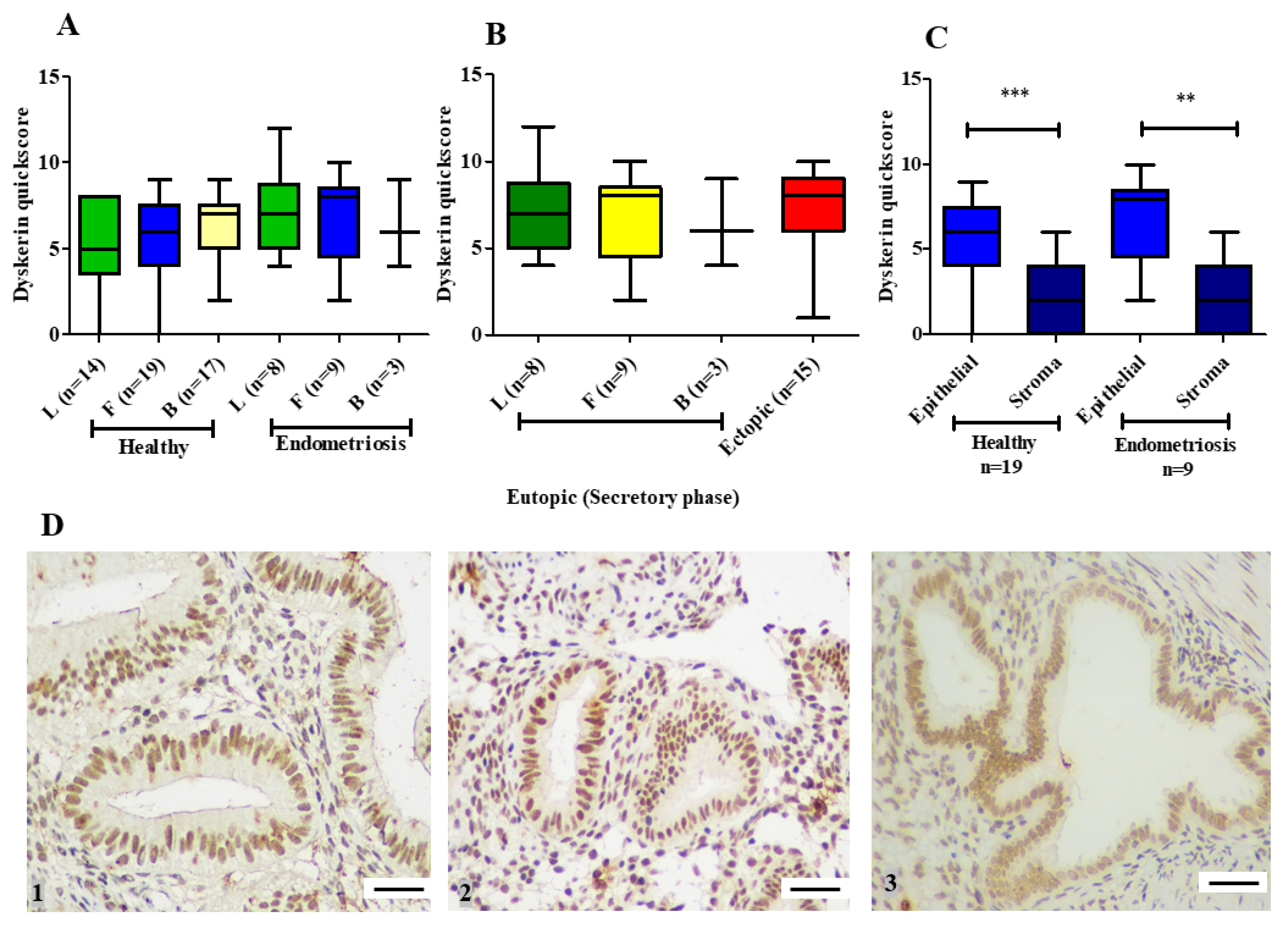
3.1.2. Dyskerin Protein
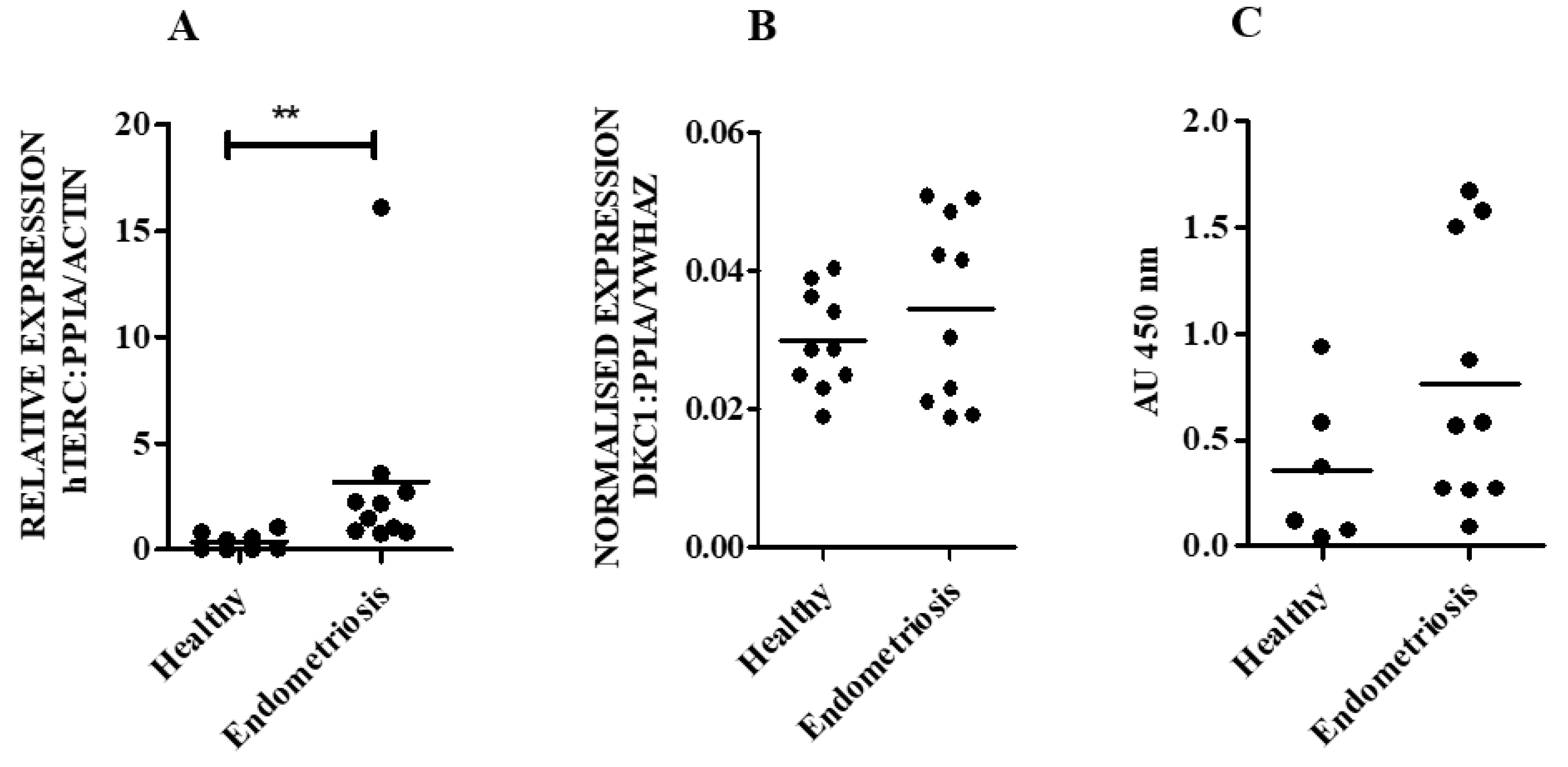
3.2. hTERC Levels Were Significantly Increased Without Affecting Dyskerin (DKC1) Levels in the Secretory Eutopic Endometrium of Women with Endometriosis
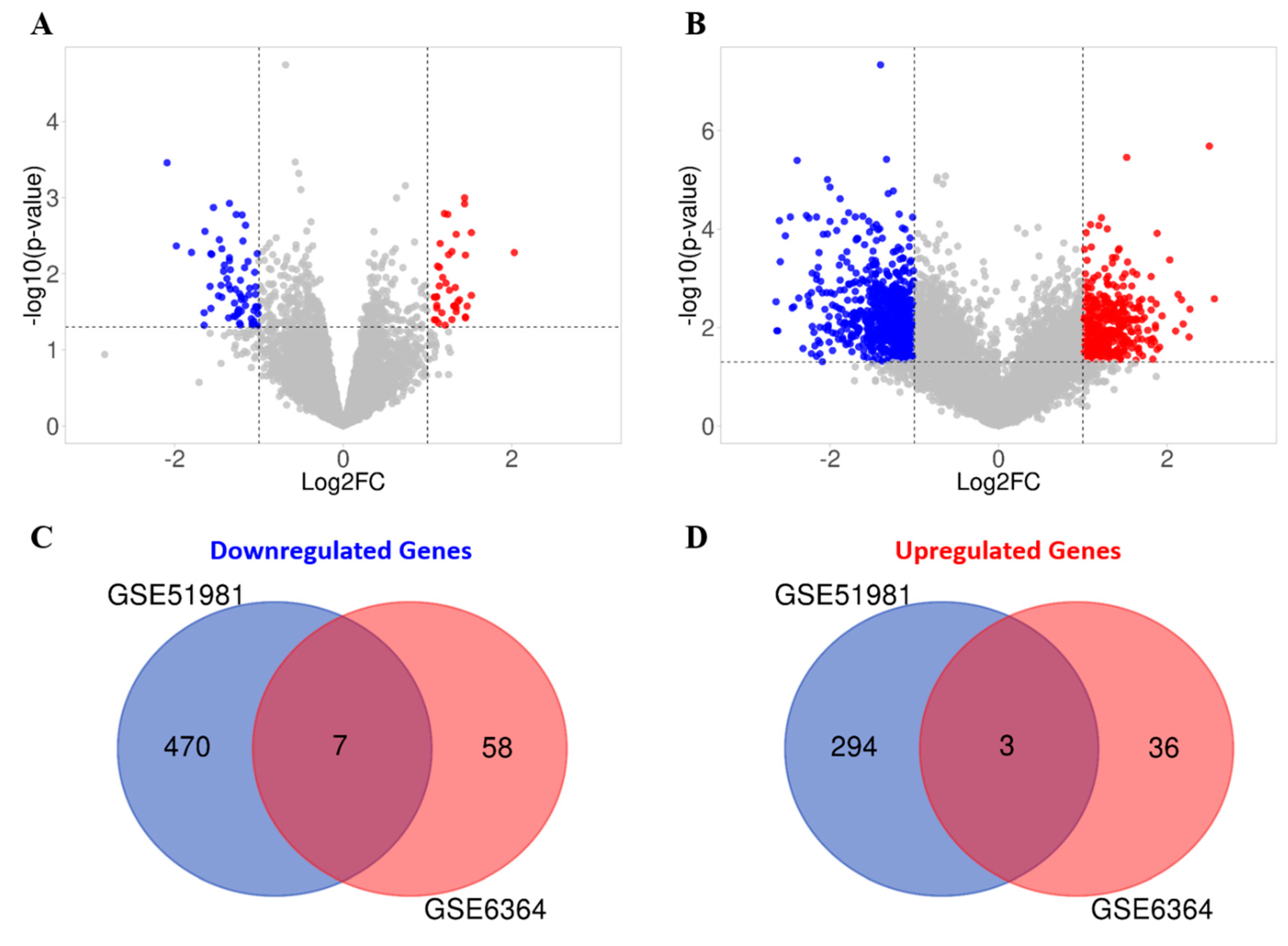
3.3. In Silico Interrogation of Publicly Available Datasets Demonstrates other Telomerase Associated Genes to Be Altered in the Mid-Secretory Endometrium of Women with Endometriosis
3.3.1. Functional Enrichment Analysis
3.3.2. Identification of Key Transcription Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H.; Gall, J.G. A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J. Mol. Biol. 1978, 120, 33–53. [Google Scholar] [CrossRef]
- Mandal, P. Recent advances of Blood telomere length (BTL) shortening: A potential biomarker for development of cancer. Pathol. Oncol. Res. 2019, 25, 1263–1265. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.B.; Graham, M.E.; Lovrecz, G.O.; Bache, N.; Robinson, P.J.; Reddel, R.R. Protein composition of catalytically active human telomerase from immortal cells. Science 2007, 315, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, A.J.; Saretzki, G.; Tempest, N.; Critchley, H.O.; Hapangama, D.K. Human endometrial epithelial telomerase is important for epithelial proliferation and glandular formation with potential implications in endometriosis. Hum. Reprod. 2015, 30, 2816–2828. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Takahashi, Y.; Morishita, S.; Hashimoto, M.; Niwa, K.; Tamaya, T. Telomerase activity in the human endometrium throughout the menstrual cycle. Mol. Hum. Reprod. 1998, 4, 173–177. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vidal, J.D.; Register, T.C.; Gupta, M.; Cline, J.M. Estrogen replacement therapy induces telomerase RNA expression in the macaque endometrium. Fertil. Steril. 2002, 77, 601–608. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Kamal, A.; Saretzki, G. Implications of telomeres and telomerase in endometrial pathology. Hum. Reprod. Update 2017, 23, 166–187. [Google Scholar] [CrossRef]
- Tanaka, M.; Kyo, S.; Takakura, M.; Kanaya, T.; Sagawa, T.; Yamashita, K.; Okada, Y.; Hiyama, E.; Inoue, M. Expression of telomerase activity in human endometrium is localized to epithelial glandular cells and regulated in a menstrual phase-dependent manner correlated with cell proliferation. Am. J. Pathol. 1998, 153, 1985–1991. [Google Scholar] [CrossRef]
- Sourial, S.; Tempest, N.; Hapangama, D.K. Theories on the Pathogenesis of Endometriosis. Int. J. Reprod. Med. 2014, 2014, 179515. [Google Scholar] [CrossRef]
- Hill, C.J.; Fakhreldin, M.; Maclean, A.; Dobson, L.; Nancarrow, L.; Bradfield, A.; Choi, F.; Daley, D.; Tempest, N.; Hapangama, D.K. Endometriosis and the Fallopian Tubes: Theories of Origin and Clinical Implications. J. Clin. Med. 2020, 9, 1905. [Google Scholar] [CrossRef]
- Parasar, P.; Ozcan, P.; Terry, K.L. Endometriosis: Epidemiology, Diagnosis and Clinical Management. Curr. Obstet. Gynecol. Rep. 2017, 6, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Metastatic or Embolic Endometriosis, due to the Menstrual Dissemination of Endometrial Tissue into the Venous Circulation. Am. J. Pathol. 1927, 3, 93–110.43. [Google Scholar] [PubMed]
- Halme, J.; Hammond, M.G.; Hulka, J.F.; Raj, S.G.; Talbert, L.M. Retrograde menstruation in healthy women and in patients with endometriosis. Obstet. Gynecol. 1984, 64, 151–154. [Google Scholar] [PubMed]
- Hapangama, D.K.; Turner, M.A.; Drury, J.; Heathcote, L.; Afshar, Y.; Mavrogianis, P.A.; Fazleabas, A.T. Aberrant expression of regulators of cell-fate found in eutopic endometrium is found in matched ectopic endometrium among women and in a baboon model of endometriosis. Hum. Reprod. 2010, 25, 2840–2850. [Google Scholar] [CrossRef] [PubMed]
- Hapangama, D.K.; Drury, J.; Da Silva, L.; Al-Lamee, H.; Earp, A.; Valentijn, A.J.; Edirisinghe, D.P.; Murray, P.A.; Fazleabas, A.T.; Gargett, C.E. Abnormally located SSEA1+/SOX9+ endometrial epithelial cells with a basalis-like phenotype in the eutopic functionalis layer may play a role in the pathogenesis of endometriosis. Hum. Reprod. 2019, 34, 56–68. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Raju, R.S.; Valentijn, A.J.; Barraclough, D.; Hart, A.; Turner, M.A.; Platt-Higgins, A.; Barraclough, R.; Rudland, P.S. Aberrant expression of metastasis-inducing proteins in ectopic and matched eutopic endometrium of women with endometriosis: Implications for the pathogenesis of endometriosis. Hum. Reprod. 2012, 27, 394–407. [Google Scholar] [CrossRef]
- Mathew, D.; Drury, J.A.; Valentijn, A.J.; Vasieva, O.; Hapangama, D.K. In silico, in vitro and in vivo analysis identifies a potential role for steroid hormone regulation of FOXD3 in endometriosis-associated genes. Hum. Reprod. 2016, 31, 345–354. [Google Scholar]
- Kim, C.M.; Oh, Y.J.; Cho, S.H.; Chung, D.J.; Hwang, J.Y.; Park, K.H.; Cho, D.J.; Choi, Y.M.; Lee, B.S. Increased telomerase activity and human telomerase reverse transcriptase mRNA expression in the endometrium of patients with endometriosis. Hum. Reprod. 2007, 22, 843–849. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Turner, M.A.; Drury, J.A.; Quenby, S.; Saretzki, G.; Martin-Ruiz, C.; Von Zglinicki, T. Endometriosis is associated with aberrant endometrial expression of telomerase and increased telomere length. Hum. Reprod. 2008, 23, 1511–1519. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Turner, M.A.; Drury, J.A.; Quenby, S.; Hart, A.; Maddick, M.; Martin-Ruiz, C.; von Zglinicki, T. Sustained replication in endometrium of women with endometriosis occurs without evoking a DNA damage response. Hum. Reprod. 2009, 24, 687–696. [Google Scholar] [CrossRef]
- Valentijn, A.J.; Palial, K.; Al-Lamee, H.; Tempest, N.; Drury, J.; Von Zglinicki, T.; Saretzki, G.; Murray, P.; Gargett, C.E.; Hapangama, D.K. SSEA-1 isolates human endometrial basal glandular epithelial cells: Phenotypic and functional characterization and implications in the pathogenesis of endometriosis. Hum. Reprod. 2013, 28, 2695–2708. [Google Scholar] [CrossRef] [PubMed]
- Bradfield, A.; Button, L.; Drury, J.; Green, D.C.; Hill, C.J.; Hapangama, D.K. Investigating the Role of Telomere and Telomerase Associated Genes and Proteins in Endometrial Cancer. Methods Protoc. 2020, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Guertler, R.; Naim, S.; Nixdorf, S.; Fedier, A.; Hacker, N.F.; Heinzelmann-Schwarz, V. Careful selection of reference genes is required for reliable performance of RT-qPCR in human normal and cancer cell lines. PLoS ONE 2013, 8, e59180. [Google Scholar] [CrossRef] [PubMed]
- Marullo, M.; Zuccato, C.; Mariotti, C.; Lahiri, N.; Tabrizi, S.J.; Di Donato, S.; Cattaneo, E. Expressed Alu repeats as a novel, reliable tool for normalization of real-time quantitative RT-PCR data. Genome Biol. 2010, 11, R9. [Google Scholar] [CrossRef]
- Turano, M.; Angrisani, A.; De Rosa, M.; Izzo, P.; Furia, M. Real-time PCR quantification of human DKC1 expression in colorectal cancer. Acta Oncol. 2008, 47, 1598–1599. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, L.; Lu, S. Role of TERRA in the regulation of telomere length. Int. J. Biol. Sci. 2015, 11, 316–323. [Google Scholar] [CrossRef]
- Chai, J.H.; Zhang, Y.; Tan, W.H.; Chng, W.J.; Li, B.; Wang, X. Regulation of hTERT by BCR-ABL at multiple levels in K562 cells. BMC Cancer 2011, 11, 512. [Google Scholar] [CrossRef]
- Romani, C.; Calza, S.; Todeschini, P.; Tassi, R.A.; Zanotti, L.; Bandiera, E.; Sartori, E.; Pecorelli, S.; Ravaggi, A.; Santin, A.D.; et al. Identification of optimal reference genes for gene expression normalization in a wide cohort of endometrioid endometrial carcinoma tissues. PLoS ONE 2014, 9, e113781. [Google Scholar] [CrossRef]
- Sadek, K.H.; Cagampang, F.R.; Bruce, K.D.; Shreeve, N.; Macklon, N.; Cheong, Y. Variation in stability of housekeeping genes in endometrium of healthy and polycystic ovarian syndrome women. Hum. Reprod. 2012, 27, 251–256. [Google Scholar] [CrossRef]
- Kamal, A.M.; Bulmer, J.N.; DeCruze, S.B.; Stringfellow, H.F.; Martin-Hirsch, P.; Hapangama, D.K. Androgen receptors are acquired by healthy postmenopausal endometrial epithelium and their subsequent loss in endometrial cancer is associated with poor survival. Br. J. Cancer 2016, 114, 688–696. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Tamaresis, J.S.; Irwin, J.C.; Goldfien, G.A.; Rabban, J.T.; Burney, R.O.; Nezhat, C.; DePaolo, L.V.; Giudice, L.C. Molecular classification of endometriosis and disease stage using high-dimensional genomic data. Endocrinology 2014, 155, 4986–4999. [Google Scholar] [CrossRef] [PubMed]
- Burney, R.O.; Talbi, S.; Hamilton, A.E.; Vo, K.C.; Nyegaard, M.; Nezhat, C.R.; Lessey, B.A.; Giudice, L.C. Gene expression analysis of endometrium reveals progesterone resistance and candidate susceptibility genes in women with endometriosis. Endocrinology 2007, 148, 3814–3826. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef]
- Ho Sui, S.J.; Mortimer, J.R.; Arenillas, D.J.; Brumm, J.; Walsh, C.J.; Kennedy, B.P.; Wasserman, W.W. oPOSSUM: Identification of over-represented transcription factor binding sites in co-expressed genes. Nucleic Acids Res. 2005, 33, 3154–3164. [Google Scholar] [CrossRef]
- Keilwagen, J.; Posch, S.; Grau, J. Accurate prediction of cell type-specific transcription factor binding. Genome Biol. 2019, 20, 9. [Google Scholar] [CrossRef]
- Avilion, A.A.; Piatyszek, M.A.; Gupta, J.; Shay, J.W.; Bacchetti, S.; Greider, C.W. Human telomerase RNA and telomerase activity in immortal cell lines and tumor tissues. Cancer Res. 1996, 56, 645–650. [Google Scholar]
- Ohyashiki, J.H.; Hisatomi, H.; Nagao, K.; Honda, S.; Takaku, T.; Zhang, Y.; Sashida, G.; Ohyashiki, K. Quantitative relationship between functionally active telomerase and major telomerase components (hTERT and hTR) in acute leukaemia cells. Br. J. Cancer 2005, 92, 1942–1947. [Google Scholar] [CrossRef]
- Bachand, F.; Autexier, C. Functional reconstitution of human telomerase expressed in Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 38027–38031. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lessey, B.A.; Kim, J.J. Endometrial receptivity in the eutopic endometrium of women with endometriosis: It is affected, and let me show you why. Fertil. Steril. 2017, 108, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Singh, V.; Tayade, C. Biomarkers in endometriosis: Challenges and opportunities. Fertil Steril 2017, 107, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Crothers, J.; Haqq, C.M.; Blackburn, E.H. Cellular and gene expression responses involved in the rapid growth inhibition of human cancer cells by RNA interference-mediated depletion of telomerase RNA. J. Biol. Chem. 2005, 280, 23709–23717. [Google Scholar] [CrossRef] [PubMed]
- Roake, C.M.; Artandi, S.E. Regulation of human telomerase in homeostasis and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 384–397. [Google Scholar] [CrossRef]
- Mochizuki, Y.; He, J.; Kulkarni, S.; Bessler, M.; Mason, P.J. Mouse dyskerin mutations affect accumulation of telomerase RNA and small nucleolar RNA, telomerase activity, and ribosomal RNA processing. Proc. Natl. Acad. Sci. USA 2004, 101, 10756–10761. [Google Scholar] [CrossRef]
- Shukla, S.; Schmidt, J.C.; Goldfarb, K.C.; Cech, T.R.; Parker, R. Inhibition of telomerase RNA decay rescues telomerase deficiency caused by dyskerin or PARN defects. Nat. Struct. Mol. Biol. 2016, 23, 286–292. [Google Scholar] [CrossRef]
- Montanaro, L.; Calienni, M.; Ceccarelli, C.; Santini, D.; Taffurelli, M.; Pileri, S.; Trere, D.; Derenzini, M. Relationship between dyskerin expression and telomerase activity in human breast cancer. Cell Oncol. 2008, 30, 483–490. [Google Scholar]
- Bellodi, C.; Krasnykh, O.; Haynes, N.; Theodoropoulou, M.; Peng, G.; Montanaro, L.; Ruggero, D. Loss of function of the tumor suppressor DKC1 perturbs p27 translation control and contributes to pituitary tumorigenesis. Cancer Res. 2010, 70, 6026–6035. [Google Scholar] [CrossRef]
- Martini, M.; Ciccarone, M.; Garganese, G.; Maggiore, C.; Evangelista, A.; Rahimi, S.; Zannoni, G.; Vittori, G.; Larocca, L.M. Possible involvement of hMLH1, p16(INK4a) and PTEN in the malignant transformation of endometriosis. Int. J. Cancer 2002, 102, 398–406. [Google Scholar] [CrossRef]
- Huschtscha, L.I.; Reddel, R.R. p16(INK4a) and the control of cellular proliferative life span. Carcinogenesis 1999, 20, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Alnafakh, R.A.A.; Adishesh, M.; Button, L.; Saretzki, G.; Hapangama, D.K. Telomerase and Telomeres in Endometrial Cancer. Front. Oncol. 2019, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, G.A.; Camargo-Kosugi, C.M.; Bonetti, T.C.; Invitti, A.L.; Girão, M.J.; Silva, I.D.; Schor, E. p27kip1 overexpression regulates VEGF expression, cell proliferation and apoptosis in cell culture from eutopic endometrium of women with endometriosis. Apoptosis 2015, 20, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.; Osterwald, S.; Deeg, K.I.; Rippe, K. PML body meets telomere: The beginning of an ALTernate ending? Nucleus 2012, 3, 263–275. [Google Scholar] [CrossRef]
- Loe, T.K.; Li, J.S.Z.; Zhang, Y.; Azeroglu, B.; Boddy, M.N.; Denchi, E.L. Telomere length heterogeneity in ALT cells is maintained by PML-dependent localization of the BTR complex to telomeres. Genes Dev. 2020, 34, 650–662. [Google Scholar] [CrossRef]
- Cloutier, P.; Poitras, C.; Durand, M.; Hekmat, O.; Fiola-Masson, É.; Bouchard, A.; Faubert, D.; Chabot, B.; Coulombe, B. R2TP/Prefoldin-like component RUVBL1/RUVBL2 directly interacts with ZNHIT2 to regulate assembly of U5 small nuclear ribonucleoprotein. Nat. Commun. 2017, 8, 15615. [Google Scholar] [CrossRef]
- Abelson, J.; Hadjivassiliou, H.; Guthrie, C. Preparation of fluorescent pre-mRNA substrates for an smFRET study of pre-mRNA splicing in yeast. Methods Enzymol. 2010, 472, 31–40. [Google Scholar]
- Ludlow, A.T.; Slusher, A.L.; Sayed, M.E. Insights into Telomerase/hTERT Alternative Splicing Regulation Using Bioinformatics and Network Analysis in Cancer. Cancers 2019, 11, 666. [Google Scholar] [CrossRef]
- Taniguchi, F.; Itamochi, H.; Harada, T.; Terakawa, N. Fibroblast growth factor receptor 2 expression may be involved in transformation of ovarian endometrioma to clear cell carcinoma of the ovary. Int. J. Gynecol. Cancer 2013, 23, 791–796. [Google Scholar] [CrossRef]
- Gadducci, A.; Zannoni, G.F. Endometriosis-associated Extraovarian Malignancies: A Challenging Question for the Clinician and the Pathologist. Anticancer Res. 2020, 40, 2429–2438. [Google Scholar] [CrossRef]
- Gatbonton, T.; Imbesi, M.; Nelson, M.; Akey, J.M.; Ruderfer, D.M.; Kruglyak, L.; Simon, J.A.; Bedalov, A. Telomere length as a quantitative trait: Genome-wide survey and genetic mapping of telomere length-control genes in yeast. PLoS Genet 2006, 2, e35. [Google Scholar]
- Askree, S.H.; Yehuda, T.; Smolikov, S.; Gurevich, R.; Hawk, J.; Coker, C.; Krauskopf, A.; Kupiec, M.; McEachern, M.J. A genome-wide screen for Saccharomyces cerevisiae deletion mutants that affect telomere length. Proc. Natl. Acad. Sci. USA 2004, 101, 8658–8663. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Tang, H.B.; Liu, N.N.; Tong, X.J.; Dang, W.; Duan, Y.M.; Fu, X.H.; Zhang, Y.; Peng, J.; Meng, F.L.; et al. Telomerase-null survivor screening identifies novel telomere recombination regulators. PLoS Genet. 2013, 9, e1003208. [Google Scholar] [CrossRef] [PubMed]
- Mahipal, A.; Tella, S.H.; Kommalapati, A.; Anaya, D.; Kim, R. FGFR2 genomic aberrations: Achilles heel in the management of advanced cholangiocarcinoma. Cancer Treat. Rev. 2019, 78, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, M.; Zhang, L.; Ding, X.; Li, W.; Lu, S. HSPC159 promotes proliferation and metastasis by inducing epithelial-mesenchymal transition and activating the PI3K/Akt pathway in breast cancer. Cancer Sci. 2018, 109, 2153–2163. [Google Scholar] [CrossRef] [PubMed]
- Dowling, P.; Pollard, D.; Larkin, A.; Henry, M.; Meleady, P.; Gately, K.; O’Byrne, K.; Barr, M.P.; Lynch, V.; Ballot, J.; et al. Abnormal levels of heterogeneous nuclear ribonucleoprotein A2B1 (hnRNPA2B1) in tumour tissue and blood samples from patients diagnosed with lung cancer. Mol. Biosyst. 2015, 11, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Konrad, L.; Dietze, R.; Riaz, M.A.; Scheiner-Bobis, G.; Behnke, J.; Horné, F.; Hoerscher, A.; Reising, C.; Meinhold-Heerlein, I. Epithelial-Mesenchymal Transition in Endometriosis-When Does It Happen? J. Clin. Med. 2020, 9, 1915. [Google Scholar] [CrossRef]
- Uziel, O.; Yosef, N.; Sharan, R.; Ruppin, E.; Kupiec, M.; Kushnir, M.; Beery, E.; Cohen-Diker, T.; Nordenberg, J.; Lahav, M. The effects of telomere shortening on cancer cells: A network model of proteomic and microRNA analysis. Genomics 2015, 105, 5–16. [Google Scholar] [CrossRef]
- Lee, O.H.; Kim, H.; He, Q.; Baek, H.J.; Yang, D.; Chen, L.Y.; Liang, J.; Chae, H.K.; Safari, A.; Liu, D.; et al. Genome-wide YFP fluorescence complementation screen identifies new regulators for telomere signaling in human cells. Mol. Cell Proteom. 2011, 10, M110.001628. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres and telomerase: Their mechanisms of action and the effects of altering their functions. FEBS Lett. 2005, 579, 859–862. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, H.; Fu, B.; Disbrow, G.L.; Apolinario, T.; Tomaic, V.; Kelley, M.L.; Baker, C.C.; Huibregtse, J.; Schlegel, R. The E6AP ubiquitin ligase is required for transactivation of the hTERT promoter by the human papillomavirus E6 oncoprotein. J. Biol. Chem. 2005, 280, 10807–10816. [Google Scholar] [CrossRef] [PubMed]
- Veldman, T.; Liu, X.; Yuan, H.; Schlegel, R. Human papillomavirus E6 and Myc proteins associate in vivo and bind to and cooperatively activate the telomerase reverse transcriptase promoter. Proc. Natl. Acad. Sci. USA 2003, 100, 8211–8216. [Google Scholar] [CrossRef] [PubMed]
- Weinrich, S.L.; Pruzan, R.; Ma, L.; Ouellette, M.; Tesmer, V.M.; Holt, S.E.; Bodnar, A.G.; Lichtsteiner, S.; Kim, N.W.; Trager, J.B.; et al. Reconstitution of human telomerase with the template RNA component hTR and the catalytic protein subunit hTRT. Nat. Genet. 1997, 17, 498–502. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Groups | No. | * Age (years) | * BMI (kg /m2) |
|---|---|---|---|
| Proliferative phase | 27 | 40(30–57) | 27(18–41) |
| Secretory phase | 23 | 41(21–48) | 26(19–40) |
| Postmenopausal | 33 | 63(40–85) | 26(20–40) |
| Endometriosis | 34 | 24(24–48) | 24(11–35) |
| Primer | Sequence | Amplicon | Annealing Temp (°C) | Efficiency (%) |
|---|---|---|---|---|
| DKC1 | F:5’-CTCGGAAGTGGGGTTTAGGT-3 | 166 | 62 | 98 |
| R:5’-ACCACTTCAGCAACCACCTC-3 | ||||
| PPIA | F:5’-AGACAAGGTCCCAAAGAC-3 | 118 | 60 | 100.10 |
| R:5’-ACCACCCTGACACATAAA-3 | ||||
| YWHAZ | F:5’-CGTTACTTGGCTGAGGTTGCC-3 | 69 | 60 | 94.30 |
| R:5’-GTATGCTTGTTGTGACTGATCGAC-3 | ||||
| TERC | F:5’-GCCTTCCACCGTTCATTCTA-3 | 220 | 60 | 86.1 |
| R:5’-CCTGAAAGGCCTGAACCTC-3 | ||||
| ACTB | F:5’-TGTACGCCAACACAGTGCTG-3 | 183 | 60 | 94.6 |
| R:5’-GCTGGAAGGTGGACAGCGA-3 |
| Inclusion | Exclusion |
|---|---|
|
|
| Commonly Upregulated Genes | ||
| Gene Symbol | Gene ID | Mean Log2FC |
| CDKN2A | 1029 | 1.255 |
| PML | 5371 | 1.295 |
| ZNHIT2 | 741 | 1.315 |
| Commonly Downregulated Genes | ||
| Gene Symbol | Gene ID | Mean Log2FC |
| UBE3A | 7337 | −1.15 |
| MCCC2 | 64087 | −1.60 |
| HSPC159 | 29094 | −1.14 |
| FGFR2 | 2263 | −1.23 |
| PIK3C2A | 5286 | −1.55 |
| RALGAPA1 | 253959 | −1.19 |
| HNRNPA2B1 | 3181 | −1.29 |
| ID | Term | p-Value | |
|---|---|---|---|
| KEGG | map05200 | Pathways in cancer | 0.001932257 |
| map04120 | Ubiquitin mediated proteolysis | 0.002022009 | |
| map05203 | Viral carcinogenesis | 0.004288773 | |
| map04144 | Endocytosis | 0.006253438 | |
| Biological Process (GO) | GO:0090398 | Cellular senescence | 6.17 × 10–5 |
| GO:200058 | Regulation of protein ubiquitination involved in ubiquitin-dependent protein catabolic process | 6.71 × 10–5 | |
| GO:0032206 | Positive regulation of telomere maintenance | 1.11 × 10–4 | |
| GO:0042325 | Regulation of phosphorylation | 0.001531221 | |
| GO:0045926 | Negative regulation of growth | 0.001556759 | |
| GO:2000109 | Regulation of macrophage apoptotic process | 0.002996578 | |
| GO:0022612 | Gland morphogenesis | 0.002996578 | |
| GO:0050657 | Nucleic acid transport | 0.003495224 | |
| GO:0006551 | Leucine metabolic process | 0.003495224 | |
| GO:0050847 | Progesterone receptor signalling pathway | 0.003495224 | |
| Molecular Function (GO) | GO:0016303 | 1-phosphatidylinositol-3-kinase activity | 2.01 × 10–4 |
| GO:0052742 | Phosphatidylinositol kinase activity | 2.83 × 10–4 | |
| GO:0035004 | Phosphatidylinositol 3-kinase activity | 6.29 × 10–4 | |
| GO:0035005 | 1-phosphatidylinositol-4-phosphate 3-kinase activity | 0.003495224 | |
| GO:1990247 | N6-methyladenosine-containing RNA binding | 0.003495224 | |
| GO:0098505 | G-rich strand telomeric DNA binding | 0.004989818 | |
| GO:0004861 | Cyclin-dependent protein serine/threonine kinase inhibitor activity | 0.004989818 | |
| GO:0032183 | SUMO binding | 0.005985094 | |
| GO:0043047 | Single-stranded telomeric DNA binding | 0.005985094 | |
| GO:0016307 | Phosphatidylinositol phosphate kinase activity | 0.007476328 | |
| Cellular Component (GO) | GO:0000781 | Chromosome, telomeric region | 0.00166095 |
| Transcription Factor | Fisher Score |
|---|---|
| SOX2 | 5.904 |
| POU5F1 | 5.826 |
| RORA_2 | 4.746 |
| HNF1A | 4.476 |
| PPARG::RXRA | 4.455 |
| Spz1 | 4.244 |
| MEF2A | 4.111 |
| MIZF | 3.906 |
| ESR2 | 3.880 |
| HIF1A::ARNT | 3.805 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alnafakh, R.; Choi, F.; Bradfield, A.; Adishesh, M.; Saretzki, G.; Hapangama, D.K. Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study. Biomedicines 2020, 8, 588. https://doi.org/10.3390/biomedicines8120588
Alnafakh R, Choi F, Bradfield A, Adishesh M, Saretzki G, Hapangama DK. Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study. Biomedicines. 2020; 8(12):588. https://doi.org/10.3390/biomedicines8120588
Chicago/Turabian StyleAlnafakh, Rafah, Fiona Choi, Alice Bradfield, Meera Adishesh, Gabriele Saretzki, and Dharani K. Hapangama. 2020. "Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study" Biomedicines 8, no. 12: 588. https://doi.org/10.3390/biomedicines8120588
APA StyleAlnafakh, R., Choi, F., Bradfield, A., Adishesh, M., Saretzki, G., & Hapangama, D. K. (2020). Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study. Biomedicines, 8(12), 588. https://doi.org/10.3390/biomedicines8120588