The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation

 ,
,  ,
,  and
and
Abstract
:1. Introduction
1.1. Relationship between the Brain and the Gut Microbiota
1.2. Blood Barriers and Their Roles in the MGB Axis
1.2.1. Intestinal Blood Barrier
1.2.2. Blood–Brain Barrier
1.3. MGB Axis: A New and Important Drug Target
1.4. The Mycobiota and Microbiota across the Lifespan
1.5. Autism and ASD in Relation to the Microbiota and Child Development
1.6. Consequences to Perturbation in the Gut Microbiota
1.7. Aging and the Microbiota through the Lifespan
1.8. The Promise of Fecal Transfer from Young Donors to Old-Aged Recipients
1.9. Methionine Cycle Genes Contribute to Epigenetic Regulation
1.10. CRISPR and Gene Engineering Prospects
1.11. Novel New Antibiotics and Synbiotics to Treat Diseases and Age-Related Syndromes
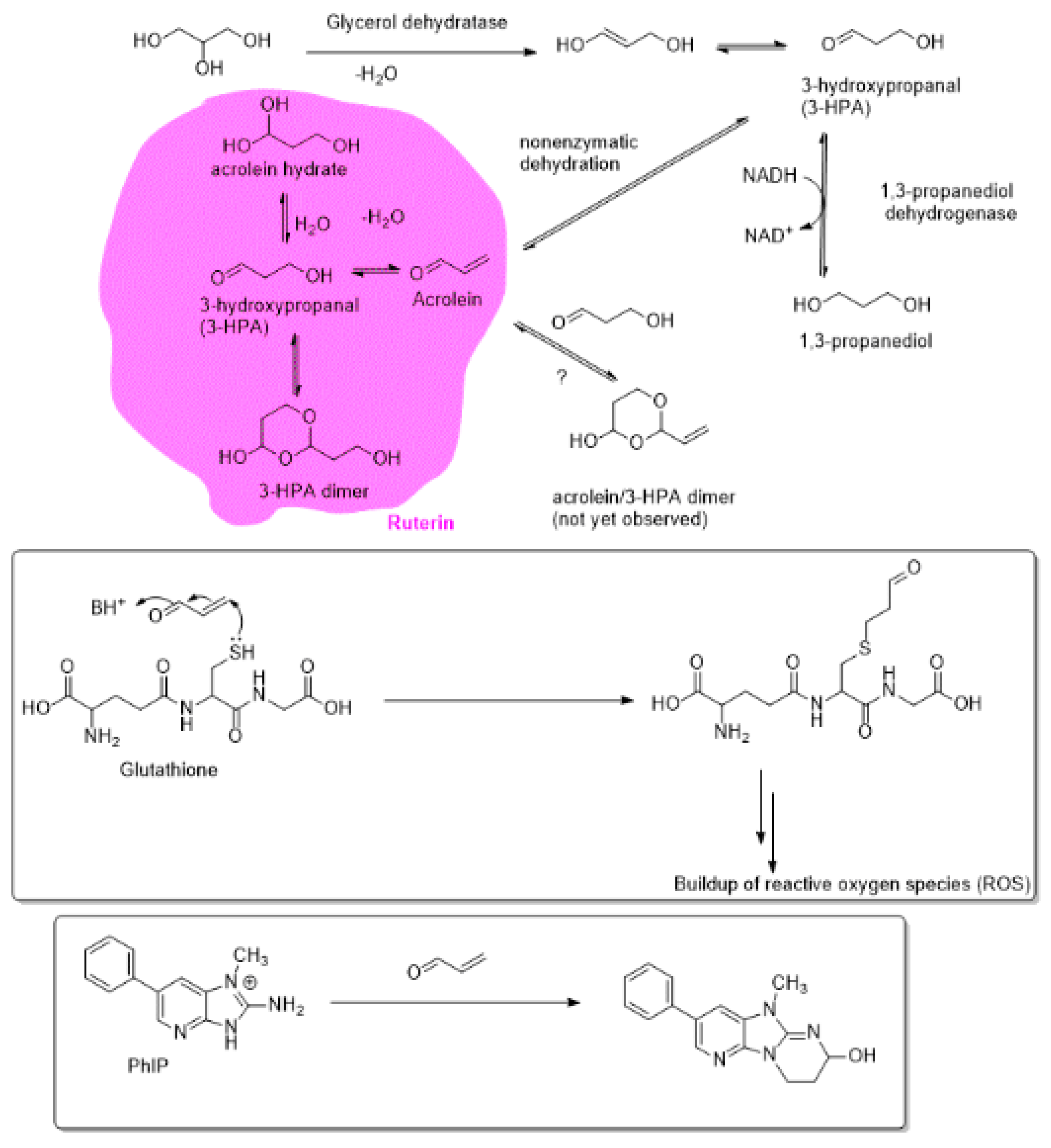
1.12. Ruterin as a Natural Source of Antibacterial Agent.
2. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
| Category | Fungal Species | Significance |
|---|---|---|
| Infant GI/Skin Mother to infant | Candida (breast (breastmilk), skin, oral, vaginal/birth and GI) Malassezia M. restricta and M. globosa, (Skin) Saccharomyces boulardii (GI/Skin) Davidiella (vaginal) Rhodotorula (skin) Pichia jadinii (Cyberlindnera) Cladosporium cladosporiodes (vaginal) Basidiomycota (Cryptococcus) Penicillium (skin) | Asthma Allergy Atopy IBD/IBS Diarrhea Emerging pathogens |
| Fecal early life | C. parapsilosis, C. tropicalis, C. albicans, C. orthopsilosis (birth mode, oral, environment), C. quercitrusa, C. diddensiae, C. krusei, C. guilliermondii, C. geocandidum Saccharomyces cerevisiae (infant GI) Saccharomycetales (overgrowth) Cryptococcus pseudolongus (C. albidosimilas and C. podzolicus) Cladosporium velox (C. sphaerospermum and C. tenuissimum) (body site) Debaryomyces renaii (body site) D. hansenii(body site) Hanseniaspora uvarum (body site) Issatchenkia orientalis (Candida krusei) Nicotiana (infant GI) Cicer (infant GI) Leptosphaerulina Aspergillus Debaryomyces Malassezia Ascomycota Eurotiomycetes Tremellomycetes Nectriaceae Trichosporon | Infant and maternal mycobiome Possible opportunistic pathogens colonization |
| Vaginal mycobiome | C. albicans S. cerevisiae C. parapsilosis | Infection Colonization |
| Infant skin | Malassezia | Colonizer/pathogenic |
| Infant oral mycobiome | Candida species such as C. albicans, C. parapsilosis, C. krusei, C. guilliermondii, C. geocandidum, C. tropicalis and C. orthopsilosis S. cerevisiae | Mucocutaneous infection Overgrowth |
| Adult fecal samples | Saccharomyces Candida Cladosporium Mucor Penicillium Wallemia Bettsia | GI Allergy Respiratory |
| Pathogenic fungi | Saccharomyces, Candida, Aspergillus, Malassezia, Penicillium | Allergy Disseminated infection |
| Adult GI mycoboiome | Candida Penicillium Aspergillus Malassezia Debaryomyces Mucor Eremothecium Pichia Cyberlindnera Saccharomyces Penicillium | Weight management/obesity |
| Adult skin mycobiome | Malassezia Candida Rhodotorula Saccharomyces Penicillium | Colonizers Pathogenic Allergy |
| Adult oral cavity | Candida Cladosporium Aspergillus Fusarium Glomus Penicillium Alternaria Cryptococcus Ophiosoma Phoma Schizosaccharomyces Zygosaccharomyces Saccharomyces | Colonizers Pathogenic Allergy |
| Age Range | Most Prevalent Phyla | Common Genera | |
|---|---|---|---|
| Infancy | 0–3 years | Actinobacteria | Propionibacterium, Rothia, Actinomyces, Bifidobacterium, Collinsella, Eggerthella, Mobiluncus |
| Proteobacteria | Escherichia, Shigella, Aggregatibacter, Eikenella | ||
| Firmicutes | Enterococcus, Gemella, Staphylococcus, Streptococcus, Anaerococcus, Granulicatella, Lactobacillus, Peptoniphilus, Veillonella, Anaerostipes, Anaerotruncus, Costridium, Megasphaera, Roseburia, Ruminococcus | ||
| Bacteroidetes | Bacteroides | ||
| Fusobacteria | Fusobacterium | ||
| Adult | 3–70 years | Firmicutes | Enterococcus, Gemella, Staphylococcus, Streptococcus, Anaerococcus, Granulicatella, Lactobacillus, Peptoniphilus, Veillonella, Anaerostipes, Anaerotruncus, Costridium, Megasphaera, Roseburia, Ruminococcus |
| Bacteroidetes | Bacteroides | ||
| Actinobacteria | Propionibacterium, Rothia, Actinomyces, Bifidobacterium, Collinsella, Eggerthella, Mobiluncus | ||
| Proteobacteria | Escherichia, Shigella, Aggregatibacter, Eikenella | ||
| Verrucomicrobia | Akkkermansia | ||
| Elderly | 70 years and older | Firmicutes | Enterococcus, Gemella, Staphylococcus, Streptococcus, Anaerococcus, Granulicatella, Lactobacillus, Peptoniphilus, Veillonella, Anaerostipes, Anaerotruncus, Costridium, Megasphaera, Roseburia, Ruminococcus |
| Actinobacteria | Propionibacterium, Rothia, Actinomyces, Bifidobacterium, Collinsella, Eggerthella, Mobiluncus | ||
| Bacteroidetes | Bacteroides | ||
| Proteobacteria | Escherichia, Shigella, Aggregatibacter, Eikenella | ||
| Verrucomicrobia | Akkermansia |
References
- Obrenovich, M. Leaky Gut, Leaky Brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef]
- Obrenovich, M.; Rai, H.; Mana, T.S.; Shola, D.; McCloskey, B. Dietary Co-Metabolism within the Microbiota-Gut-Brain-Endocrine Metabolic Interactome. BAO Microbiol. 2017, 2, 22. [Google Scholar]
- Ward, T.L.; Knights, D.; Gale, C.A. Infant fungal communities: Current knowledge and research opportunities. BMC Med. 2017, 15, 30. [Google Scholar] [CrossRef]
- Ward, T.L.; Dominguez-Bello, M.G.; Heisel, T.; Al-Ghalith, G.; Knights, D.; Gale, C.A. Development of the Human Mycobiome over the First Month of Life and across Body Sites. MSystems 2018, 3, e00140-17. [Google Scholar] [CrossRef]
- Gareau, M.G. Microbiota-gut-brain axis and cognitive function. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Mark, L., John, C., Eds.; Springer: New York, NY, USA, 2014; pp. 357–371. [Google Scholar]
- Obrenovich, M.; Flückiger, R.; Sykes, L.; Donskey, C. The co-metabolism within the gut-brain metabolic interaction: Potential targets for drug treatment and design. CNS Neurol. Disord. Drug Targets 2016, 15, 127–134. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Rashel, M.; Uchiyama, J.; Sakurai, S.; Ujihara, T.; Kuroda, M.; Ikeuchi, M.; Tani, T.; Fujieda, M.; Wakiguchi, H.; et al. Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Pini, E.; Nikkïla, J.; Monti, D.; Satokari, R.; Franceschi, C.; et al. Through ageing, and beyond: Gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 2010, 5, e10667. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Blaser, M.J.; Ley, R.E.; Knight, R. Development of the human gastrointestinal microbiota and insights from high-throughput sequencing. Gastroenterology 2011, 140, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B. Molecular Biology of the Cell, 6th ed.; Garland Science, Taylor and Francis Group: New York, NY, USA, 2015; ISBN 978-0-8153-4432-2. [Google Scholar]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood–brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Paharik, A.E.; Schreiber, H.L.; Spaulding, C.N.; Dodson, K.W.; Hultgren, S.J. Narrowing the spectrum: The new frontier of precision antimicrobials. Genome Med. 2017, 9, 110. [Google Scholar] [CrossRef]
- Braundmeier, A.G.; Lenz, K.M.; Inman, K.S.; Chia, N.; Jeraldo, P.; Walther-António, M.R.; Berg Miller, M.E.; Yang, F.; Creedon, D.J.; Nelson, H.; et al. Individualized medicine and the microbiome in reproductive tract. Front. Physiol. 2015, 6, 97. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Bernstein, C.N.; Tremlett, H.; Van Domselaar, G.; Knox, N.C. A Fungal World: Could the Gut Mycobiome Be Involved in Neurological Disease? Front. Microbiol. 2019, 9, 3249. [Google Scholar] [CrossRef]
- Sogin, M.L.; Morrison, H.G.; Huber, J.A.; Welch, D.M.; Huse, S.M.; Neal, P.R.; Arrieta, J.M.; Herndl, G.J. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. USA 2006, 103, 12115–12120. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Hogan, D.A.; Mylonakis, E. Medically important bacterial–fungal interactions. Nat. Rev. Microbiol. 2010, 8, 340. [Google Scholar] [CrossRef]
- Drell, T.; Lillsaar, T.; Tummeleht, L.; Simm, J.; Aaspõllu, A.; Väin, E.; Saarma, I.; Salumets, A.; Donders, G.G.; Metsis, M. Characterization of the vaginal micro-and mycobiome in asymptomatic reproductive-age Estonian women. PLoS ONE 2013, 8, e54379. [Google Scholar] [CrossRef]
- Rozovsky, I.; Finch, C.E.; Morgan, T.E. Age-related activation of microglia and astrocytes: In vitro studies show persistent phenotypes of aging, increased proliferation, and resistance to down-regulation. Neurobiol. Aging 1998, 19, 97–103. [Google Scholar] [CrossRef]
- Wu, Y.; Du, S.; Johnson, J.L.; Tung, H.Y.; Landers, C.T.; Liu, Y.; Seman, B.G.; Wheeler, R.T.; Costa-Mattioli, M.; Kheradmand, F.; et al. Microglia and amyloid precursor protein coordinate control of transient Candida cerebritis with memory deficits. Nat. Commun. 2019, 10, 58. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef]
- Charlson, E.S.; Diamond, J.M.; Bittinger, K.; Fitzgerald, A.S.; Yadav, A.; Haas, A.R.; Bushman, F.D.; Collman, R.G. Lung-enriched organisms and aberrant bacterial and fungal respiratory microbiota after lung transplant. Am. J. Respir. Crit. Care Med. 2012, 186, 536–545. [Google Scholar] [CrossRef]
- Seed, P.C. The human mycobiome. Cold Spring Harb. Perspect. Med. 2015, 5, a019810. [Google Scholar] [CrossRef]
- Bradford, L.L.; Ravel, J. The vaginal mycobiome: A contemporary perspective on fungi in women’s health and diseases. Virulence 2016, 8, 342–351. [Google Scholar] [CrossRef]
- Sam, Q.H.; Chang, M.W.; Chai, L.Y. The Fungal Mycobiome and Its Interaction with Gut Bacteria in the Host. Int. J. Mol. Sci. 2017, 18, 330. [Google Scholar] [CrossRef]
- Huffnagle, G.B.; Noverr, M.C. The emerging world of the fungal microbiome. Trends Microbiol. 2013, 21, 334–341. [Google Scholar] [CrossRef]
- Butel, M.J.; Waligora-Dupriet, A.J.; Wydau-Dematteis, S. The developing gut microbiota and its consequences for health. J. Dev. Orig. Health Dis. 2018, 9, 590–597. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef]
- Thum, C.; Cookson, A.L.; Otter, D.E.; McNabb, W.C.; Hodgkinson, A.J.; Dyer, J.; Roy, N.C. Can Nutritional Modulation of Maternal Intestinal Microbiota Influence the Development of the Infant Gastrointestinal Tract? J. Nutr. 2012, 142, 1921–1928. [Google Scholar] [CrossRef]
- Neu, J. Developmental aspects of maternal-fetal, and infant gut microbiota and implications for long-term health. Matern. Health Neonatol. Perinatol. 2015, 1, 6. [Google Scholar] [CrossRef]
- Jeurink, P.V.; Van Bergenhenegouwen, J.; Jimenez, E.; Knippels, L.M.; Fernández, L.; Garssen, J.; Knol, J.; Rodriguez, J.M.; Martin, R. Human milk: A source of more life than we imagine. Benef. Microbes 2012, 4, 17–30. [Google Scholar] [CrossRef]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef]
- Biagi, E.; Candela, M.; Fairweather-Tait, S.; Franceschi, C.; Brigidi, P. Ageing of the human metaorganism: The microbial counterpart. Age 2012, 34, 247–267. [Google Scholar] [CrossRef]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the human infant intestinal microbiota. PLoS Biol. 2007, 5, e177. [Google Scholar] [CrossRef]
- Bale, T.L.; Baram, T.Z.; Brown, A.S.; Goldstein, J.M.; Insel, T.R.; McCarthy, M.M.; Nemeroff, C.B.; Reyes, T.M.; Simerly, R.B.; Susser, E.S.; et al. Early life programming and neurodevelopmental disorders. Biol. Psychiatry 2010, 68, 314–319. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Shola, D.; Schroedel, K.; Agrahari, A.; Lonsdale, D. The role of trace elements, thiamin (e) in autism and autistic spectrum disorder. Front. Biosci. 2015, 7, 263–277. [Google Scholar]
- Mittal, V.A.; Ellman, L.M.; Cannon, T.D. Gene-environment interaction and covariation in schizophrenia: The role of obstetric complications. Schizophr. Bull. 2008, 34, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.L.; Levitt, P.; Stanwood, G.D. Prenatal exposure to drugs: Effects on brain development and implications for policy and education. Nat. Rev. Neurosci. 2009, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y. Neuropaediatric and neuroarchaeology: Understanding development to correct brain disorders. Acta Paediatr. 2013, 102, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M. State of the art; microbiology in health and disease. Intestinal bacterial flora in autism. Anaerobe 2011, 17, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Poschmann, J.; del Rosario, R.C.; Parikshak, N.N.; Hajan, H.S.; Kumar, V.; Ramasamy, R.; Belgard, T.G.; Elanggovan, B.; Wong, C.C.; et al. Histone acetylome-wide association study of autism spectrum disorder. Cell 2016, 167, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Mulle, J.G.; Sharp, W.G.; Cubells, J.F. The gut microbiome: A new frontier in autism research. Curr. Psychiatry Rep. 2013, 15, 337. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, J.L.; Giedd, J.N.; Gogtay, N. Neurodevelopmental model of schizophrenia: Update 2012. Mol. Psychiatry 2012, 17, 1228. [Google Scholar] [CrossRef]
- Shaw, W. Increased urinary excretion of a 3-(3-hydroxyphenyl)-3-hydroxypropionic acid (HPHPA), an abnormal phenylalanine metabolite of Clostridia spp. in the gastrointestinal tract, in urine samples from patients with autism and schizophrenia. Nutr. Neurosci. 2010, 13, 135–143. [Google Scholar] [CrossRef]
- Ming, X.; Stein, T.P.; Barnes, V.; Rhodes, N.; Guo, L. Metabolic perturbance in autism spectrum disorders: A metabolomics study. J. Proteome Res. 2012, 11, 5856–5862. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Donskey, C.J.; Schiefer, I.T.; Bongiovanni, R.; Li, L.; Jaskiw, G.E. Quantification of phenolic acid metabolites in humans by LC–MS: A structural and targeted metabolomics approach. Bioanalysis 2018, 10, 1591–1608. [Google Scholar] [CrossRef]
- De Theije, C.G.; Wopereis, H.; Ramadan, M.; van Eijndthoven, T.; Lambert, J.; Knol, J.; Garssen, J.; Kraneveld, A.D.; Oozeer, R. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav. Immun. 2014, 37, 197–206. [Google Scholar] [CrossRef]
- Douglas-Escobar, M.; Elliott, E.; Neu, J. Effect of intestinal microbial ecology on the developing brain. JAMA Pediatr. 2013, 167, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Desbonnet, L.; Clarke, G.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. Microbiota is essential for social development in the mouse. Mol. Psychiatry 2014, 19, 146. [Google Scholar] [CrossRef] [PubMed]
- Iversen, L. Neurotransmitter transporters: Fruitful targets for CNS drug discovery. Mol. Psychiatry 2000, 5, 357. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Cryan, J.F.; Ross, R.P.; Fitzgerald, G.F.; Dinan, T.G.; Stanton, C. Bacterial neuroactive compounds produced by psychobiotics. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Mark, L., John, C., Eds.; Springer: New York, NY, USA, 2014; pp. 221–239. [Google Scholar]
- Zhou, L.; Foster, J.A. Psychobiotics and the gut–brain axis: In the pursuit of happiness. Neuropsychiatr. Dis. Treat. 2015, 11, 715. [Google Scholar]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559. [Google Scholar] [CrossRef]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108, 4586–4591. [Google Scholar] [CrossRef]
- Ostan, R.; Bucci, L.; Capri, M.; Salvioli, S.; Scurti, M.; Pini, E.; Monti, D.; Franceschi, C. Immunosenescence and immunogenetics of human longevity. Neuroimmunomodulation 2008, 15, 224–240. [Google Scholar] [CrossRef]
- Biagi, E.; Candela, M.; Turroni, S.; Garagnani, P.; Franceschi, C.; Brigidi, P. Ageing and gut microbes: Perspectives for health maintenance and longevity. Pharmacol. Res. 2013, 69, 11–20. [Google Scholar] [CrossRef]
- Carter, C.J. Genetic, Transcriptome, Proteomic, and Epidemiological Evidence for Blood-Brain Barrier Disruption and Polymicrobial Brain Invasion as Determinant Factors in Alzheimer’s Disease. J. Alzheimer’s Dis. Rep. 2017, 1, 125–157. [Google Scholar] [CrossRef]
- Seidel, J.; Valenzano, D.R. The role of the gut microbiome during host ageing. F1000Research 2018, 7. [Google Scholar] [CrossRef]
- Winek, K.; Dirnagl, U.; Meisel, A. The gut microbiome as therapeutic target in central nervous system diseases: Implications for stroke. Neurotherapeutics 2016, 13, 762–774. [Google Scholar] [CrossRef]
- Smith, P.; Willemsen, D.; Popkes, M.; Metge, F.; Gandiwa, E.; Reichard, M.; Valenzano, D.R. Regulation of life span by the gut microbiota in the short-lived African turquoise killifish. Elife 2017, 6, e27014. [Google Scholar] [CrossRef]
- Hecker, M.T.; Obrenovich, M.E.; Cadnum, J.L.; Jencson, A.L.; Jain, A.K.; Ho, E.; Donskey, C.J. Fecal microbiota transplantation by freeze-dried oral capsules for recurrent Clostridium difficile infection. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2016; Volume 3. [Google Scholar]
- Conboy, I.M.; Conboy, M.J.; Wagers, A.J.; Girma, E.R.; Weissman, I.L.; Rando, T.A. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 2005, 433, 760. [Google Scholar] [CrossRef]
- Mccay, C.M.; Pope, F.; Lunsford, W.; Sperling, G.; Sambhavaphol, P. Parabiosis between old and young rats. Gerontology 1957, 1, 7–17. [Google Scholar] [CrossRef]
- Ludwig, F.C.; Elashoff, R.M. Mortality in syngeneic rat parabionts of different chronological age. Trans. N. Y. Acad. Sci. 1972, 34, 582–587. [Google Scholar] [CrossRef]
- Pesheva, E. Critical Step Found in DNA Repair, Cellular Aging, HMS Communications; The Harvard Gazette, 2017. Available online: https://news.harvard.edu/gazette/story/2017/03/harvard-scientists-pinpoint-critical-step-in-dna-repair-cellular-aging/ (accessed on 1 January 2019).
- Boukaba, A.; Sanchis-Gomar, F.; García-Giménez, J.L. Epigenetic Mechanisms as Key Regulators in Disease: Clinical Implications. Epigenetic Biomarkers and Diagnostics; Academic Press: Cambridge, MA, USA, 2016; pp. 37–66. [Google Scholar]
- Hoffmann, G.F.; Kölker, S. Defects of methionine and homocysteine metabolism. In Pediatric Neurology Part III in Handbook of Clinical Neurology; Olivier, D., Maryse, L., Harvey, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1755–1773. [Google Scholar]
- De Magalhães, J.P. The scientific quest for lasting youth: Prospects for curing aging. Rejuvenation Res. 2014, 17, 458–467. [Google Scholar] [CrossRef]
- Weil, A.A.; Hohmann, E.L. Fecal microbiota transplant: Benefits and risks. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2015; Volume 2. [Google Scholar]
- Tsukumo, D.M.; Carvalho, B.M.; Carvalho Filho, M.A.; Saad, M.J. Translational research into gut microbiota: New horizons on obesity treatment: Updated 2014. Arch. Endocrinol. Metab. 2015, 59, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Maccaferri, S.; Turroni, S.; Carnevali, P.; Brigidi, P. Functional intestinal microbiome, new frontiers in prebiotic design. Int. J. Food Microbiol. 2010, 140, 93–101. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Lee, D.; Wong, E.; Dadlani, P.; Dinh, D.; Huang, V.; Kearns, K.; Teng, S.; Chen, S.; Haliburton, J.; et al. Specific gene repression by CRISPRi system transferred through bacterial conjugation. ACS Synth. Biol. 2014, 3, 929–931. [Google Scholar] [CrossRef]
- Zuberi, A.; Misba, L.; Khan, A.U. CRISPR Interference (CRISPRi) inhibition of luxS gene expression in E. coli: An approach to inhibit biofilm. Front. Cell. Infect. Microbiol. 2017, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome–gut–brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol. Ther. 2016, 158, 52–62. [Google Scholar] [CrossRef]
- Wu, S.C.; Cao, Z.S.; Chang, K.M.; Juang, J.L. Intestinal microbial dysbiosis aggravates the progression of Alzheimer’s disease in Drosophila. Nat. Commun. 2017, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Minato, T.; Maeda, T.; Fujisawa, Y.; Tsuji, H.; Nomoto, K.; Ohno, K.; Hirayama, M. Progression of Parkinson’s disease is associated with gut dysbiosis: Two-year follow-up study. PLoS ONE 2017, 12, e0187307. [Google Scholar] [CrossRef] [PubMed]
- Spielman, L.J.; Gibson, D.L.; Klegeris, A. Unhealthy gut, unhealthy brain: The role of the intestinal microbiota in neurodegenerative diseases. Neurochem. Int. 2018, 120, 149–163. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Smith, M.A.; Siedlak, S.L.; Chen, S.G.; Jack, C.; Perry, G.; Aliev, G. Overexpression of GRK2 in Alzheimer disease and in a chronic hypoperfusion rat model is an early marker of brain mitochondrial lesions. Neurotox. Res. 2006, 10, 43–56. [Google Scholar] [CrossRef]
- Huang, Y.; Todd, N.; Thathiah, A. The role of GPCRs in neurodegenerative diseases: Avenues for therapeutic intervention. Curr. Opin. Pharmacol. 2017, 32, 96–110. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yuille, S.; Reichardt, N.; Panda, S.; Dunbar, H.; Mulder, I.E. Human gut bacteria as potent class I histone deacetylase inhibitors in vitro through production of butyric acid and valeric acid. PLoS ONE 2018, 13, e0201073. [Google Scholar] [CrossRef] [PubMed]
- Auluck, P.K.; Chan, H.E.; Trojanowski, J.Q.; Lee, V.M.; Bonini, N.M. Chaperone suppression of α-synuclein toxicity in a Drosophila model for Parkinson’s disease. Science 2002, 295, 865–868. [Google Scholar] [CrossRef]
- Jansen, M.S.; Nagel, S.C.; Miranda, P.J.; Lobenhofer, E.K.; Afshari, C.A.; McDonnell, D.P. Short-chain fatty acids enhance nuclear receptor activity through mitogen-activated protein kinase activation and histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2004, 101, 7199–7204. [Google Scholar] [CrossRef]
- Ho, L.; Ono, K.; Tsuji, M.; Mazzola, P.; Singh, R.; Pasinetti, G.M. Protective roles of intestinal microbiota derived short chain fatty acids in Alzheimer’s disease-type beta-amyloid neuropathological mechanisms. Expert Rev. Neurother. 2018, 18, 83–90. [Google Scholar] [CrossRef] [PubMed]
- MacFabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2012, 23, 19260. [Google Scholar] [CrossRef]
- Karaki, S.I.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the short-chain fatty acid receptor, GPR43, in the human colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef]
- Wang, H.; Lee, I.S.; Braun, C.; Enck, P. Effect of probiotics on central nervous system functions in animals and humans: A systematic review. J. Neurogastroenterol. Motil. 2016, 22, 589. [Google Scholar] [CrossRef]
- Engels, C.; Schwab, C.; Zhang, J.; Stevens, M.J.; Bieri, C.; Ebert, M.O.; McNeill, K.; Sturla, S.J.; Lacroix, C. Acrolein contributes strongly to antimicrobial and heterocyclic amine transformation activities of reuterin. Sci. Rep. 2016, 6, 36246. [Google Scholar] [CrossRef]
- Zhang, J.; Sturla, S.; Lacroix, C.; Schwab, C. Gut Microbial Glycerol Metabolism as an Endogenous Acrolein Source. MBio 2018, 9, e01947-17. [Google Scholar] [CrossRef]
- Vimont, A.; Fernandez, B.; Ahmed, G.; Fortin, H.P.; Fliss, I. Quantitative antifungal activity of reuterin against food isolates of yeasts and moulds and its potential application in yogurt. Int. J. Food Microbiol. 2019, 289, 182–188. [Google Scholar] [CrossRef]
- Talarico, T.L.; Dobrogosz, W.J. Purification and characterization of glycerol dehydratase from Lactobacillus reuteri. Appl. Environ. Microbiol. 1990, 56, 1195–1197. [Google Scholar]
- Toraya, T.; Eda, M.; Kamachi, T.; Yoshizawa, K. Energetic feasibility of hydrogen abstraction and recombination in coenzyme B12-dependent diol dehydratase reaction. J. Biochem. 2001, 130, 865–872. [Google Scholar] [CrossRef]
- Toraya, T. Radical catalysis in coenzyme B12-dependent isomerization (eliminating) reactions. Chem. Rev. 2003, 103, 2095–2128. [Google Scholar] [CrossRef]
- Frey, P.A.; Reed, G.H. Radical mechanisms in adenosylmethionine-and adenosylcobalamin-dependent enzymatic reactions. Arch. Biochem. Biophys. 2000, 382, 6–14. [Google Scholar] [CrossRef]
- Shibata, N.; Nakanishi, Y.; Fukuoka, M.; Yamanishi, M.; Yasuoka, N.; Toraya, T. Structural rationalization for the lack of stereospecificity in coenzyme B12-dependent diol dehydratase. J. Biol. Chem. 2003, 278, 22717–22725. [Google Scholar] [CrossRef]
- Knietsch, A.; Bowien, S.; Whited, G.; Gottschalk, G.; Daniel, R. Identification and characterization of coenzyme B12-dependent glycerol dehydratase-and diol dehydratase-encoding genes from metagenomic DNA libraries derived from enrichment cultures. Appl. Environ. Microbiol. 2003, 69, 3048–3060. [Google Scholar] [CrossRef]
- Finlay, T.H.; Valinsky, J.; Mildvan, A.S.; Abeles, R.H. Electron Spin Resonance Studies with Dioldehydrase Evidence for Radical Intermediates in reactions catalyzed by Coenzyme B12. J. Biol. Chem. 1973, 248, 1285–1290. [Google Scholar]
- Bertin, Y.; Habouzit, C.; Dunière, L.; Laurier, M.; Durand, A.; Duchez, D.; Segura, A.; Thévenot-Sergentet, D.; Baruzzi, F.; Chaucheyras-Durand, F.; et al. Lactobacillus reuteri suppresses E. coli O157: H7 in bovine ruminal fluid: Toward a pre-slaughter strategy to improve food safety? PLoS ONE 2017, 12, e0187229. [Google Scholar] [CrossRef]
- Navarro, J.B.; Mashburn-Warren, L.; Bakaletz, L.O.; Bailey, M.T.; Goodman, S.D. Enhanced probiotic potential of Lactobacillus reuteri when delivered as a biofilm on dextranomer microspheres that contain beneficial cargo. Front. Microbiol. 2017, 8, 489. [Google Scholar] [CrossRef]
- De Fouchécour, F.; Sánchez-Castañeda, A.K.; Saulou-Bérion, C.; Spinnler, H.É. Process engineering for microbial production of 3-hydroxypropionic acid. Biotechnol. Adv. 2018, 36, 1207–1222. [Google Scholar] [CrossRef]
- Zhang, J.; Empl, M.T.; Schwab, C.; Fekry, M.I.; Engels, C.; Schneider, M.; Lacroix, C.; Steinberg, P.; Sturla, S.J. Gut microbial transformation of the dietary imidazoquinoxaline mutagen MelQx reduces its cytotoxic and mutagenic potency. Toxicol. Sci. 2017, 159, 266–276. [Google Scholar] [CrossRef]
- Asare, P.T.; Greppi, A.; Stettler, M.; Schwab, C.; Stevens, M.J.; Lacroix, C. Decontamination of minimally-processed fresh lettuce using reuterin produced by Lactobacillus reuteri. Front. Microbiol. 2018, 9, 1421. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Recent Advances in Petroleum Microbiology. Microbiol. Mol. Biol. Rev. 2003, 67, 503–549. [Google Scholar] [CrossRef]
- Troise, A.D.; Buonanno, M.; Fiore, A.; Monti, S.M.; Fogliano, V. Evolution of protein bound Maillard reaction end-products and free Amadori compounds in low lactose milk in presence of fructosamine oxidase I. Food Chem. 2016, 212, 722–729. [Google Scholar] [CrossRef]
- Qian, Y.; Zheng, J.; Lin, Z. Loop engineering of amadoriase II and mutational cooperativity. Appl. Microbiol. Biotechnol. 2013, 97, 8599–8607. [Google Scholar] [CrossRef]
- Collard, F.; Fagan, R.L.; Zhang, J.; Nemet, I.; Palfey, B.A.; Monnier, V.M. The Cation—π Interaction between Lys53 and the Flavin of Fructosamine Oxidase (FAOX-II) Is Critical for Activity. Biochemistry 2011, 50, 7977–7986. [Google Scholar] [CrossRef]
- Backhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- D’Argenio, V.; Salvatore, F. The role of the gut microbiome in the healthy adult status. Clin. Chim. Acta 2015, 451, 97–102. [Google Scholar] [CrossRef]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next generation beneficial microbe: Akkermansia Muciniphilia. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, L.; Kumar, J.; Mistry, A.; Sankar Chittoor Mana, T.; Perry, G.; Reddy, V.P.; Obrenovich, M. The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation. Biomedicines 2019, 7, 24. https://doi.org/10.3390/biomedicines7020024
Jones L, Kumar J, Mistry A, Sankar Chittoor Mana T, Perry G, Reddy VP, Obrenovich M. The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation. Biomedicines. 2019; 7(2):24. https://doi.org/10.3390/biomedicines7020024
Chicago/Turabian StyleJones, Lucas, Jessica Kumar, Adil Mistry, Thriveen Sankar Chittoor Mana, George Perry, V. Prakash Reddy, and Mark Obrenovich. 2019. "The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation" Biomedicines 7, no. 2: 24. https://doi.org/10.3390/biomedicines7020024
APA StyleJones, L., Kumar, J., Mistry, A., Sankar Chittoor Mana, T., Perry, G., Reddy, V. P., & Obrenovich, M. (2019). The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation. Biomedicines, 7(2), 24. https://doi.org/10.3390/biomedicines7020024