
Selective Inhibition of Cardiac Late Na+ Current Is Based on Fast Offset Kinetics of the Inhibitor
 ,
,  , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Isolation of Cardiomyocytes
2.3. Electrophysiology
2.3.1. Action Potential Voltage Clamp
2.3.2. Conventional Voltage Clamp
2.3.3. Recording of APs from Multicellular Ventricular Preparations
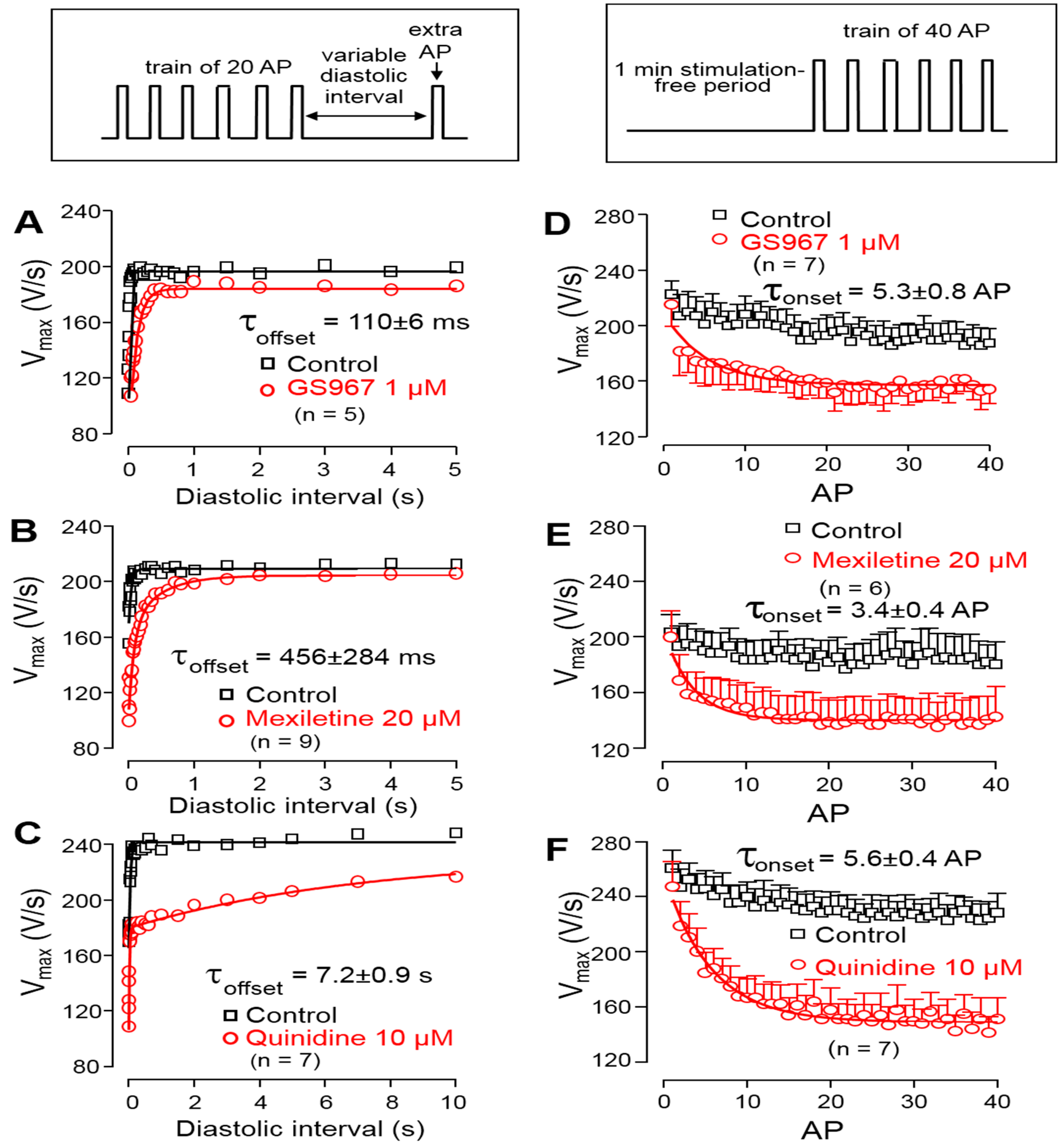
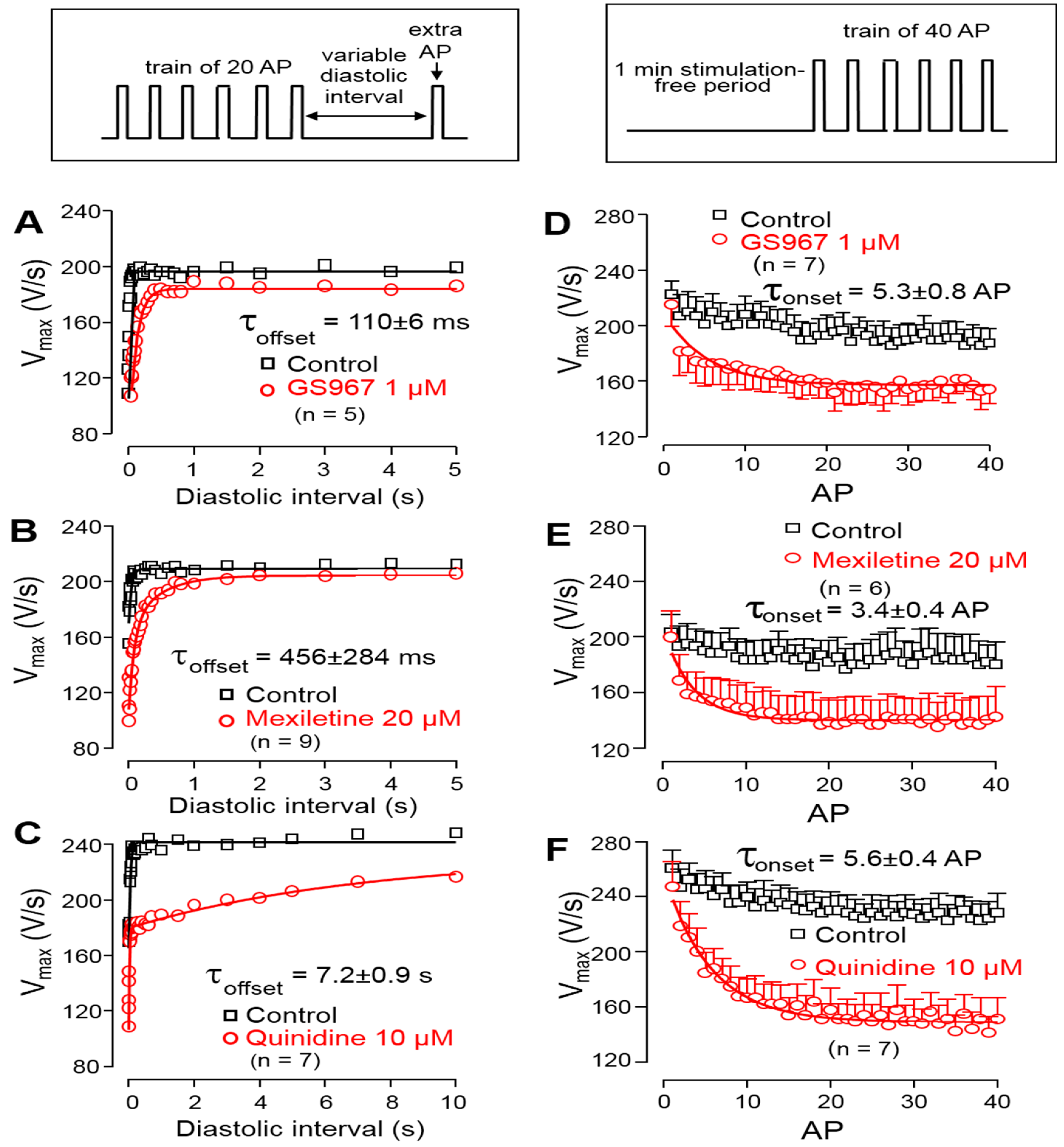
2.3.4. Determination of Offset Kinetics
2.3.5. Determination of Onset Kinetics
2.4. Statistics
3. Results
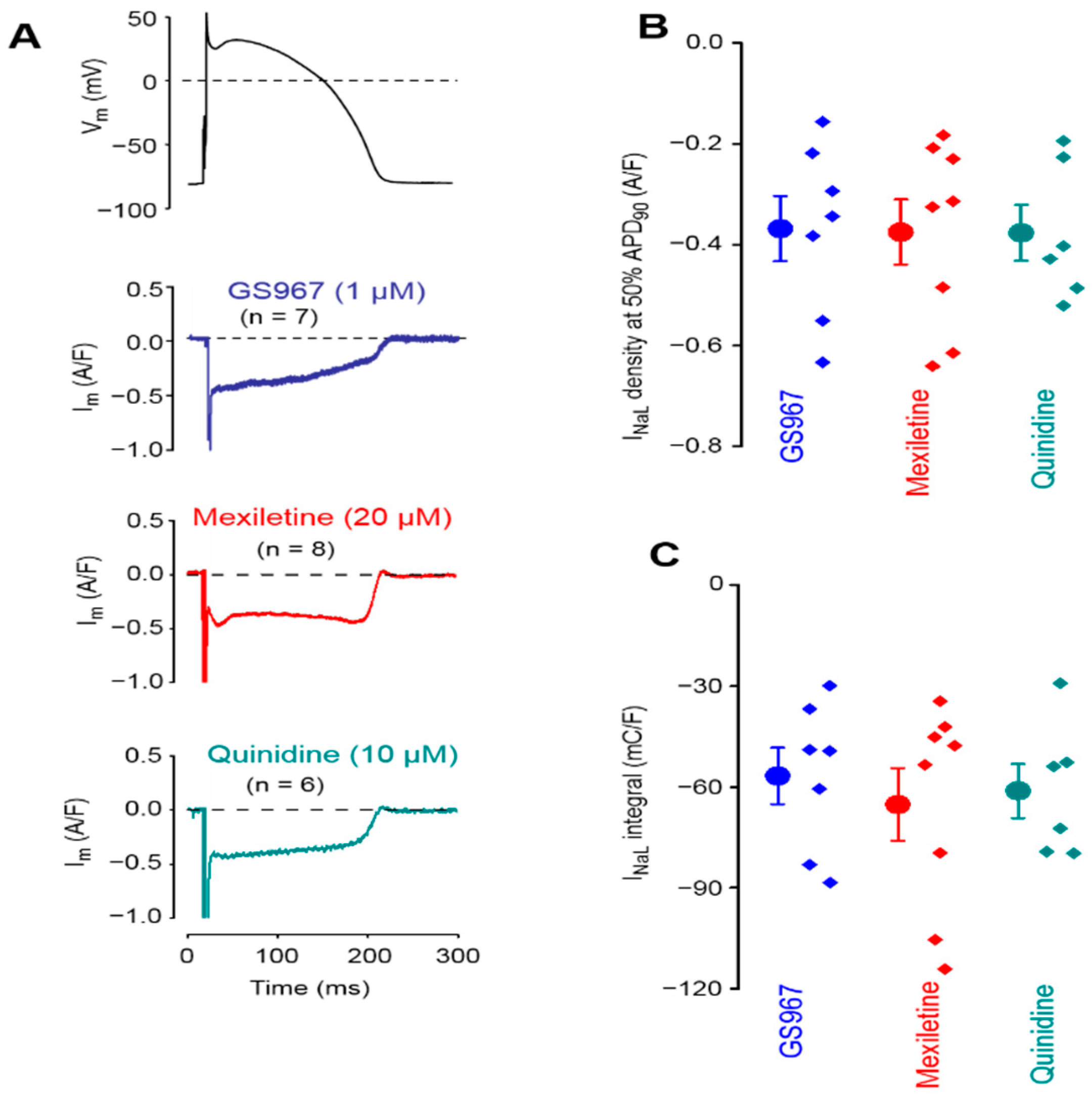
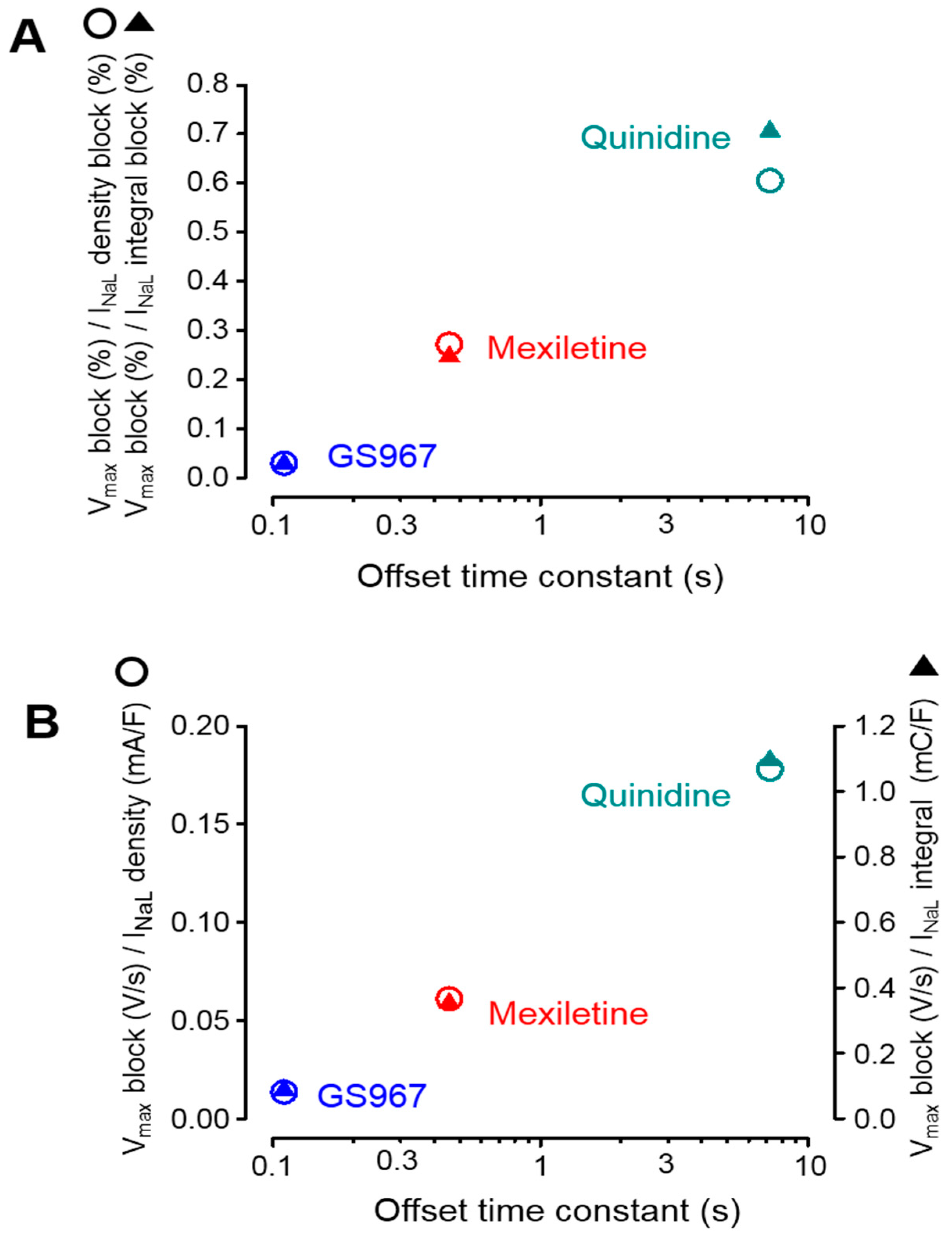
3.1. Effects of GS967, Mexiletine and Quinidine under APVC Conditions
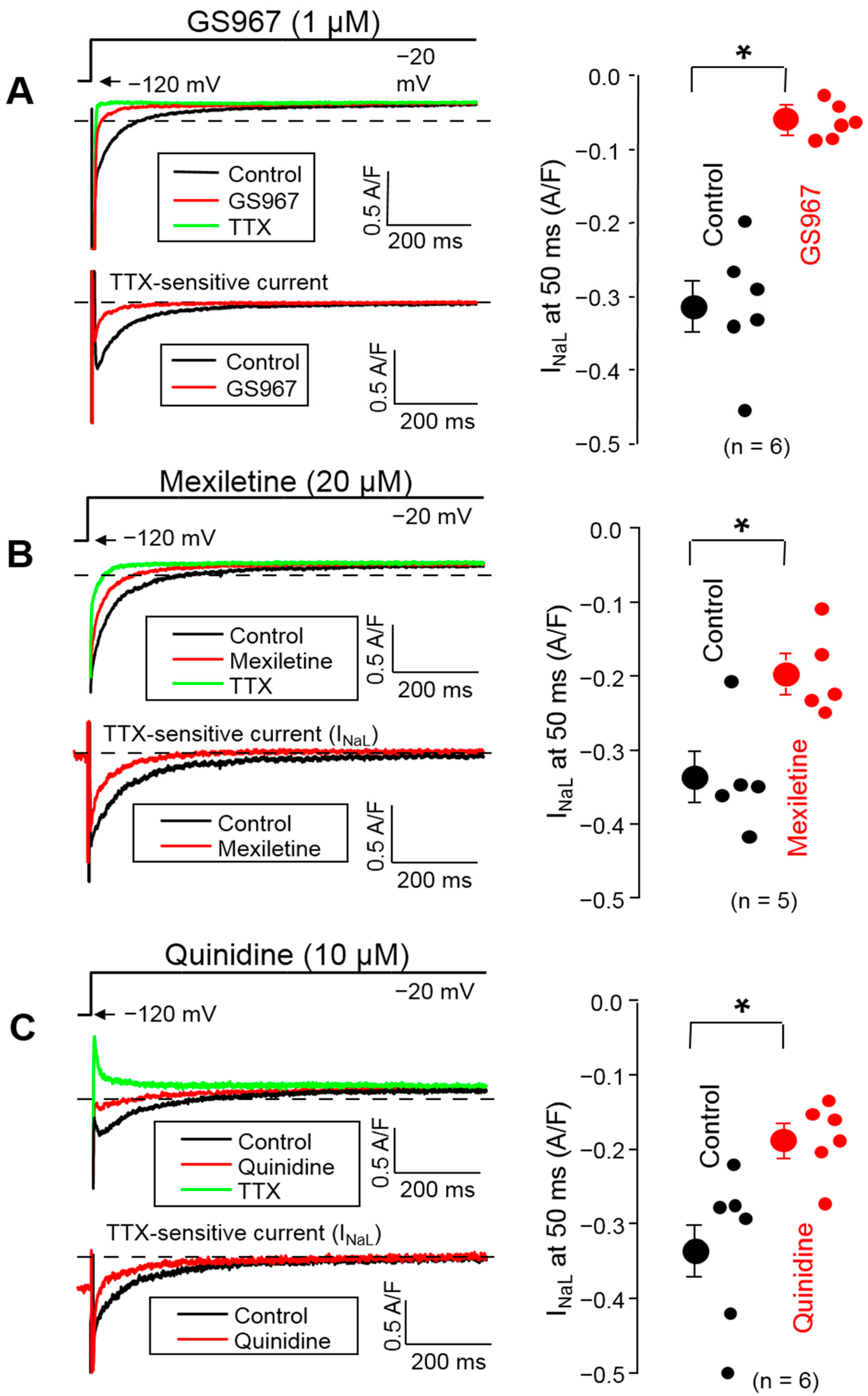
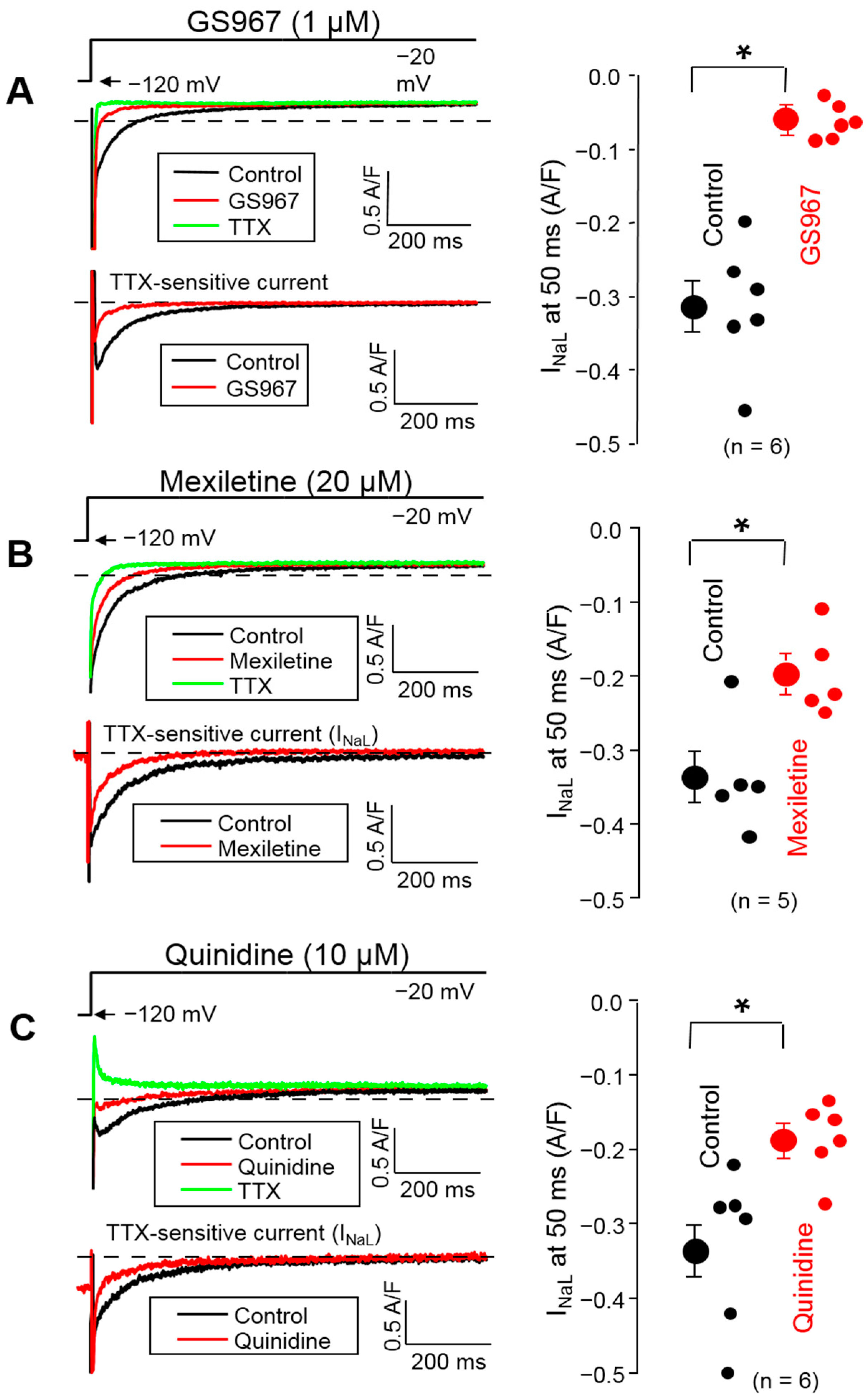
3.2. Effects of GS967, Mexiletine and Quinidine on INaL under Conventional Voltage Clamp Conditions
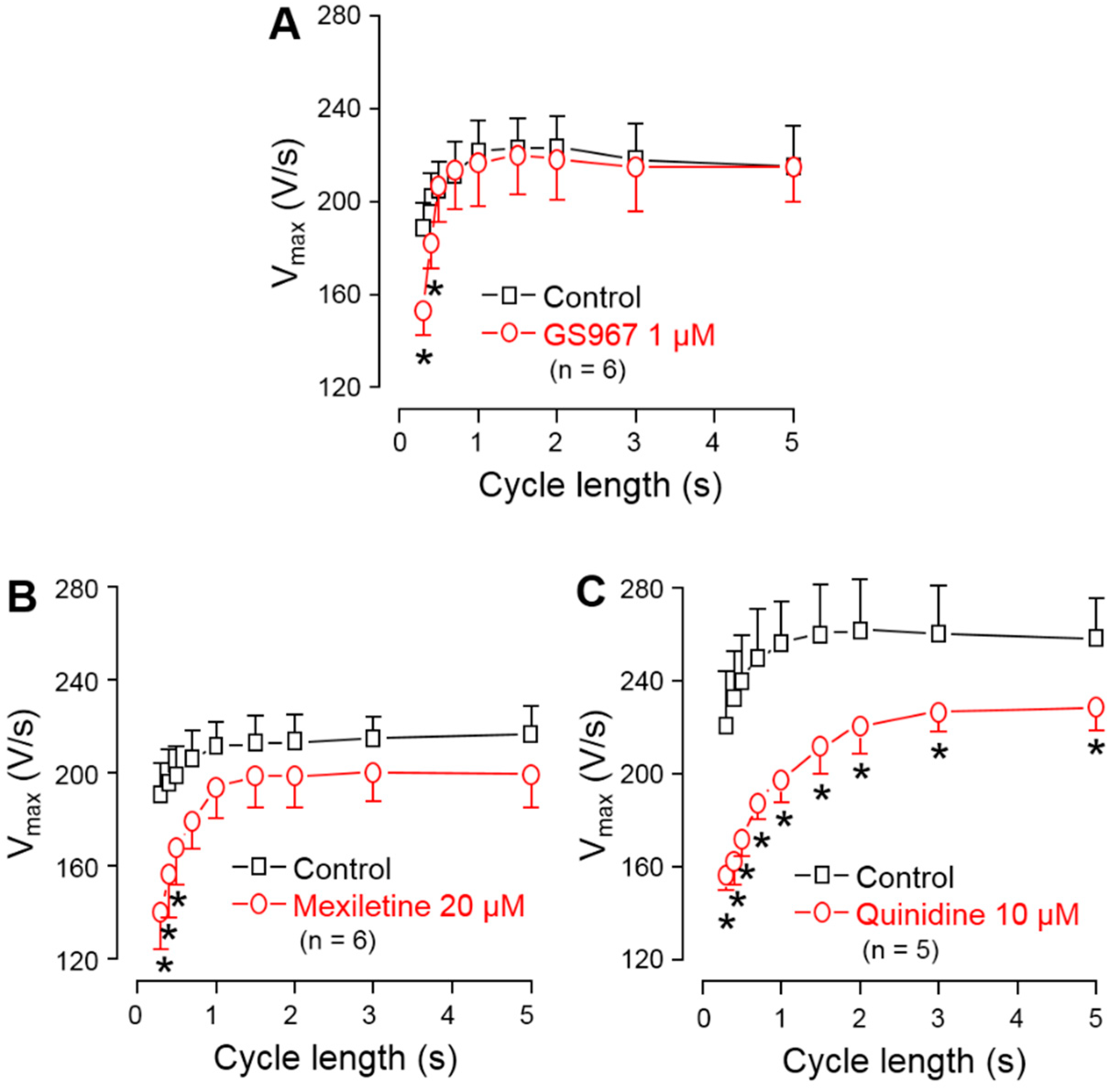
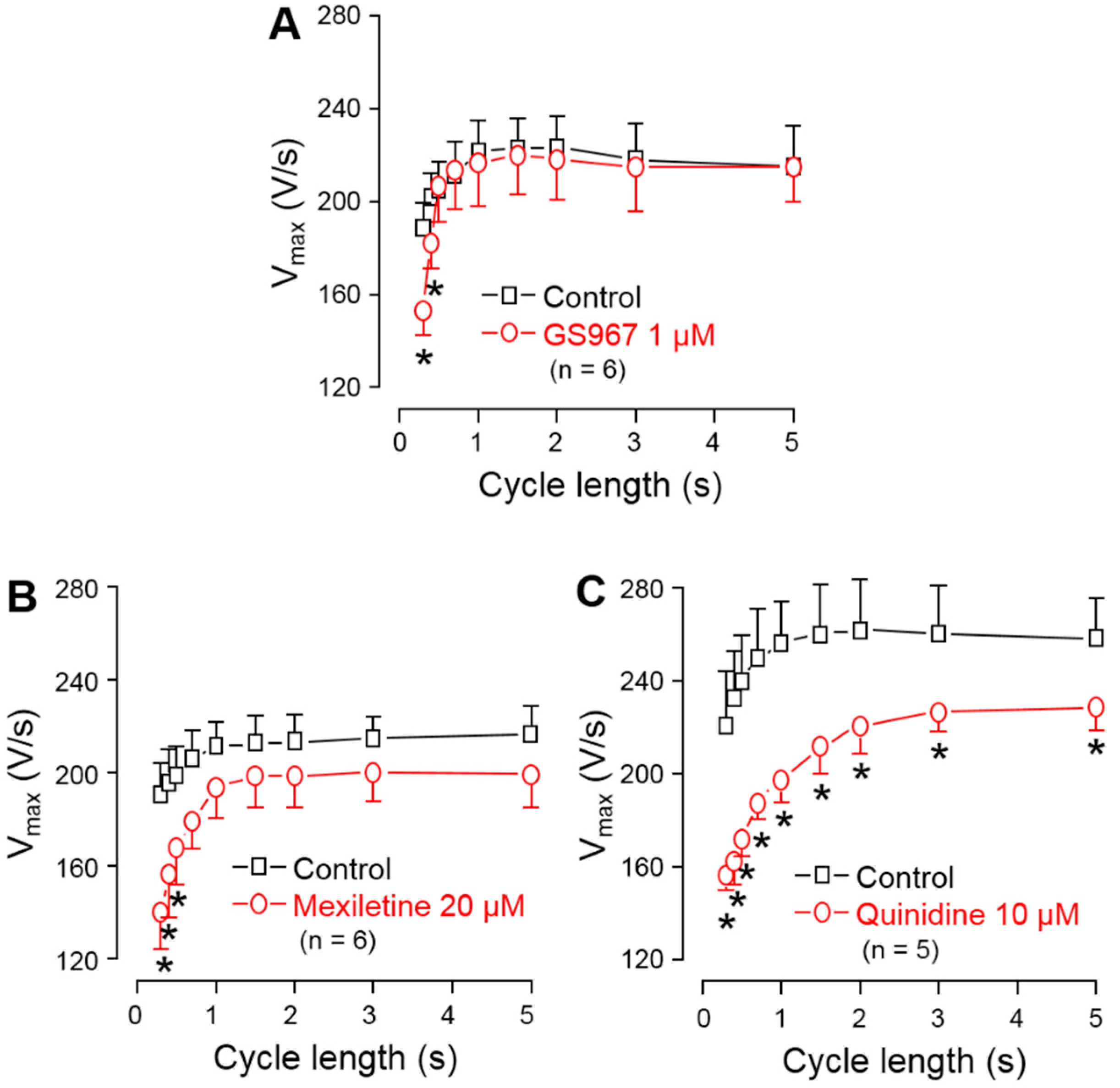
3.3. Effects of GS967, Mexiletine and Quinidine on Action Potential Upstroke
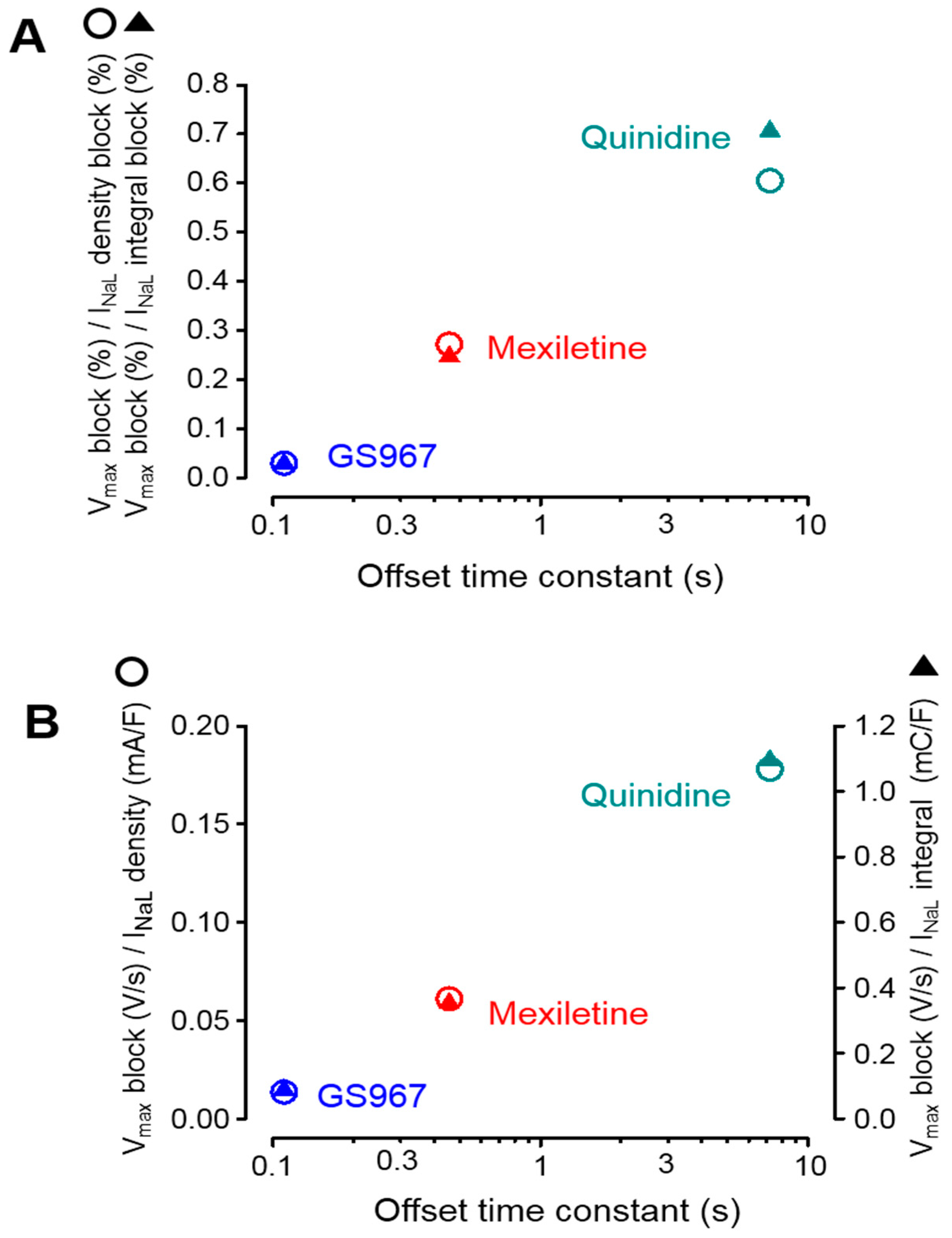
3.4. Selectivity of INaL over INaP Inhibition
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Valdivia, C.R.; Chu, W.W.; Pu, J.; Foell, J.D.; Haworth, R.A.; Wolff, M.R.; Kamp, T.J.; Makielski, J.C. Increased late sodium current in myocytes from a canine heart failure model and from failing human heart. J. Mol. Cell. Cardiol. 2005, 38, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Undrovinas, A.I.; Belardinelli, L.; Undrovinas, N.A.; Sabbah, H.N. Ranolazine improves abnormal repolarization and contraction in left ventricular myocytes of dogs with heart failure by inhibiting late sodium current. J. Cardiovasc. Electrophysiol. 2006, 17 (Suppl. S1), S169–S177. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, V.A.; Sabbah, H.N.; Undrovinas, A.I. Late Sodium Current is a Novel Target for Amiodarone: Studies in Failing Human Myocardium. J. Mol. Cell. Cardiol. 2001, 33, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Coppini, R.; Santini, L.; Olivotto, I.; Ackerman, M.J.; Cerbai, E. Abnormalities in sodium current and calcium homoeostasis as drivers of arrhythmogenesis in hypertrophic cardiomyopathy. Cardiovasc. Res. 2020, 116, 1585–1599. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.M.; Tester, D.J.; Li, R.; Sun, T.; Peterson, B.Z.; Ackerman, M.J.; Makielski, J.C.; Tan, B.-H. Mexiletine rescues a mixed biophysical phenotype of the cardiac sodium channel arising from the SCN5A mutation, N406K, found in LQT3 patients. Channels 2018, 12, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Antzelevitch, C.; Belardinelli, L. The Role of Sodium Channel Current in Modulating Transmural Dispersion of Repolarization and Arrhythmogenesis. J. Cardiovasc. Electrophysiol. 2006, 17 (Suppl. S1), S79–S85. [Google Scholar] [CrossRef]
- Maltsev, V.A.; Silverman, N.; Sabbah, H.N.; Undrovinas, A.I. Chronic heart failure slows late sodium current in human and canine ventricular myocytes: Implications for repolarization variability. Eur. J. Heart Fail. 2007, 9, 219–227. [Google Scholar] [CrossRef]
- Shyrock, J.C.; Song, Y.; Rajamani, S.; Antzelecitch, C.; Belardinelli, L. The antiarrhythmogenic consequences of increasing late INa in the cardiomyocyte. Cardiovasc. Res. 2013, 99, 600–611. [Google Scholar] [CrossRef]
- Potet, F.; Vanoye, C.G.; George, A.L., Jr. Use-Dependent Block of Human Cardiac Sodium Channels by GS967. Mol. Pharmacol. 2016, 90, 52–60. [Google Scholar] [CrossRef]
- Potet, F.; Egecioglu, D.E.; Burridge, P.W.; George, A.L., Jr. GS-967 and Eleclazine Block Sodium Channels in Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes. Mol. Pharmacol. 2020, 98, 540–547. [Google Scholar] [CrossRef]
- Ghovanloo, M.-R.; Abdelsayed, M.; Ruben, P.C. Effects of Amiodarone and N-desethylamiodarone on Cardiac Voltage-Gated Sodium Channels. Front. Pharmacol. 2016, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Salvage, S.C.; Chandrasekharan, K.H.; Jeevaratnam, K.; Dulhunty, A.F.; Thompson, A.J.; Jackson, A.P.; Huang, C.L.H. Multiple targets for flecainide action: Implications for cardiac arrhythmogenesis. Br. J. Pharmacol. 2018, 175, 1260–1278. [Google Scholar] [CrossRef] [PubMed]
- Varro, A.; Elharrar, V.; Surawicz, B. Frequency-dependent effects of several class I antiarrhythmic drugs on Vmax of action potential upstroke in canine cardiac Purkinje fibers. J. Cardiovasc. Pharmacol. 1985, 7, 482–492. [Google Scholar] [CrossRef]
- Varro, A.; Nakaya, Y.; Elharrar, V.; Surawicz, B. Effect of Antiarrhythmic Drugs on the Cycle Length-Dependent Action Potential Duration in Dog Purkinje and Ventricular Muscle Fibers. J. Cardiovasc. Pharmacol. 1986, 8, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Cardiac Arrhythmia Suppression Trial (CAST) Investigators. Preliminary report: Effect of encainide and flecainide on mortality in a randomized trial of arrhythmia suppression after myocardial infarction. N. Engl. J. Med. 1989, 321, 406–412. [Google Scholar] [CrossRef]
- Belardinelli, L.; Liu, G.; Smith-Maxwell, C.; Wang, W.Q.; El-Bizri, N.; Hirakawa, R.; Karpinski, S.; Li, H.C.; Hu, L.; Li, X.-J.; et al. A novel, potent, and selective inhibitor of cardiac late sodium current suppresses experimental arrhythmias. J. Pharmacol. Exp. Ther. 2013, 344, 23–32. [Google Scholar] [CrossRef]
- Azam, M.A.; Zamiri, N.; Massé, S.; Kusha, M.; Lai, P.F.; Nair, G.K.; Tan, N.S.; Labos, C.; Nanthakumar, K. Effects of Late Sodium Current Blockade on Ventricular Refibrillation in a Rabbit Model. Circ. Arrhythmia Electrophysiol. 2017, 10, e004331. [Google Scholar] [CrossRef]
- Del Canto, I.; Santamaría, L.; Genovés, P.; Such-Miquel, L.; Arias-Mutis, O.; Zarzoso, M.; Soler, C.; Parra, G.; Tormos, A.; Alberola, A.; et al. Effects of the inhibition of late sodium current by GS967 on stretch-induced changes in cardiac electro-physiology. Cardiovasc. Drugs Ther. 2018, 32, 413–425. [Google Scholar]
- Bossu, A.; Houtman, M.J.; Meijborg, V.M.; Varkevisser, R.; Beekman, H.D.; Dunnink, A.; de Bakker, J.M.T.; Mollova, N.; Rajamani, S.; Belardinelli, L.; et al. Selective late sodium current inhibitor GS-458967 suppresses torsades de pointes by mostly affecting perpet-uation but not initiation of the arrhythmia. Br. J. Pharmacol. 2018, 175, 2470–2482. [Google Scholar] [CrossRef]
- Horváth, B.; Hézső, T.; Szentandrássy, N.; Kistamás, K.; Árpádffy-Lovas, T.; Varga, R.; Gazdag, P.; Veress, R.; Dienes, C.; Baranyai, D.; et al. Late sodium current in human, canine and guinea pig ventricular myocardium. J. Mol. Cell. Cardiol. 2020, 139, 14–23. [Google Scholar] [CrossRef]
- Jost, N.; Virág, L.; Comtois, P.; Ördög, B.; Szuts, V.; Seprényi, G.; Bitay, M.; Kohajda, Z.; Koncz, I.; Nagy, N.; et al. Ionic mechanisms limiting cardiac repolarization reserve in humans compared to dogs. J. Physiol. 2013, 591, 4189–4206. [Google Scholar] [CrossRef] [PubMed]
- Szabó, G.; Szentandrássy, N.; Bíró, T.; Tóth, B.I.; Czifra, G.; Magyar, J.; Bányász, T.; Varró, A.; Kovács, L.; Nánási, P.P. Asymmetrical distribution of ion channels in canine and human left-ventricular wall: Epicardium versus midmyocardium. Pflüg. Arch. 2005, 450, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Szentadrassy, N.; Banyasz, T.; Biro, T.; Szabo, G.; Toth, B.I.; Magyar, J.; Lazar, J.; Varro, A.; Kovacs, L.; Nanasi, P.P. Apico-basal inhomogeneity in distribution of ion channels in canine and human ventricular myocardium. Cardiovasc. Res. 2005, 65, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Jost, N.; Acsai, K.; Horváth, B.; Bányász, T.; Baczkó, I.; Bitay, M.; Nanasi, P.P. Contribution of IKr and IK1 to ventricular repolarization in canine and human myocytes: Is there any influence of duration? Basic. Res. Cardiol. 2009, 104, 33–41. [Google Scholar] [CrossRef]
- Hegyi, B.; Bányász, T.; Izu, L.T.; Belardinelli, L.; Bers, D.M.; Chen-Izu, Y. β-adrenergic regulation of late Na+ current during cardiac action potential is mediated by both PKA and CaMKII. J. Mol. Cell. Cardiol. 2018, 123, 168–179. [Google Scholar] [CrossRef]
- Horvath, B.; Banyasz, T.; Jian, Z.; Hegyi, B.; Kistamas, K.; Nanasi, P.P.; Izu, L.T.; Chen-Izu, Y. Dynamics of the late Na+ current during cardiac action potential and its contribution to after depolarizations. J. Mol. Cell. Cardiol. 2013, 64, 59–68. [Google Scholar] [CrossRef]
- Hegyi, B.; Bossuyt, J.; Griffiths, L.G.; Shimkunas, R.; Coulibaly, Z.; Jian, Z.; Grimsrud, K.N.; Sondergaard, C.S.; Ginsburg, K.S.; Chiamvimonvat, N.; et al. Complex electrophysiological remodeling in postinfarction ischemic heart failure. Proc. Natl. Acad. Sci. USA 2018, 115, E3036–E3044. [Google Scholar] [CrossRef]
- Hézső, T.; Naveed, M.; Dienes, C.; Kiss, D.; Prorok, J.; Árpádffy-Lovas, T.; Varga, R.; Fujii, E.; Mercan, T.; Topal, L.; et al. Mexiletine-like cellular electrophysiological effects of GS967 in canine ventricular myocardium. Sci. Rep. 2021, 11, 9565. [Google Scholar] [CrossRef]
- Hegyi, B.; Horváth, B.; Váczi, K.; Gönczi, M.; Kistamás, K.; Ruzsnavszky, F.; Veress, R.; Izu, L.T.; Chen-Izu, Y.; Bányász, T.; et al. Ca2+-activated Cl− current is antiarrhythmic by reducing both spatial and temporal heterogeneity of cardiac repolarization. J. Mol. Cell. Cardiol. 2017, 109, 27–37. [Google Scholar] [CrossRef]
- Bányász, T.; Fülöp, L.; Magyar, J.; Szentandrássy, N.; Varró, A.; Nánási, P.P. Endocardial versus epicardial differences in L-type calcium current in canine ventricular myocytes studied by action potential voltage clamp. Cardiovasc. Res. 2003, 58, 66–75. [Google Scholar] [CrossRef]
- Bányász, T.; Magyar, J.; Szentandrássy, N.; Horváth, B.; Birinyi, P.; Szentmiklósi, J.; Nánási, P.P. Action potential clamp fingerprints of K+ currents in canine cardiomyocytes: Their role in ventricular re-polarization. Acta Physiol. Scand. 2007, 190, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hondeghem, L.M. Validity of Vmax as a measure of the sodium current in cardiac and nervous tissues. Biophys. J. 1978, 23, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Sheets, M.F.; Hanck, D.A.; Fozzard, H.A. Nonlinear relation between Vmax and INa in canine cardiac Purkinje cells. Circ. Res. 1988, 63, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.J.; Bean, B.P.; Tsien, R.W. Maximal upstroke velocity as an index of available sodium conductance. Comparison of maximal upstroke velocity and voltage clamp measurements of sodium current in rabbit Purkinje fibers. Circ. Res. 1984, 54, 636–651. [Google Scholar] [CrossRef]
- Hondeghem, L.M.; Katzung, B.G. Antiarrhythmic Agents: The Modulated Receptor Mechanism of Action of Sodium and Calcium Channel-Blocking Drugs. Annu. Rev. Pharmacol. Toxicol. 1984, 24, 387–423. [Google Scholar] [CrossRef]
- Grant, A.O.; Starmer, C.F.; Strauss, H.C. Antiarrhythmic drug action. Blockade of the inward sodium current. Circ. Res. 1984, 55, 427–439. [Google Scholar] [CrossRef]
- Mangold, K.E.; Brumback, B.D.; Angsutararux, P.; Voelker, T.L.; Zhu, W.; Kang, P.W.; Moreno, J.D.; Silva, J.R. Mechanisms and models of cardiac sodium channel inactivation. Channels 2017, 11, 517–533. [Google Scholar] [CrossRef]
- Karagueuzian, H.S.; Pezhouman, A.; Angelini, M.; Olcese, R. Enhanced Late Na and Ca Currents as Effective Antiarrhythmic Drug Targets. Front. Pharmacol. 2017, 8, 36. [Google Scholar] [CrossRef]
- Rougier, J.S.; Abriel, H. Role of “non-cardiac” voltage-gated sodium channels in cardiac cells. J. Mol. Cell. Cadiol. 2012, 53, 589–590. [Google Scholar] [CrossRef]
- Ahmad, S.; Tirilomis, P.; Pabel, S.; Dybkova, N.; Hartmann, N.; Molina, C.E.; Tirilomis, T.; Kutschka, I.; Frey, N.; Maier, L.S.; et al. The functional consequences of sodium channel Nav1.8 in human left ventricular hypertrophy. ESC Heart Fail. 2019, 6, 154–163. [Google Scholar] [CrossRef]
- Gaborit, N.; Le Bouter, S.; Szuts, V.; Varro, A.; Escande, D.; Nattel, S.; Demolombe, S. Regional and tissue specific transcript signatures of ion channel genes in the non-diseased human heart. J. Physiol. 2007, 582, 675–693. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.; Bertelli, S.; Pusch, M.; Gavazzo, P. Late sodium current blocker GS967 inhibits persistent currents induced by familial hemiplegic migraine type 3 mutations of the SCN1A gene. J. Headache Pain 2019, 20, 107. [Google Scholar] [CrossRef] [PubMed]
- Szél, T.; Koncz, I.; Jost, N.; Baczkó, I.; Husti, Z.; Virág, L.; Bussek, A.; Wettwer, E.; Ravens, U.; Papp, J.G.; et al. Class I/B antiarrhythmic property of ranolazine, a novel antianginal agent, in dog and human cardiac preparations. Eur. J. Pharmacol. 2011, 662, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, T.; January, C.T.; Makielski, J.C. Preferential block of late sodium current in the LQT3 DeltaKPQ mutant by the class I(C) antiarrhythmic flecainide. Mol. Pharmacol. 2000, 57, 101–107. [Google Scholar]
- Williams, E.M.V. A Classification of Antiarrhythmic Actions Reassessed After a Decade of New Drugs. J. Clin. Pharmacol. 1984, 24, 129–147. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naveed, M.; Mohammed, A.S.A.; Topal, L.; Kovács, Z.M.; Dienes, C.; Ovári, J.; Szentandrássy, N.; Magyar, J.; Bányász, T.; Prorok, J.; et al. Selective Inhibition of Cardiac Late Na+ Current Is Based on Fast Offset Kinetics of the Inhibitor. Biomedicines 2023, 11, 2383. https://doi.org/10.3390/biomedicines11092383
Naveed M, Mohammed ASA, Topal L, Kovács ZM, Dienes C, Ovári J, Szentandrássy N, Magyar J, Bányász T, Prorok J, et al. Selective Inhibition of Cardiac Late Na+ Current Is Based on Fast Offset Kinetics of the Inhibitor. Biomedicines. 2023; 11(9):2383. https://doi.org/10.3390/biomedicines11092383
Chicago/Turabian StyleNaveed, Muhammad, Aiman Saleh A. Mohammed, Leila Topal, Zsigmond Máté Kovács, Csaba Dienes, József Ovári, Norbert Szentandrássy, János Magyar, Tamás Bányász, János Prorok, and et al. 2023. "Selective Inhibition of Cardiac Late Na+ Current Is Based on Fast Offset Kinetics of the Inhibitor" Biomedicines 11, no. 9: 2383. https://doi.org/10.3390/biomedicines11092383
APA StyleNaveed, M., Mohammed, A. S. A., Topal, L., Kovács, Z. M., Dienes, C., Ovári, J., Szentandrássy, N., Magyar, J., Bányász, T., Prorok, J., Jost, N., Virág, L., Baczkó, I., Varró, A., Nánási, P. P., & Horváth, B. (2023). Selective Inhibition of Cardiac Late Na+ Current Is Based on Fast Offset Kinetics of the Inhibitor. Biomedicines, 11(9), 2383. https://doi.org/10.3390/biomedicines11092383