Protein Nanotubes: From Bionanotech towards Medical Applications




Abstract
1. Introduction
2. Protein Nanofibers and Nanotubes (NTs) from Bacterial Systems
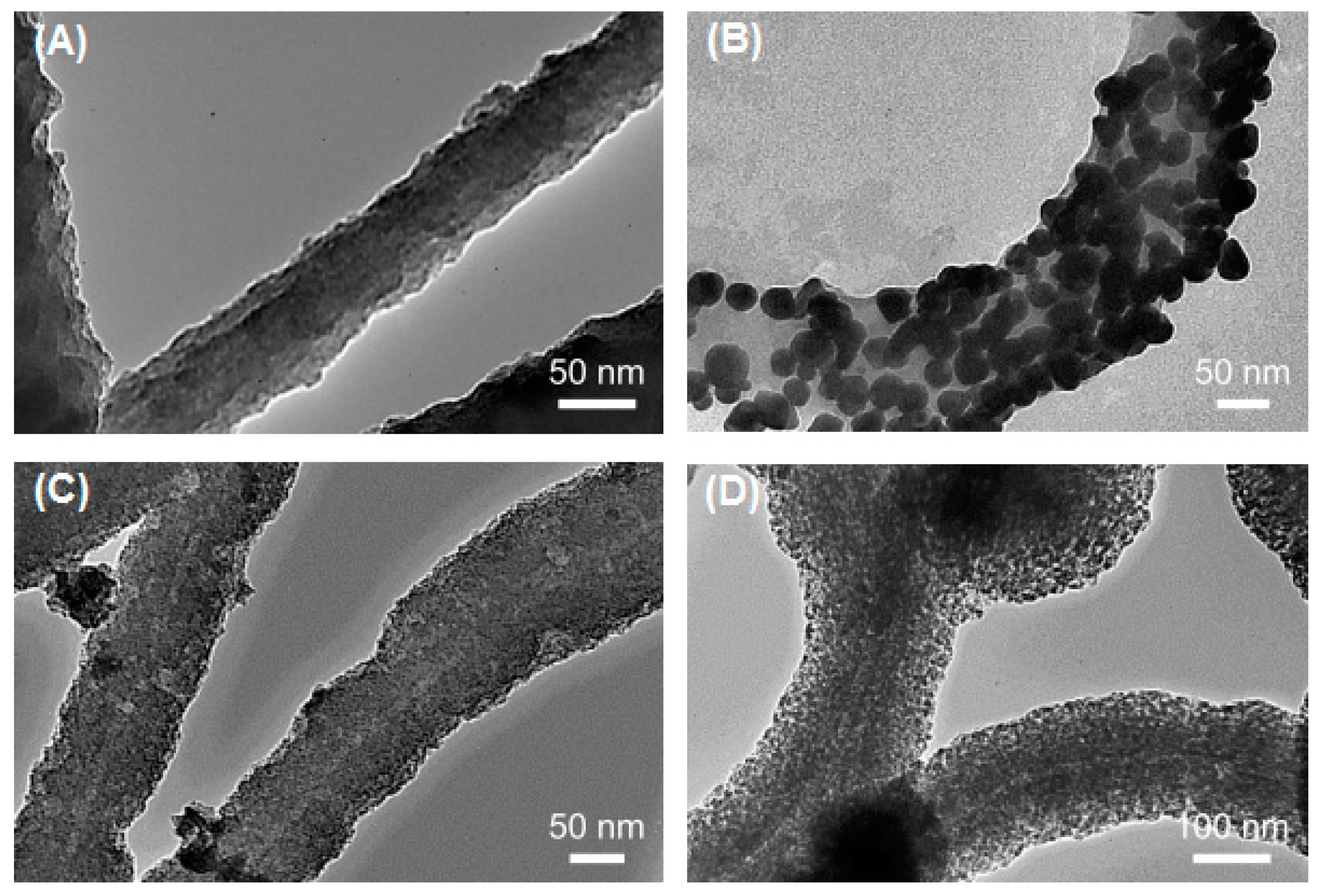
2.1. Flagella-Based Protein Nanofibers and Nanotubes
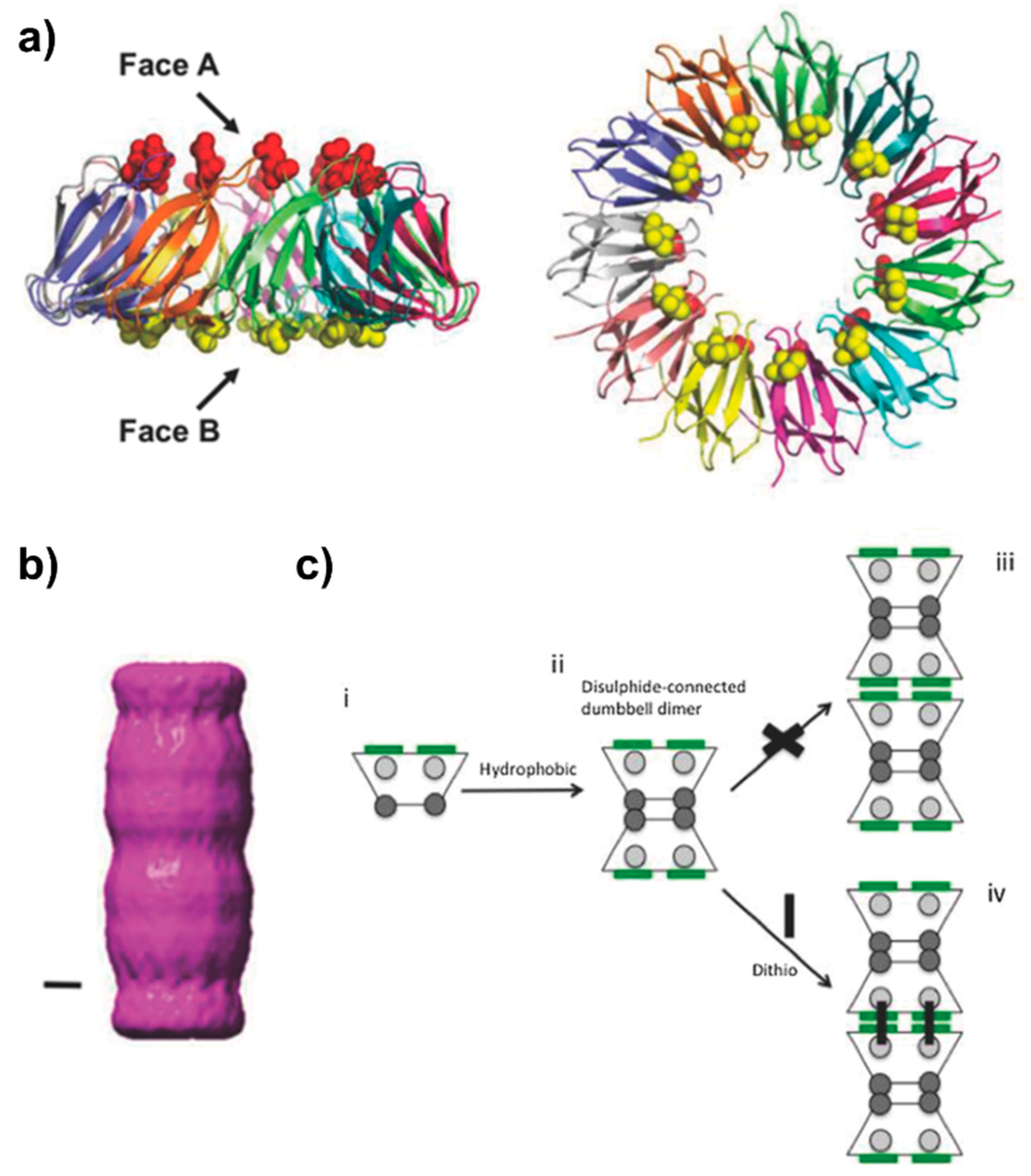
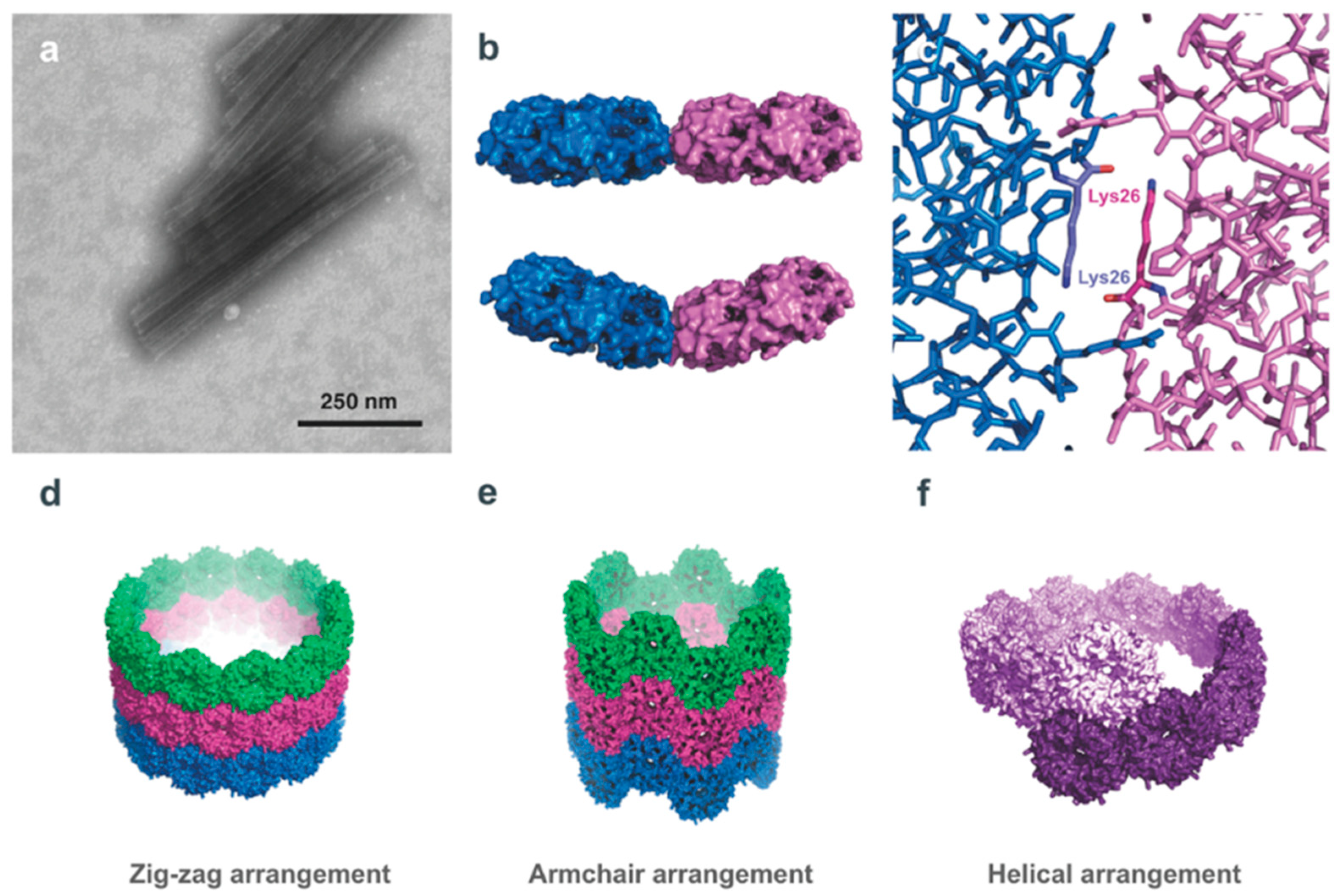
2.2. Pilin-Based Protein Nanotubes
3. Virus-Based Protein Nanotubes (PNTs)
3.1. Tobacco Mosaic Virus (TMV)
3.2. Cowpea Mosaic Virus (CPMV)
3.3. M13 Bacteriophage
4. Self-Assembling PNTs
4.1. The trp RNA Binding Attenuation Protein (TRAP) Nanotube
4.2. Microcompartment Proteins PduA and PduB
4.3. Hcp1 Nanotubes
4.4. Stable Protein 1 (SP1) Nanotubes
4.5. Self-Assembling Protein Nanoparticle (SAPN) Malaria Vaccine
4.6. Bacterial Gas Vesicles
5. Eukaryotic Systems
5.1. Kinesin-Microtubule Based Systems
5.2. Amyloid Fibrils
5.3. Silk Proteins Sericin and Fibroin
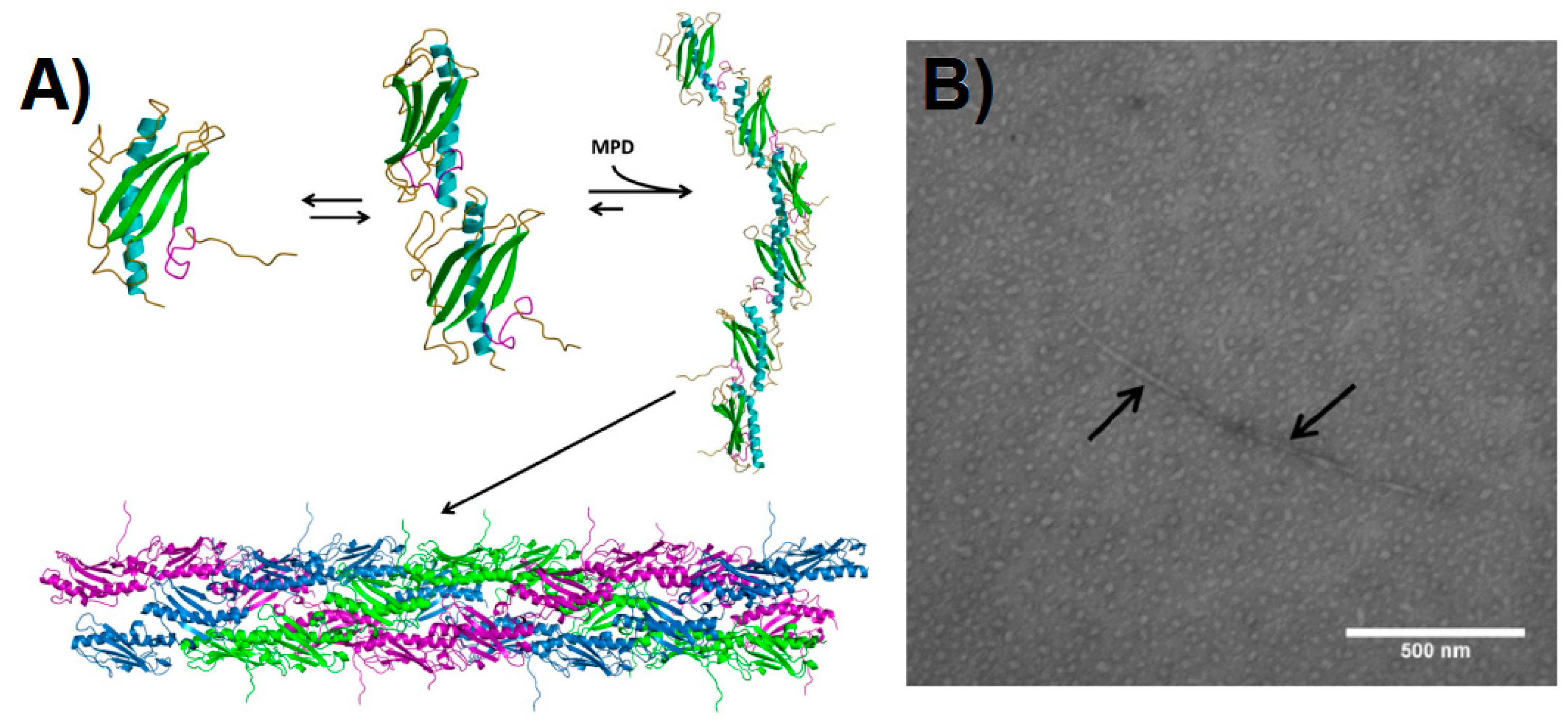
5.4. Human Insulin-Like Growth Factor Binding Protein-2 (hIGFBP-2)
6. Medical Applications of Protein and Peptide NTs
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanderson, K. Bioengineering: What to make with DNA origami. Nature 2010, 464, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Bell, N.A.W.; Keyser, U.F. Nanopores formed by DNA origami: A review. FEBS Lett. 2014, 588, 3564–3570. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, Q.; Liu, S.; Zhang, Y.; Tian, Y.; Song, C.; Wang, J.; Zou, Y.; Anderson, G.J.; Han, J.-Y.Y.; et al. A DNA nanorobot functions as a cancer therapeutic in response to a molecular trigger in vivo. Nat. Biotechnol. 2018, 36, 258–264. [Google Scholar] [CrossRef]
- Gerling, T.; Wagenbauer, K.F.; Neuner, A.M.; Dietz, H. Dynamic DNA devices and assemblies formed by shape-complementary, non–base pairing 3D components. Science 2015, 347, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Mashaghi, S.; Jadidi, T.; Koenderink, G.; Mashaghi, A. Lipid Nanotechnology. Int. J. Mol. Sci. 2013, 14, 4242–4282. [Google Scholar] [CrossRef] [PubMed]
- Tesauro, D.; Accardo, A.; Diaferia, C.; Milano, V.; Guillon, J.; Ronga, L.; Rossi, F. Peptide-Based Drug-Delivery Systems in Biotechnological Applications: Recent Advances and Perspectives. Molecules 2019, 24, 351. [Google Scholar] [CrossRef] [PubMed]
- De Frates, K.; Markiewicz, T.; Gallo, P.; Rack, A.; Weyhmiller, A.; Jarmusik, B.; Hu, X. Protein polymer-based nanoparticles: Fabrication and medical applications. Int. J. Mol. Sci. 2018, 19, 1717. [Google Scholar] [CrossRef]
- Petrov, A.; Audette, G.F. Peptide and protein-based nanotubes for nanobiotechnology. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2012, 4, 575–585. [Google Scholar] [CrossRef]
- Oliveira, S.F.; Bisker, G.; Bakh, N.A.; Gibbs, S.L.; Landry, M.P.; Strano, M.S. Protein functionalized carbon nanomaterials for biomedical applications. Carbon NY 2015, 95, 767–779. [Google Scholar] [CrossRef]
- Besteman, K.; Lee, J.; Wiertz, F.G.M.; Heering, H.A.; Dekker, C. Enzyme-Coated Carbon Nanotubes as Single-Molecule Biosensors. Nano Lett. 2003, 3, 727–730. [Google Scholar] [CrossRef]
- Unnikrishnan, B.; Palanisamy, S.; Chen, S.-M. A simple electrochemical approach to fabricate a glucose biosensor based on graphene–glucose oxidase biocomposite. Biosens. Bioelectron. 2013, 39, 70–75. [Google Scholar] [CrossRef]
- Barton, S.C.; Gallaway, J.; Atanassov, P. Enzymatic Biofuel Cells for Implantable and Microscale Devices. Chem. Rev. (Washington, DC) 2004, 104, 4867–4886. [Google Scholar] [CrossRef]
- Kannan, A.M.; Renugopalakrishnan, V.; Filipek, S.; Li, P.; Audette, G.F.; Munukutla, L. Bio-Batteries and Bio-Fuel Cells: Leveraging on Electronic Charge Transfer Proteins. J. Nanosci. Nanotechnol. 2009, 9, 1665–1678. [Google Scholar] [CrossRef] [PubMed]
- Dudzik, J.; Chang, W.-C.; Kannan, A.M.; Filipek, S.; Viswanathan, S.; Li, P.; Renugopalakrishnan, V.; Audette, G.F. Cross-linked glucose oxidase clusters for biofuel cell anode catalysts. Biofabrication 2013, 5, 035009. [Google Scholar] [CrossRef]
- Audette, G.F.; Hazes, B. Development of Protein Nanotubes from a Multi-Purpose Biological Structure. J. Nanosci. Nanotechnol. 2007, 7, 2222–2229. [Google Scholar] [CrossRef]
- Miranda, F.F.; Iwasaki, K.; Akashi, S.; Sumitomo, K.; Kobayashi, M.; Yamashita, I.; Tame, J.R.H.; Heddle, J.G. A Self-Assembled Protein Nanotube with High Aspect Ratio. Small 2009, 5, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Hopcroft, N.H.; Manfredo, A.; Wendt, A.L.; Brzozowski, A.M.; Gollnick, P.; Antson, A.A. The Interaction of RNA with TRAP: The Role of Triplet Repeats and Separating Spacer Nucleotides. J. Mol. Biol. 2004, 338, 43–53. [Google Scholar] [CrossRef]
- Nagano, S.; Banwell, E.F.; Iwasaki, K.; Michalak, M.; Pałka, R.; Zhang, K.Y.J.; Voet, A.R.D.; Heddle, J.G. Understanding the Assembly of an Artificial Protein Nanotube. Adv. Mater. Interfaces 2016, 3, 1600846. [Google Scholar] [CrossRef]
- Ballister, E.R.; Lai, A.H.; Zuckermann, R.N.; Cheng, Y.; Mougous, J.D. In vitro self-assembly of tailorable nanotubes from a simple protein building block. Proc. Natl. Acad. Sci. USA 2008, 105, 3733–3738. [Google Scholar] [CrossRef]
- Medalsy, I.; Dgany, O.; Sowwan, M.; Cohen, H.; Yukashevska, A.; Wolf, S.G.; Wolf, A.; Koster, A.; Almog, O.; Marton, I.; et al. SP1 protein-based nanostructures and arrays. Nano Lett. 2008, 8, 473–477. [Google Scholar] [CrossRef]
- Uddin, I.; Frank, S.; Warren, M.J.; Pickersgill, R.W. A Generic Self-Assembly Process in Microcompartments and Synthetic Protein Nanotubes. Small 2018, 14, 1704020. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, K.; Maki-Yonekura, S.; Namba, K. Complete atomic model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2003, 424, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.M.; Rossman, F.M.; Ferreira, J.L.; Matthews-Palmer, T.R.; Beeby, M.; Pallen, M.J. Bacterial Flagellins: Does Size Matter? Trends Microbiol. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Beatson, S.A.; Minamino, T.; Pallen, M.J. Variation in bacterial flagellins: From sequence to structure. Trends Microbiol. 2006, 14, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, K.; Maki, S.; Morgan, D.G.; DeRosier, D.J.; Vonderviszt, F.; Imada, K.; Namba, K. The bacterial flagellar cap as the rotary promoter of flagellin self-assembly. Science 2000, 290, 2148–2152. [Google Scholar] [CrossRef] [PubMed]
- Hoshikawa, H.; Kamiya, R. Elastic properties of bacterial flagellar filaments. II. Determination of the modulus of rigidity. Biophys. Chem. 1985, 22, 159–166. [Google Scholar] [CrossRef]
- Kamiya, R.; Asakura, S. Flagella transformations at alkaline pH. J. Mol. Biol. 1977, 108, 513–518. [Google Scholar] [CrossRef]
- Kamiya, R.; Asakura, S. Helical Transformations of Salmonella Flagella in vitro. J. Mol. Biol. 1976, 106, 167–186. [Google Scholar] [CrossRef]
- Kumara, M.T.; Srividya, N.; Muralidharan, S.; Tripp, B.C. Bioengineered flagella protein nanotubes with cysteine loops: Self-assembly and manipulation in an optical trap. Nano Lett. 2006, 6, 2121–2129. [Google Scholar] [CrossRef]
- Kumara, M.T.; Tripp, B.C.; Muralidharan, S. Layer-by-layer assembly of bioengineered flagella protein nanotubes. Biomacromolecules 2007, 8, 3718–3722. [Google Scholar] [CrossRef]
- Kumara, M.T.; Tripp, B.C.; Muralidharan, S. Self-Assembly of Metal Nanoparticles and Nanotubes on Bioengineered Flagella Scaffolds. Chem. Mater. 2007, 19, 2056–2064. [Google Scholar] [CrossRef]
- Jo, W.; Freedman, K.J.; Yi, D.K.; Kim, M.J. Fabrication of tunable silica-mineralized nanotubes using flagella as bio-templates. Nanotechnology 2012, 23, 55601. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.; Darmawan, M.; Kim, J.; Ahn, C.W.; Byun, D.; Baik, S.H.; Kim, M.J. Electrical property measurements of metallized flagella-templated silica nanotube networks. Nanotechnology 2013, 24, 135704. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kolappan, S.; Coureuil, M.; Yu, X.; Nassif, X.; Egelman, E.H.; Craig, L. Structure of the neisseria meningitidis type IV pilus. Nat. Commun. 2016, 7, 13015. [Google Scholar] [CrossRef] [PubMed]
- Folkhard, W.; Marvin, D.A.; Watts, T.H.; Paranchych, W. Structure of polar pili from Pseudomonas aeruginosa strains K and O. J. Mol. Biol. 1981, 149, 79–93. [Google Scholar] [CrossRef]
- Craig, L.; Taylor, R.K.; Pique, M.E.; Adair, B.D.; Arvai, A.S.; Singh, M.; Lloyd, S.J.; Shin, D.S.; Getzoff, E.D.; Yeager, M.; et al. Type IV pilin structure and assembly: X-ray and EM analyses of Vibrio cholerae toxin-coregulated pilus and Pseudomonas aeruginosa PAK pilin. Mol. Cell 2003, 11, 1139–1150. [Google Scholar] [CrossRef]
- Burrows, L.L. Pseudomonas aeruginosa Twitching Motility: Type IV Pili in Action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef]
- Harvey, H.; Habash, M.; Aidoo, F.; Burrows, L.L. Single-Residue Changes in the C-Terminal Disulfide-Bonded Loop of the Pseudomonas aeruginosa Type IV Pilin Influence Pilus Assembly and Twitching Motility. J. Bacteriol. 2009, 191, 6513–6524. [Google Scholar] [CrossRef]
- Craig, L.; Li, J. Type IV pili: Paradoxes in form and function. Curr. Opin. Struct. Biol. 2008, 18, 267–277. [Google Scholar] [CrossRef]
- Burrows, L.L. Weapons of mass retraction. Mol. Microbiol. 2005, 57, 878–888. [Google Scholar] [CrossRef]
- Craig, L.; Pique, M.E.; Tainer, J.A. Type IV pilus structure and bacterial pathogenicity. Nat. Rev. Microbiol. 2004, 2, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Hazes, B.; Sastry, P.A.; Hayakawa, K.; Read, R.J.; Irvin, R.T. Crystal structure of Pseudomonas aeruginosa PAK pilin suggests a main-chain-dominated mode of receptor binding. J. Mol. Biol. 2000, 299, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, K.V.; Irvin, R.T.; Hazes, B. Pros and cons of cryocrystallography: Should we also collect a room-temperature data set? Acta Crystallogr. Sect. D Biol. Crystallogr. 2005, 61, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Keizer, D.W.; Slupsky, C.M.; Kalisiak, M.; Campbell, A.P.; Crump, M.P.; Sastry, P.A.; Hazes, B.; Irvin, R.T.; Sykes, B.D. Structure of a Pilin Monomer from Pseudomonas aeruginosa: Implications for the assembly of pili. J. Biol. Chem. 2001, 276, 24186–24193. [Google Scholar] [CrossRef] [PubMed]
- Audette, G.F.; Irvin, R.T.; Hazes, B. Crystallographic analysis of the Pseudomonas aeruginosa strain K122-4 monomeric pilin reveals a conserved receptor-binding architecture. Biochemistry 2004, 43, 11427–11435. [Google Scholar] [CrossRef] [PubMed]
- Kao, D.J.; Churchill, M.E.A.; Irvin, R.T.; Hodges, R.S. Animal Protection and Structural Studies of a Consensus Sequence Vaccine Targeting the Receptor Binding Domain of the Type IV Pilus of Pseudomonas aeruginosa. J. Mol. Biol. 2007, 374, 426–442. [Google Scholar] [CrossRef]
- Nguyen, Y.; Jackson, S.G.; Aidoo, F.; Junop, M.; Burrows, L.L. Structural Characterization of Novel Pseudomonas aeruginosa Type IV Pilins. J. Mol. Biol. 2010, 395, 491–503. [Google Scholar] [CrossRef]
- Parge, H.E.; Forest, K.T.; Hickey, M.J.; Christensen, D.A.; Getzoff, E.D.; Tainer, J.A. Structure of the fibre-forming protein pilin at 2.6 A resolution. Nature 1995, 378, 32–38. [Google Scholar] [CrossRef]
- Piepenbrink, K.H.; Maldarelli, G.A.; Martinez de la Peña, C.F.; Dingle, T.C.; Mulvey, G.L.; Lee, A.; von Rosenvinge, E.; Armstrong, G.D.; Donnenberg, M.S.; Sundberg, E.J. Structural and Evolutionary Analyses Show Unique Stabilization Strategies in the Type IV Pili of Clostridium difficile. Structure 2015, 23, 385–396. [Google Scholar] [CrossRef]
- Piepenbrink, K.H.; Maldarelli, G.A.; Martinez de la Peña, C.F.; Mulvey, G.L.; Snyder, G.A.; De Masi, L.; von Rosenvinge, E.C.; Günther, S.; Armstrong, G.D.; Donnenberg, M.S.; et al. Structure of Clostridium difficile PilJ Exhibits Unprecedented Divergence from Known Type IV Pilins. J. Biol. Chem. 2014, 289, 4334–4345. [Google Scholar] [CrossRef]
- Wang, F.; Coureuil, M.; Osinski, T.; Orlova, A.; Altindal, T.; Gesbert, G.; Nassif, X.; Egelman, E.H.; Craig, L. Cryoelectron Microscopy Reconstructions of the Pseudomonas aeruginosa and Neisseria gonorrhoeae Type IV Pili at Sub-nanometer Resolution. Structure 2017, 25, 1423–1435. [Google Scholar] [CrossRef] [PubMed]
- Audette, G.F.; Van Schaik, E.J.; Hazes, B.; Irvin, R.T. DNA-binding protein nanotubes: Learning from nature’s nanotech examples. Nano Lett. 2004, 4, 1897–1902. [Google Scholar] [CrossRef]
- Lombardo, S.; Jasbi, S.Z.; Jeung, S.; Morin, S.; Audette, G.F. Initial Studies of Protein Nanotube Oligomerization from a Modified Gold Surface. J. Bionanosci. 2009, 3, 61–65. [Google Scholar] [CrossRef]
- Petrov, A.; Lombardo, S.; Audette, G.F. Fibril-mediated oligomerization of pilin-derived protein nanotubes. J. Nanobiotechnol. 2013, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Lento, C.; Wilson, D.J.; Audette, G.F. Dimerization of the type IV pilin from Pseudomonas aeruginosa strain K122-4 results in increased helix stability as measured by time-resolved hydrogen-deuterium exchange. Struct. Dyn. 2016, 3, 012001. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Giltner, C.L.; Van Schaik, E.J.; Bautista, D.L.; Hodges, R.S.; Audette, G.F.; Li, D.Y.; Irvin, R.T. A Novel Biometallic Interface: High Affinity Tip-AssociatedBinding by Pilin-Derived Protein Nanotubes. J. Bionanosci. 2007, 1, 73–83. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Reguera, G. Harnessing the power of microbial nanowires. Microb. Biotechnol. 2018, 1–16. [Google Scholar] [CrossRef]
- Yi, H.; Nevin, K.P.; Kim, B.-C.C.; Franks, A.E.; Klimes, A.; Tender, L.M.; Lovley, D.R. Selection of a variant of Geobacter sulfurreducens with enhanced capacity for current production in microbial fuel cells. Biosens. Bioelectron. 2009, 24, 3498–3503. [Google Scholar] [CrossRef]
- Malvankar, N.S.; Vargas, M.; Nevin, K.P.; Franks, A.E.; Leang, C.; Kim, B.-C.C.; Inoue, K.; Mester, T.; Covalla, S.F.; Johnson, J.P.; et al. Tunable metallic-like conductivity in microbial nanowire networks. Nat. Nanotechnol. 2011, 6, 573–579. [Google Scholar] [CrossRef]
- Malvankar, N.S.; Mester, T.; Tuominen, M.T.; Lovley, D.R. Supercapacitors Based on C-Type Cytochromes Using Conductive Nanostructured Networks of Living Bacteria. Chem. Phys. Chem. 2012, 13, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Giltner, C.L.; van Schaik, E.J.; Audette, G.F.; Kao, D.; Hodges, R.S.; Hassett, D.J.; Irvin, R.T. The Pseudomonas aeruginosa type IV pilin receptor binding domain functions as an adhesin for both biotic and abiotic surfaces. Mol. Microbiol. 2006, 59, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Young, M.; Willits, D.; Uchida, M.; Douglas, T. Plant viruses as biotemplates for materials and their use in nanotechnology. Annu. Rev. Phytopathol. 2008, 46, 361–384. [Google Scholar] [CrossRef] [PubMed]
- Fraenkel-Conrat, H.; Williams, R.C. Reconstitution of Active Tobacco Mosaic Virus from Its Inactive Protein and Nucleic Acid Components. Pro.c Natl. Acad. Sci. USA 1955, 41, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Perham, R.N.; Wilson, T.M. The characterization of intermediates formed during the disassembly of tobacco mosaic virus at alkaline pH. Virology 1978, 84, 293–302. [Google Scholar] [CrossRef]
- Atabekov, J.; Nikitin, N.; Arkhipenko, M.; Chirkov, S.; Karpova, O. Thermal transition of native tobacco mosaic virus and RNA-free viral proteins into spherical nanoparticles. J. Gen. Virol. 2011, 92, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Balci, S.; Bittner, A.M.; Hahn, K.; Scheu, C.; Knez, M.; Kadri, A.; Wege, C.; Jeske, H.; Kern, K. Copper nanowires within the central channel of tobacco mosaic virus particles. Electrochim. Acta 2006, 51, 6251–6257. [Google Scholar] [CrossRef]
- Balci, S.; Bittner, A.M.; Schirra, M.; Thonke, K.; Sauer, R.; Hahn, K.; Kadri, A.; Wege, C.; Jeske, H.; Kern, K. Catalytic coating of virus particles with zinc oxide. Electrochim. Acta 2009, 54, 5149–5154. [Google Scholar] [CrossRef]
- Miller, R.A.; Presley, A.D.; Francis, M.B. Self-assembling light-harvesting systems from synthetically modified tobacco mosaic virus coat proteins. J. Am. Chem. Soc. 2007, 129, 3104–3109. [Google Scholar] [CrossRef]
- Endo, M.; Fujitsuka, M.; Majima, T. Porphyrin light-harvesting arrays constructed in the recombinant tobacco mosaic virus scaffold. Chem. A Eur. J. 2007, 13, 8660–8666. [Google Scholar] [CrossRef]
- Lin, T.; Chen, Z.; Usha, R.; Stauffacher, C.V.; Dai, J.B.; Schmidt, T.; Johnson, J.E. The refined crystal structure of cowpea mosaic virus at 2.8 A resolution. Virology 1999, 265, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Shah, S.N.; Barclay, J.E.; Rallapalli, G.; Lomonossoff, G.P.; Evans, D.J. Virus-templated silica nanoparticles. Small 2009, 5, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Spall, V.E.; Findlay, K.C.; Gergerich, R.C.; Farrance, C.E.; Lomonossoff, G.P. Cowpea mosaic virus-based chimaeras. Effects of inserted peptides on the phenotype, host range, and transmissibility of the modified viruses. Virology 2003, 310, 50–63. [Google Scholar] [CrossRef]
- Lewis, J.D.; Destito, G.; Zijlstra, A.; Gonzalez, M.J.; Quigley, J.P.; Manchester, M.; Stuhlmann, H. Viral nanoparticles as tools for intravital vascular imaging. Nat. Med. 2006, 12, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Brunel, F.M.; Lewis, J.D.; Destito, G.; Steinmetz, N.F.; Manchester, M.; Stuhlmann, H.; Dawson, P.E. Hydrazone ligation strategy to assemble multifunctional viral nanoparticles for cell imaging and tumor targeting. Nano Lett. 2010, 10, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Langeveld, J.P.; Brennan, F.R.; Martinez-Torrecuadrada, J.L.; Jones, T.D.; Boshuizen, R.S.; Vela, C.; Casal, J.I.; Kamstrup, S.; Dalsgaard, K.; Meloen, R.H.; et al. Inactivated recombinant plant virus protects dogs from a lethal challenge with canine parvovirus. Vaccine 2001, 19, 3661–3670. [Google Scholar] [CrossRef]
- Huang, Y.; Chiang, C.Y.; Lee, S.K.; Gao, Y.; Hu, E.L.; De Yoreo, J.; Belcher, A.M. Programmable assembly of nanoarchitectures using genetically engineered viruses. Nano Lett. 2005, 5, 1429–1434. [Google Scholar] [CrossRef]
- Smith, G.P.; Petrenko, V.A. Phage Display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef]
- Wu, C.H.; Liu, I.J.; Lu, R.M.; Wu, H.C. Advancement and applications of peptide phage display technology in biomedical science. J. Biomed. Sci. 2016, 23, 1–14. [Google Scholar] [CrossRef]
- Rami, A.; Behdani, M.; Yardehnavi, N.; Habibi-Anbouhi, M.; Kazemi-Lomedasht, F. An overview on application of phage display technique in immunological studies. Asian Pac. J. Trop. Biomed. 2017, 7, 599–602. [Google Scholar] [CrossRef]
- Ledsgaard, L.; Kilstrup, M.; Karatt-Vellatt, A.; McCafferty, J.; Laustsen, A.H. Basics of antibody phage display technology. Toxins 2018, 10, 236. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.A.; Arbabi-Ghahroudi, M.; Scott, J.K. Beyond phage display: Non-traditional applications of the filamentous bacteriophage as a vaccine carrier, therapeutic biologic, and bioconjugation scaffold. Front. Microbiol. 2015, 6, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Solis, D.J.; Reiss, B.D.; Kottmann, S.T.; Sweeney, R.Y.; Hayhurst, A.; Georgiou, G.; Iverson, B.; Belcher, A.M. Virus-based toolkit for the directed synthesis of magnetic and semiconducting nanowires. Science 2004, 303, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, M.; Tamerler, C.; Jen, A.K.; Schulten, K.; Baneyx, F. Molecular biomimetics: Nanotechnology through biology. Nat. Mater. 2003, 2, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.T.; Kim, D.W.; Yoo, P.J.; Chiang, C.Y.; Meethong, N.; Hammond, P.T.; Chiang, Y.M.; Belcher, A.M. Virus-enabled synthesis and assembly of nanowires for lithium ion battery electrodes. Science 2006, 312, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Li, H.; Kim, W.-J.; Kang, K.; Yun, D.S.; Strano, M.S.; Ceder, G.; Belcher, A.M. Fabricating genetically engineered high-power lithium-ion batteries using multiple virus genes. Science 2009, 324, 1051–1055. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, K.; Chen, Y.; Li, S.; Nguyen, H.G.; Niu, Z.; You, S.; Mello, C.M.; Lu, X.; Wang, Q. Chemical modification of M13 bacteriophage and its application in cancer cell imaging. Bioconjug. Chem. 2010, 21, 1369–1377. [Google Scholar] [CrossRef]
- Frenkel, D.; Solomon, B. Filamentous phage as vector-mediated antibody delivery to the brain. Proc. Natl. Acad. Sci. USA 2002, 99, 5675–5679. [Google Scholar] [CrossRef]
- Dickerson, T.J.; Kaufmann, G.F.; Janda, K.D. Bacteriophage-mediated protein delivery into the central nervous system and its application in immunopharmacotherapy. Expert Opin. Biol. Ther. 2005, 5, 773–781. [Google Scholar] [CrossRef]
- Carrera, M.R.; Kaufmann, G.F.; Mee, J.M.; Meijler, M.M.; Koob, G.F.; Janda, K.D. Treating cocaine addiction with viruses. Proc. Natl. Acad. Sci. USA 2004, 101, 10416–10421. [Google Scholar] [CrossRef]
- Chen, X.P.; Antson, A.A.; Li, M.Y.P.; Baumann, C.; Dodson, E.J.; Dodson, G.G.; Gollnick, P. Regulatory features of the trp operon and the crystal structure of the trp RNA-binding attenuation protein from Bacillus stearothermophilus. J. Mol. Biol. 1999, 291, 727. [Google Scholar] [CrossRef]
- Bobik, T.A.; Havemann, G.D.; Busch, R.J.; Williams, D.S.; Aldrich, H.C. The propanediol utilization (pdu) operon of Salmonella enterica serovar Typhimurium LT2 includes genes necessary for formation of polyhedral organelles involved in coenzyme B12-dependent 1,2-propanediol degradation. J. Bacteriol. 1999, 181, 5967–5975. [Google Scholar] [CrossRef] [PubMed]
- Yeates, T.O.; Thompson, M.C.; Bobik, T.A. The protein shells of bacterial microcompartment organelles. Curr. Opin. Struct. Biol. 2011, 21, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Walter, D.; Ailion, M.; Roth, J. Genetic characterization of the pdu operon : Use of 1, 2-propanediol in Salmonella typhimurium. J. Bacteriol. 1997, 179, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Crowley, C.S.; Cascio, D.; Sawaya, M.R.; Kopstein, J.S.; Bobik, T.A.; Yeates, T.O. Structural insight into the mechanisms of transport across the Salmonella enterica Pdu microcompartment shell. J. Biol. Chem. 2010, 285, 37838–37846. [Google Scholar] [CrossRef] [PubMed]
- Dgany, O.; Gonzalez, A.; Sofer, O.; Wang, W.; Zolotnitsky, G.; Wolf, A.; Shoham, Y.; Altman, A.; Wolf, S.G.; Shoseyov, O.; et al. The structural basis of the thermostability of SP1, a novel plant (Populus tremula) boiling stable protein. J. Biol. Chem. 2004, 279, 51516–51523. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Han, J.; Zhang, H.; Zhao, L.; Si, C.; Zhang, X.; Hou, C.; Luo, Q.; Xu, J.; Liu, J. Quantum-dot-induced self-assembly of cricoid protein for light harvesting. ACS Nano. 2014, 8, 3743–3751. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Zhang, X.; Si, C.; Gao, Y.; Zhao, L.; Hou, C.; Shoseyov, O.; Luo, Q.; Liu, J. Construction of a highly stable artificial glutathione peroxidase on a protein nanoring. Org. Biomol. Chem. 2014, 12, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Fan, Q.; Zhao, L.; Qiao, Q.; Zhang, X.; Hou, C.; Xu, J.; Luo, Q.; Liu, J. The construction of functional protein nanotubes by small molecule-induced self-assembly of cricoid proteins. Chem. Commun. 2016, 52, 4092–4095. [Google Scholar] [CrossRef]
- Hughes, A.L. Circumsporozoite protein genes of malaria parasites (Plasmodium spp.): Evidence for positive selection on immunogenic regions. Genetics 1991, 127, 345–353. [Google Scholar]
- Guo, Q.; Dasgupta, D.; Doll, T.A.; Burkhard, P.; Lanar, D.E. Expression, purification and refolding of a self-assembling protein nanoparticle (SAPN) malaria vaccine. Methods 2013, 60, 242–247. [Google Scholar] [CrossRef]
- Pfeifer, F. Distribution, formation and regulation of gas vesicles. Nat. Rev. Microbiol. 2012, 10, 705–715. [Google Scholar] [CrossRef]
- Shapiro, M.G.; Goodwill, P.W.; Neogy, A.; Yin, M.; Foster, F.S.; Schaffer, D.V.; Conolly, S.M. Biogenic gas nanostructures as ultrasonic molecular reporters. Nat. Nanotechnol. 2014, 9, 311–316. [Google Scholar] [CrossRef]
- Lakshmanan, A.; Farhadi, A.; Nety, S.P.; Lee-Gosselin, A.; Bourdeau, R.W.; Maresca, D.; Shapiro, M.G. Molecular Engineering of Acoustic Protein Nanostructures. ACS Nano 2016, 10, 7314–7322. [Google Scholar] [CrossRef]
- Valenzuela, K.; Raghavan, S.; Deymier, P.A.; Hoying, J. Formation of copper nanowires by electroless deposition using microtubules as templates. J. Nanosci. Nanotechnol. 2008, 8, 3416–3421. [Google Scholar] [CrossRef][Green Version]
- Paparcone, R.; Cranford, S.W.; Buehler, M.J. Self-folding and aggregation of amyloid nanofibrils. Nanoscale 2011, 3, 1748. [Google Scholar] [CrossRef]
- Scheibel, T.; Parthasarathy, R.; Sawicki, G.; Lin, X.M.; Jaeger, H.; Lindquist, S.L. Conducting nanowires built by controlled self-assembly of amyloid fibers and selective metal deposition. Proc. Natl. Acad. Sci. USA 2003, 100, 4527–4532. [Google Scholar] [CrossRef]
- Kurland, N.E.; Kundu, J.; Pal, S.; Kundu, S.C.; Yadavalli, V.K. Self-assembly mechanisms of silk protein nanostructures on two-dimensional surfaces. Soft Matter. 2012, 8, 4952–4959. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, T.; Ban, H.; Liu, L. Hydrogen bonding-assisted thermal conduction in β-sheet crystals of spider silk protein. Nanoscale 2014, 6, 7786–7791. [Google Scholar] [CrossRef]
- Joseph, B.; Raj, S.J. Therapeutic applications and properties of silk proteins from Bombyx mori. Front. Life Sci. 2012, 6, 55–60. [Google Scholar] [CrossRef]
- Spiess, K.; Lammel, A.; Scheibel, T. Recombinant spider silk proteins for applications in biomaterials. Macromol. Biosci. 2010, 10, 998–1007. [Google Scholar] [CrossRef]
- Miyairi, S.; Sugiura, M. Properties of P-Glucosidase Immobilized in Sericin Membrane. J. Ferment. Technol. 1978, 56, 303–308. [Google Scholar]
- Zhang, Y. Application of natural silk sericin protein in biomaterials. Biotechnol. Adv. 2002, 20, 91–100. [Google Scholar] [CrossRef]
- Min, B.; Lee, G.; Hyun, S.; Sik, Y.; Seung, T.; Ho, W. Electrospinning of silk fibroin nanofibers and its effect on the adhesion and spreading of normal human keratinocytes and fibroblasts in vitro. Biomaterials 2004, 25, 1289–1297. [Google Scholar] [CrossRef]
- Roloff, F.; Strauß, S.; Vogt, P.M.; Bicker, G.; Radtke, C. Spider Silk as Guiding Biomaterial for Human Model Neurons. Biomed Res. Int. 2014, 2014, 906819. [Google Scholar] [CrossRef]
- Steins, A.; Dik, P.; Müller, W.H.; Vervoort, S.J.; Reimers, K. In Vitro Evaluation of Spider Silk Meshes as a Potential Biomaterial for Bladder Reconstruction. PLoS ONE 2015, 10, e0145240. [Google Scholar] [CrossRef]
- Huang, W.; Rollett, A.; Kaplan, D.L. Silk-elastin-like protein biomaterials for the controlled delivery of therapeutics. Expert Opin. Drug Deliv. 2015, 12, 779–791. [Google Scholar] [CrossRef]
- Xia, X.X.; Xu, Q.; Hu, X.; Qin, G.; Kaplan, D.L. Tunable self-assembly of genetically engineered silk-elastin-like protein polymers. Biomacromolecules 2011, 12, 3844–3850. [Google Scholar] [CrossRef]
- Xia, X.X.; Wang, M.; Lin, Y.; Xu, Q.; Kaplan, D.L. Hydrophobic drug-triggered self-assembly of nanoparticles from silk-elastin-like protein polymers for drug delivery. Biomacromolecules 2014, 15, 908–914. [Google Scholar] [CrossRef]
- Seabra, A.B.; Durán, N. Biological applications of peptides nanotubes: An overview. Peptides 2013, 39, 47–54. [Google Scholar] [CrossRef]
- Žganec, M.; Žerovnik, E. Amyloid fibrils compared to peptide nanotubes. Biochim. Biophys. Acta. Gen. Subj. 2014, 1840, 2944–2952. [Google Scholar] [CrossRef]
- Scanlon, S.; Aggeli, A. Self-assembling peptide nanotubes. Nano Today 2008, 3, 22–30. [Google Scholar] [CrossRef]
- Gao, X.; Matsui, H. Peptide-Based Nanotubes and Their Applications in Bionanotechnology. Adv. Mater. 2005, 17, 2037–2050. [Google Scholar] [CrossRef]
- Hsieh, W.H.; Liaw, J. Applications of cyclic peptide nanotubes (cPNTs). J. Food Drug Anal. 2019, 27, 32–47. [Google Scholar] [CrossRef]
- Brea, R.J.; Reiriz, C.; Granja, J.R. Towards functional bionanomaterials based on self-assembling cyclic peptide nanotubes. Chem. Soc. Rev. 2010, 39, 1448–1456. [Google Scholar] [CrossRef]
- Chen, K.H.; Corro, K.A.; Le, S.P.; Nowick, J.S. X-ray Crystallographic Structure of a Giant Double-Walled Peptide Nanotube Formed by a Macrocyclic β-Sheet Containing Aβ16-22. J. Am. Chem. Soc. 2017, 139, 8102–8105. [Google Scholar] [CrossRef]
- Swain, M.; Thirupathi, R.; Krishnarjuna, B.; Eaton, E.M.; Kibbey, M.M.; Rosenzweig, S.A.; Atreya, H.S. Spontaneous and reversible self-assembly of a polypeptide fragment of insulin-like growth factor binding protein-2 into fluorescent nanotubular structures. Chemic 2010, 46, 216–218. [Google Scholar] [CrossRef][Green Version]
- Asampille, G.; Verma, B.K.; Swain, M.; Shettar, A.; Rosenzweig, S.A.; Kondaiah, P.; Atreya, H.S. An ultra-stable redox-controlled self-assembling polypeptide nanotube for targeted imaging and therapy in cancer. J. Nanobiotechnol. 2018, 16, 1–15. [Google Scholar] [CrossRef]
- Kuang, Z.; Yao, S.; Keizer, D.W.; Wang, C.C.; Bach, L.A.; Forbes, B.E.; Wallace, J.C.; Norton, R.S. Structure, Dynamics and Heparin Binding of the C-terminal Domain of Insulin-like Growth Factor-binding Protein-2 (IGFBP-2). J. Mol. Biol. 2006, 2, 690–704. [Google Scholar] [CrossRef]
- Bellis, S.L. Biomaterials Advantages of RGD peptides for directing cell association with biomaterials. Biomaterials 2011, 32, 4205–4210. [Google Scholar] [CrossRef]
- Kibbey, M.M.; Jameson, M.J.; Eaton, E.M.; Rosenzweig, S.A. Insulin-Like Growth Factor Binding Protein-2: Contributions of the C-Terminal Domain to Insulin-Like Growth Factor-1 Binding. Mol. Pharmacol. 2006, 69, 833–845. [Google Scholar] [CrossRef]
- Ngweniform, P.; Li, D.; Mao, C. Self-assembly of drug-loaded liposomes on genetically engineered protein nanotubes: A potential anti-cancer drug delivery vector. Soft Matter. 2009, 5, 954–956. [Google Scholar] [CrossRef]
- Fan, Y.W.; Sun, Z.; Wang, R.; Abbott, C.; Moradian-Oldak, J. Enamel inspired nanocomposite fabrication through amelogenin supramolecular assembly. Biomaterials 2007, 28, 3034–3042. [Google Scholar] [CrossRef]
- Fincham, A.G.; Moradian-Oldak, J.; Simmer, J.P. The Structural Biology of the Developing Dental Enamel Matrix. J. Struct. Biol. 1999, 126, 270–299. [Google Scholar] [CrossRef]
- Wang, L.; Guan, X.; Yin, H.; Moradian-Oldak, J.; Nancollas, G.H. Mimicking the self-organized microstructure of tooth enamel. J. Phys. Chem. C 2008, 112, 5892–5899. [Google Scholar] [CrossRef]
- Graveland-Bikker, J.F.; de Kruif, C.G. Unique milk protein based nanotubes: Food and nanotechnology meet. Trends Food Sci. Technol. 2006, 17, 196–203. [Google Scholar] [CrossRef]
- Graveland-Bikker, J.F.; Schaap, I.A.T.; Schmidt, C.F.; De Kruif, C.G. Structural and mechanical study of a self-assembling protein nanotube. Nano Lett. 2006, 6, 616–621. [Google Scholar] [CrossRef]
- Tarhan, Ö.; Tarhan, E.; Harsa, Ş. Investigation of the structure of alpha-lactalbumin protein nanotubes using optical spectroscopy. J. Dairy Res. 2014, 81, 98–106. [Google Scholar] [CrossRef]
- Hu, B.; Huang, Q.R. Biopolymer based nano-delivery systems for enhancing bioavailability of nutraceuticals. Chin. J. Polym. Sci. 2013, 31, 1190–1203. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PNT Origin | Application | Reference |
|---|---|---|
| Bacterial | ||
| Flagella (FliC) | Anionic FliC PNTs coordinated with ZnPc-loaded cationic liposomes are used for targeted drug delivery of cancer cells | [132] |
| Gas vesicles | GvpC is engineered to alter surface properties, the display of ligands, and the attachment of fluorescent proteins to enable enhanced ultrasound imaging | [104] |
| Viral | ||
| Cowpea mosaic virus (CPMV) capsid | Labeled with fluorescent dye for non-immunogenic intravital visualization of vasculature | [74] |
| Labeled with a peptide and a fluorophore for tumor cell detection | [75] | |
| Attachment of viral antigens as a vaccine delivery vehicle | [76] | |
| M13 Bacteriophage | Folic acid and fluorophore attachment allows visualization of cancer cells overexpressing the folate receptor | [87] |
| Detection of Alzheimer’s through fusion of M13 minor coat protein pIII with I123 radiolabeled anti-β-amyloid antibody | [88,89] | |
| Attachment of GNC 92H2 murine monoclonal antibody to the pVIII coat protein for treatment of cocaine addictions | [90] | |
| Eukaryotic | ||
| Self-Assembled Protein Nanoparticle (SAPN) | Self-assembled spherical nanoparticles presenting P. falciparum antigenic epitopes for use as a malarial vaccine | [101] |
| α-lactalbumin | PNTs with utility as viscosifiers, gelation agents for food and encapsulating vitamins or enzymes for drug delivery | [136] |
| Amelogenin | Amelogenin nanorods facilitate enamel growth in dental repair | [135] |
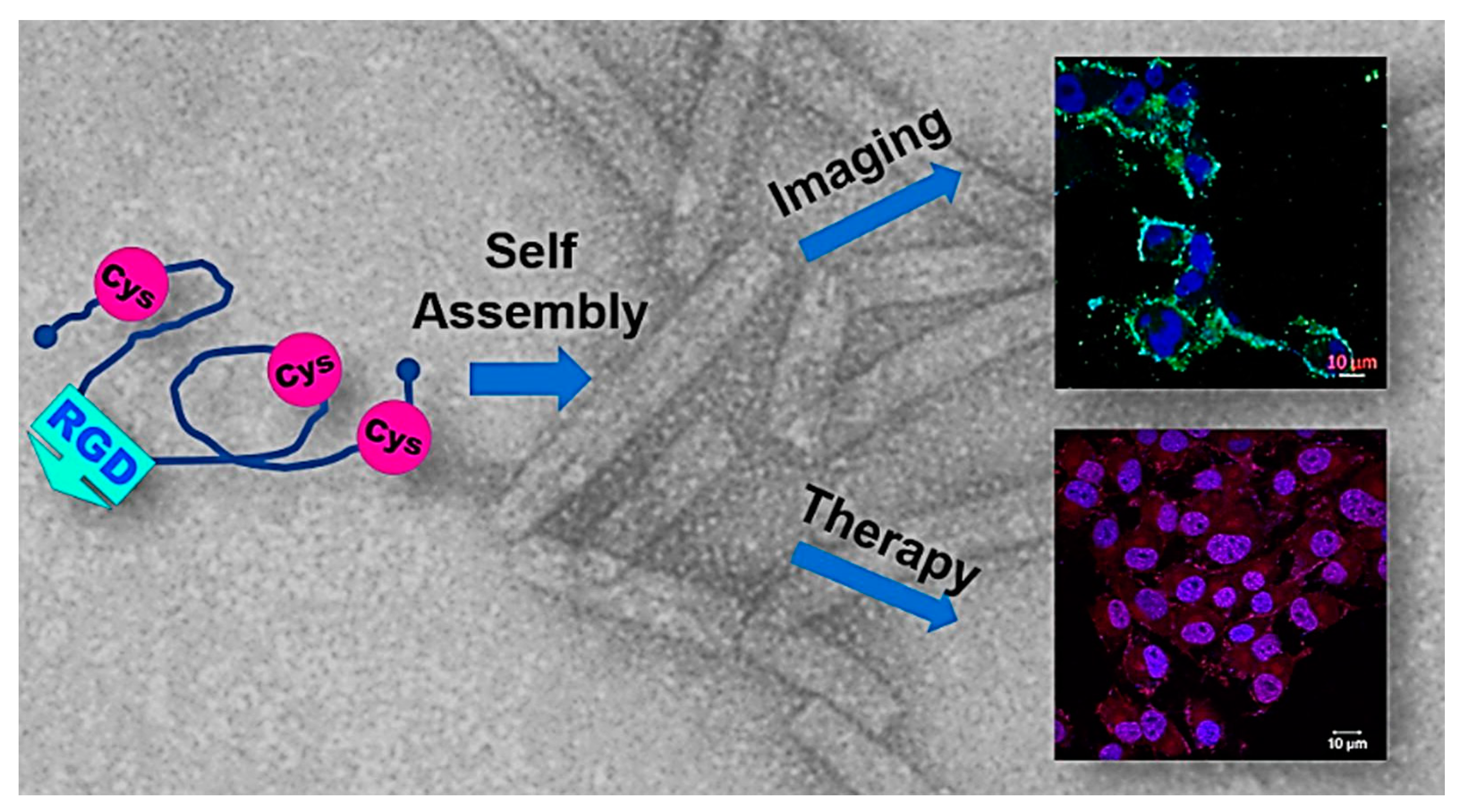
| Human insulin-like growth factor binding protein-2 (hIGFPB-2) | Cell targeting with DOX and imaging agents | [127,128] |
| Silk Proteins | B. mori silk processed to immobilize enzymes for the production of a biocatalytic surface | [109] |
| Adaptable ADF3 and ADF4 fibroin forms morphologies suitable as drug delivery vehicles, and for cell adhesion during tissue engineering | [111] | |
| Nephila spider silk is used as a biocompatible scaffold, shown to guide axonal regeneration and allow tissue development for the reconstruction of a human bladder | [115,116] | |
| Silk-elastin-like protein polymers tuned to form a nanoparticle capable of targeting cancer cells with doxorubicin | [119] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Audette, G.F.; Yaseen, A.; Bragagnolo, N.; Bawa, R. Protein Nanotubes: From Bionanotech towards Medical Applications. Biomedicines 2019, 7, 46. https://doi.org/10.3390/biomedicines7020046
Audette GF, Yaseen A, Bragagnolo N, Bawa R. Protein Nanotubes: From Bionanotech towards Medical Applications. Biomedicines. 2019; 7(2):46. https://doi.org/10.3390/biomedicines7020046
Chicago/Turabian StyleAudette, Gerald F., Ayat Yaseen, Nicholas Bragagnolo, and Raj Bawa. 2019. "Protein Nanotubes: From Bionanotech towards Medical Applications" Biomedicines 7, no. 2: 46. https://doi.org/10.3390/biomedicines7020046
APA StyleAudette, G. F., Yaseen, A., Bragagnolo, N., & Bawa, R. (2019). Protein Nanotubes: From Bionanotech towards Medical Applications. Biomedicines, 7(2), 46. https://doi.org/10.3390/biomedicines7020046